Variability in Anthocyanins, Phenolic Compounds and Antioxidant Capacity in the Tassels of Collected Waxy Corn Germplasm

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Experimental Design
2.2. Chemicals and Reagents
2.3. Sample Preparation and Extraction
2.4. Total Anthocyanin Content (TAC)
2.5. Total Phenolic Content (TPC)
2.6. Antioxidant Capacity by DPPH and TEAC Assay
2.7. Statistical Analysis
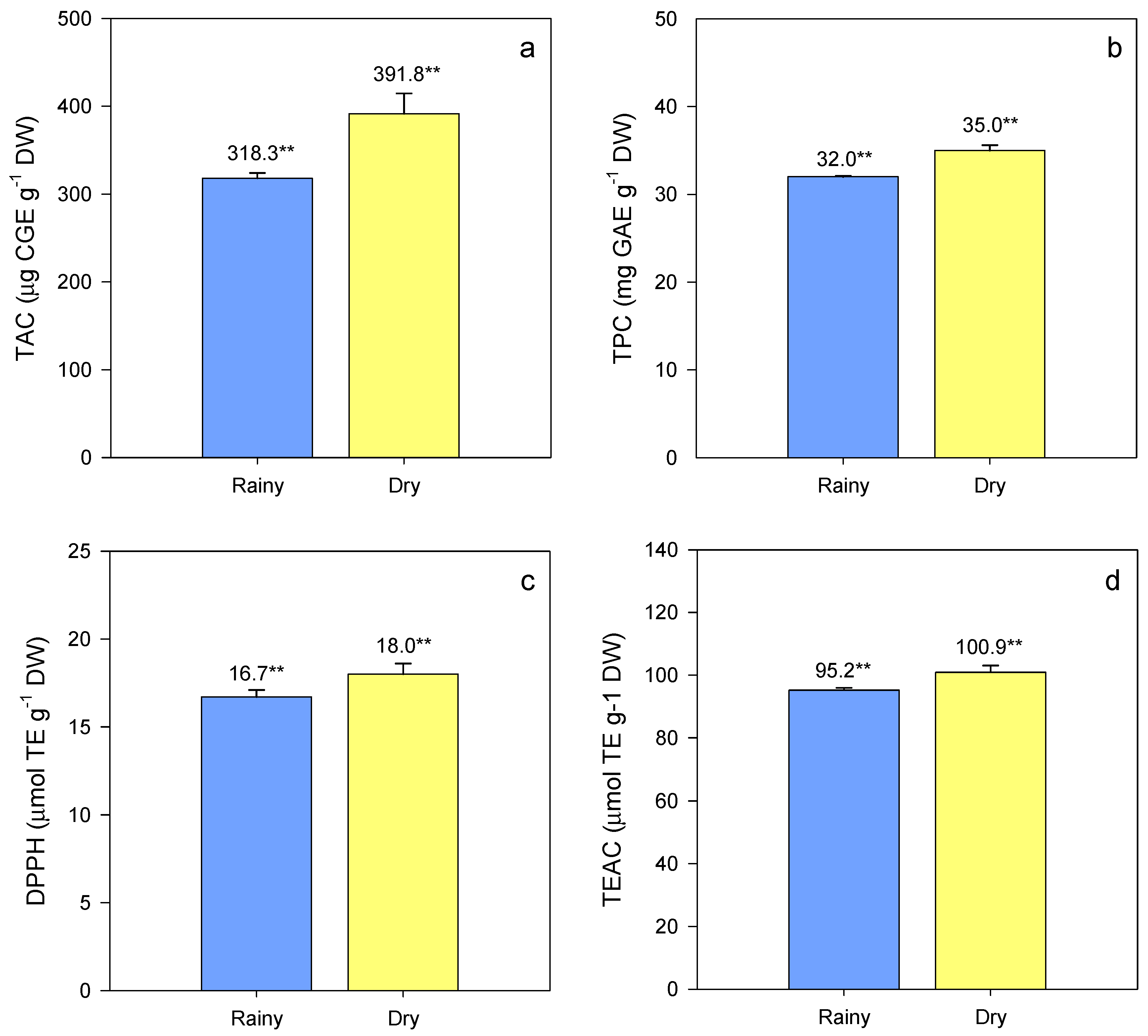
3. Results and Discussion
3.1. Phytochemicals and Antioxidant Variation in Waxy Corn Germplasm
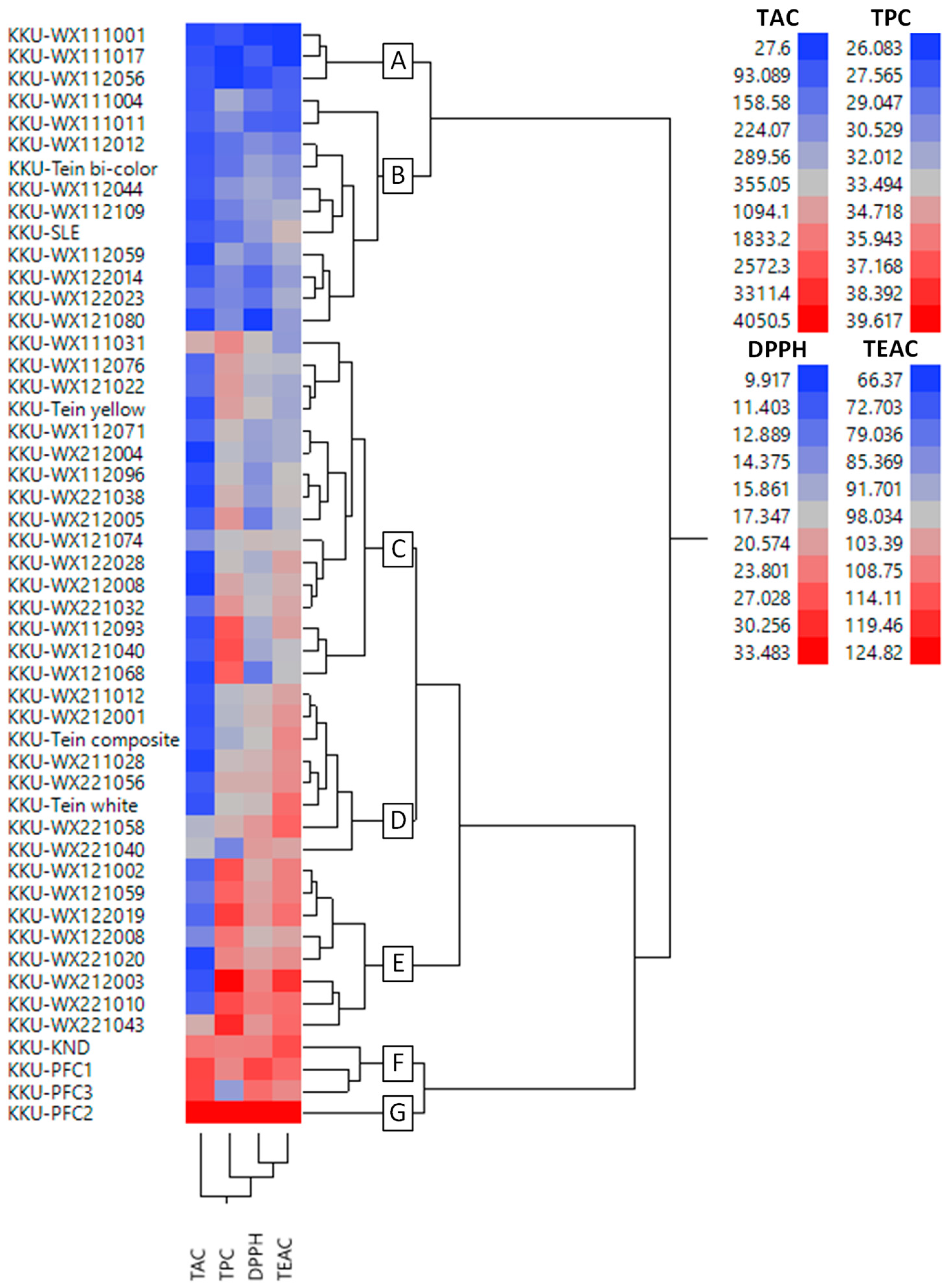
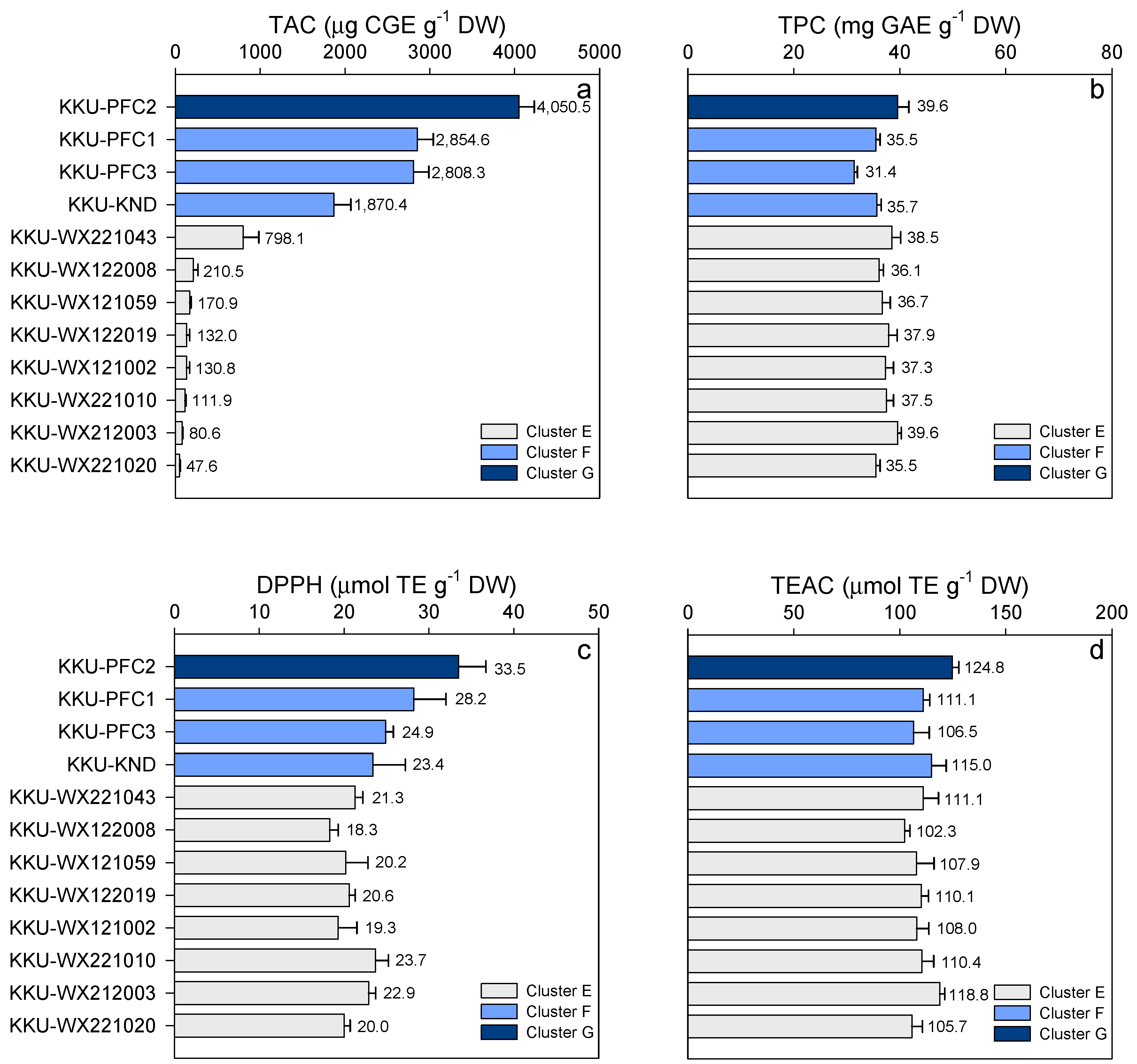
3.2. Cluster Analysis
3.3. Pearson Correlation Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lertrat, K.; Thongnarin, N. Novel approach to eating quality improvement in local waxy corn: Improvement of sweet taste in local waxy corn variety with mixed kernels from super sweet corn. Acta Hortic. 2008, 769, 145–150. [Google Scholar] [CrossRef]
- Simla, S.; Boontang, S.; Harakotr, B. Anthocyanin content, total phenolic content, and antiradical capacity in different ear components of purple waxy corn at two maturation stages. Aust. J. Crop Sci. 2016, 10, 675–682. [Google Scholar] [CrossRef]
- Duangpapeng, P.; Ketthaisong, D.; Lomthaisong, K.; Lertrat, K.; Scott, M.P.; Suriharn, B. Corn tassel: A new source of phytochemicals and antioxidant potential for value-added product development in the agro-industry. Agronomy 2018, 8, 242. [Google Scholar] [CrossRef]
- Lopez-Martinez, L.X.; Oliart-Ros, R.M.; Valerio-Alfaro, G.; Lee, C.-H.; Parkin, K.L.; Garcia, H.S. Antioxidant activity, phenolic compounds and anthocyanins content of eighteen strains of Mexican maize. LWT-Food Sci. Technol. 2009, 42, 1187–1192. [Google Scholar] [CrossRef]
- Žilić, S.; Serpen, A.; Akıllıoğlu, G.; Gökmen, V.; Vančetović, J. Phenolic compounds, carotenoids, anthocyanins, and antioxidant capacity of colored maize (Zea mays L.) kernels. J. Agric. Food Chem. 2012, 60, 1224–1231. [Google Scholar] [CrossRef] [PubMed]
- Harakotr, B.; Suriharn, B.; Tangwongchai, R.; Scott, M.P.; Lertrat, K. Anthocyanins and antioxidant activity in coloured waxy corn at different maturation stages. J. Funct. Foods 2014, 9, 109–118. [Google Scholar] [CrossRef] [Green Version]
- Petroni, K.; Pilu, R.; Tonelli, C. Anthocyanins in corn: A wealth of genes for human health. Planta 2014, 240, 901–911. [Google Scholar] [CrossRef] [PubMed]
- Thiraphatthanavong, P.; Wattanathorn, J.; Muchimapura, S.; Thukham-mee, W.; Lertrat, K.; Suriharn, B. The combined extract of purple waxy corn and ginger prevents cataractogenesis and retinopathy in streptozotocin-diabetic rats. Oxid. Med. Cell. Longev. 2014. [Google Scholar] [CrossRef]
- Berardo, N.; Mazzinelli, G.; Valoti, P.; Laganà, P.; Redaelli, R. Characterization of maize germplasm for the chemical composition of the grain. J. Agric. Food Chem. 2009, 57, 2378–2384. [Google Scholar] [CrossRef]
- González-Muñoz, A.; Quesille-Villalobos, A.M.; Fuentealba, C.; Shetty, K.; Gálvez Ranilla, L. Potential of Chilean native corn (Zea mays L.) accessions as natural sources of phenolic antioxidants and in vitro bioactivity for hyperglycemia and hypertension management. J. Agric. Food Chem. 2013, 61, 10995–11007. [Google Scholar] [CrossRef]
- Harakotr, B.; Suriharn, B.; Scott, M.P.; Lertrat, K. Genotypic variability in anthocyanins, total phenolics, and antioxidant activity among diverse waxy corn germplasm. Euphytica 2014, 203, 237–248. [Google Scholar] [CrossRef] [Green Version]
- Menkir, A.; Maziya-Dixon, B.; Mengesha, W.; Rocheford, T.; Alamu, E.O. Accruing genetic gain in pro-vitamin A enrichment from harnessing diverse maize germplasm. Euphytica 2017, 213. [Google Scholar] [CrossRef]
- Kirisattayakul, W.; Wattanathorn, J.; Iamsaard, S.; Jittiwat, J.; Suriharn, B.; Lertrat, K. Neuroprotective and memory-enhancing effect of the combined extract of purple waxy corn cob and pandan in ovariectomized rats. Oxid. Med. Cell. Longev. 2017. [Google Scholar] [CrossRef]
- Intuyod, K.; Priprem, A.; Limphirat, W.; Charoensuk, L.; Pinlaor, P.; Pairojkul, C.; Lertrat, K.; Pinlaor, S. Anti-inflammatory and anti-periductal fibrosis effects of an anthocyanin complex in Opisthorchis viverrini-infected hamsters. Food Chem. Toxicol. 2014, 74, 206–215. [Google Scholar] [CrossRef]
- Intuyod, K.; Priprem, A.; Pairojkul, C.; Hahnvajanawong, C.; Vaeteewoottacharn, K.; Pinlaor, P.; Pinlaor, S. Anthocyanin complex exerts anti-cholangiocarcinoma activities and improves the efficacy of drug treatment in a gemcitabine-resistant cell line. Int. J. Oncol. 2018, 52, 1715–1726. [Google Scholar] [CrossRef] [PubMed]
- Mohsen, S.M.; Ammar, A.S.M. Total phenolic contents and antioxidant activity of corn tassel extracts. Food Chem. 2009, 112, 595–598. [Google Scholar] [CrossRef]
- Komosinska-Vassev, K.; Olczyk, P.; Kaźmierczak, J.; Mencner, L.; Olczyk, K. Bee pollen: Chemical composition and therapeutic application. Evid.-Based Complement. Altern. Med. 2015. [Google Scholar] [CrossRef]
- Khider, M.; Elbanna, K.; Mahmoud, A.; Owayss, A.A. Egyptian honeybee pollen as antimicrobial, antioxidant agents, and dietary food supplements. Food Sci. Biotechnol. 2013, 22, 1461–1469. [Google Scholar] [CrossRef]
- Žilić, S.; Vančetović, J.; Janković, M.; Maksimović, V. Chemical composition, bioactive compounds, antioxidant capacity and stability of floral maize (Zea mays L.) pollen. J. Funct. Foods 2014, 10, 65–74. [Google Scholar] [CrossRef]
- Wang, L.; Yu, Y.; Fang, M.; Zhan, C.; Pan, H.; Wu, Y.; Gong, Z. Antioxidant and antigenotoxic activity of bioactive extracts from corn tassel. J. Huazhong Univ. Sci. Technol. Med. Sci. 2014, 34, 131–136. [Google Scholar] [CrossRef]
- Wille, J.J.; Berhow, M.A. Bioactives derived from ripe corn tassels: A possible new natural skin whitener, 4-hydroxy−1-oxindole-3-acetic acid. Curr. Bioact. Compd. 2011, 7, 126–134. [Google Scholar] [CrossRef]
- Prohens, J. Plant breeding: A success story to be continued thanks to the advances in genomics. Front. Plant Sci. 2011, 2, 51. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Fan, G.; Gu, Z.; Han, Y.; Chen, Z. Optimization extraction of anthocyanins from purple corn (Zea mays L.) cob using tristimulus colorimetry. Eur. Food Res. Technol. 2008, 227, 409–415. [Google Scholar] [CrossRef]
- Giusti, M.M.; Wrolstad, R.E. Characterization and measurement of anthocyanins by UV-visible spectroscopy. In Current Protocols in Food Analytical Chemistry; Wrolstad, R.E., Acree, T.E., An, H., Decker, E.A., Penner, M.A., Reid, D.S., Schwartz, S.J., Shoemaker, C.F., Sporns, P., Eds.; John Wiley & Sons Inc.: New York, NY, USA, 2001; pp. F1.2.1–F1.2.13. [Google Scholar]
- Hu, Q.-P.; Xu, J.-G. Profiles of carotenoids, anthocyanins, phenolics, and antioxidant activity of selected color waxy corn grains during maturation. J. Agric. Food Chem. 2011, 59, 2026–2033. [Google Scholar] [CrossRef] [PubMed]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Gomaz, K.A.; Gomez, A.A. Statistical Procedures for Agricultural Research, 2nd ed.; John Wiley and Sons: Singapore, 1984; p. 680. [Google Scholar]
- Reddivari, L.; Hale, A.L.; Miller, J.C. Genotype, location, and year influence antioxidant activity, carotenoid content, phenolic content, and composition in specialty potatoes. J. Agric. Food Chem. 2007, 55, 8073–8079. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.-H.; McClung, A.M.; Bergman, C.J. Phenolic content, anthocyanins and antiradical capacity of diverse purple bran rice genotypes as compared to other bran colors. J. Cereal Sci. 2017, 77, 110–119. [Google Scholar] [CrossRef]
- Khamphasan, P.; Lomthaisong, K.; Harakotr, B.; Ketthaisong, D.; Scott, M.P.; Lertrat, K.; Suriharn, B. Genotypic variation in anthocyanins, phenolic compounds, and antioxidant activity in cob and husk of purple field corn. Agronomy 2018, 8, 271. [Google Scholar] [CrossRef]
- Fonseca, A.; Westgate, M. Relationship between desiccation and viability of maize pollen. Field Crops Res. 2005, 94, 114–125. [Google Scholar] [CrossRef]
- Van Bilsen, D.G.J.L.; Hoekstra, F.A.; Crowe, L.M.; Crowe, J.H. Altered phase behavior in membranes of aging dry pollen may cause imbibitional leakage. Plant Physiol. 1994, 104, 1193–1199. [Google Scholar] [CrossRef]
- Cheynier, V.; Comte, G.; Davies, K.M.; Lattanzio, V.; Martens, S. Plant phenolics: Recent advances on their biosynthesis, genetics, and ecophysiology. Plant Physiol. Biochem. 2013, 72, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Khampas, S.; Lertrat, K.; Lomthaisong, K.; Simla, S.; Suriharn, B. Effect of location, genotype and their interactions for anthocyanins and antioxidant activities of purple waxy corn cobs. Turk. J. Field Crops 2015, 20, 15–23. [Google Scholar] [CrossRef]
- Mohamed, G.; Lertrat, K.; Suriharn, B. Phenolic compound, anthocyanin content, and antioxidant activity in some parts of purple waxy corn across maturity stages and locations. Int. Food Res. J. 2017, 24, 490–497. [Google Scholar]
- Heraty, J.M.; Ellstrand, N.C. Maize germplasm conservation in Southern California’s urban gardens: Introduced diversity beyond ex situ and in situ management. Econ. Bot. 2016, 70, 37–48. [Google Scholar] [CrossRef]
- Lu, Y.; Yan, J.; Guimarães, C.T.; Taba, S.; Hao, Z.; Gao, S.; Chen, S.; Li, J.; Zhang, S.; Vivek, B.S.; et al. Molecular characterization of global maize breeding germplasm based on genome-wide single nucleotide polymorphisms. Theor. Appl. Genet. 2009, 120, 93–115. [Google Scholar] [CrossRef]
- Adewale, B.D.; Dumet, D.J.; Vroh-Bi, I.; Kehinde, O.B.; Ojo, D.K.; Adegbite, A.E.; Franco, J. Morphological diversity analysis of African yam bean (Sphenostylis stenocarpa Hochst. ex A. Rich.) harms and prospects for utilization in germplasm conservation and breeding. Genet. Resour. Crop Evol. 2011, 59, 927–936. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Source of Variation | df | Phytochemicals | Antioxidant Capacity | ||
|---|---|---|---|---|---|
| TAC | TPC | DPPH | TEAC | ||
| Season (S) | 1 | 404,390 ** (0.2) a | 638.6 ** (8.7) | 112.1 * (1.6) | 2484.9 * (3.2) |
| Genotype (G) | 49 | 3,864,741 ** (97.5) | 63.4 ** (42.3) | 121.5 ** (83.5) | 946.9 ** (59.6) |
| S × G | 49 | 82,648 ** (2.1) | 65.8 ** (43.8) | 20.2 ** (13.9) | 568.7 ** (35.8) |
| Error | 196 | 1519 (0.2) | 1.8 (4.7) | 0.1 (0.3) | 2.7 (0.7) |
| C.V. (%) | 10.98 | 3.98 | 1.97 | 1.66 |
| Clusters | No. Genotype | Phytochemicals | Antioxidant Capacity | ||
|---|---|---|---|---|---|
| TAC (µg CGE g−1 DW) | TPC (mg GAE g−1 DW) | DPPH (µmol TE g−1 DW) | TEAC (µmol TE g−1 DW) | ||
| A | 3 | 61.9 ± 19.0 | 28.4 ± 7.5 | 11.9 ± 3.0 | 75.3 ± 24.7 |
| B | 11 | 92.0 ± 36.2 | 29.8 ± 4.3 | 13.4 ± 2.7 | 85.9 ± 14.9 |
| C | 16 | 135.6 ± 179.4 | 34.8 ± 3.5 | 16.0 ± 2.2 | 96.8 ± 9.5 |
| D | 8 | 139.7 ± 161.5 | 33.0 ± 3.4 | 19.0 ± 2.8 | 106.3 ± 5.2 |
| E | 8 | 210.3 ± 238.8 | 37.4 ± 3.8 | 20.8 ± 2.2 | 109.3 ± 10.6 |
| F | 3 | 2,511.1 ± 180.7 | 34.2 ± 4.3 | 25.5 ± 3.6 | 110.9 ± 10.7 |
| G | 1 | 4,050.5 ± 180.7 | 39.6 ± 2.1 | 33.5 ± 3.2 | 124.8 ± 3.0 |
| Parameters | TAC | TPC | DPPH |
|---|---|---|---|
| TPC | 0.19 ** | ||
| DPPH | 0.70 ** | 0.60 ** | |
| TEAC | 0.33 ** | 0.76 ** | 0.73 ** |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duangpapeng, P.; Lertrat, K.; Lomthaisong, K.; Paul Scott, M.; Suriharn, B. Variability in Anthocyanins, Phenolic Compounds and Antioxidant Capacity in the Tassels of Collected Waxy Corn Germplasm. Agronomy 2019, 9, 158. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9030158
Duangpapeng P, Lertrat K, Lomthaisong K, Paul Scott M, Suriharn B. Variability in Anthocyanins, Phenolic Compounds and Antioxidant Capacity in the Tassels of Collected Waxy Corn Germplasm. Agronomy. 2019; 9(3):158. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9030158
Chicago/Turabian StyleDuangpapeng, Prakasit, Kamol Lertrat, Khomsorn Lomthaisong, Marvin Paul Scott, and Bhalang Suriharn. 2019. "Variability in Anthocyanins, Phenolic Compounds and Antioxidant Capacity in the Tassels of Collected Waxy Corn Germplasm" Agronomy 9, no. 3: 158. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9030158