1. Introduction
Misuse of agricultural fertilizers to boost plant production systems has occurred, even if the increase in fertilizers has not always corresponded to an increase in productivity. Due to the limited mineral uptake capacity of the shallow root system, especially in vegetable cropping systems, most of the applied mineral fertilizers have been lost in the soil, causing its pollution and degradation [
1,
2]. In the greenhouse sector, where the fertilizer inputs per unit area are much higher than any other agricultural system, this situation appears particularly serious [
2,
3,
4]. Hence, sustainable agricultural intensification can be achieved only by improving plant crops’ resource use efficiency (RUE) while reducing the use of chemical fertilizers and their environmental impact [
5,
6]. A promising and environmentally friendly approach could be the use of plant biostimulants (PB), which are defined at European level as:
products stimulating plant nutrition processes independently of the product’s nutrient content, with the aim of improving one or more of the following characteristics of the plant: (1) nutrient use efficiency, (2) tolerance to abiotic stress, (3) crop quality traits, or (4) availability of confined nutrients in the soil and rhizosphere.
Among PB, natural bioactive substances (seaweed and plant extracts, protein hydrolysates, and humic and fulvic acids) and beneficial microorganisms (mycorrhizal fungi and plant growth-promoting rhizobacteria belonging to the genera
Azospirillum,
Azotobacter, and
Rhizobium spp.) have been shown to improve plant productivity and tolerance to abiotic stressors by modifying and regulating the activities of carbon and nitrogen metabolism enzymes, hormonal activities, and secondary metabolism [
7,
8,
9,
10,
11]. In addition to the benefits of PB on yield and abiotic stress tolerance, biostimulants have the capacity to improve nutrient use efficiency (NUE), which is considered an important claim for both economic and environmental reasons [
8]. PB have been proven to boost NUE by improving uptake, assimilation, and use of macro- and micronutrients [
12,
13,
14,
15,
16]. This latter effect has been mainly related to root morphological changes, including increases in density, length, surface area, number of lateral roots, and root exudation of low-molecular-weight compounds, which facilitate the formation of micro-aggregates that enhance microbial activity and, in particular, nitrogen and phosphorus uptake [
1,
13,
17]. PB can increase the activity of the plasma membrane H
+-ATPase acidifying rhizosphere and increase the solubility of micronutrients [
14]. The positive effects of PB on plant RUE also include the improvement of nutritional and nutraceutical quality of plants, increasing the synthesis and accumulation of bioactive molecules with antioxidant activity like carotenoids, ascorbic acid, or phenolic compounds [
9,
18]. However, the responses induced by PBs, like the commercial vegetal-derived biostimulant produced through water extraction and fermentation of tropical plant biomass (Auxym
®, Italpollina, Rivoli Veronese, Italy), which highly improves the mineral status of spinach and tomato [
1,
16], have been investigated only in control conditions (i.e., absence of stress). To the best of our knowledge, no data are available concerning the effect of these vegetal-derived biostimulants (containing mainly phytohormones, amino acids, peptides, vitamins, and microelements) on yield and functional quality of leafy vegetable crops under different levels of nutrients in the media solution, even if improving plant crops’ RUE while reducing the use of chemical fertilizers is a priority.
Jute plants (
Corchorus sp.), shrubs belonging to the
Malvaceae family that grow spontaneously in the wild, are cultivated in all tropical areas because of their potential and multiple uses both for industrial processing and food [
19]. In the Middle East and North Africa, the species
Corchorus olitorius L. (
n = 24), known by the common name of molokhia, is consumed as a leafy vegetable, fresh or dry, accompanying soups or as part of traditional dishes of Arab and African culture [
19,
20,
21,
22]. It has a high nutritional value as source of essential amino acids, such as methionine [
23]; however, it has important nutraceutical properties, containing high concentrations of β-carotene, riboflavin, ascorbic acid, folic acid, iron, tocopherols, and phenolic compounds [
24]. Given the presence of these antioxidant metabolites, its use has been associated with the prevention of chronic diseases such as cancer, diabetes, hypertension, and heart disease [
24,
25,
26]. Even the use of a nutrient solution with halved nutrient concentrations does not negatively affect the growth and post-harvest performance of jute plants [
23]. Therefore, it would be particularly interesting to verify whether the use of a bio-activator of vegetal origin could better modulate the yield performance, physiological parameters and phytochemical quality of jute in conditions of optimal or suboptimal supply of mineral nutrients.
Considering this background, the aim of this study was to assess the effects of three different nutrient solution concentrations (full-, half-, and quarter-strength), in the absence or presence of a tropical plant extract (PE), to analyze the interactive response of nutrient solution concentration × PE in Jute. We were able to evaluate the ability of this biostimulant to favor plant growth and RUE under optimal and sub-optimal nutrient regimens. Treated and untreated jute plants were compared in terms of morpho-physiological parameters, mineral profile, and the contents of carbohydrates, proteins, and amino acids, including essential amino acids, chlorophyll, anthocyanins, and polyphenols.
2. Materials and Methods
2.1. Plant Material, Glasshouse Conditions, and Experimental Design
The experiment was conducted in the 2017 summer-autumn growing season in a glasshouse situated at the experimental station of the Department of Agricultural Sciences, University of Naples Federico II, located in Bellizzi (Salerno), Southern Italy (43°31′ N, 14°58′ E; 60 m above sea level). The crop species tested for the present experiment was jute (Corchorus olitorius L.). Seeds of Jute (Nehme Establishment for Trade and Agriculture, Batroun, Lebanon) were germinated in vermiculite on August 14, 2017. Thirty days after sowing (September 12, at the two true-leaf stage) the jute seedling were transplanted into plastic pots containing 1.5 L of a peat:vermiculite mixture in a 2:1 volume ratio. Plastic pots were placed on three 1.8-m-wide and 7-m-long benches in single rows. Jute seedlings were spaced 20 cm between the plants with 30 cm between the rows, giving a plant density of 16.6 plants per square meter. Plants were grown under natural light conditions. Inside the glasshouse, the daily air temperature was maintained between 18 °C and 28 °C. Night air temperature was always higher than 16 °C and the relative humidity was 58% and 78% during the day and night, respectively.
Six treatments derived from a factorial combination of three nutrient concentrations (25%, 50%, or 100% of a full strength nutrient solution) and two biostimulant applications (control and or foliar application of a tropical plant extract Auxym®). The treatments were arranged in a randomized complete-block design with three replications per treatment, for a total of 18 experimental units. Each experimental unit consisted of 10 plants, yielding a total number of 180 jute plants.
2.2. Bistimulant Characteristics and Application and Nutrient Solution Management
The tropical plant extract (PE) biostimulant Auxym
® was provided by Italpollina Company (Rivoli Veronese, Italy). The commercial PE biostimulant was produced through water extraction and fermentation of tropical plant biomass. The commercial biostimulant Auxym
® contains mainly phytohormones (auxins and cytokinins), amino acids, peptides, vitamins (C and E, niacin, pyridoxine, riboflavin, and thiamin), and essential micronutrients (B, Cu, Fe, Mn, and Zn). The exact concentrations of the different compounds were reported in detail by Colla et al. [
4]. The treated jute plants were uniformly sprayed (i.e., foliar application) nine days after transplanting (DAT) at weekly intervals (four times during the growing cycle) at an application rate of 2 mL·L
−1 using a 4-L stainless steel sprayer.
The full-strength nutrient solution used in the current glasshouse experiment was a modified Hoagland Arnon formulation used previously by Rouphael et al. [
27] with a composition of 14.4 mM N–NO
3−, 2.0 mM S, 1.2 mM P, 6.0 mM K, 4.5 mM Ca, 2.3 mM Mg, 20 µM Fe, 10 µM Mn, 2 µM Cu, 5 µM Zn, 20 µM B, and 0.5 µM Mo. The electrical conductivity (EC) and pH of the full-strength nutrient solution were 2.0 dS·m
−1 and 6.0 ± 0.2, respectively. The quarter-strength and half-strength solutions had 25% and 50% of the macronutrients concentration, respectively, and the same micronutrient concentrations of the full-strength nutrient solution. The EC of the half- and quarter-strength nutrient solutions were 1.0 ± 0.1 and 0.5 ± 0.1 dS·m
−1, respectively. The full-, half-, and quarter-strength nutrient solutions were pumped from 50-L plastic tanks through a drip irrigation system through auto-compensated drippers having a flow rate of 2 L·h
−1. Irrigation scheduling in all treatments involved using low-tension tensiometers that monitor irrigation based on growing media matric potential as described in detail by Rouphael et al. [
11]. The low-tension tensiometers were connected to an electronic programmer to manage the beginning and the end of the fertigation, which are associated with the high (−10 kPa) and low (−1 kPa) tension set points of the major substrates used in horticulture.
2.3. Biometric Measurements
On October 11, 2017 (31 DAT), six jute plants per replicate were harvested and the following biometric measurements were recorded: plant height, number of leaves per plant, and the total leaf area using an electronic area meter (Li-Cor3000, Li-Cor, Lincoln, NE, USA). The fresh leaf tissue was first weighed then dried at 70 °C for 72 h until they reached a constant weight. After complete drying, leaf samples were weighed again with an analytical balance for dry biomass determination and the dry samples were conserved for mineral analysis. The leaf dry matter percentage was also calculated.
2.4. Photosynthetic Activity and Soil Plant Analysis Development Index
One day before harvest (30 DAT), the net carbon dioxide assimilation rate (A
CO2), was measured with a portable gas exchange analyzer (LCA-4; ADC BioScientific Ltd., Hoddesdon, UK) equipped with a broadleaf chamber (cuvette window area: 6.25 cm
2). This measurement was recorded within 2 h across solar noon (i.e., between 11:00 a.m. and 1:00 p.m.) on eight healthy and fully expanded jute leaves per treatment. Photosynthetically active radiation, relative humidity, and carbon dioxide concentration (585 ± 10 μmol·m
−2·s
−1, 58 ± 1.5%, and 331 ± 0.5 ppm, respectively) were set at ambient value and the flow rate of air was 400 mL·s
−1, as described by Carillo et al. [
28].
On the same date, the Soil Plant Analysis Development (SPAD) index was measured on fully expanded jute leaves by means of a portable SPAD-502 chlorophyll meter (Konica Minolta, Hino-shi Tokyo, Japan) as described by Kumar et al. [
29]. Fifteen leaves were randomly measured and averaged to a single SPAD value for each replicate (i.e., experimental unit).
2.5. Mineral Composition Analysis
The desiccated jute leaf tissue was ground in a Wiley Mill (Model 4, Thomas Scientific, San Mateo, CA, USA) to pass through an 841-µm screen and used for mineral content analysis as described in detail by Rouphael et al. [
10]. Briefly, 0.25 g of desiccated tissue was suspended in 50 mL of ultrapure water (Milli-Q. Merck Millipore, Darmstadt, Germany) and subjected to three freeze-thaw cycles in liquid N, followed by incubation in a shaking 80 °C water bath (ShakeTemp SW22, Julabo, Seelbach, Germany) for 10 min. The suspension was centrifuged at 6,000 rpm for 10 min (R-10 M, Remi Elektrotechnik Limited, Mumbay, India), then filtered through 0.20-μm filter paper. Nitrate, P, K, S, Ca, Mg, S, and Na were separated and quantified by ion chromatography (ICS-3000, Dionex, Sunnyvale, CA, USA) coupled to a conductivity detector. An IonPac CG12A (4 × 250 mm, Dionex, Sunnyvale, CA, USA) guard column and IonPac CS12A (4 × 250 mm, Dionex, Sunnyvale, CA, USA) analytical column were used for the potassium, calcium, magnesium, and sodium analysis. For nitrate, phosphorus, and sulfur quantification, an IonPac AG11-HC guard (4 × 50 mm, Dionex, Sunnyvale, CA, USA) column and IonPac AS11-HC analytical column (4 × 250 mm, Dionex, Sunnyvale, CA, USA) were adopted.
2.6. Chlorophyll Analysis
Chlorophylls (µg·g
−1 fresh weight; fw) were extracted by homogenization of frozen powdered jute leaf tissues (1.0 g) in methanol. The resulting extracts were centrifuged at 4800×
g for 15 min and chlorophyll a and chlorophyll b contents were assessed by recording the absorbance of the supernatant at 665 and 652 nm in a polypropylene microplate by a microplate reader (Synergy HT, BioTEK Instruments, Bad Friedrichshall, Germany) based on the procedure used by Wellburn [
30]. The total chlorophyll content was calculated as the sum of chlorophyll a and b.
2.7. Starch and Soluble Carbohydrates Analysis
Soluble sugars (µmol·g
−1 fw) were quantified in ethanolic extracts of frozen powdered leaf tissues, and the starch in the pellet of the same ethanolic extract after glucose hydrolysis was quantified by an enzymatic assay coupled with reduction of pyridine nucleotides as described by Carillo et al. [
31]. The increase in absorbance at 340 nm was recorded by a FLX-Xenius spectrophotometer (SAFAS, Monaco City, Monaco).
2.8. Polyphenols and Anthocyanins Analysis
The total polyphenols (mg·g
−1 fw) content was determined by the Folin–Ciocalteu method described by Singleton et al. [
31]. Frozen powdered jute leaf tissues (20 mg) were extracted in 800 μL of 60% methanol (
v/
v); 100 μL of extract was mixed with 50 μL of the Folin–Ciocalteu reagent diluted with distilled water (1:1
v/
v) and 650 μL of 3% (
w/
v) sodium carbonate. After 90 min at room temperature, the absorbance at 760 nm was determined in a microplate reader FLX-Xenius spectrophotometer (SAFAS, Monaco City, Monaco). The polyphenols concentration is expressed as gallic acid equivalents (GAE) as described in Carillo et al. [
28].
Anthocyanins (mg·g
−1 fw) were determined in frozen powdered leaf tissues (40 mg) extracted twice in 200 µL of 40% (
v/
v) ethanol on ice for 20 min, according to Carillo et al. [
28]. Two aliquots of 150 µL of clear mixed supernatants or 40% (
v/
v) ethanol (as blank) were added to 75 µL of 25 mM KCl (pH 1.0) or 75 µL of 400 mM sodium acetate (pH 4.5) and the absorbance was determined at 520 and 700 nm using a FLX-Xenius spectrophotometer (SAFAS, Monaco City, Monaco). The anthocyanins are expressed as mg cyanidin-3-glucoside (C3G) equivalents according to Giusti and Wrolstad [
32].
2.9. Soluble Proteins and Free and Total Amino Acids Analysis
Primary amino acids and proline (µmol·g
−1 fw) were extracted from 50 mg of frozen powdered leaf tissues in 1 mL of 60% (
v/
v) ethanol overnight at 4 °C, following Carillo et al. [
33]. Amino acids were estimated by high performance liquid chromatography (HPLC) after pre-column derivatization with o-phthaldialdehyde (OPA), according to Woodrow et al. [
34]. Proline was determined by mixing 50 μL of ethanol extract with 100 μL of a reaction mix prepared with ninhydrin 1% (
w/
v) in acetic acid 60% (
v/
v) and ethanol 20% (
v/
v) at 95 °C for 20 min and reading the absorbance at 520 nm using a Synergy HT microplate reader (Bio-Tek, Bad Friedrichshall, Germany), according to Woodrow et al. [
34].
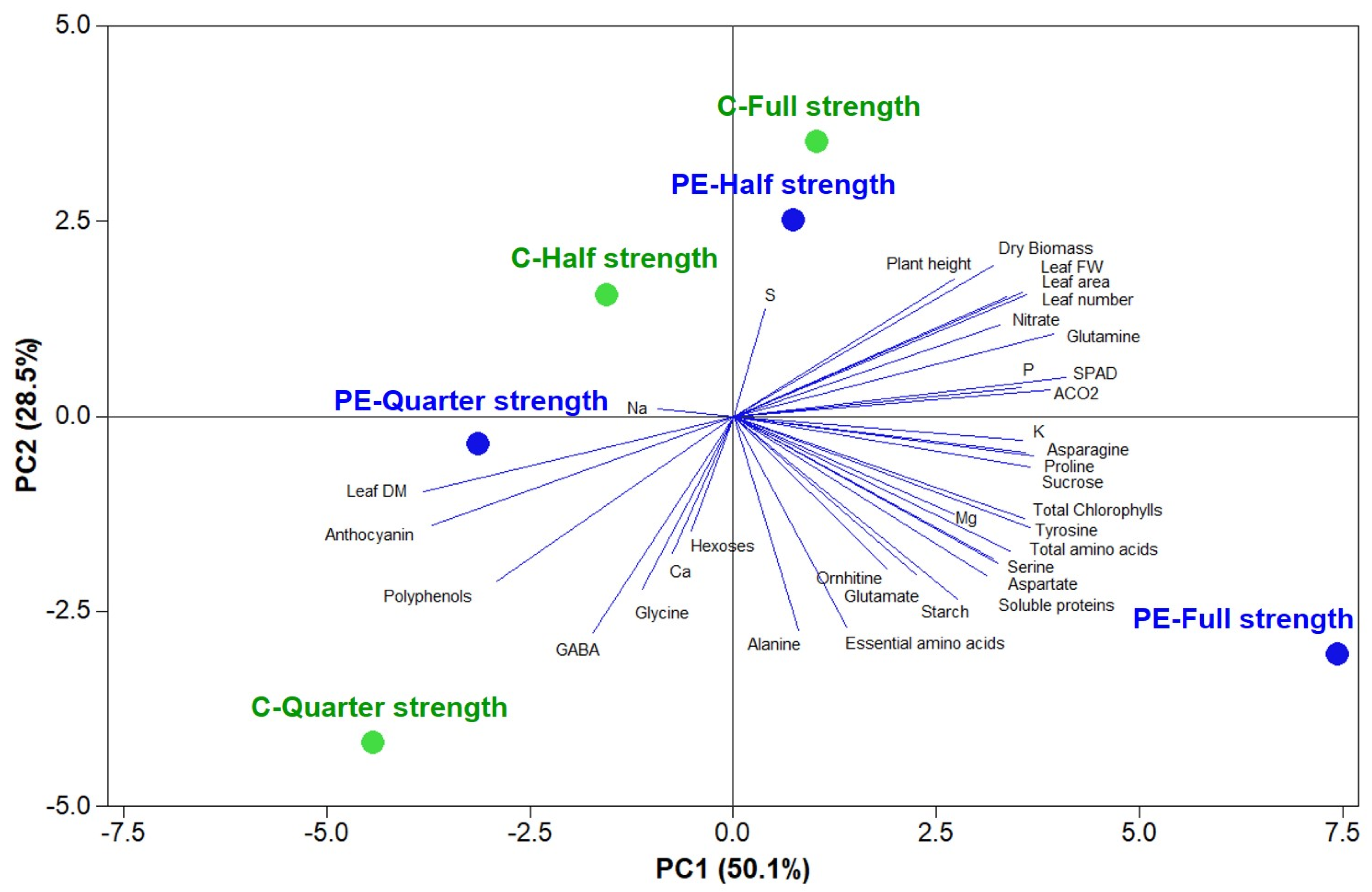
2.10. Statistics, Principal Component Analysis, and Heat Map
All data were subjected to two-way analysis of variance (ANOVA) using the software package SPSS 13 for Windows 2001 (
www.ibm.com/software/analytics/spss). The means were separated by Tukey’s honest significant difference (HSD) test (significance level 0.05). Biostimulant application main effects were compared using the
t-test. Principal component analysis (PCA) was performed on morpho-physiological and phytochemical parameters as well as on the mineral composition to individuate the dominant parameters that most effectively discriminated between nutrient solution concentrations and biostimulant application by using Minitab® 18 statistical software (Minitab LLC, State College, PA, USA). The score plot and loading matrix were also determined based on the first and second principal components (PCs). A heat map generated in Excel (Microsoft Inc., Redmond, Washington) was used to summarize the plant responses to different nutrient strengths and PE. Results were calculated as log
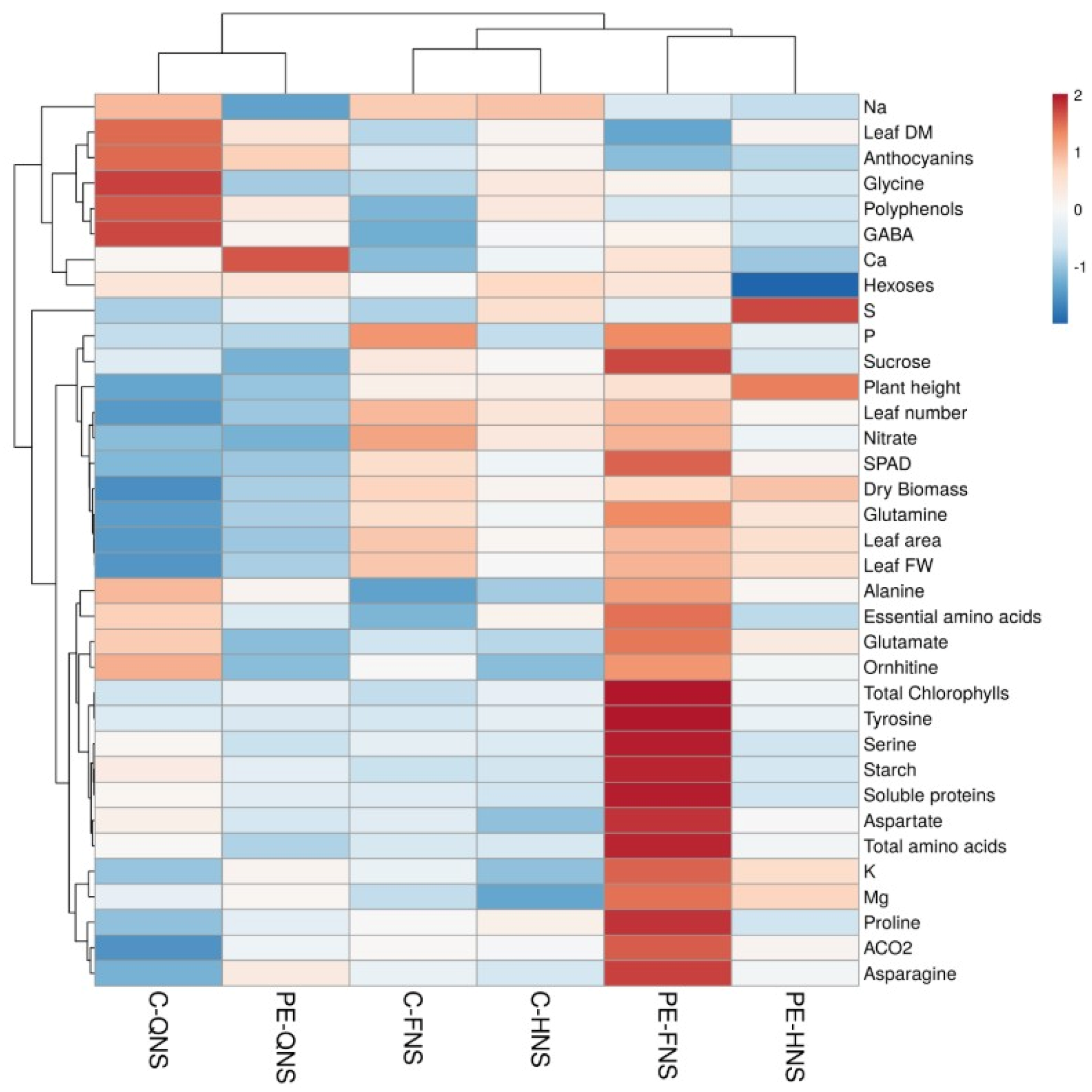
2 (half-strength and quarter-strength solutions/full-strength solution, or PE/control for plants grown at different strength solutions) and are visualized using a false color scale, with red indicating an increase and blue a decrease of values. No differences are denoted by white squares. A different heatmap summarizing all the plant responses to PE and different nutrient strength treatments was generated using the
https://biit.cs.ut.ee/clustvis/ online program package with Euclidean distance as the similarity measure and hierarchical clustering with complete linkage. Data were visualized using a false color scale, with red indicating an increase and blue a decrease of values [
16].
4. Discussion
The horticulture industry uses the highest amount of fertilizer per unit area of any other agricultural system [
4,
27]. However, to sustainably increase agricultural productivity, it is necessary to improve the plant RUE and reduce the use and environmental impact of chemical fertilizers [
6]. For improving sustainable cultivation and safe use in the future, it is necessary to find a compromise between the minimum quantity of fertilizer and the stability of yield quantity and quality. The use of natural plant biostimulants can facilitate this process since they are able to improve uptake, assimilation, and use of macro- and micronutrients, and increase plant yield to improve plant productivity even under reduced nutrient fertilization [
9,
13,
17].
Significant changes in morphological and physiological traits were observed; in particular, the half- and quarter-strength NS sharply reduced leaf area per plant thus decreasing the jute fresh yield and total dry biomass [
26]. The lower macronutrient concentration in the NS negatively affected nitrate and phosphorus content. Due to reduced nitrogen availability, at quarter-strength NS, the CO
2 assimilation rate decreased. Lower leaf nitrogen availability can affect photosynthesis through nitrogen remobilization from photosynthetic enzymes and pigments, decreasing photosynthetic rate and ATP synthesis [
35].
Alanine and GABA strongly increased under the lowest macronutrient concentration in the NS, even if the increase in GABA was not statistically significantly. In addition to their use for protein synthesis, amino acids are precursors of several biosynthetic pathways and play crucial roles in plant stress responses [
36]. From this viewpoint, the strong increase in alanine and GABA under quarter-strength NS could be an important mechanism of biochemical pH-stat for the viability of jute plant under nitrogen limitation. A decrease in ATP can impair the control of plant cell cytoplasmic pH because of the reduced activity of the plasma membrane H
+-ATPase that exerts the ATP-dependent electrogenic extrusion of H
+ ions [
37]. Alanine and GABA syntheses, arising from the decarboxylation of malate to pyruvate as a result of malic enzyme activity for alanine and decarboxylation of glutamate to GABA, are proton-consuming reactions buffering cytosolic acidosis [
38,
39]. Alanine can be easily transaminated to pyruvate and transported into the mitochondrial matrix to be converted to acetyl-Coenzyme A (acetyl-CoA), whereas the GABA shunt can supply NADH and/or succinate to the tricarboxylic acid (TCA) cycle to increase the production of ATP [
40]. High concentrations of GABA and calcium may interact to function as signal of the N status of plants [
41].
Essential amino acids significantly increased under both lower macronutrient concentration in the NS, potentially functioning as antioxidants to protect plants by photo-oxidation due to the lower efficiency of photosynthetic system and the pH impairment. The increase in essential amino acids could also be dependent on an increase of carbohydrates, e.g., starch, in particular at quarter-strength NS. Both the low nitrate content and the decrease in ATP concentration due to the slowing down of the photosynthetic process could be responsible for the accumulation of starch that cannot be used as source of carbon skeletons for the production of primary amino acids glutamine and glutamate [
34,
42]. At half- and quarter-NS, conditions increasing leaf cell susceptibility to light-induced oxidative damage, we found an increase in chlorophyll b, anthocyanins, and polyphenols. Chlorophyll b, as a constituent of the light-harvesting complexes II (LHCII), is pivotal in the dissipation of excess excitation energy [
43], whereas anthocyanins and polyphenols can function as both sunscreens and scavengers of reactive oxygen species (ROS) for protecting young expanding leaves and senescing tissues more susceptible to light damage [
44].
In the current study, weekly foliar application of the commercial tropical plant extract improved fresh yield in jute plants under sub-optimal nutrient regimens (half- and quarter-strength) compared to control treatment. Higher photosynthetic activity, SPAD index, and enhanced leaf nutritional status, reflected by higher K and Mg and lower Na concentrations, were observed in PE-treated jute plants, indicating a more efficient accumulation and translocation of assimilates to photosynthetic sinks irrespective of macronutrient concentration in the nutrient solution, thus allowing improved crop performance. Our results are in line with the previous findings on the effect of commercial vegetal-derived biostimulant produced through water extraction and fermentation of tropical plant biomass (Auxym
®) on tomato [
1], spinach [
16], and lettuce [
45]. A presumed direct mechanism behind the stimulation of jute fresh yield production in response to foliar application of PE under both half- and quarter-strength NS could involve the increased presence of signaling molecules, such as amino acids (glutamic and aspartic acids involved in the stimulation of C and N metabolism), peptides (root-promoting hair peptides), and phytohormones (auxins and cytokinins), which are typical compounds in the commercial PE Auxym
®. The former signaling molecules, which are easily perceived by leaf tissue, may elicit endogenous phytohormone biosynthesis, promoting plant growth and thus crop productivity [
9]. In addition to the direct mechanism of the commercial PE biostimulant, indirect modes of action on crop productivity have been demonstrated when PE was applied to plants [
1]. Foliar application of PE has been shown to improve the uptake, translocation, and accumulation of macronutrients in tomato [
1] and spinach [
16]. The improved nutrient uptake performance of PE-treated plants has been attributed to several mechanisms including (1) modification of root system architecture (higher root biomass, root surface, and length and number of lateral roots) and (2) to the presence of bioactive molecules (amino acids, peptides, and vitamins), which may have boosted sink strength and affected the movement of nutrient substrates (i.e., including minerals) within the plant [
1,
8,
9,
16].
At full-strength NS, PE increased not only the number of active photosystems, as proven by the increase in chlorophyll a, but also that of light harvesting complex (LHC), as shown by the strong increase in chlorophyll b. Photosystems, in particular photosystem II (PSII), operates during day under light notwithstanding the continuous environmental changes, in particular the spectral quality and intensity of light. In higher plants, PSII is highly efficient at low irradiance, whereas under high light, by means of a LHCII, non-photochemical quenching (NPQ) dissipates the excess of light as heat [
46]. The increase in chlorophyll b can increase the quantum yield of O
2 evolution and photosynthetic capacity, decreasing the sensitivity to photo-inhibition [
47]. Therefore, PE was able to mediate, at full-strength NS, an increase in jute plant adaptation to fluctuating light conditions by strongly increasing chlorophyll b content, and allowing the plants to fine-tune photosynthesis in a fluctuating environment, increasing its efficiency, as also demonstrated by the strong increase in starch. This determined a higher efficiency in using nitrate that appeared decreased compared to the control because it was more efficiently reduced and assimilated. Accordingly, the soluble protein and total amino acid contents increased. In particular, there was a strong increase in the primary amino acids glutamate, aspartate, asparagine, and alanine, which are the primary products of nitrogen assimilation involved in transport and storage of organic nitrogen [
36].
At quarter-strength NS and PE treatment, nitrogen was remobilized from free amino acids, in particular glutamate, but not from chlorophylls. While chlorophyll a content did not differ from that at full-strength without PE, chlorophyll b was significantly higher than at full-strength without PE. This allowed the efficient functioning of the photosynthetic process that could satisfy the demand of ATP for nitrogen assimilation in amino acids and use of starch, which decreased. The only amino acids that accumulated were the amides glutamine and asparagine under quarter-strength NS. Amides are not only used as nitrogen storage and transport compounds [
48], but are also involved in osmotic adjustment and macromolecules protection [
34].
5. Conclusions
Breeding methods and new biotechnological advances have been placed alongside studies on the potential role of plant biostimulants, which have been proven to increase uptake, assimilation, and use of macro- and micronutrients, and therefore RUE. Our results demonstrate that both in tropical PE-treated and untreated jute plants, growth parameters and marketable fresh yield decreased in response to a reduction in macronutrient concentration in the nutrient solution. Reduced productivity was accompanied by a reduction in macronutrient concentrations (nitrate and phosphorus) in leaf tissue as well as by a significant reduction in net CO2 assimilation, SPAD index, chlorophyll a, and starch, especially under severe nutrient stress conditions (i.e., quarter-strength). The foliar application of PE boosted the marketable yield under half- and quarter-strength NS compared to the control. Our data showed that the use of a tropical plant extract biostimulant enhanced photosynthetic activity and the SPAD index, and improved leaf nutritional status, reflected in higher potassium and magnesium and lower sodium concentrations, irrespective of the macronutrient concentration in the nutrient solution. Finally, the use of vegetal-derived biostimulant Auxym® reprogrammed the use of nitrate, not only improving the nitrogen use efficiency of jute plants, but also completely changing the distribution patterns of organic nitrogen and carbon in leaf tissues both at high- and low-strength nutrition, improving metabolism and supporting leaf growth and expansion even under sub-optimal nutrient conditions.

 ,
, 







{kind=link}
{kind=link}
{kind=link}