Effect of Gibberellic Acid on Growth, Yield, and Quality of Leaf Lettuce and Rocket Grown in a Floating System


Abstract
:1. Introduction
2. Materials and Methods
2.1. Leafy Vegetable Cultivation
2.2. Statistics and Principal Component Analysis
3. Results
3.1. Morpho–Physiological Parameters and Yield of Leaf Lettuce
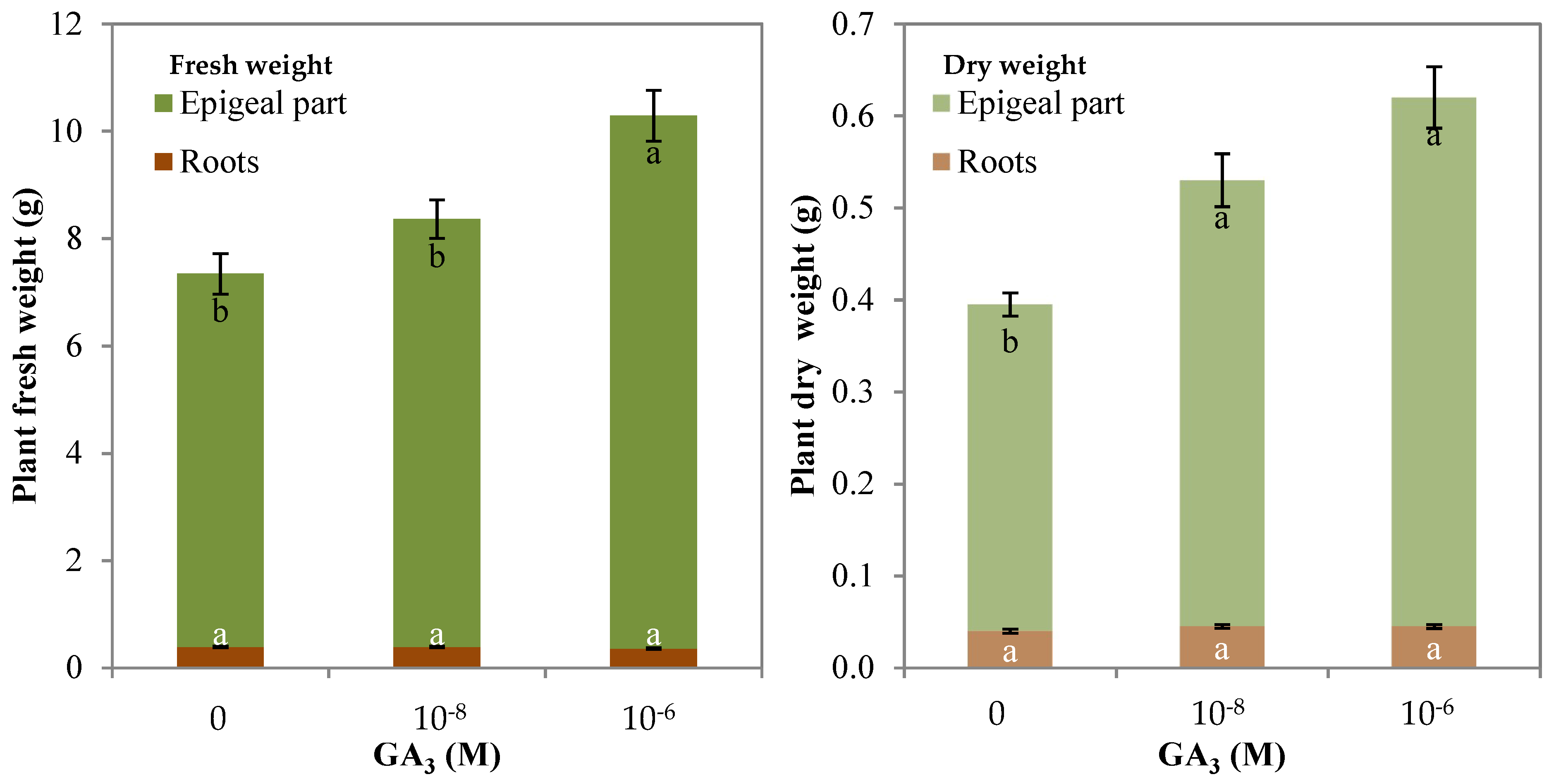
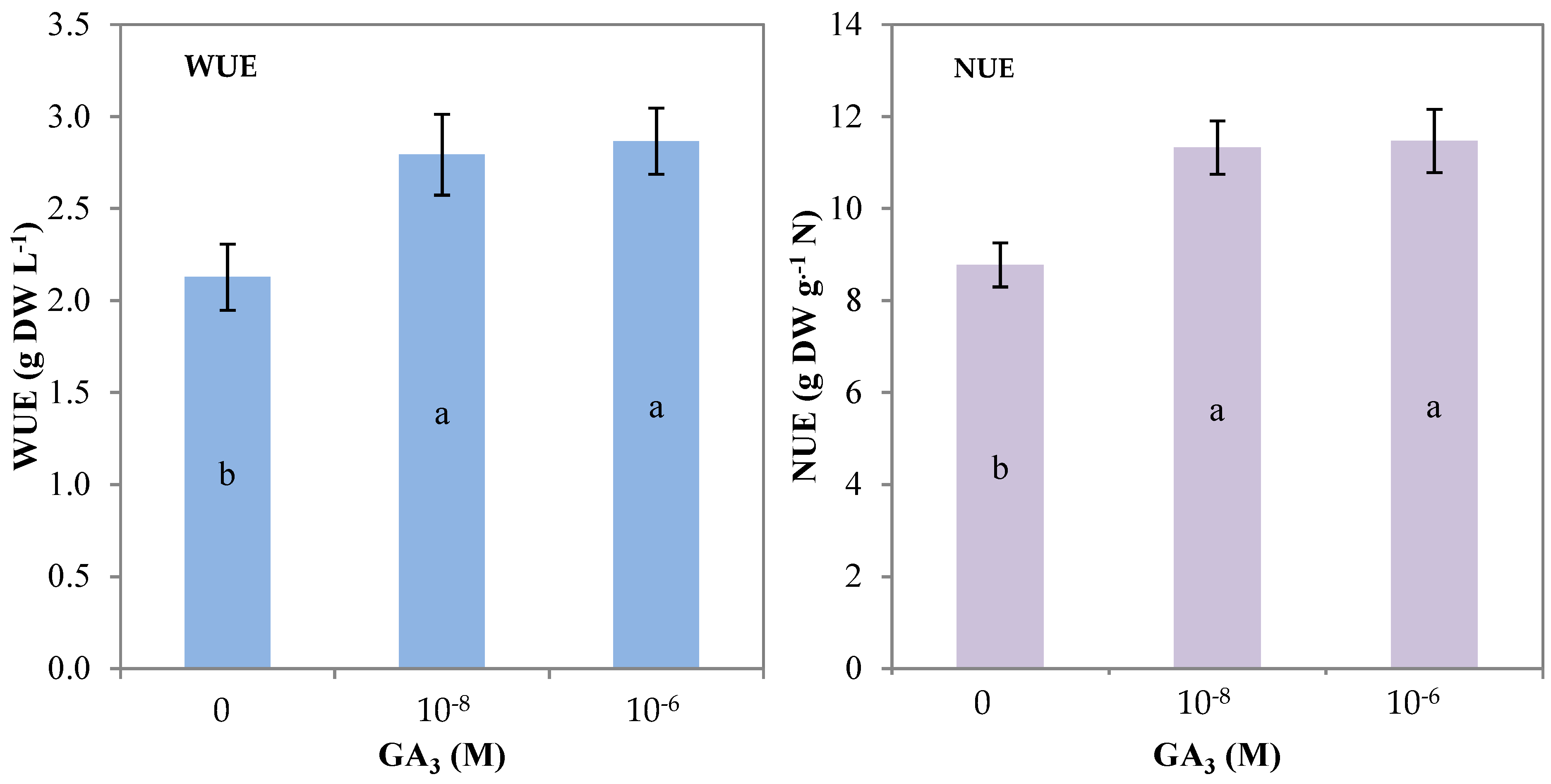
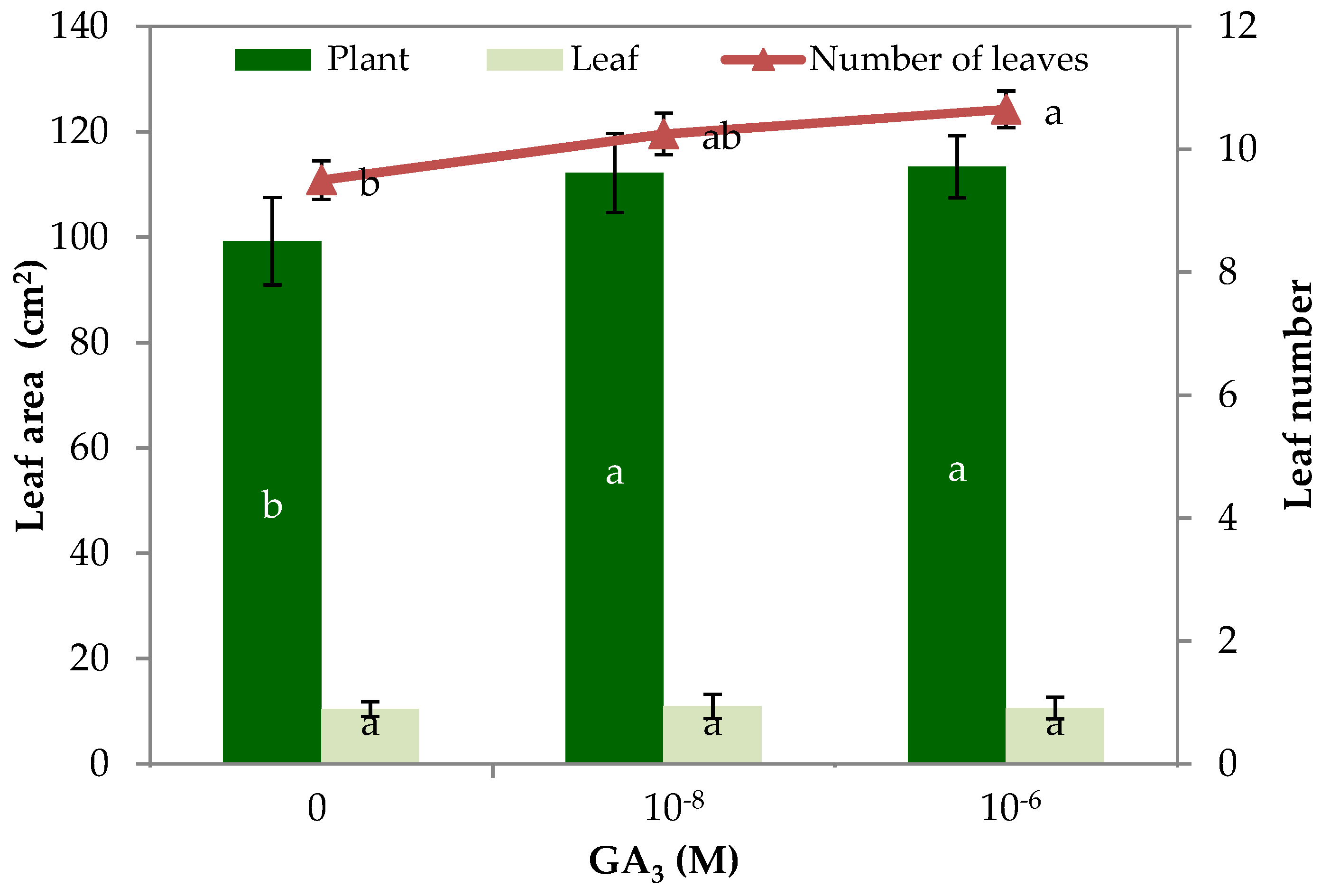
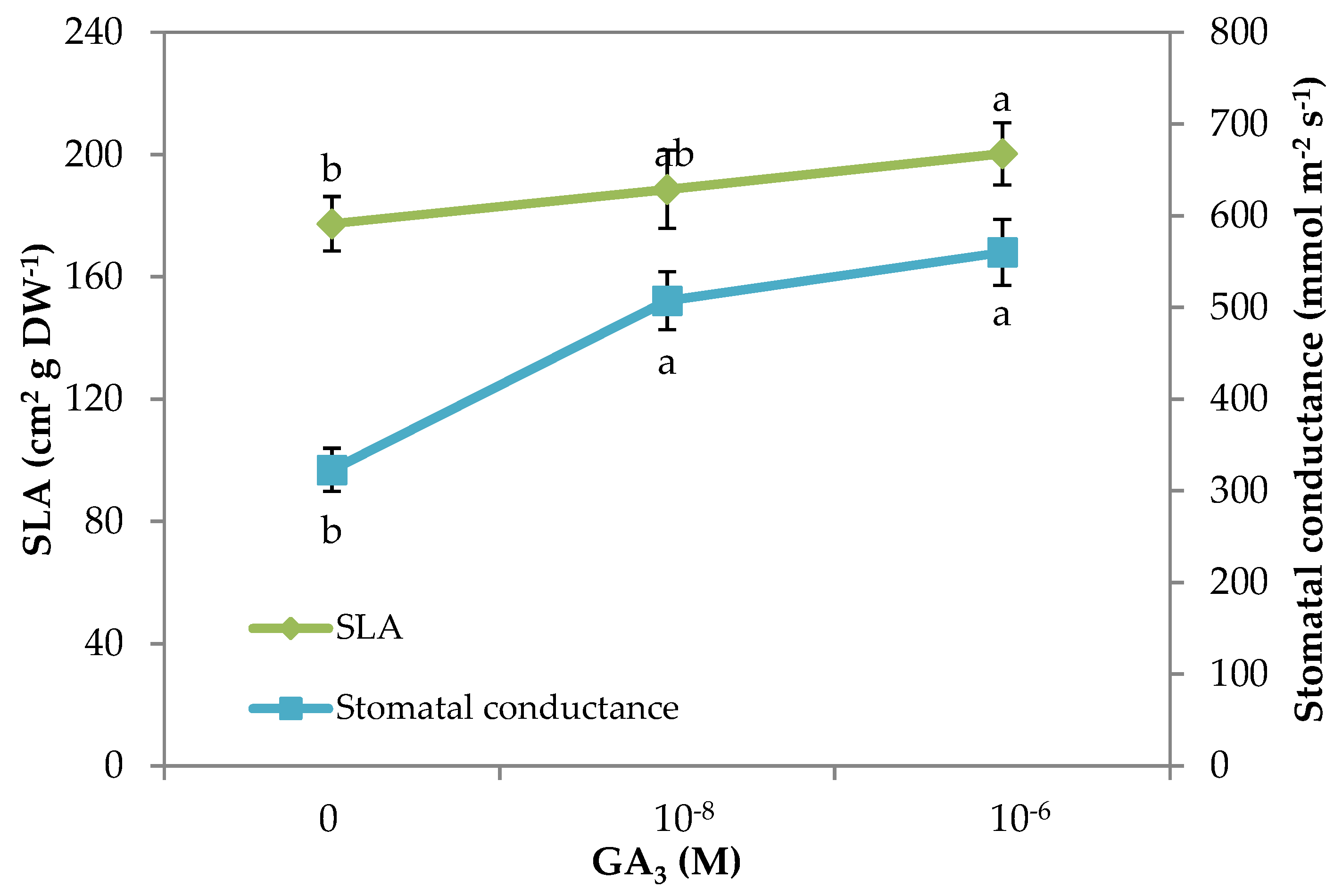
3.2. Morpho–Physiological Parameters and Yield of Rocket
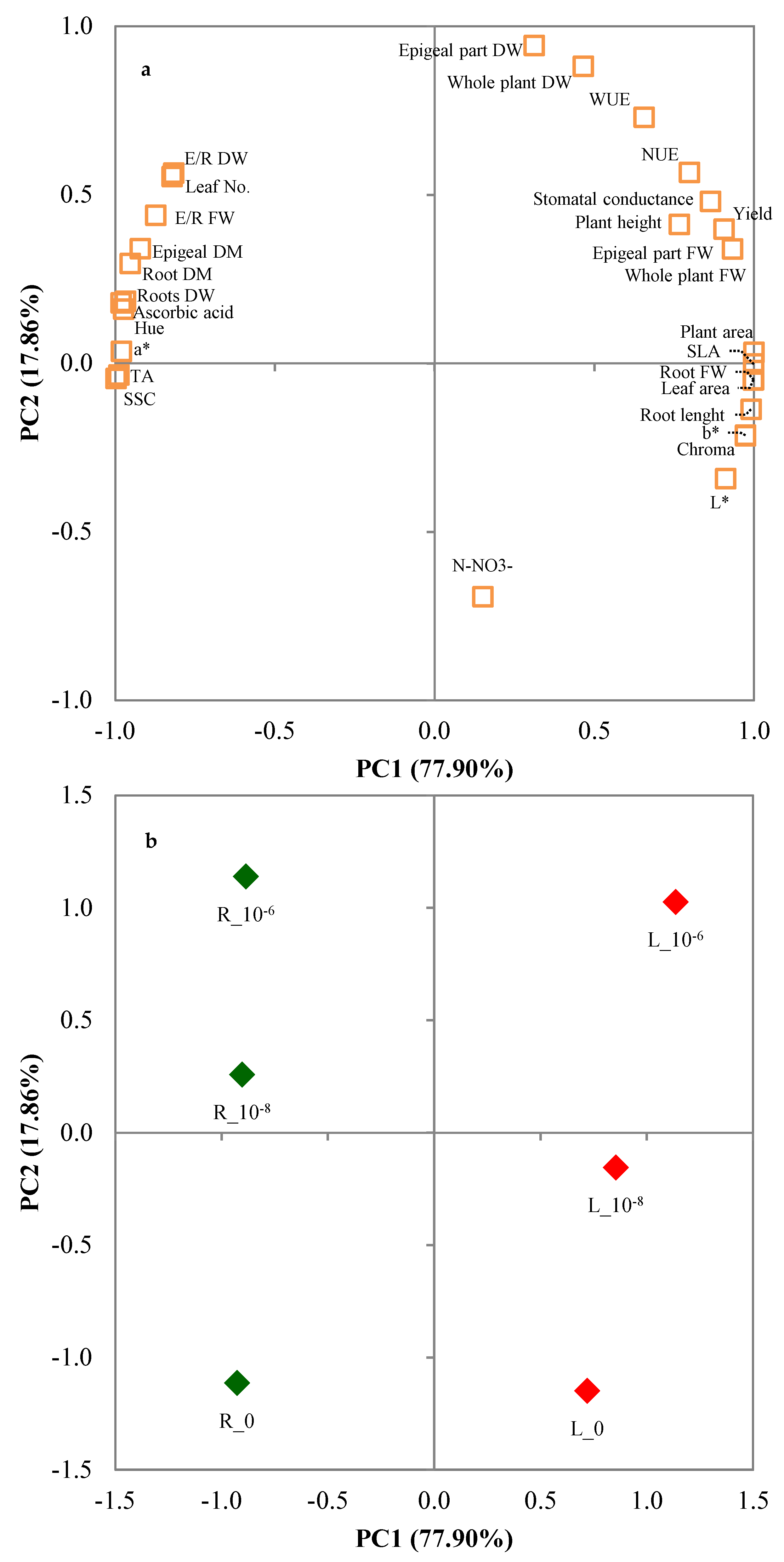
3.3. Principal Components Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Lynch, M.F.; Tauxe, R.V.; Hedberg, C.W. The growing burden of foodborne outbreaks due to contaminated fresh produce: Risks and opportunities. Epidemiol. Infect. 2009, 137, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Settanni, L.; Miceli, A.; Francesca, N.; Cruciata, M.; Moschetti, G. Microbiological investigation of Raphanus sativus L. grown hydroponically in nutrient solutions contaminated with spoilage and pathogenic bacteria. Int. J. Food Microbiol. 2013, 160, 344–352. [Google Scholar] [CrossRef] [PubMed]
- Tomasi, N.; Pinton, R.; Dalla Costa, L.; Cortella, G.; Terzano, R.; Mimmo, T.; Scampicchio, M.; Cesco, S. New ’solutions’ for floating cultivation system of ready-to-eat salad: A review. Trends Food Sci. Technol. 2015, 46, 267–276. [Google Scholar] [CrossRef]
- Khan, I.; Tango, C.N.; Miskeen, S.; Lee, B.H.; Oh, D.-H. Hurdle technology: A novel approach for enhanced food quality and safety—A review. Food Control 2017, 73, 1426–1444. [Google Scholar] [CrossRef]
- Miceli, A.; Settanni, L. Influence of agronomic practices and pre-harvest conditions on the attachment and development of Listeria monocytogenes in vegetables. Ann. Microbiol. 2019, 69, 185–199. [Google Scholar] [CrossRef]
- Soerjomataram, I.; Oomen, D.; Lemmens, V.; Oenema, A.; Benetou, V.; Trichopoulou, A.; Coebergh, J.W.; Barendregt, J.; de Vries, E. Increased consumption of fruit and vegetables and future cancer incidence in selected European countries. Eur. J. Cancer 2010, 46, 2563–2580. [Google Scholar] [CrossRef] [PubMed]
- Karam, M.C.; Petit, J.; Zimmer, D.; Djantou, E.B.; Scher, J. Effects of drying and grinding in production of fruit and vegetable powders: A review. J. Food Eng. 2016, 188, 32–49. [Google Scholar] [CrossRef]
- Chakraborty, I.; Chattopadhyay, A. Pre- and Post-Harvest Losses in Vegetables. In Advances in Postharvest Technologies of Vegetable Crops; Singh, B., Singh, S., Koley, T.K., Eds.; CRC Press Taylor & Francis: Boca Raton, FL, USA, 2018. [Google Scholar]
- Lee, I.J. Practical application of plant growth regulator on horticultural crops. J. Hort. Sci. 2003, 10, 211–217. [Google Scholar]
- Miceli, A.; Sabatino, L.; Moncada, A.; Vetrano, F.; D’Anna, F. Nursery and Field Evaluation of Eggplant Grafted onto Unrooted Cuttings of Solanum torvum Sw. Sci. Hortic. 2014, 178, 203–210. [Google Scholar] [CrossRef]
- Miceli, A.; Romano, C.; Moncada, A.; Piazza, G.; Torta, L.; D’Anna, F.; Vetrano, F. Yield and quality of mini-watermelon as affected by grafting and mycorrhizal inoculum. J. Agr. Sci. Tech. 2016, 18, 505–516. [Google Scholar]
- Moncada, A.; Miceli, A.; Vetrano, F.; Mineo, V.; Planeta, D.; D’Anna, F. Effect of Grafting on Yield and Quality of Eggplant (Solanum melongena L.). Sci. Hortic. 2013, 149, 108–114. [Google Scholar] [CrossRef]
- Rouphael, Y.; Schwarz, D.; Krumbein, A.; Colla, G. Impact of Grafting on Product Quality of Fruit Vegetables. Sci. Hortic. 2010, 127, 172–179. [Google Scholar] [CrossRef]
- Basra, A. Plant Growth Regulators in Agriculture and Horticulture: Their Role and Commercial Uses; CRC Press Inc.: Boca Raton, FL, USA, 2000. [Google Scholar]
- Hooley, R. Gibberellins: Perception, transduction and responses. Plant Mol. Biol. 1994, 26, 1529–1555. [Google Scholar] [CrossRef] [PubMed]
- Hedden, P.; Sponsel, V. A Century of Gibberellin Research. J. Plant Growth Regul. 2015, 34, 740–760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, T.P. Gibberellin Signal Transduction in Stem Elongation & Leaf Growth. In Plant Hormones; Davis, P.J., Ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2004; pp. 308–328. [Google Scholar]
- Rodrigues, T.D.J.; Leite, I.C. Fisiologia vegetal-hormônios das plantas. Jaboticaba 2004, 19–37. [Google Scholar]
- Azuma, T.; Ueno, S.; Uchida, N.; Yasuda, T. Gibberellin-induced elongation and osmoregulation in internodes of floating rice. Physiol. Plant. 1997, 99, 517–522. [Google Scholar] [CrossRef]
- Bhaskar, S.; Vasantha Kumar, T.; Srivastava, H.C. Influence of growth regulators on production of herbage and oil in patchouli (Pogostemon patchouli). Ind. Perfum 1997, 41, 98–101. [Google Scholar]
- Brock, T.G. Combined effects of hormones and light during growth promotion in primary leaves of Phaseolus vulgaris. Can. J. Bot. 1993, 71, 501–550. [Google Scholar] [CrossRef]
- Emongor, V.E. Effect of benzyladenine and gibberellins on growth, yield and yield components of common bean (Phaseolus vulgaris). UNISWA Res. J. Agric. Sci. Technol. 2002, 6, 65–72. [Google Scholar] [CrossRef]
- Gupta, V.N.; Datta, S.K. Influence of gibberellic acid on growth and flowering in chrysanthemum (Chrysanthemum morifolium Ramat) cv. Jayanti. Ind. J. Plant Physiol. 2001, 6, 420–422. [Google Scholar]
- Khan, N.A.; Samiullah, M. Comparative effect of modes of gibberellic acid application on photosynthetic biomass distribution and productivity of rapeseed-mustard. Physiol. Mol. Biol. Plants 2003, 9, 141–145. [Google Scholar]
- Richards, D.E.; King, K.E.; Ait-Ali, T.; Harberd, N.P. How gibberellin regulates plant growth and development: A molecular genetic analysis of gibberellin signaling. Annu. Rev. Plant Biol. 2001, 52, 67–88. [Google Scholar] [CrossRef] [PubMed]
- Takei, K.; Takahashi, T.; Sugiyama, T.; Yamaya, T.; Sakakibara, H. Multiple routes communicating nitrogen availability from roots to shoots: A signal transduction pathway mediated by cytokinin. J. Exp. Bot. 2002, 53, 971–977. [Google Scholar] [CrossRef] [PubMed]
- Wareing, P.F.; Phillips, I.D.J. Growth and Differentiation in Plants, 3rd ed.; Pergamon Press: Oxford, UK, 1981. [Google Scholar]
- Yang, Y.Z.; Lin, D.C.; Guo, Z.Y. Promotion of fruit development in cucumber (Cucumis sativus) by thidiazuron. Sci. Hortic. 1992, 50, 47–51. [Google Scholar] [CrossRef]
- Robert, E.C. Introduction: Nature, Occurrence and Functioning of Plant Hormones. In Biochemistry and Molecular Biology of Plant Hormones; Hooykaas, P.J.J., Hall, M.A., Libbenga, K.R., Eds.; Elsevier: Amsterdam, The Netherlands; New York, NY, USA; London, UK, 1999; pp. 3–22. [Google Scholar]
- Shah, S.H.; Ahmad, I.; Samiullah, M. Effect of gibberellic acid spray on growth.; nutrient uptake and yield attributes during various growth stages of black cumin (Nigella sativa L.). Asian J. Plant Sci. 2006, 5, 881–884. [Google Scholar]
- Janick, J. Horticultural Science; Freeman & Co.: San Francisco, CA, USA, 1979. [Google Scholar]
- Khan, M.M.A.; Gautam, C.; Mohammad, F.; Siddiqui, M.H.; Naeem, M.; Khan, M.N. Effect of gibberellic acid spray on performance of tomato. Turk. J. Biol. 2006, 30, 11–16. [Google Scholar]
- Khan, N.A.; Ansari, H.R. Samiullah Effect of gibberellic acid spray during ontogeny of mustard on growth, nutrient uptake and yield characteristics. J. Agron. Crop Sci. 1998, 181, 61–73. [Google Scholar] [CrossRef]
- King, R.W.; Evans, L.T. Gibberellins and flowering of grasses and cereals: Prizing open the lid of the “florigen black box”. Annu. Rev. Plant Biol. 2003, 54, 307–328. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, R.K.; Prabhakar, B.S. Effect of gibberellin on rice yield response to nitrogen applied at heading and quality of seeds. Plant Soil 1980, 55, 153–156. [Google Scholar] [CrossRef]
- Pal, P.; Yadav, K.; Kumar, K.; Singh, N. Effect of gibberellic acid and potassium foliar sprays on productivity and physiological and biochemical parameters of parthenocarpic cucumber cv. ’seven star F1’. J. Hortic. Res. 2016, 24, 93–100. [Google Scholar] [CrossRef]
- Singh, S.P. Response of tomatoes to growth substances—A review. Adv. Hort. Forest. 1995, 4, 73–84. [Google Scholar]
- Shah, S.H. Effects of salt stress on mustard as affected by gibberellic acid application. Gen Appl. Plant Physiol. 2007, 33, 97–106. [Google Scholar]
- Maggio, A.; Barbieri, G.; Raimondi, G.; De Pascale, S. Contrasting effects of GA3 treatments on tomato plants exposed to increasing salinity. J. Plant Growth Regul. 2010, 29, 63–72. [Google Scholar] [CrossRef]
- Khan, N.A.; Mir, R.; Khan, M.; Javid, S. Effects of gibberellic acid spray on nitrogen yield efficiency of mustard grown with different nitrogen levels. Plant Growth Regul. 2002, 38, 243–247. [Google Scholar] [CrossRef]
- Gelmesa, D.; Abebie, B.; Desalegn, L. Effects of Gibberellic acid and 2, 4-dichlorophenoxyacetic acid spray on fruit yield and quality of tomato (Lycopersicon esculentum Mill.). J. Plant Breed. Crop Sci. 2010, 2, 316–324. [Google Scholar]
- Sonneveld, C.; Voogt, W. Plant Nutrition of Greenhouse Crops; Springer: New York, NY, USA, 2009. [Google Scholar]
- Goto, E.; Both, A.J.; Albright, L.D.; Langhans, R.W.; Leed, A.R. Effect of dissolved oxygen concentration on lettuce growth in floating hydroponics. Acta Hortic. 1996, 440, 205–210. [Google Scholar] [CrossRef] [PubMed]
- Fageria, N.K.; Baligar, V.C.; Li, Y.C. The role of nutrient efficient plants in improving crop yields in the twenty first century. J. Plant Nutr. 2008, 31, 1121–1157. [Google Scholar] [CrossRef]
- McGuire, R.G. Reporting of objective color measurements. Hortic. Sci. 1992, 27, 1254–1255. [Google Scholar] [CrossRef]
- Da Silva Vieira, M.R.; Citadini, V.; Lima, G.P.P.; de Souza, A.V.; de Souza Alves, L. Use of gibberellin in floriculture. Afr. J. Biotechnol. 2010, 9, 9118–9121. [Google Scholar]
- Dayan, J.; Voronin, N.; Gong, F.; Sun, T.; Hedden, P.; Fromm, H.; Aloni, R. Leaf-induced gibberellin signaling is essential for internode elongation, cambial activity, and fiber differentiation in tobacco stems. Plant Cell 2012, 24, 66–79. [Google Scholar] [CrossRef]
- Vivanco, J.M.; Flores, H.E. Control of Root Formation by Plant Growth Regulators. In Plant Growth Regulators in Agriculture and Horticulture: Their Role and Commercial Uses; Basra, A., Ed.; CRC Press Inc.: Boca Raton, FL, USA, 2000; pp. 1–26. [Google Scholar]
- Buchanan, B.B.; Gruissem, W.; Jones, R.L. Biochemistry and Molecular Biology of Plants; American Society of Plant Physiologists: Rockville, MD, USA, 2000. [Google Scholar]
- Arteca, R.N. Plant Growth Substances: Principles and Applications; Chapman and Hall Inc.: New York, NY, USA, 1996. [Google Scholar]
- Liu, P.B.W.; Loy, B. Action of gibberellic acid on cell proliferation in the subapical shoot meristem of watermelon seedlings. Am. J. Bot. 1976, 63, 700–704. [Google Scholar] [CrossRef]
- Moore, T.C. Biochemistry and Physiology of Plant Hormones; Springer-Verlag Inc.: New York, NY, USA, 1989. [Google Scholar]
- Ogasawara, N.; Hiramasu, T.; Ishiyama, K.; Fushimi, H.; Suzuki, H.; Takagi, H. Effects of gibberellic acid and temperature on growth and root carbohydrates of Delphinium seedlings. Plant Growth Regul. 2001, 33, 181–187. [Google Scholar] [CrossRef]
- King, R.W.; Moritz, T.; Evans, L.T.; Junttila, O.; Herlt, A.J. Long-day induction of flowering in Lolium temulentum involves sequential increases in specific gibberellins at the shoot apex. Plant Physiol. 2001, 127, 624–632. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, S.; Böhlenius, H.; Moritz, T.; Nilsson, O. GA4 is the active gibberellin in the regulation of LEAFY transcription and Arabidopsis floral initiation. Plant Cell 2006, 18, 2172–2181. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, S.; Lin, J.; Wang, Y.; Fang, X. Research progress on cloning and functional analysis of plant height genes in rice (Oryza sativa L.). Chin. Agr. Sci. Bull. 2014, 30, 78–89. [Google Scholar]
- Achard, P.; Gusti, A.; Cheminant, S.; Alioua, M.; Dhondt, S.; Coppens, F.; Beemster, G.T.; Genschik, P. Gibberellin signaling controls cell proliferation rate in Arabidopsis. Curr. Biol. 2009, 19, 1188–1193. [Google Scholar] [CrossRef]
- Ubeda-Tomás, S.; Federici, F.; Casimiro, I.; Beemster, G.T.; Bhalerao, R.; Swarup, R.; Doerner, P.; Haseloff, J.; Bennett, M.J. Gibberellin signaling in the endodermis controls Arabidopsis root meristem size. Curr. Biol. 2009, 19, 1194–1199. [Google Scholar] [CrossRef] [PubMed]
- Broughton, W.J.; McComb, A.J. Changes in the pattern of enzyme development in gibberellin-treated pea internodes. Ann. Bot. 1971, 35, 213–228. [Google Scholar] [CrossRef]
- Johri, M.M.; Varner, J.E. Enhancement of RNA synthesis in isolated pea nuclei by gibberellic acid. Proc. Natl. Acad. Sci. USA 1968, 59, 269–279. [Google Scholar] [CrossRef]
- Mozer, T.J. Control of protein synthesis in barley aleurone layers by the plant hormones, gibberellic acid and abscisic acid. Cell 1980, 20, 479–485. [Google Scholar] [CrossRef]
- Pain, S.K.; Dutta, J.K. Studies on growth and metabolism of Zea mays L. I. The effect of application of gibberellic acid on the growth and metabolism of seedlings. Indian Biol. 1977, 9, 38–43. [Google Scholar]
- Roth Benjerano, N.; Lips, S.H. Hormonal regulation of nitrate reductase activity in leaves. New Phytol. 1970, 69, 165–169. [Google Scholar] [CrossRef]
- Evins, W.H.; Varner, J.E. Hormonal control of polyribosome formation in barley aleurone layers. Plant Physiol. 1972, 49, 348–352. [Google Scholar] [CrossRef] [PubMed]
- Broughton, W.J. Influence of gibberellic acid on nucleic acid synthesis in dwarf pea internodes. Biochem. Biophys. Acta 1968, 155, 308–310. [Google Scholar] [CrossRef]
- Glasziou, K.T. Control of enzyme formation and inactivation in plants. Annu. Rev. Plant Physol. 1969, 20, 63–88. [Google Scholar] [CrossRef]
- Seitz, K.; Lang, A. Invertase activity and cell growth in lentil epicotyls. Plant Physiol. 1968, 43, 1075–1082. [Google Scholar] [CrossRef] [PubMed]
- Crozier, A.; Turnbull, C.G.N. Gibberellins: Biochemistry and action in extension growth. What’s New in Plant Physiol. 1984, 15, 9–12. [Google Scholar]
- Wood, A.; Paleg, L.G. The influence of GA3 on the permeability of model membrane systems. Plant Physiol. 1972, 50, 103–108. [Google Scholar] [CrossRef]
- Wood, A.; Paleg, L.G. Alteration of liposomal membrane fluidity by gibberellic acid. Aust. J. Plant Physiol. 1974, 1, 31–40. [Google Scholar] [CrossRef]
- Al-Wakeel, S.A.M.; Hamed, A.A.; Dadoura, S.S. Interactive effects of water stress and gibberellic acid on mineral composition of fenugreek plant. Egypt. J. Physiol. Sci. 1995, 18, 269–282. [Google Scholar]
- Ansari, H.R. Effect of Some Phytohormones and NPK on Growth and Metabolism of Mustard. Ph.D. Thesis, ; Aligarh Muslim University, Aligarh, India, 1996. [Google Scholar]
- Aloni, B.; Daie, J.; Wyse, R.E. Enhancement of (14C)-sucrose export from source leaves of Vicia faba by gibberellic acid. Plant Physiol. 1986, 82, 962–966. [Google Scholar] [CrossRef] [PubMed]
- Daie, J.; Watts, M.; Aloni, B.; Wyse, R.E. In vitro and in vivo modification of sugar transport and translocation in celery by phytohormones. Plant Sci. 1986, 46, 35–41. [Google Scholar] [CrossRef]
- Estruch, J.J.; Pereto, J.G.; Vercher, Y.; Beltrán, J.P. Sucrose loading in isolated veins of Pisum sativum: Regulation by abscisic acid, gibberellic acid and cell turgor. Plant Physiol. 1989, 91, 259–265. [Google Scholar] [CrossRef] [PubMed]
- Mulligan, D.R.; Patrick, J.W. Gibberellic-acid-promoted transport of assimilates in stems of Phaseolus vulgaris L.: Localised versus remote site (S) of action. Planta 1979, 145, 233–238. [Google Scholar] [CrossRef] [PubMed]
- Hedden, P.; Thomas, S.G. Gibberellin biosynthesis and its regulation. Biochem. J. 2012, 444, 11–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugiura, D.; Kojima, M.; Sakakibara, H. Phytohormonal Regulation of Biomass Allocation and Morphological and Physiological Traits of Leaves in Response to Environmental Changes in Polygonum cuspidatum. Front. Plant Sci. 2016, 7, 1189. [Google Scholar] [CrossRef] [PubMed]
- Sugiura, D.; Sawakami, K.; Kojima, M.; Sakakibara, H.; Terashima, I.; Tateno, M. Roles of gibberellins and cytokinins in regulation of morphological and physiological traits in Polygonum cuspidatum responding to light and nitrogen availabilities. Funct. Plant Bio. 2015, 42, 397–409. [Google Scholar] [CrossRef]
- Bultynck, L.; Lambers, H. Effects of applied gibberellic acid and paclobutrazol on leaf expansion and biomass allocation in two Aegilops species with contrasting leaf elongation rates. Physiol. Plant. 2004, 122, 143–151. [Google Scholar] [CrossRef]
- Tang, X.M.; Liu, C.; Zhong, R.C.; Jiang, J.; Han, Z.Q.; He, L.Q.; Li, Z.; Xiong, F.Q.; Tang, R.H. Comparative studies on the chemical regulation of peanuts by paclobutrazol, mepiquat chloride and chlorocholine chloride. J. South. Agric. 2011, 42, 603–605. [Google Scholar]
- Xie, B.T.; Wang, Q.M.; Zhang, H.Y.; Li, A.X.; Hou, F.Y.; Wang, B.Q.; Dong, S.X.; Zhang, L.M. The effect of plant growth regulators on the yield and phytohormone concentration in sweet potato. Acta Agric. Boreali-Sin. 2016, 31, 155–161. [Google Scholar]
- Zhang, Y. Research status of plant growth regulators on crop regulation. Mod. Agric. 2016, 5, 31–34. [Google Scholar]
- Han, H.Y.; Wang, F.A.; Chen, B.; Li, B.A.; Zhang, W.F.; Tian, X.L.; Li, Z. Effect of nitrogen fertilizer on plant growth and yield formation of cotton applied with fortified DPC. J. Chin. Agric. Univ. 2017, 22, 12–20. [Google Scholar]
- Zhang, M.; Liu, M.; Zhang, Y.; Ji, Y.; Zhao, M.; Wu, Z. Effect of different plant growth regulator added in nutrient solution on growth and development of summer tomato seedling. North Hortic. 2017, 6, 8–13. [Google Scholar]
- Khan, N.A. Effect of gibberellic acid on carbonic anhydrase, photosynthesis, growth and yield of mustard. Biol. Plant. 1996, 38, 145. [Google Scholar] [CrossRef]
- Sultemeyer, D.; Schmidt, C.; Fock, H.P. Carbonic anhydrases in higher plants and aquatic microorganisms. Physiol. Plant. 1993, 88, 179–190. [Google Scholar] [CrossRef]
- Kachru, R.B.; Anderson, L.E. Chloroplast and cytoplasmic enzymes. Planta 1974, 118, 235–240. [Google Scholar] [CrossRef]
- Khan, N.A. Variation in carbonic anhydrase activity and its relationship with photosynthesis and dry mass of mustard. Photosynthetica 1994, 30, 317–320. [Google Scholar]
- Yuan, L.; Xu, D.Q. Stimulation effect of gibberellic acid short-term treatment on leaf photosynthesis related to the increase in Rubisco content in broad bean and soybean. Photosynth. Res. 2001, 68, 39–47. [Google Scholar] [CrossRef]
- Ihl, M.; Shene, C.; Scheuermann, E.; Bifani, V. Correlation for pigment content through color determination using tristimulus values in a green leafy vegetable, Swiss chard. J. Sci. Food Agric. 1994, 66, 527–531. [Google Scholar] [CrossRef]
- Madeira, A.C.; Ferreira, A.; de Varennes, A.; Vieira, M.I. SPAD meter versus tristimulus colorimeter to estimate chlorophyll content and leaf color in sweet pepper. Commun. Soil Sci. Plant Anal. 2003, 34, 2461–2470. [Google Scholar] [CrossRef]
- Magdah, G.M. Impact of biostimulant and synthetic hormone gibberellic acid on molecular structure of Solanum melongena L. J. Mol. Biol. Res. 2016, 6, 100–110. [Google Scholar] [CrossRef]
- Mbandlwa, N.P.; Fotouo, M.H.; Maboko, M.M.; Sivakumar, D. Stomatal conductance, leaf chlorophyll content, growth, and yield of sweet pepper in response to plant growth regulators. Int. J. Veg. Sci. 2019, 1–11. [Google Scholar] [CrossRef]
- Wheeler, A.W.; Humphies, E.C. Effect of gibberellic acid on growth, gibberellin content, and chlorophyll content of leaves of potato (Solanum tuberosum). J. Exp. Bot. 1963, 14, 132–136. [Google Scholar] [CrossRef]
- Mahindre, P.B.; Jawarkar, A.K.; Ghawade, S.M.; Tayade, V.D. Effect of different concentration of plant growth regulators on growth and quality of green chilli. J. Pharm. Phytochem. 2018, 1, 3040–3042. [Google Scholar]
- Göring, H.; Koshuchowa, S.; Deckert, C. Influence of gibberellic acid on stomatal movement. Bioche. Physiol. Pflanz. 1990, 186, 367–374. [Google Scholar] [CrossRef]
- Chanda, S.V.; Sood, C.R.; Reddy, V.S.; Singh, Y.D. Influence of plant growth regulators on some enzymes of nitrogen assimilation in mustard seedlings. J. Plant Nutr. 1998, 21, 1765–1777. [Google Scholar] [CrossRef]
- Lee, S.K.; Kader, A.A. Preharvest and postharvest factors influencing vitamin C content of horticultural crops. Postharvest Biol. Technol. 2000, 20, 207–220. [Google Scholar] [CrossRef] [Green Version]
- Stefanelli, D.; Goodwin, I.; Jones, R. Minimal nitrogen and water use in horticulture, effects on quality and content of selected nutrients. Food Res. Int. 2010, 43, 1833–1843. [Google Scholar] [CrossRef]
- Miceli, A.; Miceli, C. Effect of Nitrogen Fertilization on the Quality of S wiss Chard at Harvest and during Storage as Minimally Processed Produce. J. Food Qual. 2014, 37, 125–134. [Google Scholar] [CrossRef]
- Mozafar, A. Plant Vitamins. Agronomic, Physiological and Nutritional Aspects; CRC Press Inc.: Boca Raton, FL, USA, 1994. [Google Scholar]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GA3 (M) | |||
|---|---|---|---|
| 0 | 10−8 | 10−6 | |
| Plant height (cm) | 25.0 b 1 | 25.2 b | 31.8 a |
| Root length (cm) | 31.1 a | 30.7 a | 32.0 a |
| Plant fresh weight | |||
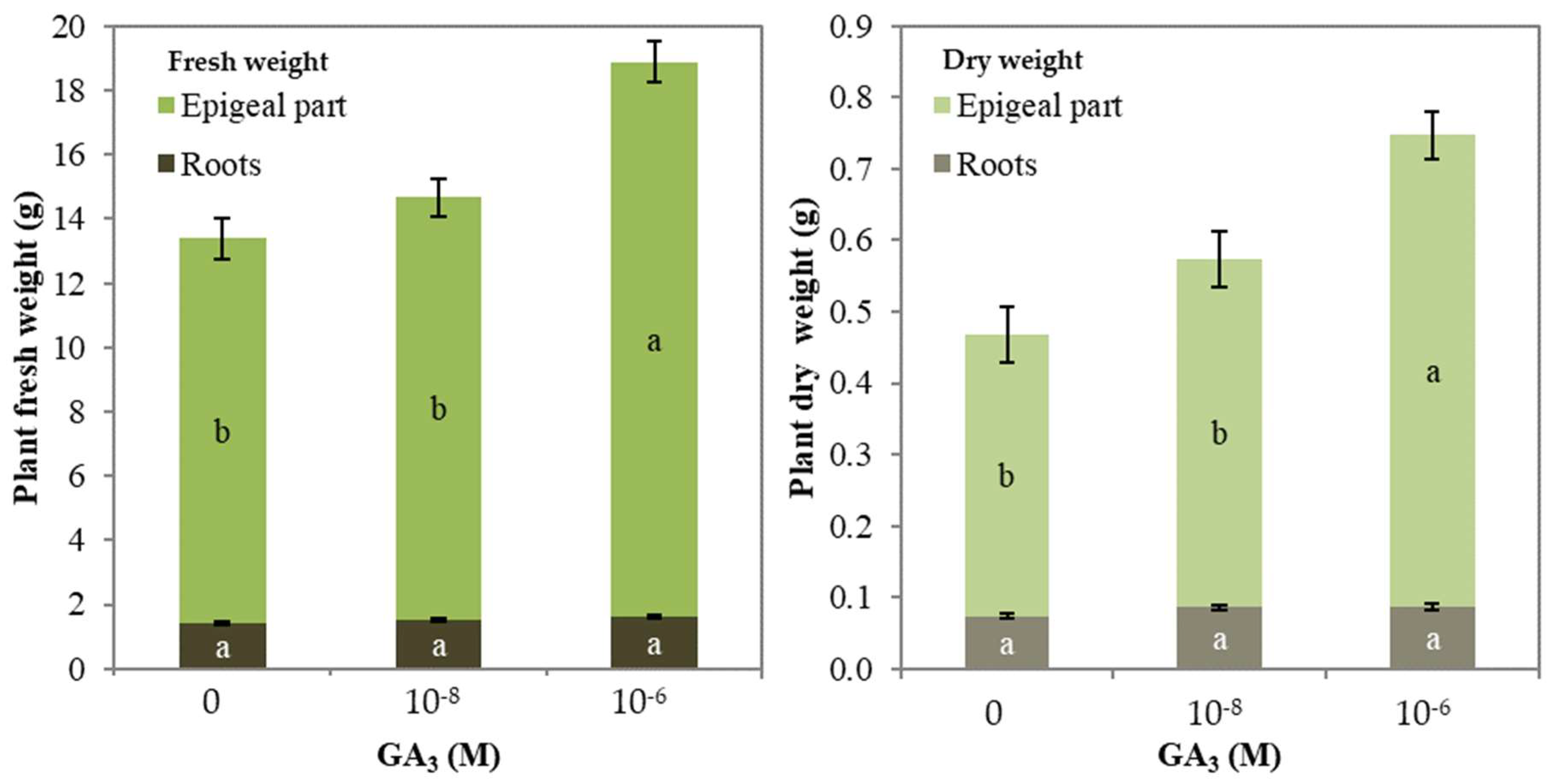
| Whole plant (g) | 13.4 b | 14.7 b | 18. 9a |
| Epigeal part (g) | 12.0 b | 13.2 b | 17.3 a |
| Roots (g) | 1.4 a | 1.5 a | 1.6 a |
| Epigeal/Roots Ratio | 8.3 b | 8.8 b | 10.7 a |
| Plant dry weight | |||
| Whole plant (g) | 0.47 b | 0.57 b | 0.75 a |
| Epigeal part (g) | 0.39 b | 0.49 b | 0.66 a |
| Roots (g) | 0.07 a | 0.09 a | 0.09 a |
| Epigeal/Roots Ratio | 5.3 b | 5.7 b | 7.6 a |
| Epigeal dry matter (%) | 3.3 b | 3.7 a | 3.8 a |
| Root dry matter (%) | 5.2 b | 5.4 ab | 5.7 a |
| Yield (kg m−2) | 4.8 b | 5.3 b | 6.9 a |
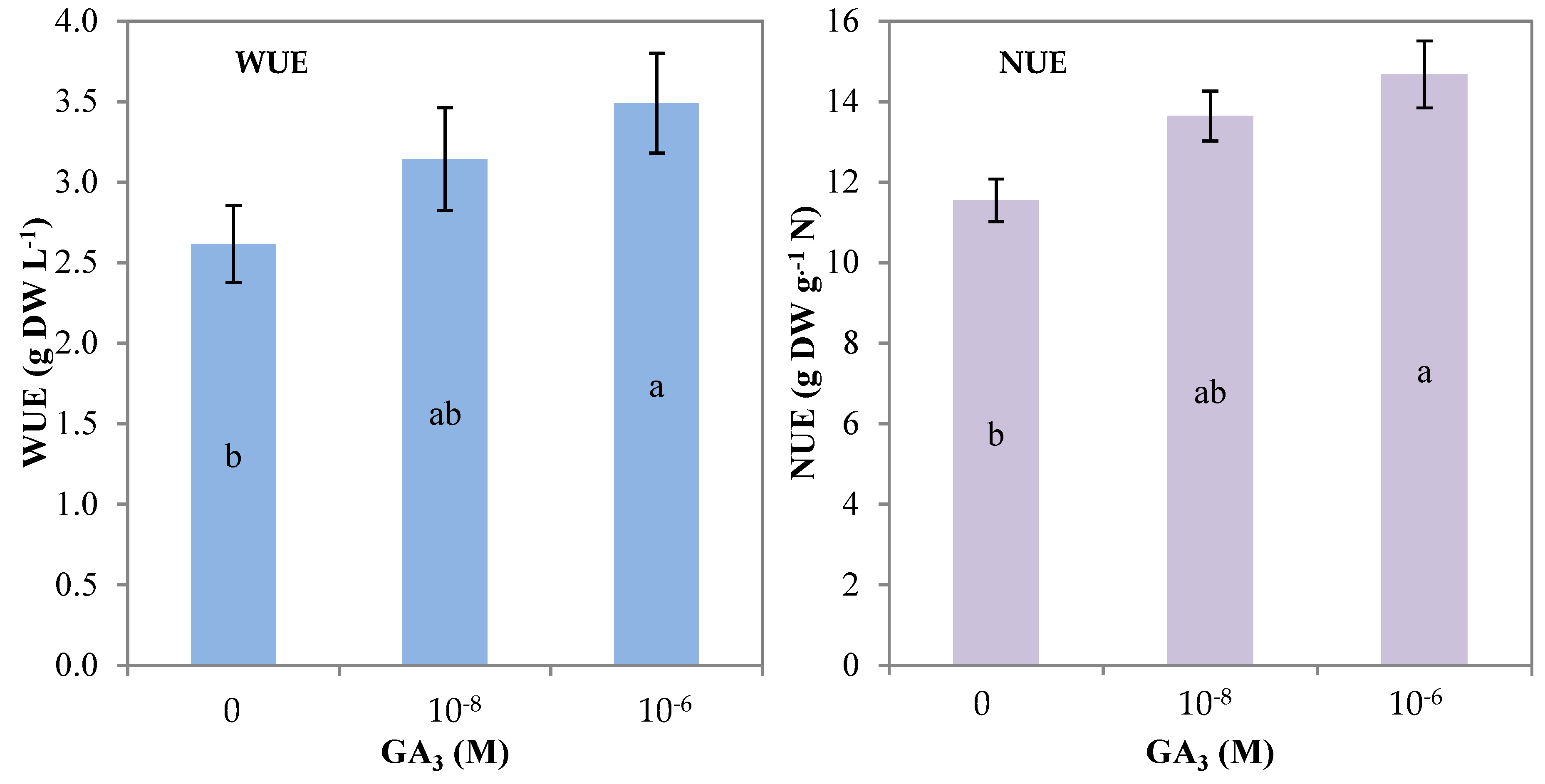
| WUE (g DW L−1 H2O) | 2.6 b | 3.1 ab | 3.5 a |
| NUE (g DW g−1 N) | 11.6 b | 13.6 ab | 14.7 a |
| GA3 (M) | |||
|---|---|---|---|
| 0 | 10−8 | 10−6 | |
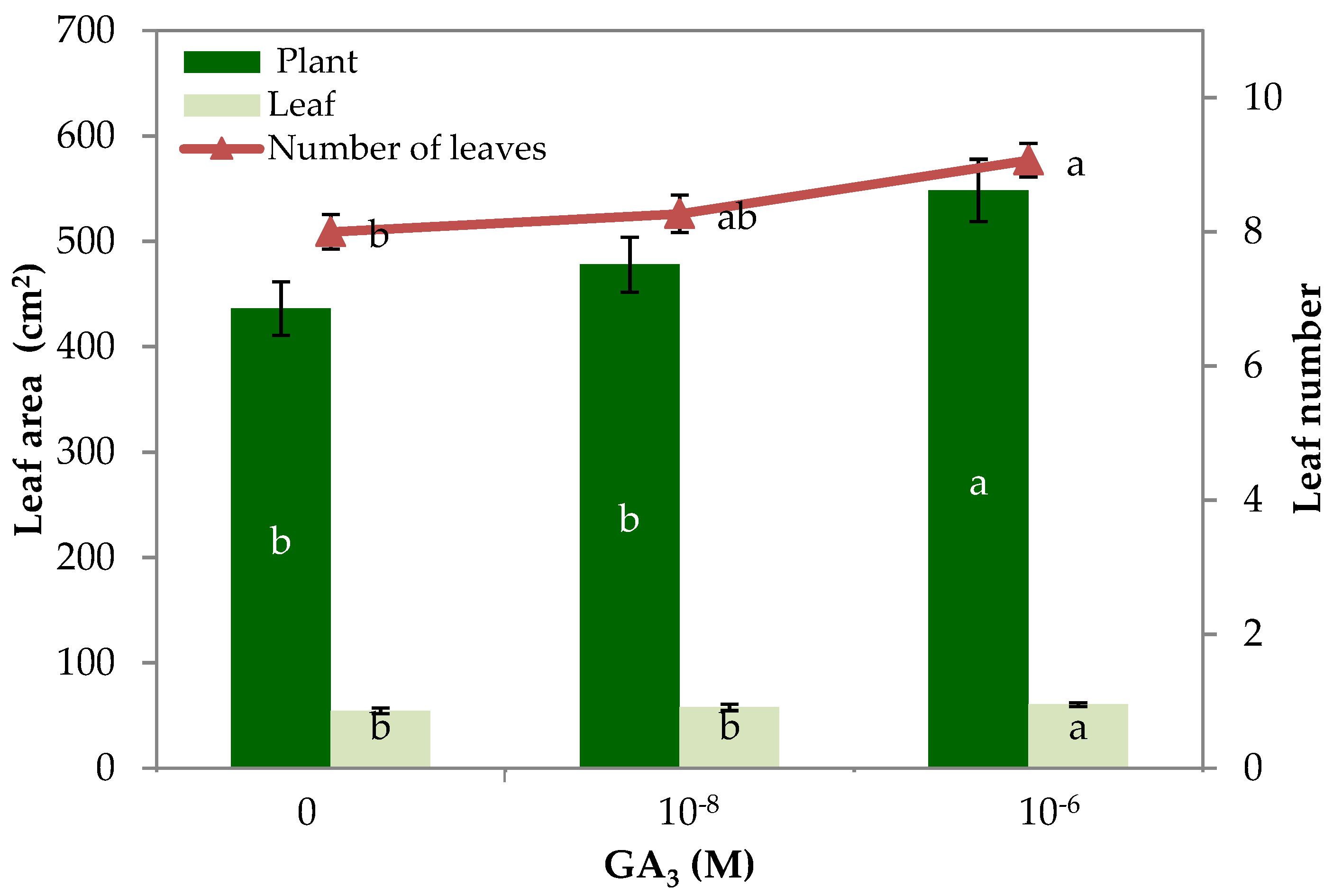
| Number of leaves | 8.0 b 1 | 8.3 ab | 9.1 a |
| Leaf width (cm) | 8.8 a | 9.4 a | 8.7 a |
| Leaf area (cm2 plant−1) | 436.1 b | 477.8 b | 548.3 a |
| Leaf area (cm2 leaf−1) | 54. 5 b | 57.8 b | 60.5 a |
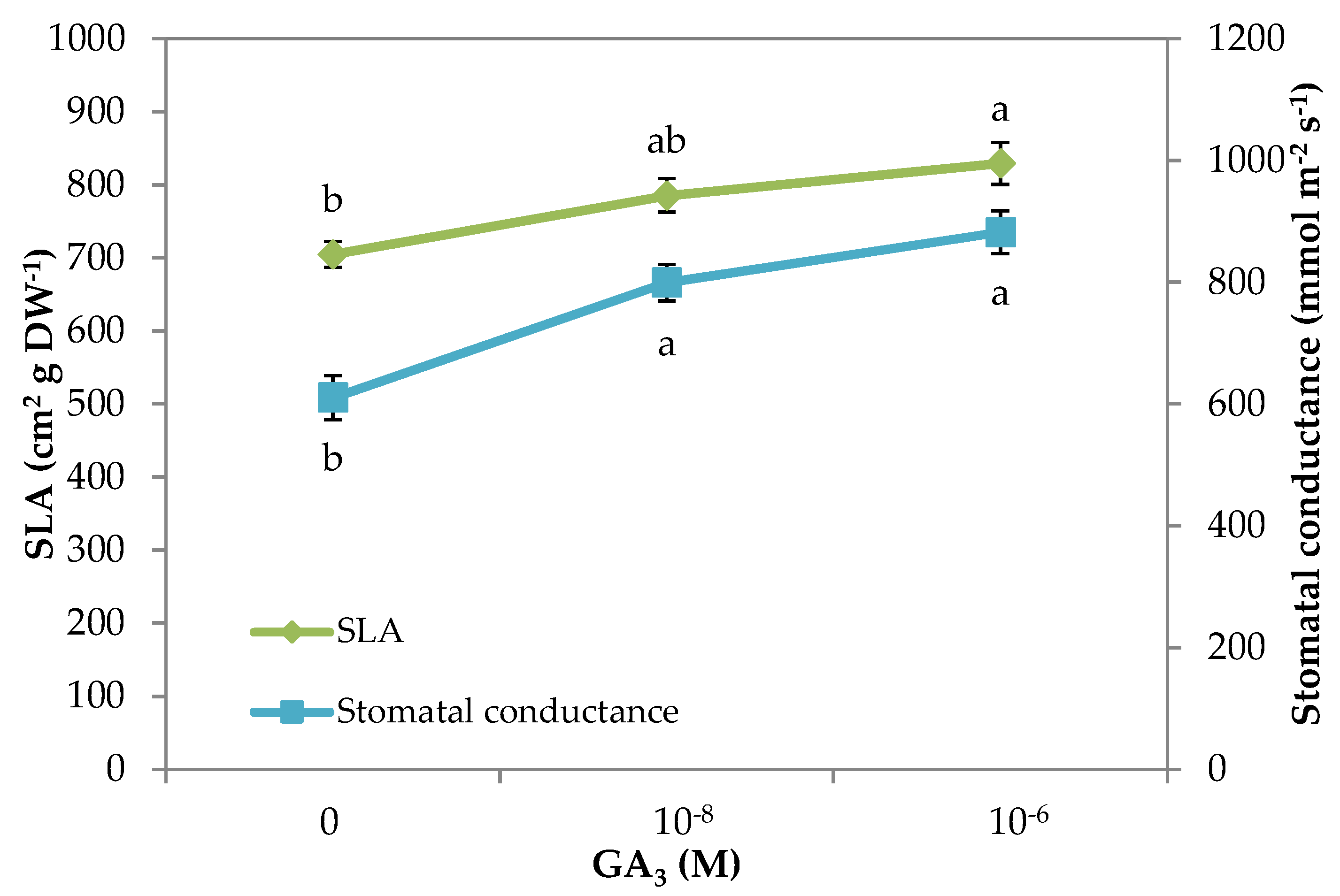
| Specific Leaf Area (cm2 g DW−1) | 704.5 b | 785.2 ab | 829.1 a |
| Stomatal conductance (mmol m−2 s−1) | 610.0 b | 799.0 a | 882.0 a |
| L* | 54.8 a | 51.0 b | 50.9 b |
| a* | -21.7 a | -22.0 a | -21.7 a |
| b* | 39.0 a | 38.2 ab | 37.2 b |
| Chroma | 44.7 a | 44.1 ab | 43.1 b |
| Hue° | 119.1 b | 120.0 ab | 120.3 a |
| Soluble solid content (°Brix) | 2.9 a | 3.1 a | 2.5 a |
| Titratable acidity (mg 100 g−1 FW) 2 | 30.7 a | 28.8 a | 25.9 a |
| Ascorbic Acid (mg 100 g−1 FW) | 78.5 a | 53.0 a | 58.0 a |
| N-NO3− (mg kg−1 FW) | 2535.0 a | 2305.0 a | 2505.0 a |
| GA3 (M) | |||
|---|---|---|---|
| 0 | 10−8 | 10−6 | |
| Plant height (cm) | 21.9 b 1 | 23.1 ab | 24.0 a |
| Root length (cm) | 21.0 a | 19.9 a | 19.1 a |
| Plant fresh weight | |||
| Whole plant (g) | 7.3 b | 8.4 b | 10.3 a |
| Epigeal part (g) | 7.0 b | 8.0 b | 9.9 a |
| Roots (g) | 0.39 a | 0.39 a | 0.36 a |
| Ratio Epigeal/Roots | 17.9 b | 20.5 b | 27.9 a |
| Plant dry weight | |||
| Whole plant (g) | 0.39 b | 0.53 a | 0.62 a |
| Epigeal part (g) | 0.36 b | 0.49 a | 0.58 a |
| Roots (g) | 0.04 a | 0.05 a | 0.05 a |
| Ratio Epigeal/Roots | 9.0 b | 10.8 ab | 12.9 a |
| Epigeal dry matter (%) | 5.1 b | 6.1 a | 5.8 a |
| Root dry matter (%) | 10.2 b | 11.6 ab | 12.7 a |
| Yield (kg m−2) | 2.8 b | 3.2 ab | 4.0 a |
| WUE (g DW L−1 H2O) | 2.1 b | 2.8 a | 2.9 a |
| NUE (g DW g−1 N) | 8.8 b | 11.3 a | 11.5 a |
| GA3 (M) | |||
|---|---|---|---|
| 0 | 10−8 | 10−6 | |
| Number of leaves | 9.5 b 1 | 10.3 ab | 10.7 a |
| Leaf blade length (cm) | 11.1 a | 11.6 a | 11.6 a |
| Petiole length (cm) | 8.7 a | 8.8 b | 10.2 a |
| Leaf area (cm2 plant−1) | 99.2 b | 112.2 a | 113.3 a |
| Leaf area (cm2 leaf−1) | 10.4 a | 10.9 a | 10.6 a |
| Specific Leaf Area (cm2 g−1 DW) | 177.4 b | 188.6 ab | 200.3 a |
| Stomatal conductance (mmol m−2 s−1) | 322.9 b | 507.4 a | 560.1 a |
| L* | 44.7 a | 43.5 ab | 43.1 b |
| a* | -19.4 b | -19.1 ab | -18.6 a |
| b* | 26.2 a | 25.5 a | 24.3 a |
| Chroma | 32.6 a | 31.8 a | 30.6 a |
| Hue° | 126.8 a | 127.2 a | 127.5 a |
| Soluble solid content (°Brix) | 5.8 a | 5.2 a | 5.4 a |
| Titratable acidity (mg 100 g−1 FW) 2 | 46.1 a | 44.8 a | 44.8 a |
| Ascorbic Acid (mg 100 g−1 FW) | 173.0 b | 221.0 a | 218.0 a |
| N-NO3− (mg kg−1 FW) | 2714.9 a | 2364.9 ab | 2069.9 b |
| Variables | PC1 | PC2 |
|---|---|---|
| Plant height | 0.766 | 0.412 |
| Root length | 0.990 | −0.137 |
| Whole plant fresh weight | 0.932 | 0.339 |
| Epigeal part fresh weight | 0.905 | 0.398 |
| Root fresh weight | 0.999 | −0.049 |
| E/R fresh weight | −0.874 | 0.439 |
| Whole plant dry weight | 0.465 | 0.881 |
| Epigeal part dry weight | 0.311 | 0.943 |
| Roots dry weight | −0.982 | 0.179 |
| E/R dry weight | −0.823 | 0.554 |
| Epigeal dry weight | −0.922 | 0.340 |
| Root dry weight | −0.954 | 0.296 |
| Yield | 0.905 | 0.398 |
| Water Use Efficiency | 0.655 | 0.729 |
| Nitrogen Use Efficiency | 0.797 | 0.566 |
| Leaf No. | −0.818 | 0.564 |
| Plant area | 0.999 | 0.032 |
| Leaf area | 0.998 | −0.050 |
| Specific Leaf Area | 0.999 | −0.002 |
| Stomatal conductance | 0.863 | 0.480 |
| L* | 0.911 | −0.342 |
| a* | −0.982 | 0.035 |
| b* | 0.973 | −0.213 |
| Chroma | 0.973 | −0.216 |
| Hue | −0.975 | 0.160 |
| Soluble solid content | −0.989 | −0.035 |
| Titratable acidity | −0.999 | −0.045 |
| Ascorbic acid | −0.968 | 0.184 |
| N-NO3− | 0.151 | −0.693 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miceli, A.; Moncada, A.; Sabatino, L.; Vetrano, F. Effect of Gibberellic Acid on Growth, Yield, and Quality of Leaf Lettuce and Rocket Grown in a Floating System. Agronomy 2019, 9, 382. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9070382
Miceli A, Moncada A, Sabatino L, Vetrano F. Effect of Gibberellic Acid on Growth, Yield, and Quality of Leaf Lettuce and Rocket Grown in a Floating System. Agronomy. 2019; 9(7):382. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9070382
Chicago/Turabian StyleMiceli, Alessandro, Alessandra Moncada, Leo Sabatino, and Filippo Vetrano. 2019. "Effect of Gibberellic Acid on Growth, Yield, and Quality of Leaf Lettuce and Rocket Grown in a Floating System" Agronomy 9, no. 7: 382. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9070382