Application of Proximal Optical Sensors to Fine-Tune Nitrogen Fertilization: Opportunities for Woody Ornamentals

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling Site, Planting Material and Experimental Design
2.2. Plant Measurements
2.3. Optical Measurements at Leaf Level
2.4. Optical Measurements at Canopy Level
2.5. Statistical Analysis
3. Results
3.1. Effects of Fertilization Levels on PlantGgrowth, Quality (Height and Biomass) and N Uptake.
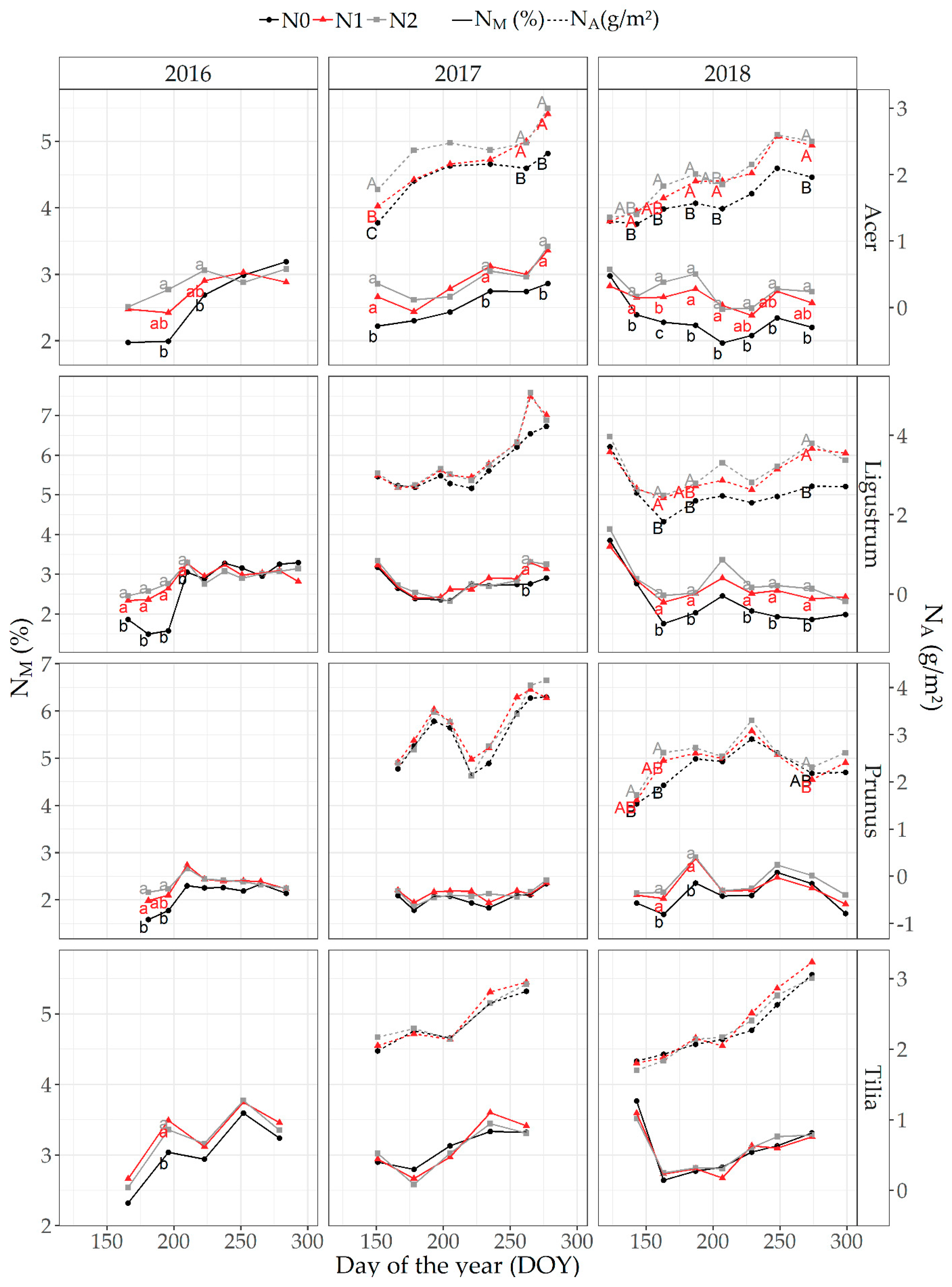
3.2. Seasonal Changes in LMA, Mass- and Area-Based Leaf N Content and Optical Leaf Measurements
3.2.1. A. pseudoplatanus
3.2.2. L. ovalifolium
3.2.3. P. laurocerasus
3.2.4. T. cordata
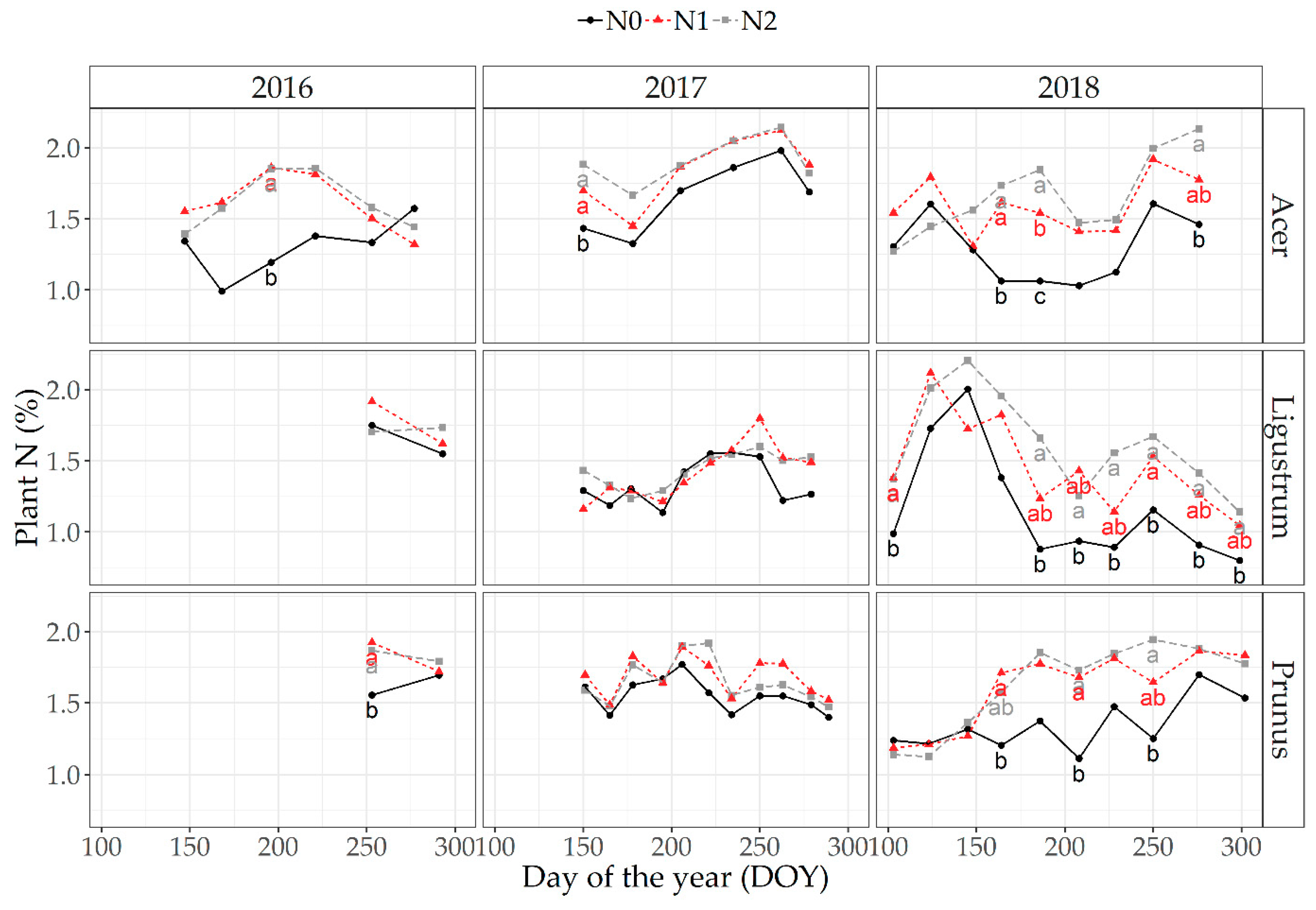
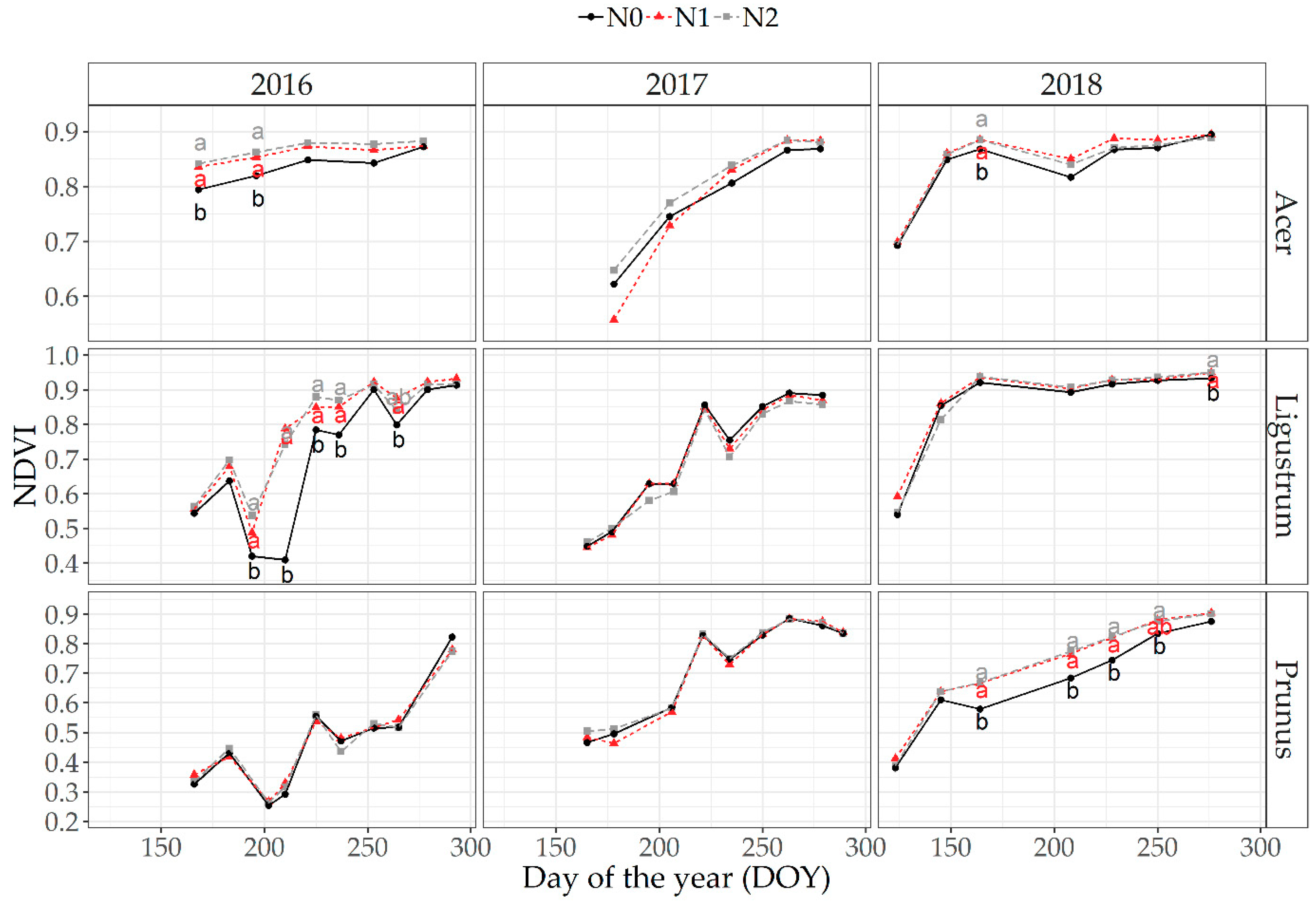
3.3. Seasonal Changes in Plant Biomass, Plant N Content and Optical Plant Measurements (NDVI)
3.4. Absolute/Correlation Approach
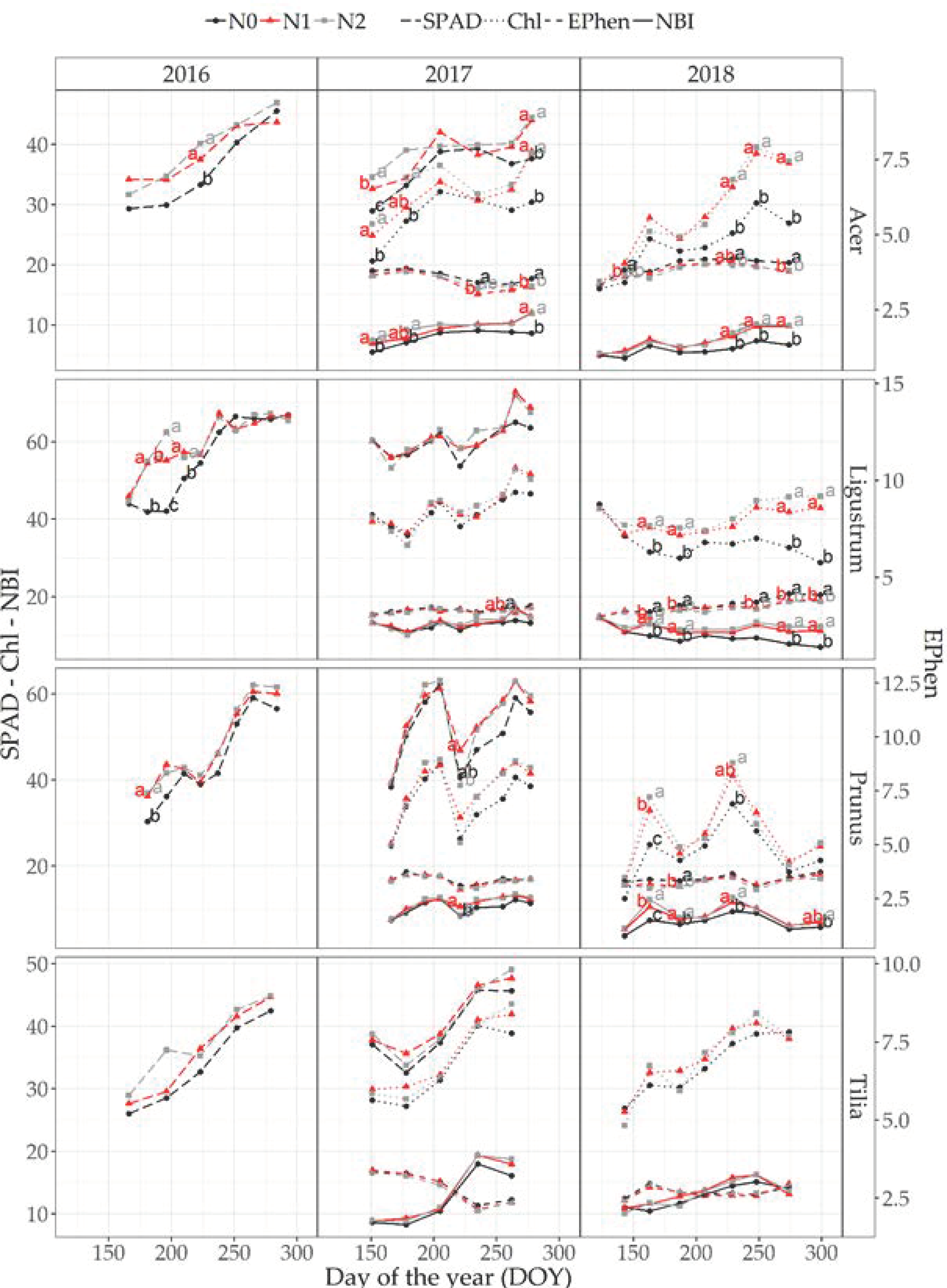
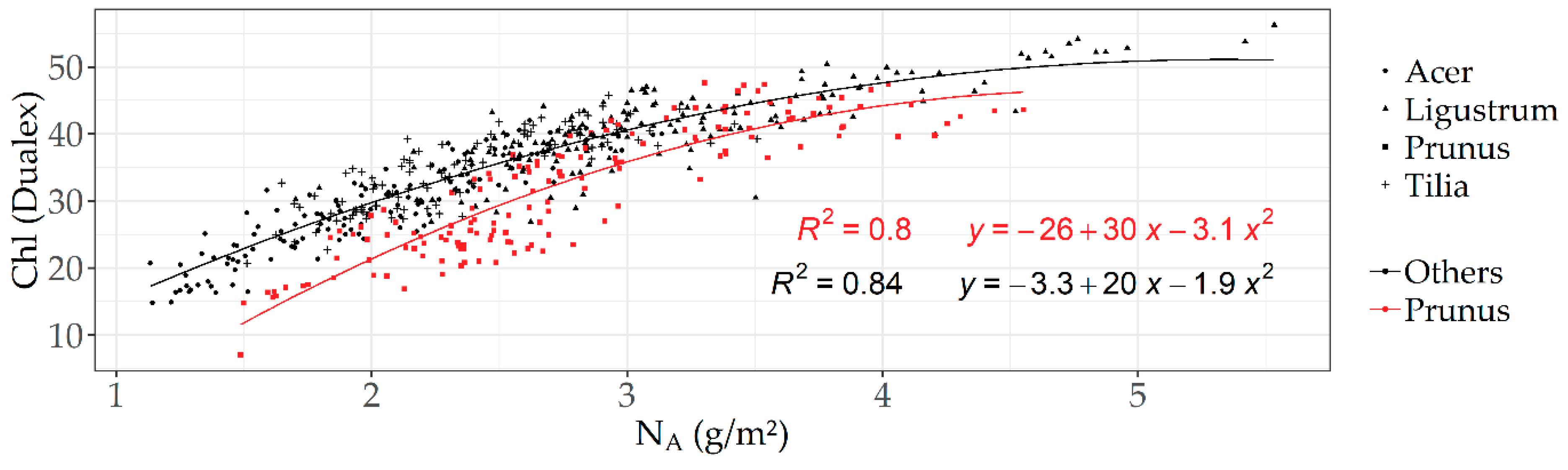
3.4.1. Optical Measurements at Leaf Level
3.4.2. Optical Measurements at Canopy Level
3.5. Relative/Saturation Index Approach
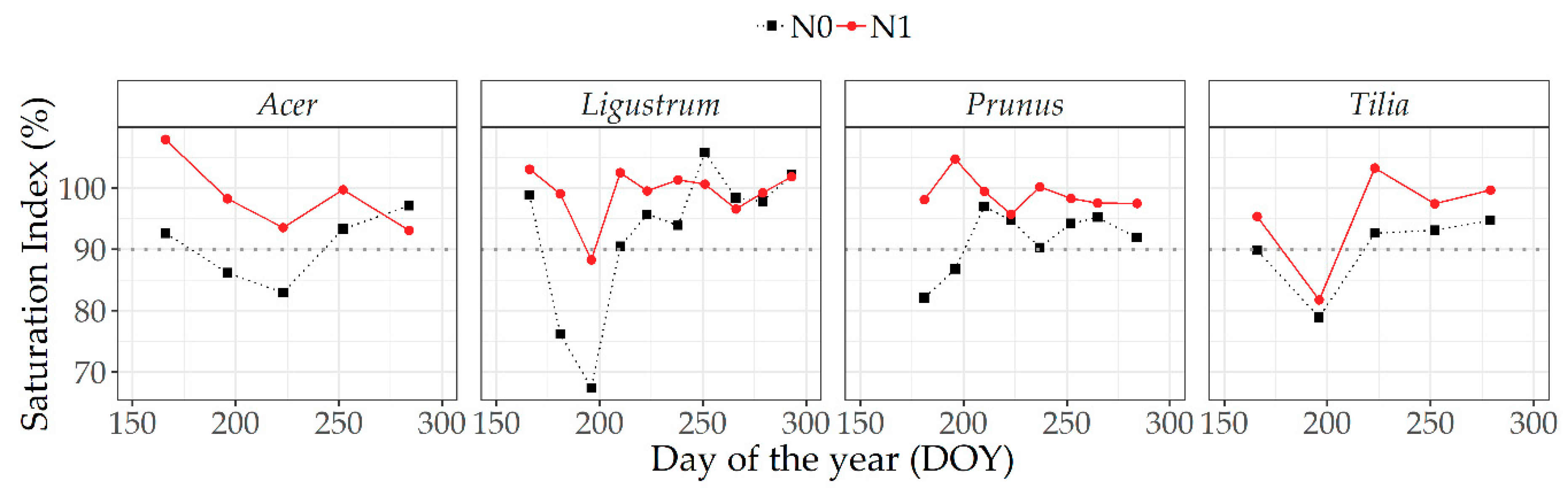
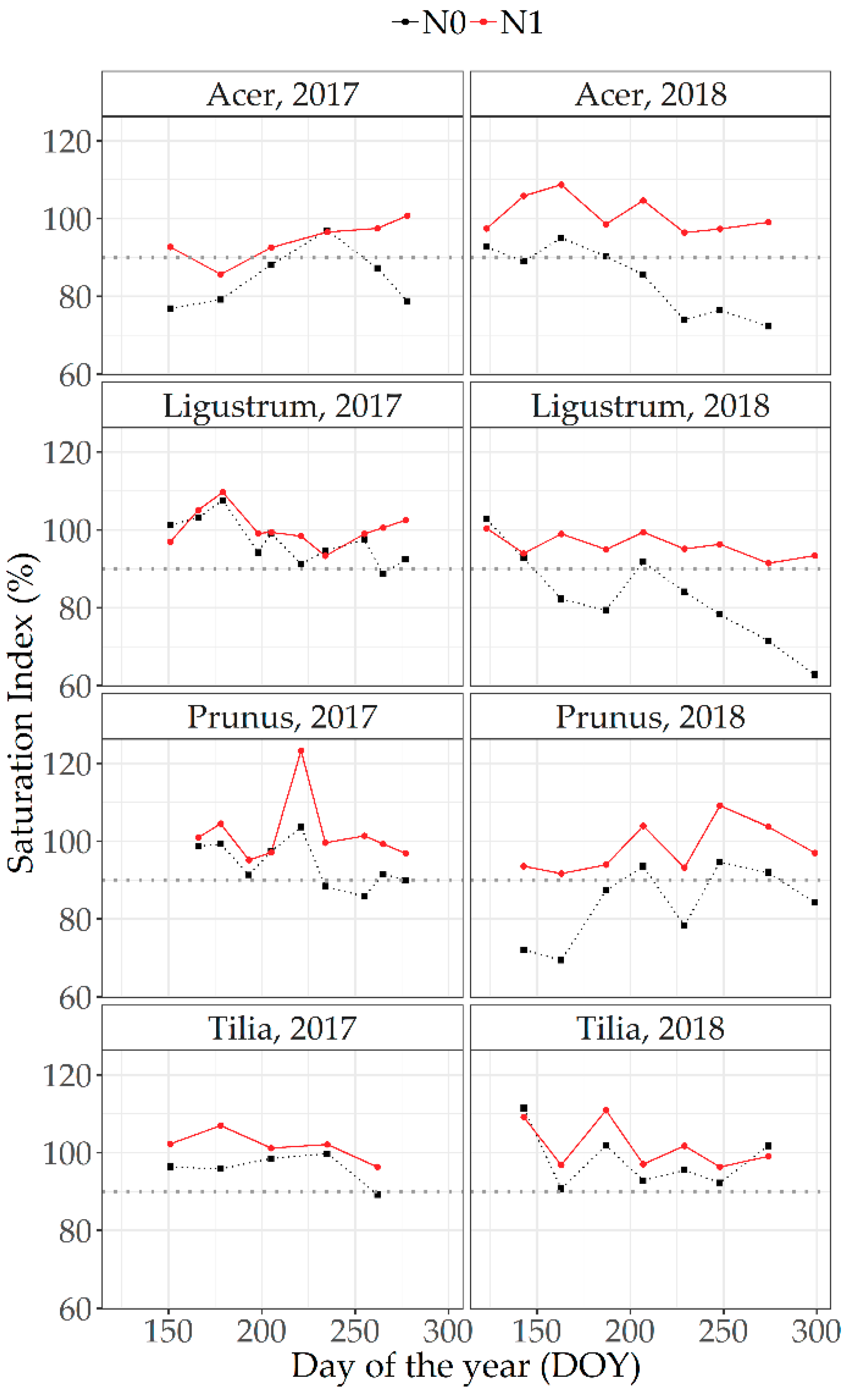
3.5.1. Optical Measurements at Leaf Level
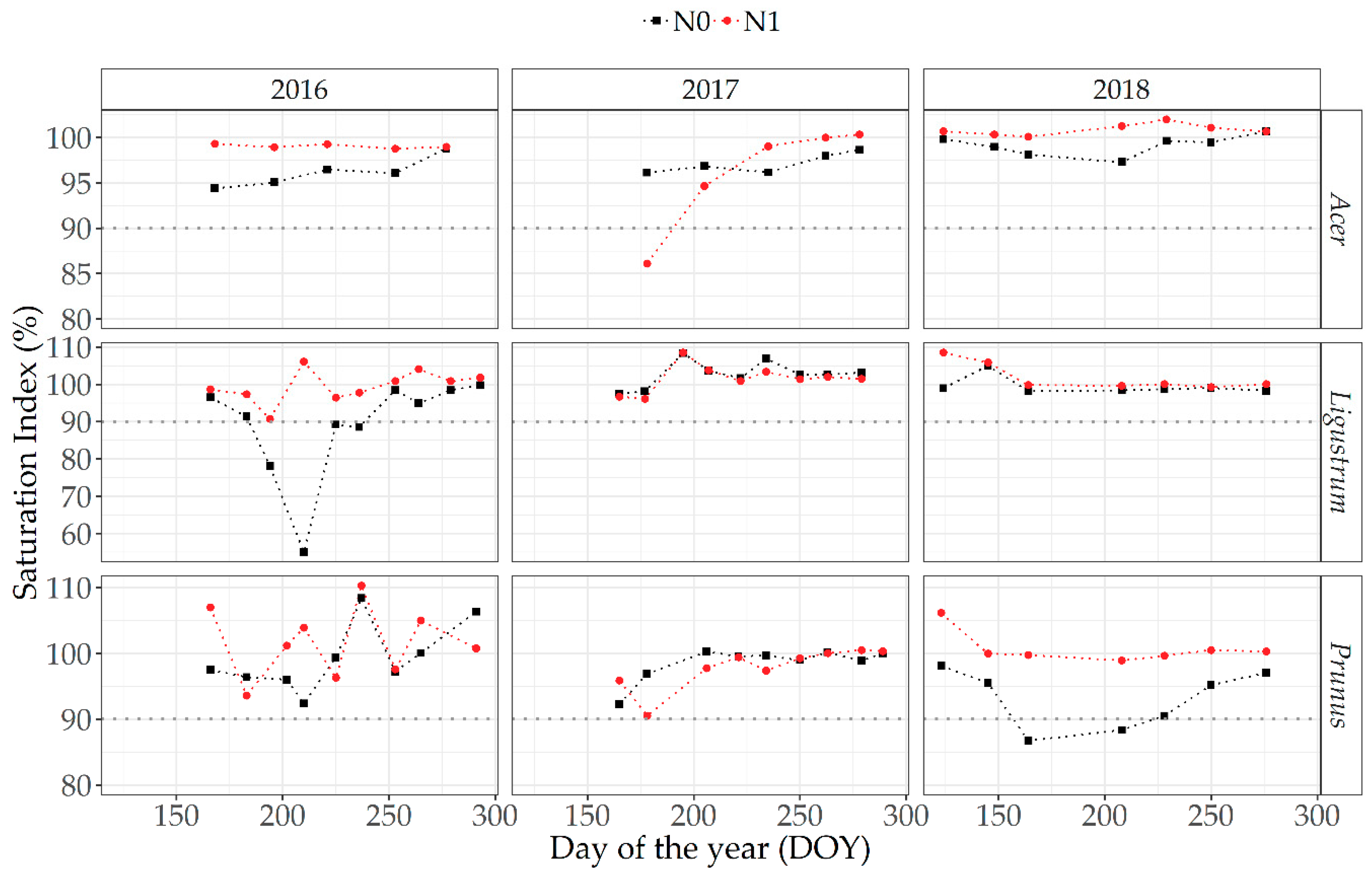
3.5.2. Optical Measurements at Canopy Level
4. Discussion
4.1. Effects of Fertilization Levels
4.2. Absolute/Correlation Approach
4.3. Relative/Saturation Index Approach
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Council of the European Communities Council directive 91/676/EEC concerning the protection of waters against pollution caused by nitrates from agricultural sources. Off. J. Eur. Commun. 1991, 34, 1–8.
- Vlaamse Landmaatschappij Mestrapport 2018. Available online: https://www.vlm.be/nl/SiteCollectionDocuments/Publicaties/mestbank/Mestrapport_2018.pdf (accessed on 20 April 2019).
- Reavis, R.; Whitcomb, C.E. Effects of pruning and fertilization on establishment of bareroot deciduous trees. J. Arboric. 1981, 7, 1978–1980. [Google Scholar]
- Ponder, H.G.; Gilliam, C.H.; Wilkinson, E. Influence of Trickle Irrigation and Nitrogen Rates to Acer rubrum L. J. Environ. Hortic. 1984, 2, 40–43. [Google Scholar]
- Hensley, D.L.; McNiel, R.E.; Sundheim, R. Management influences growth of transplanted Magnolia grandiflora. J. Arboric. 1988, 14, 204–207. [Google Scholar]
- Gilman, E.F.; Marshall, M.D. Fertilizer rate and number of applications impact growth of trees in field soil. Arboric. Urban For. 2014, 40, 178–185. [Google Scholar]
- Gilman, E.F.; Yeager, T.H. Fertilizer type and nitrogen rate affects field-grown Laurel oak and Japanese ligustrum. Proc. Fla. State Hortic. Soc. 1990, 103, 370–372. [Google Scholar]
- Rose, M.A. Nutrient use patterns in woody perennials: Implications for increasing fertilizer efficiency in field-grown and landscape ornamentals. HortTechnology 1999, 9, 613–617. [Google Scholar] [CrossRef]
- Alt, D. N-Fertilization of Nursery Crops in the Field—A Review, Part 1. Gartenbauwwissenschaft 1998, 63, 165–170. [Google Scholar]
- Gilliam, C.H.; Wright, R.D. Timing of fertilizer application in relation to growth flushes of “Helleri” holly (Ilex crenata Thunb.). HortScience 1978, 13, 300–301. [Google Scholar]
- Kuehny, J.S.; Miller, W.B.; DeCoteau, D.R. Changes in Carbohydrate and Nitrogen Relationships during Episodic Growth of Ligustrum japonicum Thunb. J. Am. Soc. Hortic. Sci. 1997, 122, 634–641. [Google Scholar] [CrossRef]
- Hershey, D.R.; Paul, J.L. Ion absorption by a woody plant with episodic growth. HortScience 1983, 18, 357–359. [Google Scholar]
- Evans, C.A.; Miller, E.K.; Friedland, A.J. Effect of nitrogen and light on nutrient concentrations and associated physiological responses in birch and fir seedlings. Plant Soil 2001, 236, 197–207. [Google Scholar] [CrossRef]
- Peoples, M.B.; Dalling, M.J. The Interplay between Proteolysis and Amino Acid Metabolism during Senescence and Nitrogen Reallocation. In Senescence and Aging in Plants; Elsevier BV: Amsterdam, The Netherlands, 1988; pp. 181–217. [Google Scholar]
- Driessche, R.V.D. Prediction of mineral nutrient status of trees by foliar analysis. Bot. Rev. 1974, 40, 347–394. [Google Scholar] [CrossRef]
- Louis, J.; Meyer, S.; Maunoury-Danger, F.; Fresneau, C.; Meudec, E.; Cerovic, Z.G. Seasonal changes in optically assessed epidermal phenolic compounds and chlorophyll contents in leaves of sessile oak (Quercus petraea): Towards signatures of phenological stage. Funct. Plant Boil. 2009, 36, 732–741. [Google Scholar] [CrossRef]
- Neilsen, D.; Hogue, E.J.; Neilsen, G.H.; Parchomchuk, P. Using SPAD-502 Values to Assess the Nitrogen Status of Apple Trees. HortScience 1995, 30, 508–512. [Google Scholar] [CrossRef] [Green Version]
- Turner, F.; Jund, M. Assessing the nitrogen requirements of rice crops with a chlorophyll meter. Aust. J. Exp. Agric. 1994, 34, 1001–1005. [Google Scholar] [CrossRef]
- Xiong, D.; Chen, J.; Yu, T.; Gao, W.; Ling, X.; Li, Y.; Peng, S.; Huang, J. SPAD-based leaf nitrogen estimation is impacted by environmental factors and crop leaf characteristics. Sci. Rep. 2015, 5, 13389. [Google Scholar] [CrossRef] [Green Version]
- Peng, S.; Garcia, F.V.; Laza, R.C.; Cassman, K.G. Adjustment for specific leaf weight improves chlorophyll meter’s estimate of rice leaf nitrogen concentration. Agron. J. 1993, 85, 987–990. [Google Scholar] [CrossRef]
- Debaeke, P.; Rouet, P.; Justes, E. Relationship between the Normalized SPAD Index and the Nitrogen Nutrition Index: Application to Durum Wheat. J. Plant Nutr. 2006, 29, 75–92. [Google Scholar] [CrossRef]
- Schröder, J.; Neeteson, J.; Oenema, O.; Struik, P. Does the crop or the soil indicate how to save nitrogen in maize production? Field Crop Res. 2000, 66, 151–164. [Google Scholar] [CrossRef]
- Blackmer, T.M.; Schepers, J.S.; E Varvel, G.; Meyer, G.E. Analysis of Aerial Photography for Nitrogen Stress within Corn Fields. Agron. J. 1996, 88, 729–733. [Google Scholar] [CrossRef] [Green Version]
- Westerveld, S.M.; McKeown, A.W.; McDonald, M.R.; Scott-Dupree, C.D. Assessment of Chlorophyll and Nitrate Meters as Field Tissue Nitrogen Tests for Cabbage, Onions, and Carrots. HortTechnology 2004, 14, 179–188. [Google Scholar] [CrossRef]
- Varvel, G.E.; Schepers, J.S.; Francis, D.D. Ability for In-Season Correction of Nitrogen Deficiency in Corn Using Chlorophyll Meters. Soil Sci. Soc. Am. J. 1997, 61, 1233–1239. [Google Scholar] [CrossRef]
- Goulas, Y.; Cerovic, Z.G.; Cartelat, A.; Moya, I. Dualex: A new instrument for field measurements of epidermal ultraviolet absorbance by chlorophyll fluorescence. Appl. Opt. 2004, 43, 4488–4496. [Google Scholar] [CrossRef] [PubMed]
- Cerovic, Z.G.; Milhade, C.; Obert, M.; Debuisson, S.; Le Moigne, M.; Ben Ghozlen, N. Nondestructive Diagnostic Test for Nitrogen Nutrition of Grapevine (Vitis vinifera L.) Based on Dualex Leaf-Clip Measurements in the Field. J. Agric. Food Chem. 2015, 63, 3669–3680. [Google Scholar] [CrossRef] [PubMed]
- Deering, D.W.; Haas, R.H.; Welch, R.I.; Harlan, J.C.; Whitney, P.R. Applied Regional Monitoring of the Vernal Advancement and Retrogradation (Green Wave Effect) of Natural Vegetation in the Great Plains Corridor; NASA/GSFC Type III Final Report; Texas A&M University: College Station, TX, USA, 1977. [Google Scholar]
- Danson, F.M.; Plummer, S.E. Red-edge response to forest leaf area index. Int. J. Remote. Sens. 1995, 16, 183–188. [Google Scholar] [CrossRef]
- Percival, G.C.; Keary, I.P.; Noviss, K. The potential of a chlorophyll content SPAD meter to quantify nutrient stress in foliar tissue of sycamore (Acer pseudoplatanus), English oak (Quercus robur), and European beech (Fagus sylvatica). Arboric. Urban For. 2008, 34, 89–100. [Google Scholar]
- Meyer, S.; Cerovic, Z.G.; Goulas, Y.; Montpied, P.; Demotes-Mainard, S.; Bidel, L.P.; Moya, I.; Dreyer, E. Relationships between optically assessed polyphenols and chlorophyll contents, and leaf mass per area ratio in woody plants: A signature of the carbon-nitrogen balance within leaves? Plant Cell Environ. 2006, 29, 1338–1348. [Google Scholar] [CrossRef] [PubMed]
- Demotes-Mainard, S.; Boumaza, R.; Meyer, S.; Cerovic, Z.G. Indicators of nitrogen status for ornamental woody plants based on optical measurements of leaf epidermal polyphenol and chlorophyll contents. Sci. Hortic. 2008, 115, 377–385. [Google Scholar] [CrossRef]
- Official Methods of Analysis Method 968.06-1969. In Official Methods of Analysis of AOAC International; Horwitz, W. (Ed.) Association of Official Analytical Chemists: Arlington, VA, USA, 2000. [Google Scholar]
- Kipp, S.; Mistele, B.; Schmidhalter, U. The performance of active spectral reflectance sensors as influenced by measuring distance, device temperature and light intensity. Comput. Electron. Agric. 2014, 100, 24–33. [Google Scholar] [CrossRef]
- Beneduzzi, H.M.; Souza, E.G.; Bazzi, C.L.; Schenatto, K. Temporal variability in active reflectance sensor-measured ndvi in soybean and wheat crops. Engenharia Agrícola 2017, 37, 771–781. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- Poorter, H.; Niinemets, Ü.; Poorter, L.; Wright, I.J.; Villar, R. Causes and consequences of variation in leaf mass per area (LMA): A meta-analysis. New Phytol. 2009, 182, 565–588. [Google Scholar] [CrossRef] [PubMed]
- Koch, K.; Bhushan, B.; Barthlott, W. Diversity of structure, morphology and wetting of plant surfaces. Soft Matter 2008, 4, 1943–1963. [Google Scholar] [CrossRef]
- Jetter, R.; Schaffer, S.; Riederer, M. Leaf cuticular waxes are arranged in chemically and mechanically distinct layers: Evidence from Prunus laurocerasus L. Plant Cell Environ. 2000, 23, 619–628. [Google Scholar] [CrossRef]
- Villar, R.; Ruiz-Robleto, J.; Ubera, J.L.; Poorter, H. Exploring variation in leaf mass per area (LMA) from leaf to cell: An anatomical analysis of 26 woody species. Am. J. Bot. 2013, 100, 1969–1980. [Google Scholar] [CrossRef] [PubMed]
- Jones, C.G.; Hartley, S.E. A Protein Competition Model of Phenolic Allocation. Oikos 1999, 86, 27–44. [Google Scholar] [CrossRef]
- van Den Burg, J. Minerale voeding van bomen: Bladmonsteranalyse als basis voor een bemestingsadvies. In Boom en Bodem: Bijdragen aan het Symposium Boom en Bodem, Gehouden op 19 en 20 November 1987 te Wageningen en Rhenen; Rijksinstituut voor Onderzoek in de Bos- en Landschapsbouw “De Dorschkamp”: Wageningen, The Netherlands, 1988; pp. 86–110. [Google Scholar]
- Padilla, F.M.; Peña-Fleitas, M.T.; Gallardo, M.; Thompson, R.B. Threshold values of canopy reflectance indices and chlorophyll meter readings for optimal nitrogen nutrition of tomato. Ann. Appl. Biol. 2015, 166, 271–285. [Google Scholar] [CrossRef]
- Padilla, F.M.; Peña-Fleitas, M.; Gallardo, M.; Thompson, R.B. Proximal optical sensing of cucumber crop N status using chlorophyll fluorescence indices. Eur. J. Agron. 2016, 73, 83–97. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 2016 | 2017 | 2018 | |||||||
|---|---|---|---|---|---|---|---|---|---|
| N Treatment | N0 | N1 | N2 | N0 | N1 | N2 | N0 | N1 | N2 |
| Species/Fertilizer | CAN (3) | Tropicote® (4) | Tropicote® (3) | ||||||
| A. pseudoplatanus | 0 | 89 | 179 | 0 | 130 | 260 | 0 | 100 | 150 |
| L. ovalifolium | 0 | 89 | 179 | 0 | 33 | 67 | 0 | 75 | 113 |
| P. laurocerasus | 0 | 89 | 179 | 0 | 33 | 67 | 0 | 75 | 113 |
| T. cordata | 0 | 89 | 179 | 0 | 33 | 67 | 0 | 33 | 50 |
| Year | A. pseudoplatanus | L. ovalifolium | P. laurocerasus | T. cordata |
|---|---|---|---|---|
| 2016 | 1 | 1 | 1 | 1 (2) 1 |
| 2017 | 3 | 2 | 1 (2) 1 | 1 |
| 2018 | 5 (10) 1 | 2 (5) 1 | 2 (5) 1 | 1 (2) 1 |
| Species | Year | DOY | N Treatment | Applied N (kg ha−1) | Height (cm) | Aboveground Biomass (g plant−1) | Aboveground N Uptake (kg ha−1) |
|---|---|---|---|---|---|---|---|
| A. pseudoplatanus | 2016 | 277 | N0 | 0 | 72.0 ± 3.8b | 26.5 ± 4.4a | 131.2 ± 35.9a |
| N1 | 89 | 98.5 ± 6.9a | 48.9 ± 7.7a | 208.9 ± 37.5a | |||
| N2 | 179 | 88.9 ± 3.2ab | 42.6 ± 5.9a | 198.5 ± 35.1a | |||
| 2017 | 278 | N0 | 0 | 46.3 ± 2.4a | 17.2 ± 1.7a | 75.4 ± 10.0a | |
| N1 | 130 | 52.2 ± 6.4a | 20.4 ± 3.6a | 102 ± 21.7a | |||
| N2 | 260 | 58.2 ± 9.7a | 22.9 ± 2.9a | 111.2 ± 16.2a | |||
| 2018 | 276 | N0 | 0 | 60.5 ± 2.5a | 15.2 ± 2.1a | 44.0 ± 9.9b | |
| N1 | 100 | 72.7 ± 6.7a | 23.0 ± 2.0a | 96.6 ± 9.5ab | |||
| N2 | 150 | 73.7 ± 9.8a | 25.5 ± 4.9a | 134.4 ± 27.0a | |||
| L. ovalifolium | 2016 | 293 | N0 | 0 | 45.6 ± 4.4a | 32.5 ± 4.0a | 17.8 ± 5.8a |
| N1 | 89 | 56.2 ± 2.9a | 65.2 ± 10.1a | 97.2 ± 21.6a | |||
| N2 | 179 | 53.4 ± 7.2a | 58.7 ± 20.6a | 90.5 ± 47.9a | |||
| 2017 | 279 | N0 | 0 | 24.9 ± 0.7a | 48.2 ± 11.9a | 85.1 ± 18.0a | |
| N1 | 33 | 19.9 ± 2.4a | 52.0 ± 7.6a | 123.5 ± 18.7a | |||
| N2 | 67 | 22.4 ± 2.0a | 55.2 ± 13.9a | 136.5 ± 37.0a | |||
| 2018 | 276 | N0 | 0 | 65.3 ± 3.9a | 121.0 ± 16a | 164.2 ± 31.0a | |
| N1 | 75 | 71.7 ± 4.9a | 146.3 ± 11.2a | 271.6 ± 10.6a | |||
| N2 | 113 | 77.6 ± 5.6a | 124.1 ± 4.6a | 252.4 ± 6.8a | |||
| P. laurocerasus | 2016 | 291 | N0 | 0 | 41.1 ± 3.5a | 48.1 ± 7.5a | 26.6 ± 6.9a |
| N1 | 89 | 49.0 ± 3.8a | 50.4 ± 1.9a | 28.9 ± 4.4a | |||
| N2 | 179 | 43.9 ± 2.9a | 58.6 ± 2.6a | 37.4 ± 3.0a | |||
| 2017 | 289 | N0 | 0 | 38.9 ± 1.7a | 114.8 ± 9.6a | 109.4 ± 8.1b | |
| N1 | 33 | 37.9 ± 1.3a | 136.0 ± 13.3a | 146.3 ± 16.1ab | |||
| N2 | 67 | 42.6 ± 5.4a | 183.7 ± 26.1a | 193.4 ± 21.7a | |||
| 2018 | 302 | N0 | 0 | 29.0 ± 2.9a | 106.1 ± 1.9a | 113.6 ± 20.3a | |
| N1 | 75 | 33.8 ± 1.1a | 121.4 ± 19.4a | 137.6 ± 32.1a | |||
| N2 | 113 | 32.8 ± 2.1a | 134.8 ± 3.6a | 175.4 ± 17.2a | |||
| T. cordata | 2016 | 281 | N0 | 0 | 81.6 ± 5.6ab | 75.0 ± 5.8a | 19.7 ± 1.7a |
| N1 | 89 | 91.6 ± 7.9a | 101.7 ± 17.8a | 27.4 ± 4.2a | |||
| N2 | 179 | 60.0 ± 5.0b | 69.4 ± 19.8a | 18.7 ± 5.3a | |||
| 2017 | 277 | N0 | 0 | 179.2 ± 15.7a | 171.2 ± 43.9a | 13.0 ± 9.6a | |
| N1 | 33 | 214.3 ± 16.3a | 339.0 ± 83.3a | 46.9 ± 15.6a | |||
| N2 | 67 | 172.2 ± 8.2a | 175.9 ± 14.3a | 14.1 ± 1.2a | |||
| 2018 | 276 | N0 | 0 | 266.9 ± 15.5b | 437.4 ± 2.1a | 53.4 ± 1.9a | |
| N1 | 33 | 328.8 ± 4.8a | 497.6 ± 76.7a | 58.6 ± 13.9a | |||
| N2 | 50 | 299.7 ± 17.3ab | 422.1 ± 63.1a | 51.1 ± 8.5a |
| A. pseudoplatanus | L. ovalifolium | P. laurocerasus | T. cordata | Overall | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2017 | 2018 | 2 years 1 | 2017 | 2018 | 2 years 1 | 2017 | 2018 | 2 years 1 | 2017 | 2018 | 2 years 1 | 2017 | 2018 | 2 years 1 | |||||
| SPAD vs. NM | 0.66 *** | 0.76 *** | 0.49 *** | 0.66 *** | 0.30 ** | 0.12 ns | 0.80 *** | 0.66 *** | -0.04 ns | 0.10 * | |||||||||
| SPAD/LMA vs. NM | 0.27 ns | 0.39 *** | 0.25 * | 0.72 *** | 0.75 *** | ||||||||||||||
| SPAD vs. NA | 0.80 *** | 0.87 *** | 0.84 *** | 0.83 *** | 0.86 *** | 0.86 *** | |||||||||||||
| Chl vs. NM | 0.60 *** | −0.04 ns | 0.28 ** | 0.45 *** | 0.49 *** | 0.48 *** | 0.32 ** | 0.07 ns | −0.02 ns | 0.81 *** | −0.04 ns | 0.31 *** | 0.22 *** | 0.30 *** | 0.26 *** | ||||
| Chlad vs. NM | 0.59 *** | −0.03 ns | 0.28 ** | 0.44 *** | 0.51 *** | 0.48 *** | 0.32 ** | 0.07 ns | −0.03 ns | 0.81 *** | −0.05 ns | 0.31 ** | 0.21 *** | 0.30 *** | 0.25 *** | ||||
| Chlab vs. NM | 0.61 *** | −0.04 ns | 0.28 ** | 0.45 *** | 0.47 *** | 0.47 *** | 0.31 ** | 0.07 ns | −0.02 ns | 0.80 *** | −0.04 ns | 0.32 *** | 0.24 *** | 0.30 *** | 0.26 *** | ||||
| Chl/LMA vs. NM | 0.65 *** | 0.56 *** | 0.63 *** | 0.31 ** | 0.80 *** | 0.73 *** | 0.32 ** | 0.50 *** | 0.40 *** | 0.89 *** | 0.49 *** | 0.64 *** | 0.85 *** | 0.75 *** | 0.79 *** | ||||
| Chl vs. NA | 0.89 *** | 0.93 *** | 0.91 *** | 0.84 *** | 0.77 *** | 0.83 *** | 0.83 *** | 0.85 *** | 0.87 *** | 0.90 *** | 0.80 *** | 0.82 *** | 0.86 *** | 0.79 *** | 0.84 *** | ||||
| EPhen vs. NM | −0.72 *** | −0.66 *** | −0.72 *** | −0.35 *** | −0.60 *** | −0.56 *** | −0.09 ns | −0.53 *** | −0.36 *** | −0.83 *** | −0.33 ** | −0.55 *** | −0.49 *** | −0.53 *** | −0.51 *** | ||||
| EPhenad vs. NM | −0.77 *** | −0.58 *** | −0.71 *** | −0.14 ns | −0.73 *** | −0.55 *** | −0.20 ns | −0.45 *** | −0.32 *** | −0.80 *** | 0.09 ns | −0.37 *** | −0.02 ns | −0.31 *** | −0.19 *** | ||||
| EPhenab vs. NM | −0.64 *** | −0.64 *** | −0.68 *** | −0.34 ** | −0.51 *** | −0.49 *** | −0.03 ns | −0.53 *** | −0.36 *** | −0.80 *** | −0.64 *** | −0.70 *** | 0.74 *** | −0.64 *** | −0.68 *** | ||||
| EPhen/LMA vs. NM | −0.54 *** | 0.28 * | −0.17 ns | −0.35 ** | 0.16 ns | 0.01 ns | −0.17 ns | 0.17 ns | 0.21 ** | −0.66 *** | 0.30 * | −0.11 ns | 0.28 *** | 0.13 * | 0.18 *** | ||||
| EPhen vs. NA | −0.38 ** | 0.38 ** | −0.17 ns | 0.19 ns | −0.02 ns | −0.04 ns | −0.04 ns | 0.23 ns | 0.32 *** | −0.81 *** | 0.35 ** | −0.25 ** | 0.00 ns | −0.08 ns | −0.07 ns | ||||
| NBI vs. NM | 0.80 *** | 0.12 ns | 0.50 *** | 0.58 *** | 0.72 *** | 0.66 *** | 0.35 ** | 0.19 ns | 0.05 ns | 0.84 *** | 0.05 ns | 0.47 *** | 0.46 *** | 0.51 *** | 0.47 *** | ||||
| NBI vs. NA | 0.89 *** | 0.90 *** | 0.90 *** | 0.79 *** | 0.61 *** | 0.72 *** | 0.72 *** | 0.79 *** | 0.83 *** | 0.86 *** | 0.68 *** | 0.68*** | 0.64 *** | 0.64 *** | 0.67 *** | ||||
| Chl vs. SPAD | 0.90 *** | 0.88 *** | 0.99 *** | 0.94 *** | 0.89 *** | ||||||||||||||
| NDVI vs. | A. pseudoplatanus | L. ovalifolium | P. laurocerasus | Overall | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2016 | 2017 | 2018 | 3 years | 2017 | 2018 | 2 years | 2017 | 2018 | 2 years | 2017 | 2018 | 3 years | |
| Biomass, kg plant−1 | 0.67 *** | 0.78 *** | 0.55 *** | 0.50 *** | 0.77 *** | 0.71 *** | 0.75 *** | 0.69 *** | 0.82 *** | 0.72 *** | 0.44 *** | 0.41 *** | 0.38 *** |
| Plant N (%) | 0.55 *** | 0.74 *** | 0.08 ns | 0.17 * | 0.47 *** | −0.44 *** | 0.03 ns | −0.19 ns | 0.66 *** | 0.28 *** | 0.31 *** | 0.08 ns | 0.15 ** |
| N uptake, kg ha−1 | 0.72 *** | 0.79 *** | 0.49 *** | 0.53 *** | 0.75 *** | 0.59 *** | 0.69 *** | 0.69 *** | 0.81 *** | 0.74 *** | 0.69 *** | 0.59 *** | 0.61 *** |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bracke, J.; Elsen, A.; Adriaenssens, S.; Schoeters, L.; Vandendriessche, H.; Van Labeke, M.-C. Application of Proximal Optical Sensors to Fine-Tune Nitrogen Fertilization: Opportunities for Woody Ornamentals. Agronomy 2019, 9, 408. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9070408
Bracke J, Elsen A, Adriaenssens S, Schoeters L, Vandendriessche H, Van Labeke M-C. Application of Proximal Optical Sensors to Fine-Tune Nitrogen Fertilization: Opportunities for Woody Ornamentals. Agronomy. 2019; 9(7):408. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9070408
Chicago/Turabian StyleBracke, Jolien, Annemie Elsen, Sandy Adriaenssens, Lore Schoeters, Hilde Vandendriessche, and Marie-Christine Van Labeke. 2019. "Application of Proximal Optical Sensors to Fine-Tune Nitrogen Fertilization: Opportunities for Woody Ornamentals" Agronomy 9, no. 7: 408. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9070408