Solution NMR Structure of the SH3 Domain of Human Caskin1 Validates the Lack of a Typical Peptide Binding Groove and Supports a Role in Lipid Mediator Binding

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Protein Expression and Purification
2.2. NMR Spectroscopy
3. Results
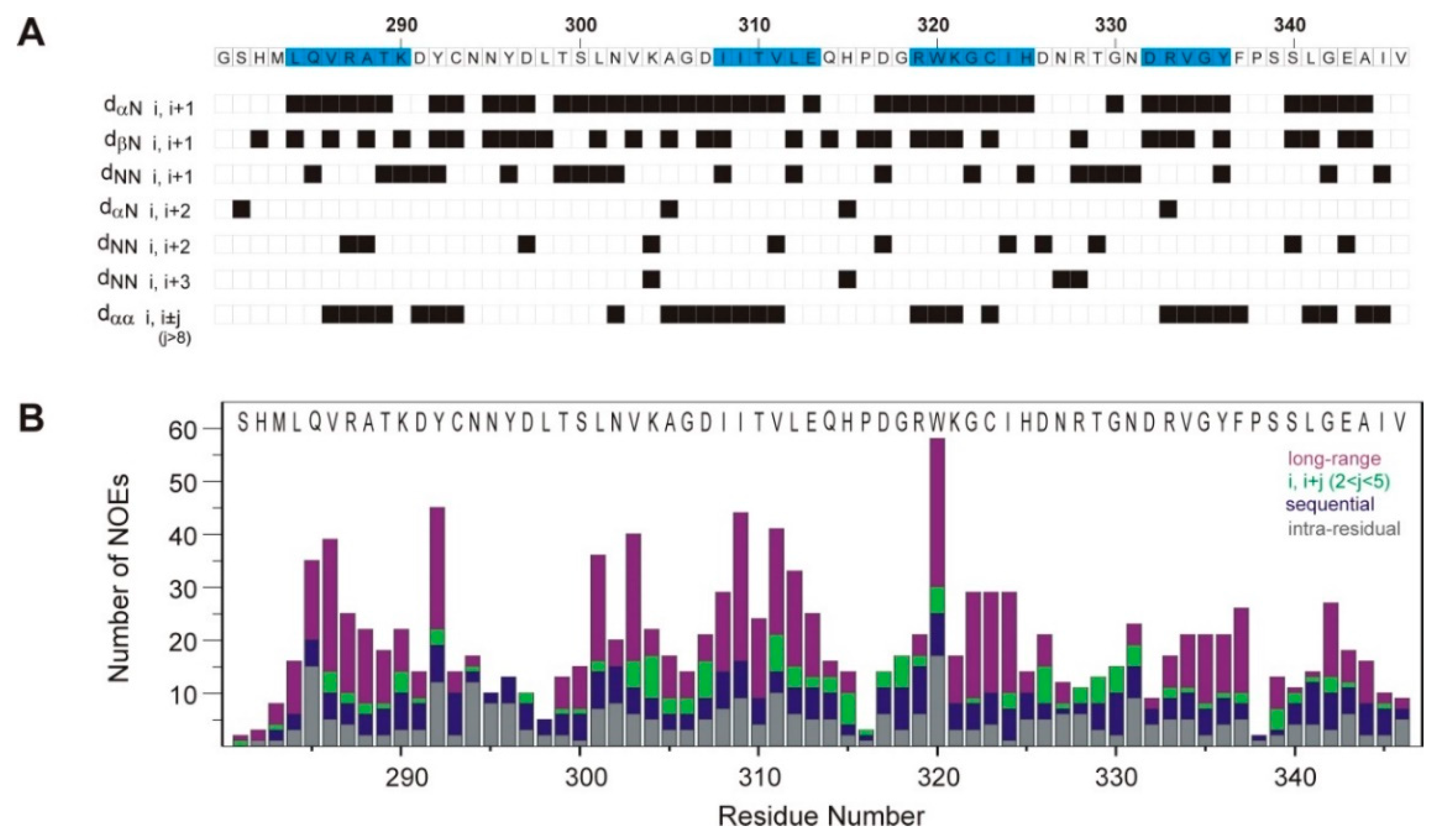
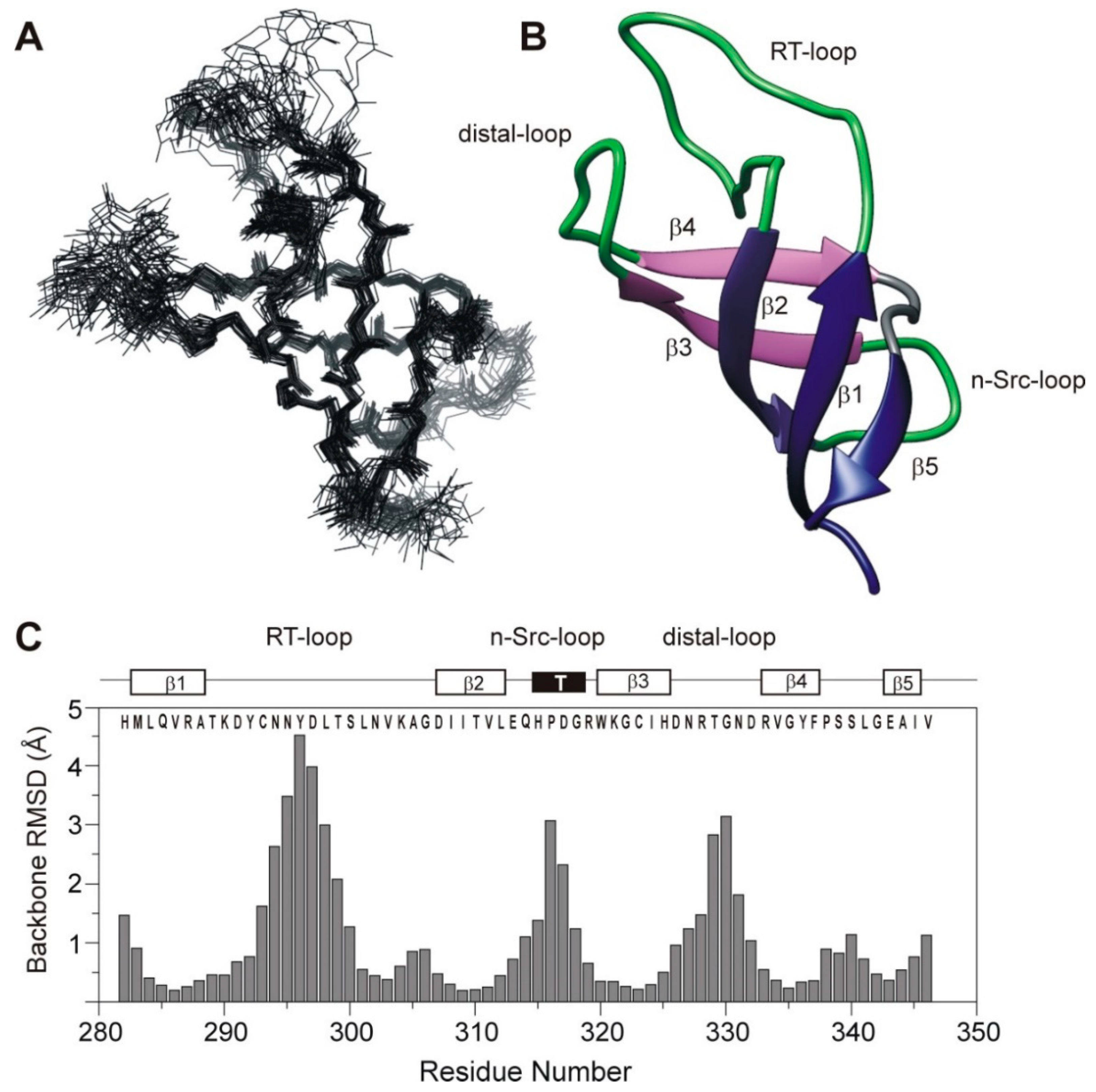
3.1. 3D Solution Structure of the SH3 Domain of Human Caskin1
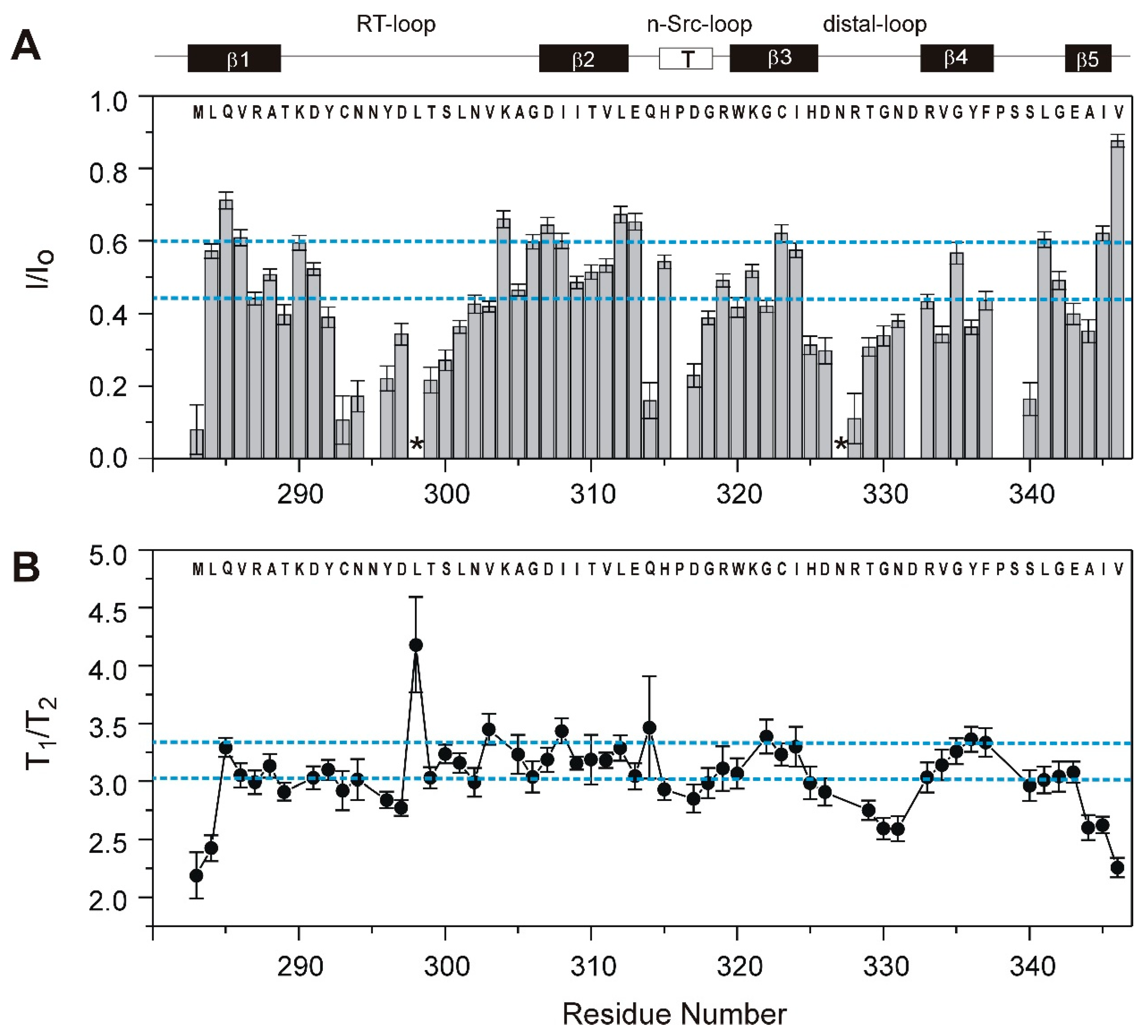
3.2. Solvent Exchange and Backbone 15N Relaxation
3.3. Mapping LPA-Induced Chemical Shift Perturbations on the NMR Structure of Human Caskin1 SH3 Domain
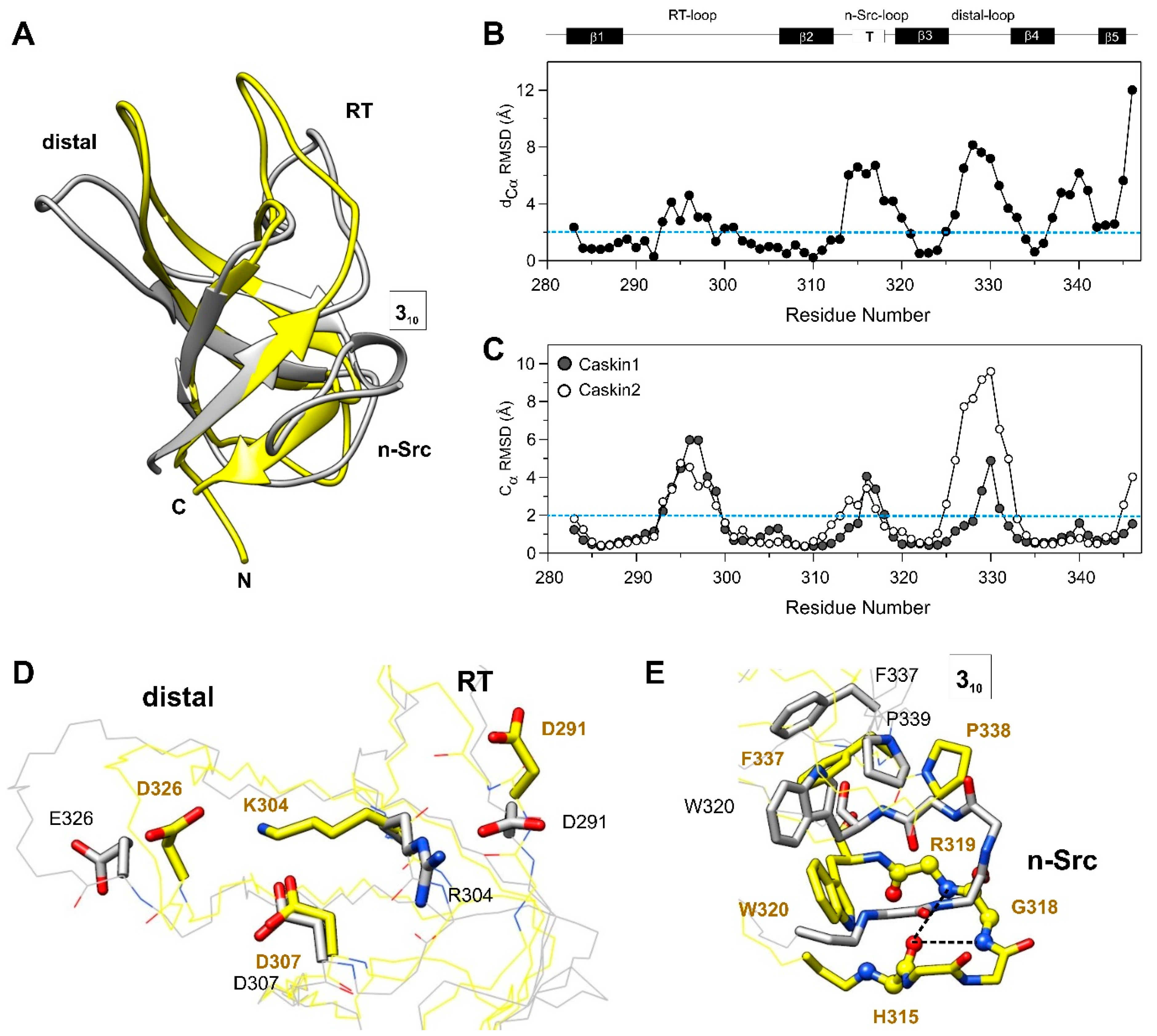
3.4. Comparison with the SH3 Domain of Human Caskin2
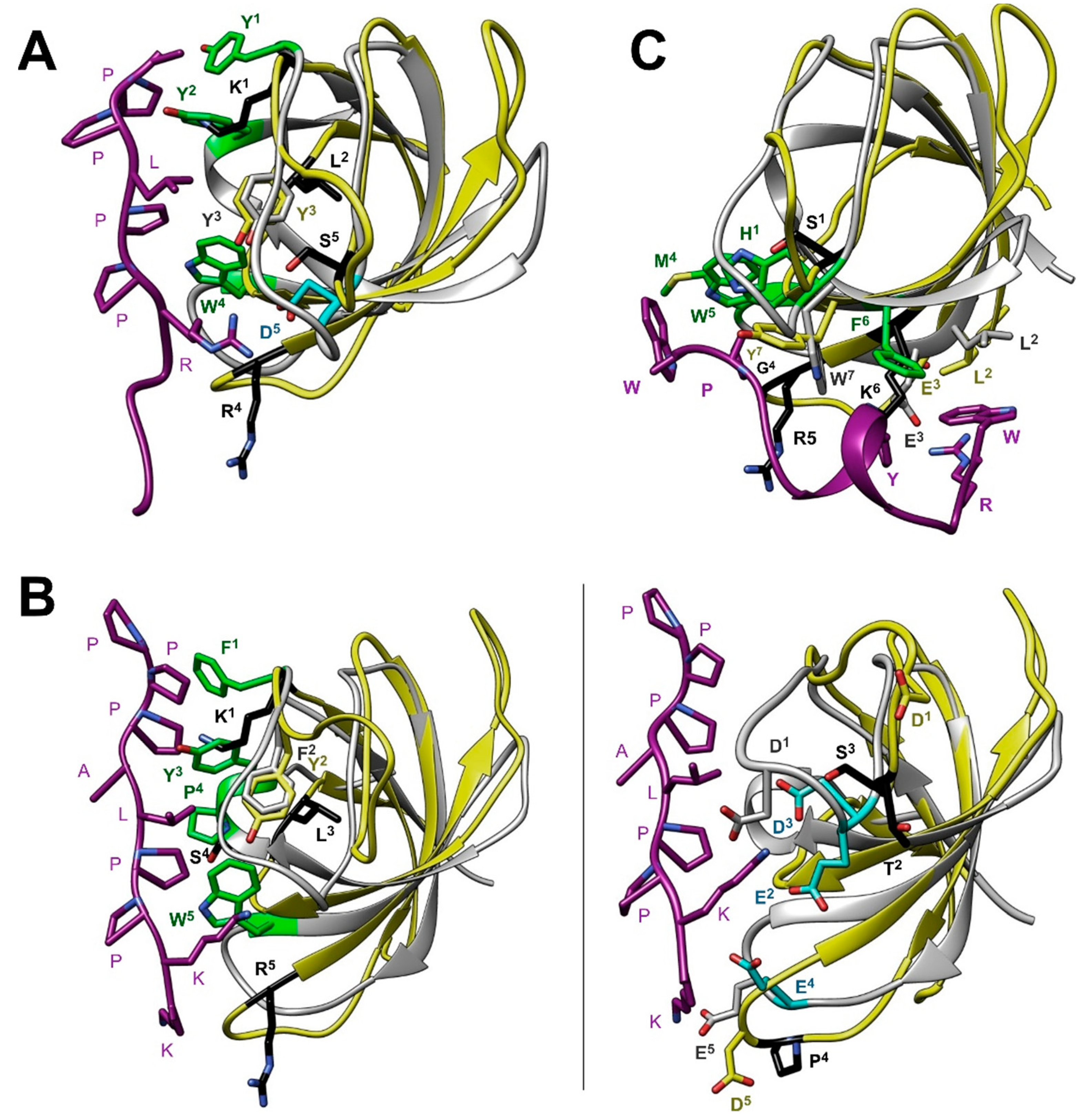
3.5. Comparison with other SH3 Domains
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tabuchi, K.; Biederer, T.; Butz, S.; Sudhof, T.C. CASK participates in alternative tripartite complexes in which Mint 1 competes for binding with caskin 1, a novel CASK-binding protein. J. Neurosci. 2002, 22, 4264–4273. [Google Scholar] [CrossRef] [PubMed]
- Balazs, A.; Csizmok, V.; Buday, L.; Rakacs, M.; Kiss, R.; Bokor, M.; Udupa, R.; Tompa, K.; Tompa, P. High levels of structural disorder in scaffold proteins as exemplified by a novel neuronal protein, CASK-interactive protein1. FEBS J. 2009, 276, 3744–3756. [Google Scholar] [CrossRef] [PubMed]
- Stafford, R.L.; Ear, J.; Knight, M.J.; Bowie, J.U. The molecular basis of the Caskin1 and Mint1 interaction with CASK. J. Mol. Biol. 2011, 412, 3–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weng, Y.L.; Liu, N.; DiAntonio, A.; Broihier, H.T. The cytoplasmic adaptor protein Caskin mediates Lar signal transduction during Drosophila motor axon guidance. J. Neurosci. 2011, 31, 4421–4433. [Google Scholar] [CrossRef] [Green Version]
- Pesti, S.; Balazs, A.; Udupa, R.; Szabo, B.; Fekete, A.; Bogel, G.; Buday, L. Complex formation of EphB1/Nck/Caskin1 leads to tyrosine phosphorylation and structural changes of the Caskin1 SH3 domain. Cell Commun. Signal. 2012, 10, 36. [Google Scholar] [CrossRef] [Green Version]
- Stein, E.; Lane, A.A.; Cerretti, D.P.; Schoecklmann, H.O.; Schroff, A.D.; Van Etten, R.L.; Daniel, T.O. Eph receptors discriminate specific ligand oligomers to determine alternative signaling complexes, attachment, and assembly responses. Genes Dev. 1998, 12, 667–678. [Google Scholar] [CrossRef] [Green Version]
- Daimon, C.M.; Jasien, J.M.; Wood, W.H., 3rd; Zhang, Y.; Becker, K.G.; Silverman, J.L.; Crawley, J.N.; Martin, B.; Maudsley, S. Hippocampal Transcriptomic and Proteomic Alterations in the BTBR Mouse Model of Autism Spectrum Disorder. Front. Physiol. 2015, 6, 324. [Google Scholar] [CrossRef] [Green Version]
- Lv, X.; Zhao, K.; Lan, Y.; Li, Z.; Ding, N.; Su, J.; Lu, H.; Song, D.; Gao, F.; He, W. miR-21a-5p Contributes to Porcine Hemagglutinating Encephalomyelitis Virus Proliferation via Targeting CASK-Interactive Protein1 In vivo and vitro. Front. Microbiol. 2017, 8, 304. [Google Scholar] [CrossRef]
- Datta, A.; Jingru, Q.; Khor, T.H.; Teo, M.T.; Heese, K.; Sze, S.K. Quantitative neuroproteomics of an in vivo rodent model of focal cerebral ischemia/reperfusion injury reveals a temporal regulation of novel pathophysiological molecular markers. J. Proteome Res. 2011, 10, 5199–5213. [Google Scholar] [CrossRef]
- Middleton, F.A.; Carrierfenster, K.; Mooney, S.M.; Youngentob, S.L. Gestational ethanol exposure alters the behavioral response to ethanol odor and the expression of neurotransmission genes in the olfactory bulb of adolescent rats. Brain Res. 2009, 1252, 105–116. [Google Scholar] [CrossRef] [Green Version]
- Crockett, D.K.; Lin, Z.; Elenitoba-Johnson, K.S.; Lim, M.S. Identification of NPM-ALK interacting proteins by tandem mass spectrometry. Oncogene 2004, 23, 2617–2629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katano, T.; Takao, K.; Abe, M.; Yamazaki, M.; Watanabe, M.; Miyakawa, T.; Sakimura, K.; Ito, S. Distribution of Caskin1 protein and phenotypic characterization of its knockout mice using a comprehensive behavioral test battery. Mol. Brain 2018, 11, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bencsik, N.; Pusztai, S.; Borbély, S.; Fekete, A.; Dülk, M.; Kis, V.; Pesti, S.; Vas, V.; Szűcs, A.; Buday, L.; et al. Dendritic spine morphology and memory formation depend on postsynaptic Caskin proteins. Sci. Rep. 2019, 9, 16843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oyazato, Y.; Iijima, K.; Emi, M.; Sekine, T.; Kamei, K.; Takanashi, J.; Nakao, H.; Namai, Y.; Nozu, K.; Matsuo, M. Molecular analysis of TSC2/PKD1 contiguous gene deletion syndrome. Kobe J. Med. Sci. 2011, 57, E1–E10. [Google Scholar]
- Boehm, D.; Bacher, J.; Neumann, H.P. Gross genomic rearrangement involving the TSC2-PKD1 contiguous deletion syndrome: Characterization of the deletion event by quantitative polymerase chain reaction deletion assay. Am. J. Kidney Dis. 2007, 49, e11–e21. [Google Scholar] [CrossRef] [PubMed]
- Ren, R.; Mayer, B.J.; Cicchetti, P.; Baltimore, D. Identification of a ten amino acid proline-rich SH3 binding site. Science 1993, 259, 1157–1161. [Google Scholar] [CrossRef]
- Saksela, K.; Permi, P. SH3 domain ligand binding: What’s the consensus and where’s the specificity? FEBS Lett. 2012, 586, 2609–2614. [Google Scholar] [CrossRef] [Green Version]
- Kwan, J.J.; Donaldson, L.W. A lack of peptide binding and decreased thermostability suggests that the CASKIN2 scaffolding protein SH3 domain may be vestigial. BMC Str. Biol. 2016, 16, 14. [Google Scholar] [CrossRef] [Green Version]
- Mayer, B.J. SH3 domains: Complexity in moderation. J. Cell. Sci. 2001, 114, 1253–1263. [Google Scholar]
- Koprivanacz, K.; Toke, O.; Besztercei, B.; Juhasz, T.; Radnai, L.; Mero, B.; Mihaly, J.; Peter, M.; Balogh, G.; Vigh, L.; et al. The SH3 domain of Caskin1 binds to lysophosphatidic acid suggesting a direct role for the lipid in intracellular signaling. Cell. Signal. 2017, 32, 66–75. [Google Scholar] [CrossRef]
- Moolenaar, W.H.; van Meeteren, L.A.; Giepmans, B.N. The ins and outs of lysophosphatidic acid signaling. Bioessays 2004, 26, 870–881. [Google Scholar] [CrossRef] [PubMed]
- Xiang, H.; Lu, Y.; Shao, M.; Wu, T. Lysophosphatidic Acid Receptors: Biochemical and Clinical Implications in Different Diseases. J. Cancer 2020, 11, 3519–3535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kay, L.E.; Keifer, P.; Saarinen, T. Pure absorption gradient enhanced heteronuclear single quantum correlation spectroscopy with improved sensitivity. J. Am. Chem. Soc. 1992, 114, 10663–10665. [Google Scholar] [CrossRef]
- Wittekind, M.; Mueller, L. HNCACB, a high-sensitivity 3D NMR experiment to correlate amide-proton and nitrogen resonances with the alpha- and beta-carbon resonances in proteins. J. Magn. Reson. Ser. B 1993, 101, 201–205. [Google Scholar] [CrossRef]
- Grzesiek, S.; Bax, A. Correlating backbone amide and side chain resonances in larger proteins by multiple relayed triple resonance NMR. J. Am. Chem. Soc. 1992, 114, 6291–6293. [Google Scholar] [CrossRef]
- Grzesiek, S.; Bax, A. Amino acid type determination in the sequential assignment procedure of uniformly 13C/15 N-enriched proteins. J. Biomol. NMR 1993, 3, 185–204. [Google Scholar] [CrossRef]
- Grzesiek, S.; Anglister, J.; Bax, A. Correlation of backbone amide and aliphatic side-chain resonances in 13C/15N-enriched proteins by isotropic mixing of 13C magnetization. J. Magn. Res. 1993, 101, 114–119. [Google Scholar] [CrossRef] [Green Version]
- Kay, L.E.; Xu, G.Y.; Singer, A.U.; Muhandiram, D.R.; Forman-Kay, J.D. A gradient enhanced HCCH-TOCSY experiment for recording side-chain proton and carbon-13 correlations in water samples of proteins. J. Magn. Res. B 1993, 101, 333–337. [Google Scholar] [CrossRef]
- Markley, J.L.; Bax, A.; Arata, Y.; Hilbers, C.W.; Kaptein, R.; Sykes, B.D.; Wright, P.E.; Wüthrich, K. Recommendations for the presentation of NMR structures of proteins and nucleic acids. J. Mol. Biol. 1998, 280, 933–952. [Google Scholar] [CrossRef]
- Wishart, D.S.; Sykes, B.D. The 13C chemical-shift index: A simple method for the identification of protein secondary structure using 13C chemical-shift data. J. Biomol. NMR 1994, 4, 171–180. [Google Scholar] [CrossRef]
- Muhandiram, D.R.; Farrow, N.A.; Xu, G.Y.; Smallcombe, S.H.; Kay, L.E. A gradient 13C NOESY-HSQC experiment for recording NOESY spectra of 13C-labeled protein dissolved in H2O. J. Magn. Reson. B 1993, 102, 317–321. [Google Scholar] [CrossRef]
- Marion, D.; Kay, L.E.; Sparks, S.W.; Torchia, D.A.; Bax, A. Three-dimensional heteronuclear NMR of nitrogen-15 labeled proteins. J. Am. Chem. Soc. 1989, 111, 1515–1517. [Google Scholar] [CrossRef]
- Zwahlen, C.; Gardner, K.H.; Sarma, S.P.; Horita, D.A.; Byrd, R.A.; Kay, L.E. An NMR experiment for measuring methyl–methyl NOEs in 13C-labeled proteins with high resolution. J. Am. Chem. Soc. 1998, 120, 7617–7625. [Google Scholar] [CrossRef]
- Kumar, A.; Ernst, R.R.; Wüthrich, K. A two-dimensional nuclear Overhauser enhancement (2D NOE) experiment for the elucidation of complete proton-proton cross-relaxation networks in biological macromolecules. Biochem. Biophys. Res. Commun. 1980, 95, 1–6. [Google Scholar] [CrossRef]
- Rieping, W.; Habeck, M.; Bardiaux, B.; Bernard, A.; Malliavin, T.E.; Nilges, M. ARIA2: Automated NOE assignment and data integration in NMR structure calculation. Bioinformatics 2007, 23, 381–382. [Google Scholar] [CrossRef] [Green Version]
- Mareuil, F.; Malliavin, T.E.; Nilges, M.; Bardiaux, B. Improved reliability, accuracy and quality in automated structure calculation with ARIA. J. Biomol. NMR 2015, 62, 425–438. [Google Scholar] [CrossRef] [Green Version]
- Laskowski, R.A.; MacArthur, M.W.; Moss, D.S.; Thornton, J.M. PROCHECK: A program to check the stereochemical quality of protein structures. J. Appl. Chrystallogr. 1993, 26, 283–291. [Google Scholar] [CrossRef]
- Davis, I.W.; Leaver-Fay, A.; Chen, V.B.; Block, J.N.; Kapral, G.J.; Wang, X.; Murray, L.W.; Arendall, W.B., 3rd; Snoeyink, J.; Richardson, J.S.; et al. MolProbity: All-atom contacts and structure validation for proteins and nucleic acids. Nucleic Acids Res. 2007, 35, W375–W383. [Google Scholar] [CrossRef] [Green Version]
- Kay, L.; Nicholson, L.; Delaglio, F.; Bax, A.; Torchia, D. Pulse sequences for removal of the effects of cross-correlation between dipolar and chemical-shift anisotropy relaxation mechanism on the measurement of heteronuclear T1 and T2 values in proteins. J. Magn. Res. 1992, 97, 359–375. [Google Scholar] [CrossRef]
- Farrow, N.A.; Muhandiram, R.; Singer, A.U.; Pascal, S.M.; Kay, C.M.; Gish, G.; Shoelson, S.E.; Pawson, T.; Forman-Kay, J.D.; Kay, L.E. Backbone dynamics of a free and phosphopeptide-complexed Src homology 2 domain studied by 15N NMR relaxation. Biochemistry 1994, 33, 5984–6003. [Google Scholar] [CrossRef]
- Gryk, M.R.; Finucaine, M.D.; Zheng, Z.; Jardetzky, O. Solution dynamics of the trp repressor: A study of amide proton exchange by T1 relaxation. J. Mol. Biol. 1995, 246, 618–627. [Google Scholar] [CrossRef]
- Musacchio, A. How SH3 domains recognize proline. Adv. Protein Chem. 2002, 61, 211–268. [Google Scholar]
- Englander, S.W.; Kallenbach, N.R. Hydrogen exchange and structural dynamics of proteins and nucleic acids. Q. Rev. Biophys. 1983, 16, 521–655. [Google Scholar] [CrossRef] [PubMed]
- Gryk, M.R.; Jardetzky, O. AV77 hinge mutation stabilizes the helix-turnhelix domain of trp repressor. J. Mol. Biol. 1996, 255, 204–214. [Google Scholar] [CrossRef] [PubMed]
- Korzhnev, D.M.; Salvatella, X.; Vendruscolo, M.; Di Nardo, A.A.; Davidson, A.R.; Dobson, C.M.; Kay, L.E. Low-populated folding intermediates of Fyn SH3 characterized by relaxation dispersion NMR. Nature 2004, 430, 586–590. [Google Scholar] [CrossRef] [PubMed]
- Dayhoff, M.O.; Schwartz, R.M.; Orcutt, B.C. A model of evolutionary change in proteins. In Atlas of Protein Sequence and Structure; Dayhoff, M.O., Ed.; National Biomedical Science Foundation: Washington, DC, USA, 1978; Volume 5, Suppl. 3, pp. 345–352. [Google Scholar]
- Feng, S.; Chen, J.K.; Yu, H.; Simon, J.A.; Schreiber, S.L. Two binding orientations for peptides to the Src SH3 domain: Development of a general model for SH3–ligand interactions. Science 1994, 266, 1241–1247. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Knudsen, B.; Feller, S.M.; Zheng, J.; Sali, A.; Cowburn, D.; Hanafusa, H.; Kuriyan, J. Structural basis for the specific interaction of lysine-containing proline-rich peptides with the N-terminal SH3 domain of c-Crk. Structure 1995, 3, 215–226. [Google Scholar] [CrossRef] [Green Version]
- Teyra, J.; Huang, H.; Jain, S.; Guan, X.; Dong, A.; Liu, Y.; Tempel, W.; Min, J.; Tong, Y.; Kim, P.M.; et al. Comprehensive analysis of the human SH3 domain reveals a wide variety of non-canonical specificities. Structure 2017, 25, 1598–1610. [Google Scholar] [CrossRef] [Green Version]
- Li, S.S. Specificity and versatility of SH3 and other proline-recognition domains: Structural basis and implications for cellular signal transduction. Biochem. J. 2005, 390, 641–653. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.; Chen, J.K.; Feng, S.; Dalgarno, D.C.; Brauer, A.W.; Schreiber, S.L. Structural basis for the binding of proline-rich peptides to SH3 domains. Cell 1994, 76, 933–945. [Google Scholar] [CrossRef]
- Lim, W.A.; Richards, F.M.; Fox, R.O. Structural determinants of peptide-binding orientation and of sequence specificity in SH3 domains. Nature 1994, 372, 375–379. [Google Scholar] [CrossRef] [PubMed]
- Moncalian, G.; Cardenes, N.; Deribe, Y.L.; Spinola-Amilibia, M.; Dikic, I.; Bravo, J. Atypical polyproline recognition by the CMS N-terminal Src homology 3 domain. J. Biol. Chem. 2006, 281, 38845–38853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mongiovi, A.M.; Romano, P.R.; Panni, S.; Mendoza, M.; Wong, W.T.; Musacchio, A.; Cesareni, G.; Di Fiore, P.P. A novel peptide-SH3 interaction. EMBO J. 1999, 18, 5300–5309. [Google Scholar] [CrossRef] [Green Version]
- Kato, M.; Miyazawa, K.; Kitamura, N. A deubiquitinating enzyme UBPY interacts with the Src homology 3 domain of Hrs-binding protein via a novel binding motif PX(V/I)(D/N)RXXKP. J. Biol. Chem. 2000, 275, 37481–37487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Q.; Berry, D.; Nash, P.; Pawson, T.; McGlade, C.J.; Li, S.S. Structural basis for specific binding of the Gads SH3 domain to an RxxK motif-containing SLP-76 peptide: A novel mode of peptide recognition. Mol. Cell. 2003, 11, 471–481. [Google Scholar] [CrossRef]
- Harkiolaki, M.; Lewitzky, M.; Gilbert, R.J.; Jones, E.Y.; Bourette, R.P.; Mouchiroud, G.; Sondermann, H.; Moarefi, I.; Feller, S.M. Structural basis for SH3 domain-mediated high-affinity binding between Mona/Gads and SLP-76. EMBO J. 2003, 22, 2571–2582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, H.; Freund, C.; Duke-Cohan, J.S.; Musacchio, A.; Wagner, G.; Rudd, C.E. SH3 domain recognition of a proline-independent tyrosine-based RKxxYxxY motif in immune cell adaptor SKAP55. EMBO J. 2000, 19, 2889–2899. [Google Scholar] [CrossRef] [Green Version]
- Duke-Cohan, J.S.; Kang, H.; Liu, H.; Rudd, C.E. Regulation and function of SKAP-55 non-canonical motif binding to the SH3c domain of adhesion and degranulationpromoting adaptor protein. J. Biol. Chem. 2006, 281, 13743–13750. [Google Scholar] [CrossRef] [Green Version]
- Merő, B.; Radnai, L.; Gógl, G.; Tőke, O.; Leveles, I.; Koprivanacz, K.; Szeder, B.; Dülk, M.; Kudlik, G.; Vas, V.; et al. Structural insights into the regulation of SH3 domain function by phosphorylation of conserved tyrosine residues within the ligand binding groove. J. Biol. Chem. 2019, 294, 4608–4620. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Hicke, L.; Radhakrishnan, I. Structural basis for ubiquitin recognition by SH3 domains. J. Mol. Biol. 2007, 73, 190–196. [Google Scholar] [CrossRef] [Green Version]
- Chan, B.; Lanyi, A.; Song, H.K.; Griesbach, J.; Simarro-Grande, M.; Poy, F.; Howie, D.; Sumegi, J.; Terhorst, C.; Eck, M.J. SAP couples Fyn to SLAM immune receptors. Nat. Cell. Biol. 2003, 5, 155–160. [Google Scholar] [CrossRef]
- Heuer, K.; Arbuzova, A.; Strauss, H.; Kofler, M.; Freund, C. The helically extended SH3 domain of the T cell adaptor protein ADAP is a novel lipid interaction domain. J. Mol. Biol. 2005, 348, 1025–1035. [Google Scholar] [CrossRef] [PubMed]
- Heuer, K.; Sylvester, M.; Kliche, S.; Pusch, R.; Thiemke, K.; Schraven, B.; Freund, C. Lipid-binding hSH3 domains in immune cell adapter proteins. J. Mol. Biol. 2006, 361, 94–104. [Google Scholar] [CrossRef] [PubMed]
- Pérez, Y.; Maffei, M.; Igea, A.; Amata, I.; Gairí, M.; Nebreda, A.R.; Bernadó, P.; Pons, M. Lipid binding by the Unique and SH3 domains of c-Src suggests a new regulatory mechanism. Sci. Rep. 2013, 3, 1295. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Distance restraints from NOEs | |
| unambiguous | 809 |
| intraresidue | 307 |
| sequential | 152 |
| i–i + j, where j = 2, 3, or 4 | 74 |
| i–i + j, where j > 4 | 276 |
| ambiguous | 180 |
| Ensemble RMSD values | |
| All backbone atoms | 1.3 Å |
| All heavy atoms | 1.8 Å |
| All backbone atoms in ordered regions | 0.8 Å |
| All heavy atoms in ordered regions | 1.3 Å |
| Statistics | |
| Ramachandran plot statistics (ordered protein regions) | |
| Residues in most favored regions [A, B, L], % | 81.5 |
| Residues in additionally allowed regions [a, b, l, p], % | 18.3 |
| Residues in generously allowed regions [~a, ~b, ~l, ~p], % | 0.2 |
| Residues in disallowed regions, % | 0.0 |
| Main-chain statistics | |
| SD of ω angle, degrees | 3.9 |
| Bad contacts/100 residues | 0 |
| Cα chirality, SD of ζ angle, degrees | 1.2 |
| SD of H-bond energy, kcal/mol | 0.9 |
| Overall G-factor | −0.1 |
| Side-chain statistics | |
| χ-1 gauche minus SD, degrees | 7.3 |
| χ-1 trans SD, degrees | 9.1 |
| χ-1 gauche plus SD, degrees | 9.7 |
| χ-1 pooled SD, degrees | 10.7 |
| χ-2 trans SD, degrees | 12.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tőke, O.; Koprivanacz, K.; Radnai, L.; Merő, B.; Juhász, T.; Liliom, K.; Buday, L. Solution NMR Structure of the SH3 Domain of Human Caskin1 Validates the Lack of a Typical Peptide Binding Groove and Supports a Role in Lipid Mediator Binding. Cells 2021, 10, 173. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10010173
Tőke O, Koprivanacz K, Radnai L, Merő B, Juhász T, Liliom K, Buday L. Solution NMR Structure of the SH3 Domain of Human Caskin1 Validates the Lack of a Typical Peptide Binding Groove and Supports a Role in Lipid Mediator Binding. Cells. 2021; 10(1):173. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10010173
Chicago/Turabian StyleTőke, Orsolya, Kitti Koprivanacz, László Radnai, Balázs Merő, Tünde Juhász, Károly Liliom, and László Buday. 2021. "Solution NMR Structure of the SH3 Domain of Human Caskin1 Validates the Lack of a Typical Peptide Binding Groove and Supports a Role in Lipid Mediator Binding" Cells 10, no. 1: 173. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10010173