
Ejaculate Allocation and Sperm Characteristics Differ among Alternative Male Types in a Species of Fish with Cooperation and Competition among Unrelated Males

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site and General Information
2.2. Absolute and Relative Allocation to Gonads
2.3. Ejaculate Production and Regeneration
2.4. Sperm Morphology
3. Results
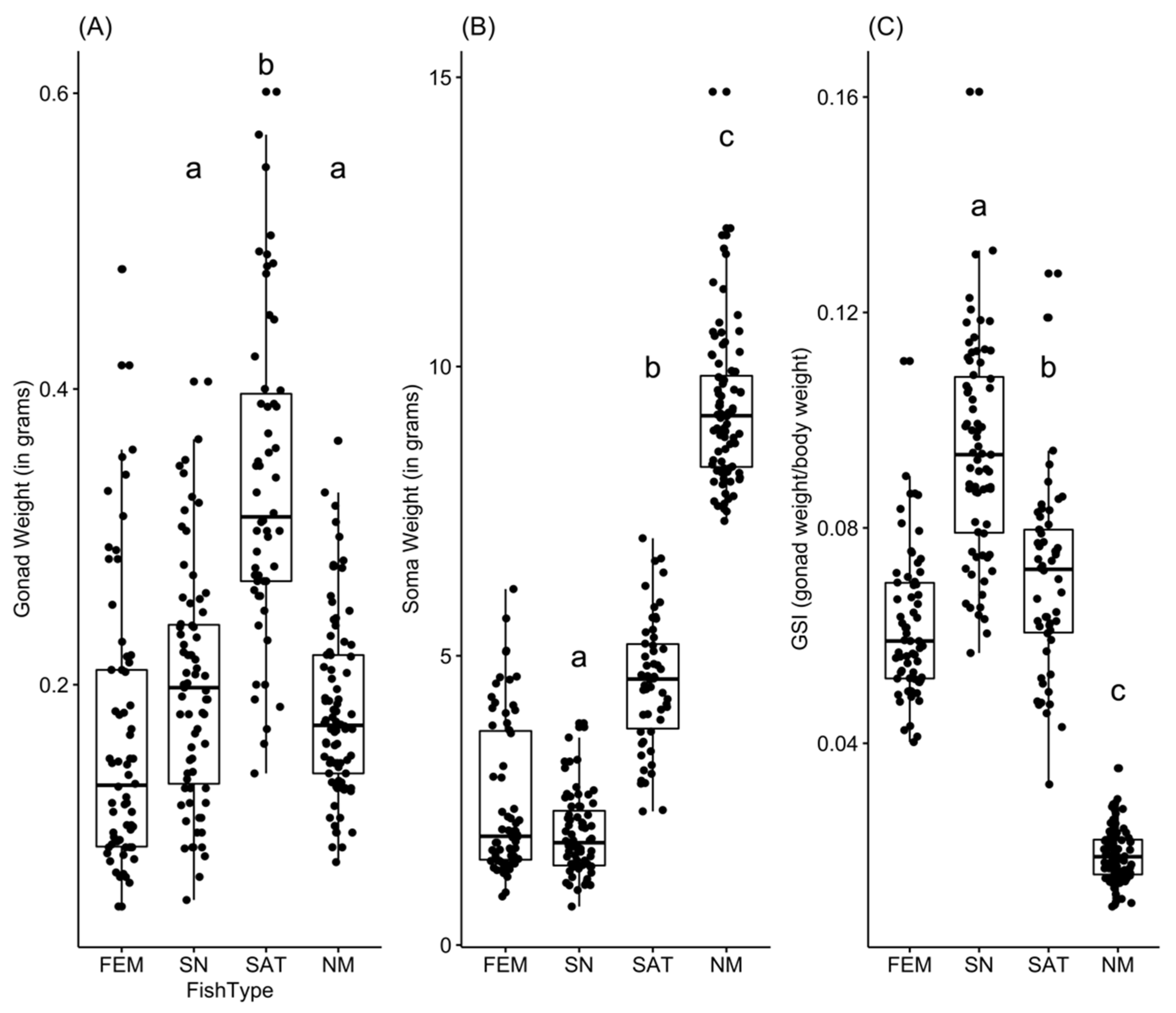
3.1. Comparing Absolute Testes Weight and Soma Weight among Male Types
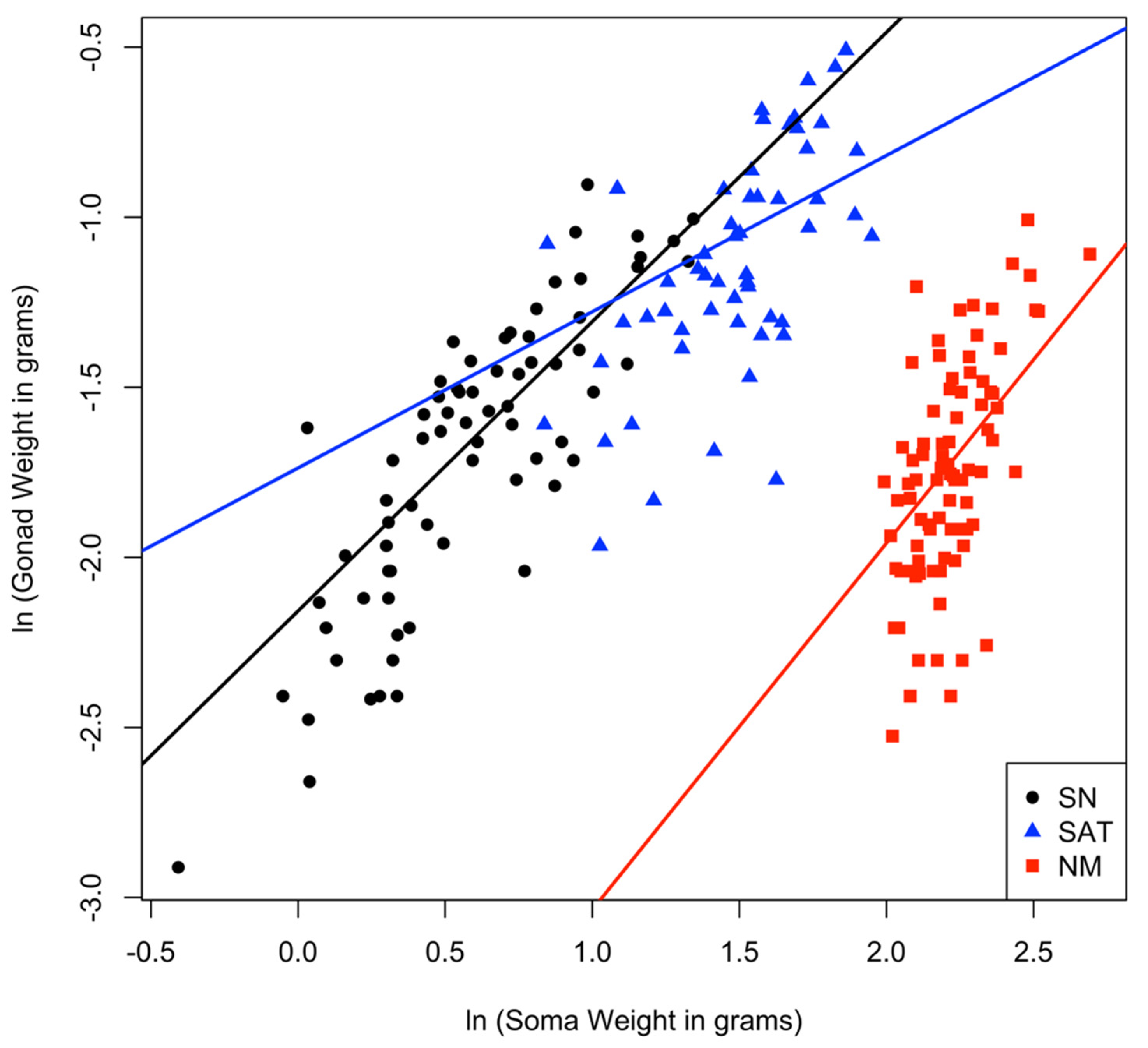
3.2. Comparing Relative Testes Weight among Male Types
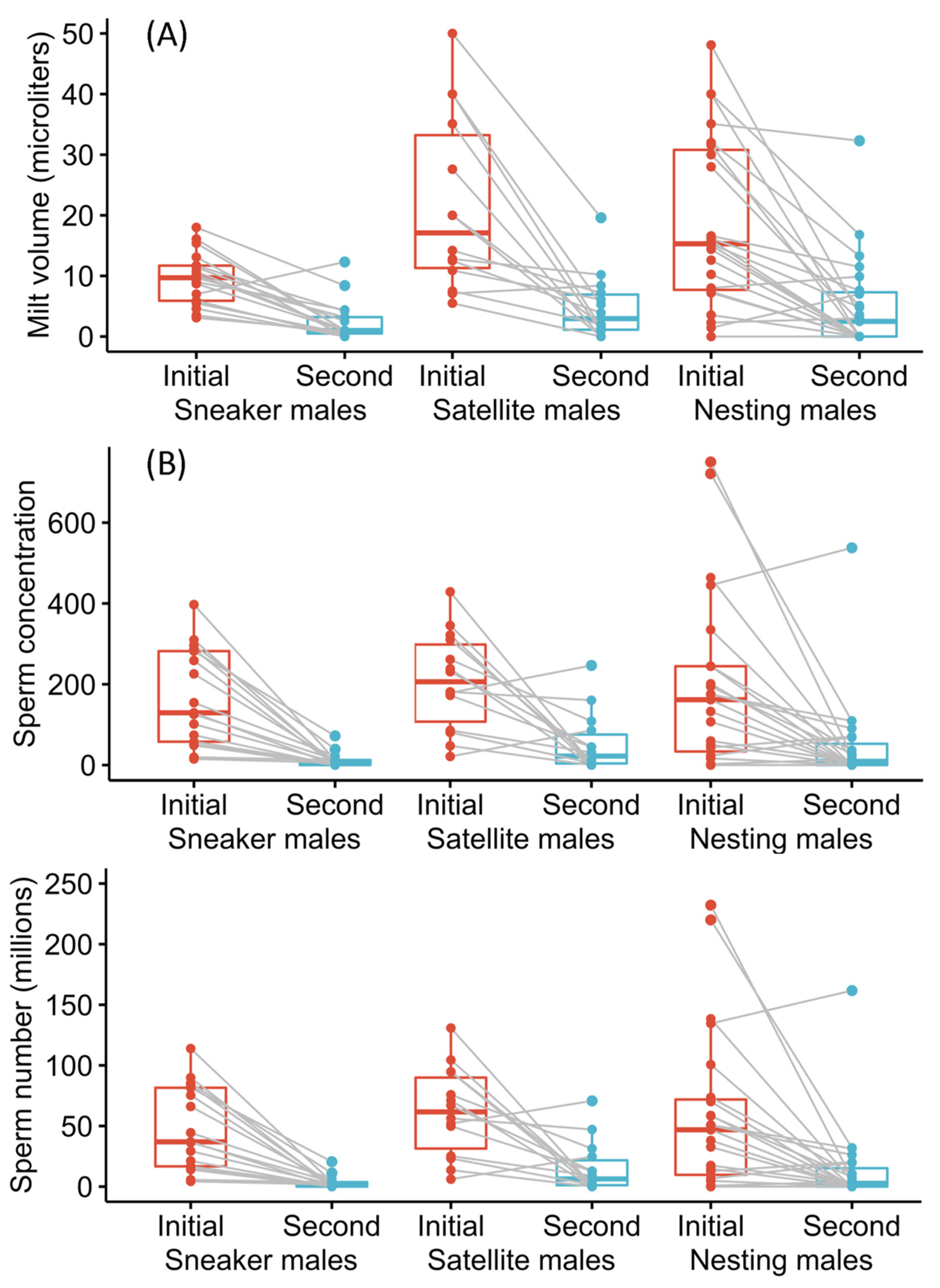
3.3. Ejaculate Production and Regeneration
3.3.1. Initial Ejaculate Production
3.3.2. Ejaculate Regeneration
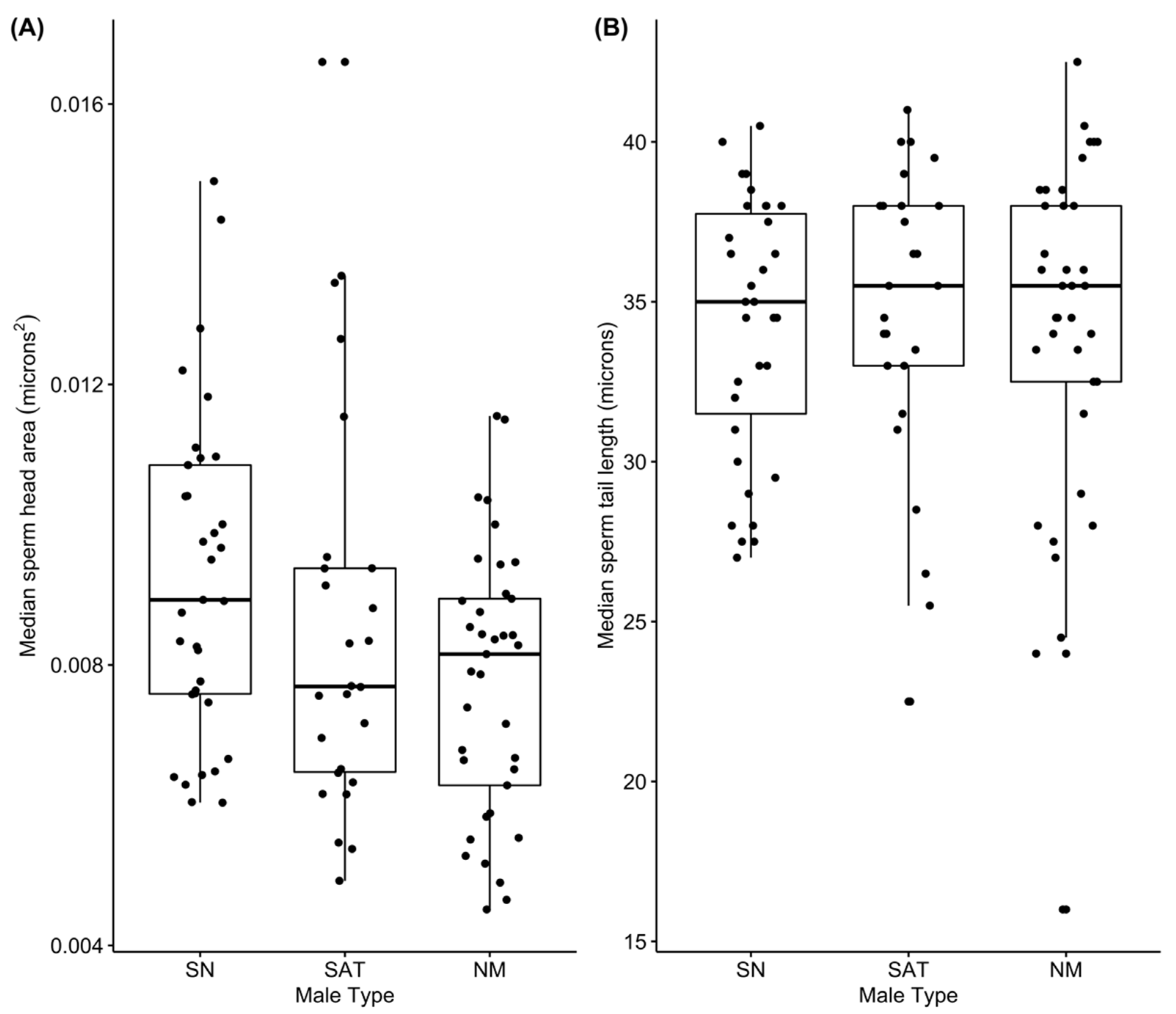
3.4. Sperm Morphology
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Parker, G.A. Sperm competition games: Raffles and roles. Proc. R. Soc. B 1990, 242, 120–126. [Google Scholar]
- Parker, G.A. Sperm competition games: Sneaks and extra-pair copulations. Proc. R. Soc. B 1990, 242, 127–133. [Google Scholar]
- Gage, M.J.; Stockley, P.; Parker, G.A. Effects of alternative male mating strategies on characteristics of sperm production in the Atlantic salmon (Salmo salar): Theoretical and empirical investigations. Philos. Trans. R. Soc. 1995, 350, 391–399. [Google Scholar]
- Stockley, P.; Gage, M.J.G.; Parker, G.A.; Møller, A.P. Sperm competition in fishes: The evolution of testis size and ejaculate characteristics. Am. Nat. 1997, 149, 933–954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parker, G.A. Sperm Competition and the Evolution of Ejaculates: Towards a Theory Base. In Sperm Competition and Sexual Selection; Birkhead, T.R., Møller, A.P., Eds.; Academic Press: London, UK, 1998; pp. 3–54. [Google Scholar]
- Birkhead, T.R.; Pizzari, T. Postcopulatory sexual selection. Nat. Rev. Genet. 2002, 3, 262–273. [Google Scholar] [CrossRef]
- Birkhead, T.R.; Hosken, D.J.; Pitnick, S. Sperm Biology: An Evolutionary Perspective; Elsevier Press: Burlington, VT, USA, 2009. [Google Scholar]
- Parker, G.A.; Pizzari, T. Sperm competition and ejaculate economics. Biol. Rev. 2010, 85, 897–934. [Google Scholar] [CrossRef]
- Baker, J.; Humphries, S.; Ferguson-Gow, H.; Meade, A.; Venditty, C. Rapid decreases in relative testes mass among monogamous birds but not in other vertebrates. Ecol. Lett. 2020, 23, 283–292. [Google Scholar] [CrossRef]
- Pitnick, S.; Hosken, D.J.; Birkhead, T.R. Sperm morphological diversity. In Sperm Biology: An Evolutionary Perspective; Birkhead, T.R., Hosken, D.J., Pitnick, S., Eds.; Elsevier Press: Burlington, VT, USA, 2009; pp. 69–149. [Google Scholar]
- Lüpold, S.; Pitnick, S. Sperm form and function: What do we know about the role of sexual selection? Reproduction 2018, 155, R229–R243. [Google Scholar] [CrossRef]
- Swanson, W.J.; Vacquier, V.D. The rapid evolution of reproductive proteins. Nat. Rev. Genet. 2002, 3, 137–144. [Google Scholar] [CrossRef]
- Ramm, S.A.; Oliver, P.L.; Ponting, C.P.; Stockley, P.; Emes, R.D. Sexual selection and the adaptive evolution of mammalian ejaculate proteins. Mol. Biol. Evol. 2007, 25, 207–219. [Google Scholar] [CrossRef] [Green Version]
- Rowe, M.; Whittington, E.; Borziak, K.; Ravinet, M.; Eroukhmanoff, F.; Sætre, G.; Dorus, S. Molecular diversification of the seminal fluid proteome in a recently diverged passerine species pair. Mol. Biol. Evol. 2020, 37, 488–506. [Google Scholar] [CrossRef]
- Harcourt, A.H.; Harvey, P.H.; Larson, S.G.; Short, R.V. Testis weight, body weight and breeding system in primates. Nature 1981, 293, 55–57. [Google Scholar] [CrossRef]
- Møller, A.P. Sperm competition, sperm depletion, paternal care, and relative testis size in birds. Am. Nat. 1991, 137, 882–906. [Google Scholar] [CrossRef]
- Jennions, M.D.; Passmore, N.E. Sperm competition in frogs: Testis size and a ‘sterile male’ experiment on Chiromantis xerampelina (Rhacophoridae). Biol. J. Linn. Soc. 1993, 40, 211–220. [Google Scholar]
- Hosken, D.J. Sperm competition in bats. Proc. R. Soc. B 1997, 264, 385–392. [Google Scholar] [CrossRef]
- Taborsky, M. Sneakers, satellites, and helpers: Parasitic and cooperative behavior in fish reproduction. Adv. Study Behav. 1994, 23, 1–100. [Google Scholar]
- Taborsky, M.; Schutz, D.; Goffinet, O.; van Doorn, G.S.; Schütz, D.; Goffinet, O.; Sander van Doorn, G. Alternative male morphs solve sperm performance/longevity trade-off in opposite directions. Sci. Adv. 2018, 4, eaap8563. [Google Scholar] [CrossRef] [Green Version]
- Petersen, C.W.; Warner, R.R. Sperm competition in fishes. In Sperm Competition and Sexual Selection; Birkhead, T.R., Møller, A.P., Eds.; Academic Press: London, UK, 1998; pp. 435–463. [Google Scholar]
- Schradin, C.; Kinahan, A.A.; Pillay, N. Cooperative breeding in groups of synchronously mating females and evolution of large testes to avoid sperm depletion in African striped mice. Biol. Reprod. 2009, 81, 111–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kustra, M.C.; Alonzo, S.H. Sperm and alternative reproductive tactics: A review of existing theory and empirical data. Philos. Trans. R. Soc. B 2020, 375, 20200075. [Google Scholar] [CrossRef] [PubMed]
- Cornwallis, C.K.; Birkhead, T.R. Changes in sperm quality and numbers in response to experimental manipulation of male social status and female attractiveness. Am. Nat. 2007, 170, 758–770. [Google Scholar] [CrossRef] [PubMed]
- Awata, S.; Takeyama, T.; Makino, Y.; Kitamura, Y.; Kohda, M. Cooperatively breeding cichlid fish adjust their testis size but not sperm traits in relation to sperm competition risk. Behav. Ecol. Sociobiol. 2008, 62, 1701–1710. [Google Scholar] [CrossRef]
- Kustan, J.M.; Maruska, K.P.; Fernald, R.D. Subordinate male cichlids retain reproductive competence during social suppression. Proc. R. Soc. B 2012, 279, 434–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelly, C.D.; Jennions, M.D. Sexual selection and sperm quantity: Meta-analyses of strategic ejaculation. Biol. Rev. 2011, 86, 863–884. [Google Scholar] [CrossRef]
- Miller, J.S.; Bose, A.P.H.; Fitzpatrick, J.L.; Balshine, S. Sperm maturation and male tactic-specific differences in ejaculates in the plainfin midshipman fish Porichthys notatus. J. Fish. Biol. 2019, 94, 434–445. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.S.; Mazzoldi, C.; Rasotto, M.B.; Balshine, S. Differential investment in male accessory glands: Lessons from a marine fish with alternative reproductive tactics. Mar. Biol. 2019, 166, 37. [Google Scholar] [CrossRef]
- Magris, M. Strategic adjustment of ejaculate quality in response to variation of the socio-sexual environment. Behav. Ecol. Sociobiol. 2021, 75, 91. [Google Scholar] [CrossRef]
- Fitzpatrick, J.L.; Montgomerie, R.; Desjardins, J.K.; Stiver, K.A.; Kolm, N.; Balshine, S. Female promiscuity promotes the evolution of faster sperm in cichlid fishes. Proc. Natl. Acad. Sci. USA 2009, 106, 1128–1132. [Google Scholar] [CrossRef] [Green Version]
- Fitzpatrick, J.L.; Lüpold, S. Sexual selection and the evolution of sperm quality. Mol. Hum. Reprod. 2014, 20, 1180–1189. [Google Scholar] [CrossRef] [Green Version]
- Lüpold, S. Ejaculate quality and constraints in relation to sperm competition levels among eutherian mammals. Evolution 2013, 67, 3052–3060. [Google Scholar] [CrossRef]
- Simmons, L.W.; Roberts, J.D.; Dziminski, M.A. Egg jelly influences sperm motility in the externally fertilizing frog, Crinia Georgiana. J. Evol. Biol. 2009, 22, 225–229. [Google Scholar] [CrossRef]
- Hettyey, A.; Roberts, J.D. Sperm traits in the quacking frog (Crinia georgiana), a species with plastic alternative mating tactics. Behav. Ecol. Sociobiol. 2007, 61, 1303–1310. [Google Scholar] [CrossRef]
- Montgomerie, R.; Fitzpatrick, J.L. Testes, Sperm, and Sperm Competition. In Reproductive Biology and Phylogeny of Fishes (Agnathans and Bony Fishes); Jamieson, B.G.M., Ed.; CRC Press: Boca Rotan, FL, USA, 2009; pp. 1–53. [Google Scholar]
- Sefc, K. Mating and parental care in Lake Tanganyika’s cichlids. Int. J. Evol. Biol. 2011, 2011, 470875. [Google Scholar] [CrossRef] [Green Version]
- Awata, S.; Heg, D.; Munehara, H.; Kohdaa, M. Testis size depends on social status and the presence of male helpers in the cooperatively breeding cichlid Julidochromis ornatus. Behav. Ecol. 2006, 17, 372–379. [Google Scholar] [CrossRef] [Green Version]
- Fitzpatrick, J.L.; Desjardins, J.K.; Stiver, K.A.; Montgomerie, R.; Balshine, S. Male reproductive suppression in the cooperatively breeding fish Neolamprologus pulcher. Behav. Ecol. 2006, 17, 25–33. [Google Scholar] [CrossRef]
- Díaz-Muñoz, S.L.; DuVal, E.H.; Krakauer, A.H.; Lacey, E.A. Cooperating to compete: Altruism, sexual selection and causes of male reproductive cooperation. Anim. Behav. 2014, 88, 67–78. [Google Scholar] [CrossRef] [Green Version]
- Stiver, K.A.; Alonzo, S.H. Does the risk of sperm competition help explain cooperation between reproductive competitors? A study in the ocellated wrasse (Symphodus ocellatus). Am. Nat. 2013, 181, 357–368. [Google Scholar] [CrossRef] [PubMed]
- Foster, K.R.; Pizzari, T. Sperm Sociality: Cooperation, altruism, and spite. PLoS Biol. 2008, 6, e130. [Google Scholar]
- Fisher, H.S.; Hoekstra, H.E. Competition drives cooperation among closely related sperm of deer mice. Nature 2010, 463, 801–803. [Google Scholar] [CrossRef]
- Fisher, H.S.; Giomi, L.; Hoekstra, H.E.; Mahadevan, L. The dynamics of sperm cooperation in a competitive environment. Proc. R. Soc. B 2014, 281, 20140296. [Google Scholar] [CrossRef] [Green Version]
- Sutter, A.; Immler, S. Within-ejaculate sperm competition. Philos. Trans. R. Soc. B 2020, 375, 20200066. [Google Scholar] [CrossRef]
- Lejeune, P. Etude écoéthologique des comportements reproducteur et sociaux des Labridae méditerranéens des genres Symphodus (Rafinesque 1810) et Coris (Lacepede 1802). Cah. D’ethologie Appl. 1985, 5, 1–208. [Google Scholar]
- Alonzo, S.H. Uncertainty in territory quality affects the benefits of usurpation in a Mediterranean wrasse. Behav. Ecol. 2004, 15, 278–285. [Google Scholar] [CrossRef] [Green Version]
- Alonzo, S.H.; Warner, R.R. A trade-off generated by sexual conflict: Mediterranean wrasse males refuse present mates to increase future success. Behav. Ecol. 1999, 10, 105–111. [Google Scholar] [CrossRef] [Green Version]
- Alonzo, S.H.; Warner, R.R. Dynamic games and field experiments examining intra- and intersexual conflict: Explaining counter-intuitive mating behavior in a Mediterranean wrasse, Symphodus ocellatus. Behav. Ecol. 2000, 11, 56–70. [Google Scholar] [CrossRef]
- Taborsky, M.; Hudde, B.; Wirtz, P. 1987. Reproductive behavior and ecology of Symphodus (Crenilabrus) Ocellatus, a European wrasse with 4 types of male behavior. Behaviour 1987, 102, 82–118. [Google Scholar] [CrossRef]
- Hellmann, J.K.; Stiver, K.S.; Marsh-Rollo, S.E.; Alonzo, S.H. Defense against outside competition is linked to cooperation and conflict in male-male partnerships. Behav. Ecol. 2020, 31, 432–439. [Google Scholar] [CrossRef]
- Alonzo, S.H.; Taborsky, M.; Wirtz, P. Male alternative reproductive behaviors in a Mediterranean wrasse, Symphodus ocellatus: Evidence from otoliths for separate life-history pathways. Evol. Ecol. Res. 2000, 2, 997–1007. [Google Scholar]
- Stiver, K.A.; Harris, R.M.; Townsend, J.P.; Hofmann, H.A.; Alonzo, S.H. Neural gene expression profiles and androgen levels underlie alternative reproductive tactics in the ocellated wrasse, Symphodus ocellatus. Ethology 2015, 120, 1–16. [Google Scholar] [CrossRef]
- Nugent, B.M.; Stiver, K.A.; Alonzo, S.H.; Hofmann, H.A. Neuroendocrine profiles associated with discrete behavioral variation in Symphodus ocellatus, a species with male alternative reproductive tactics. Mol. Ecol. 2016, 25, 5212–5227. [Google Scholar] [CrossRef]
- Dean, R.; Wright, A.E.; Marsh-Rollo, S.E.; Nugent, B.M.; Kindsvater, H.K.; Alonzo, S.H.; Mank, J.E. Sperm competition shapes gene expression and sequence evolution in the ocellated wrasse. Mol. Ecol. 2017, 26, 505–518. [Google Scholar] [CrossRef] [Green Version]
- Alonzo, S.H.; Warner, R.R. Allocation to mate guarding or increased sperm production in a Mediterranean wrasse. Am. Nat. 2000, 156, 266–275. [Google Scholar] [CrossRef]
- Warner, R.R.; Lejeune, P. Sex change limited by paternal care: A test using four Mediterranean labrid fishes, genus Symphodus. Mar. Biol. 1985, 87, 89–100. [Google Scholar] [CrossRef]
- Tomkins, J.L.; Simmons, L.W. Measuring relative investment: A case study of testes investment in species with alternative male reproductive tactics. Animal. Behav. 2002, 63, 1009–1016. [Google Scholar] [CrossRef] [Green Version]
- Alonzo, S.H.; Heckman, K.L. The unexpected but understandable dynamics of mating, paternity and paternal care in the ocellated wrasse. Proc. R. Soc. B 2010, 277, 115–122. [Google Scholar] [CrossRef] [Green Version]
- Milazzo, M.; Cattano, C.; Alonzo, S.H.; Gristina, M.; Rodolfo-Metalpa, R.; Sinopoli, M.; Spatafora, D.; Stiver, K.A.; Hall-Spencer, J.M. Ocean acidification affects fish spawning but not paternity at CO2 seeps. Proc. R. Soc. B 2016, 283, 20161021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stiver, K.S.; Kindsvater, H.K.; Tamburello, N.; Heckman, K.L.; Klein, J.; Alonzo, S.H. Intentional multiple mating by females in a species where sneak fertilization circumvents female choice for parental males. J. Fish. Biol. 2018, 93, 324–333. [Google Scholar] [CrossRef] [PubMed]
- Nugent, B.M.; Stiver, K.A.; Hofmann, H.A.; Alonzo, S.H. Experimentally-induced variation in neuroendocrine processes affects male reproductive behavior, sperm characteristics, and social interactions. Mol. Ecol. 2019, 28, 3464–3481. [Google Scholar] [CrossRef] [PubMed]
- Alonzo, S.H.; Stiver, K.A.; Marsh-Rollo, S.E. Ovarian fluid allows directional cryptic female choice despite external fertilization. Nat. Commun. 2016, 7, 12452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tabachnick, B.G.; Fidell, L.S. Using Multivariate Statistics, 4th ed.; Allyn and Bacon: Boston, MA, USA, 2001. [Google Scholar]
- Field, A.; Miles, J.; Field, Z. Discovering Statistics Using R; Sage Publications Ltd.: London, UK, 2012. [Google Scholar]
- Zuur, A.; Ieno, E.N.; Walker, N.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R; Springer Science & Business Media: New York, NY, USA, 2009. [Google Scholar]
- Neff, B.D.; Fu, P.; Gross, M.R. Sperm investment and alternative mating tactics in bluegill sunfish (Lepomis macrochirus). Behav. Ecol. 2003, 14, 634–641. [Google Scholar] [CrossRef]
- Kvarnemo, C.; Svensson, O.; Manson, W. Investment in testes, sperm-duct glands and lipid reserves differs between male morphs but not between early and late breeding season in Pomatoschistus Minutus. J. Fish. Biol. 2010, 76, 1609–1625. [Google Scholar] [CrossRef]
- Awata, S.; Munehara, H.; Kohda, M. Social system and reproduction of helpers in a cooperatively breeding cichlid fish (Julidochromis ornatus) in Lake Tanganyika: Field observations and parentage analyses. Behav. Ecol. Sociobiol. 2005, 58, 506–516. [Google Scholar] [CrossRef]
- Küpper, C.; Stocks, M.; Risse, J.E.; dos Remedios, N.; Farrell, L.L.; McRae, S.B.; Morgan, T.C.; Karlionova, N.; Pinchuk, P.; Verkuil, Y.I.; et al. A supergene determines highly divergent male reproductive morphs in the ruff. Nat. Genet. 2016, 48, 79–83. [Google Scholar] [CrossRef] [Green Version]
- Stiver, K.A.; Fitzpatrick, J.L.; Desjardins, J.K.; Balshine, S. Mixed parentage in Neolamprologus pulchar groups. J. Fish. Biol. 2009, 74, 1129–1135. [Google Scholar] [CrossRef]
- Locatello, L.; Poli, F.; Rasotto, M.B. Tactic-specific differences in seminal fluid influence sperm performance. Proc. R. Soc. B 2013, 280, 20122891. [Google Scholar] [CrossRef] [PubMed]
- Arnqvist, G.; Rowe, L. Sexual Conflict; Princeton University Press: Princeton, NJ, USA, 2005. [Google Scholar]
- Alonzo, S.H.; Pizzari, T. Male fecundity stimulation: Conflict and cooperation within and between the sexes: Model analyses and coevolutionary dynamics. Am. Nat. 2010, 175, 174–185. [Google Scholar] [CrossRef] [PubMed]
- Alonzo, S.H.; Pizzari, T. Selection on female remating interval is influenced by male sperm competition strategies and ejaculate characteristics. Philos. Trans. R. Soc. B 2013, 368, 20120044. [Google Scholar] [CrossRef] [Green Version]
- Carazo, P.; Tan, C.K.W.; Allen, F.; Wigby, S.; Pizzari, T. Within-group male relatedness reduces harm to females in Drosophila. Nature 2014, 505, 672–67530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rankin, D.J. Kin selection and the evolution of sexual conflict. J. Evol. Biol. 2011, 24, 71–81. [Google Scholar] [CrossRef]
- Pitcher, T.E.; Dunn, P.O.; Whittingham, L.A. Sperm competition and the evolution of testes size in birds. J. Evol. Biol. 2005, 18, 557–567. [Google Scholar] [CrossRef] [PubMed]
- Fitzpatrick, J.L. Sperm competition and fertilization mode in fishes. Philos. Trans. R. Soc. B 2020, 375, 20200074. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Estimate | Standard Error | t-Value | p-Value |
|---|---|---|---|---|
| Intercept | −2.16 | 0.08 | −28.4 | <0.001 |
| Male type: satellite | 0.42 | 0.21 | 1.98 | 0.0488 |
| Male type: nesting | −1.96 | 0.46 | −4.28 | <0.001 |
| ln(soma weight) | 0.85 | 0.08 | 10.53 | <0.001 |
| ln(soma weight) × satellite | −0.39 | 0.15 | −2.57 | 0.0108 |
| ln(soma weight) × nesting | 0.23 | 0.22 | 1.05 | 0.2930 |
| Variable | Estimate | Standard Error | df | t-Value | Prob > |t| | |
|---|---|---|---|---|---|---|
| Sperm Cell Head Area (mm2) | Intercept | 0.000009529 | 3.939 × 10−7 | 42.77 | 24.193 | <0.001 |
| Type: satellite | −0.000004820 | 6.037 × 10−7 | 42.82 | −0.798 | 0.429 | |
| Type: nesting | −0.000001770 | 5.495 × 10−7 | 42.77 | −3.221 | 0.002 | |
| Sperm Tail Length (mm) | Intercept | 0.03442 | 0.00113 | 97.33 | 30.232 | <0.001 |
| Type: satellite | 0.00128 | 0.00175 | 97.89 | 0.731 | 0.467 | |
| Type: nesting | −0.00084 | 0.00159 | 97.33 | −0.527 | 0.600 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alonzo, S.H.; Stiver, K.A.; Kindsvater, H.K.; Marsh-Rollo, S.E.; Nugent, B.; Kazancıoğlu, E. Ejaculate Allocation and Sperm Characteristics Differ among Alternative Male Types in a Species of Fish with Cooperation and Competition among Unrelated Males. Cells 2021, 10, 2612. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10102612
Alonzo SH, Stiver KA, Kindsvater HK, Marsh-Rollo SE, Nugent B, Kazancıoğlu E. Ejaculate Allocation and Sperm Characteristics Differ among Alternative Male Types in a Species of Fish with Cooperation and Competition among Unrelated Males. Cells. 2021; 10(10):2612. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10102612
Chicago/Turabian StyleAlonzo, Suzanne H., Kelly A. Stiver, Holly K. Kindsvater, Susan E. Marsh-Rollo, Bridget Nugent, and Erem Kazancıoğlu. 2021. "Ejaculate Allocation and Sperm Characteristics Differ among Alternative Male Types in a Species of Fish with Cooperation and Competition among Unrelated Males" Cells 10, no. 10: 2612. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10102612