
PARP1 as a Marker of an Aggressive Clinical Phenotype in Cutaneous Melanoma—A Clinical and an In Vitro Study

 , , , , , ,
, , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Confocal Microscopy and Image Analysis
2.3. Cell Lysate Preparation
2.4. Western Blotting Analysis
2.5. RNA Isolation and Reverse Transcription
2.6. Real-Time PCR Gene Expression Analysis
2.7. Immunohistochemistry
2.8. Patients
2.9. Statistical Analysis
3. Results
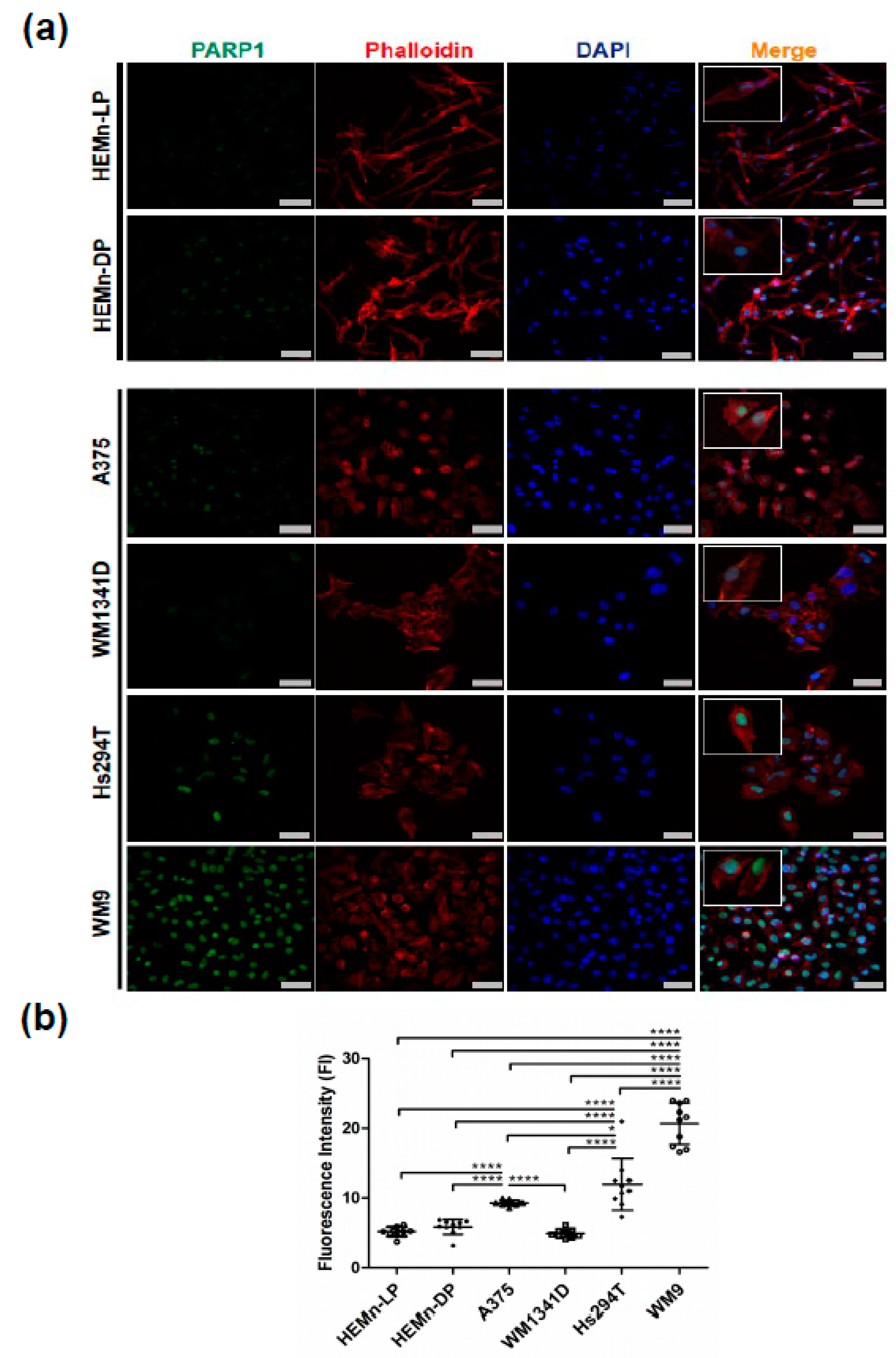
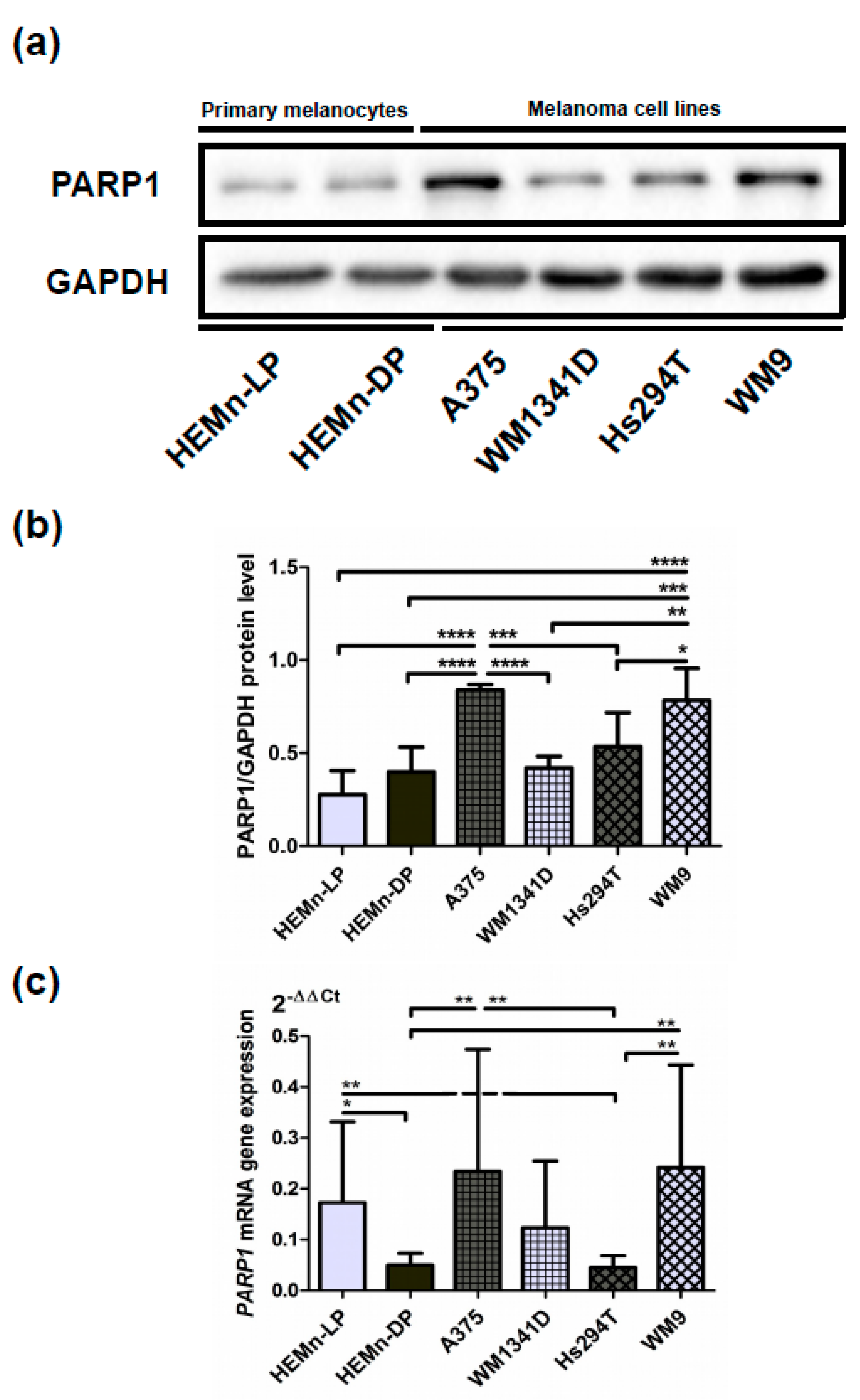
3.1. PARP1 in Normal Melanocyte and Malignant Melanoma Cell Lines
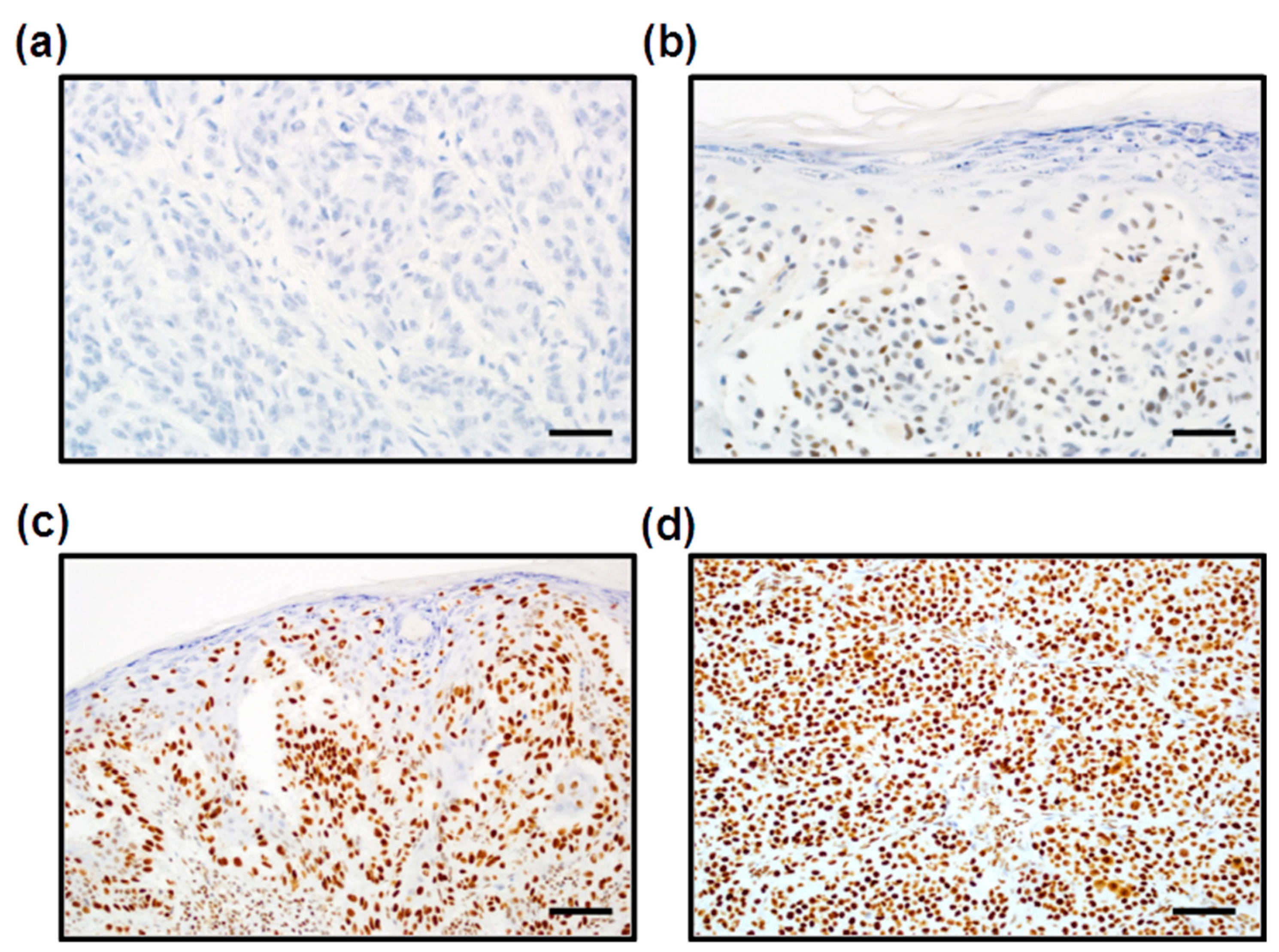
3.2. Expression of PARP1 in Cutaneous Melanoma Patients
3.3. Correlation between PARP1 Expression and Clinicopathologic Parameters of Primary Tumors
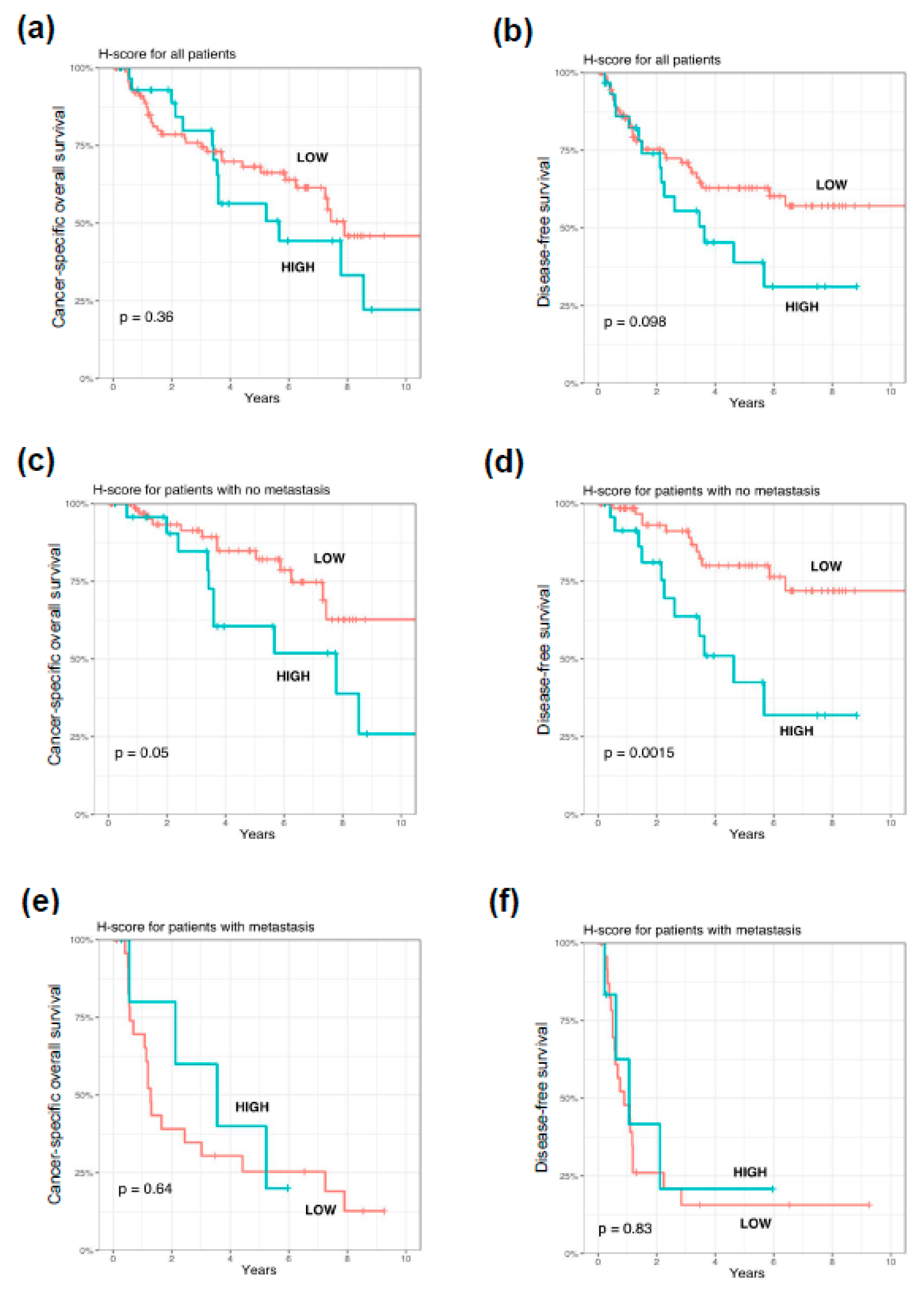
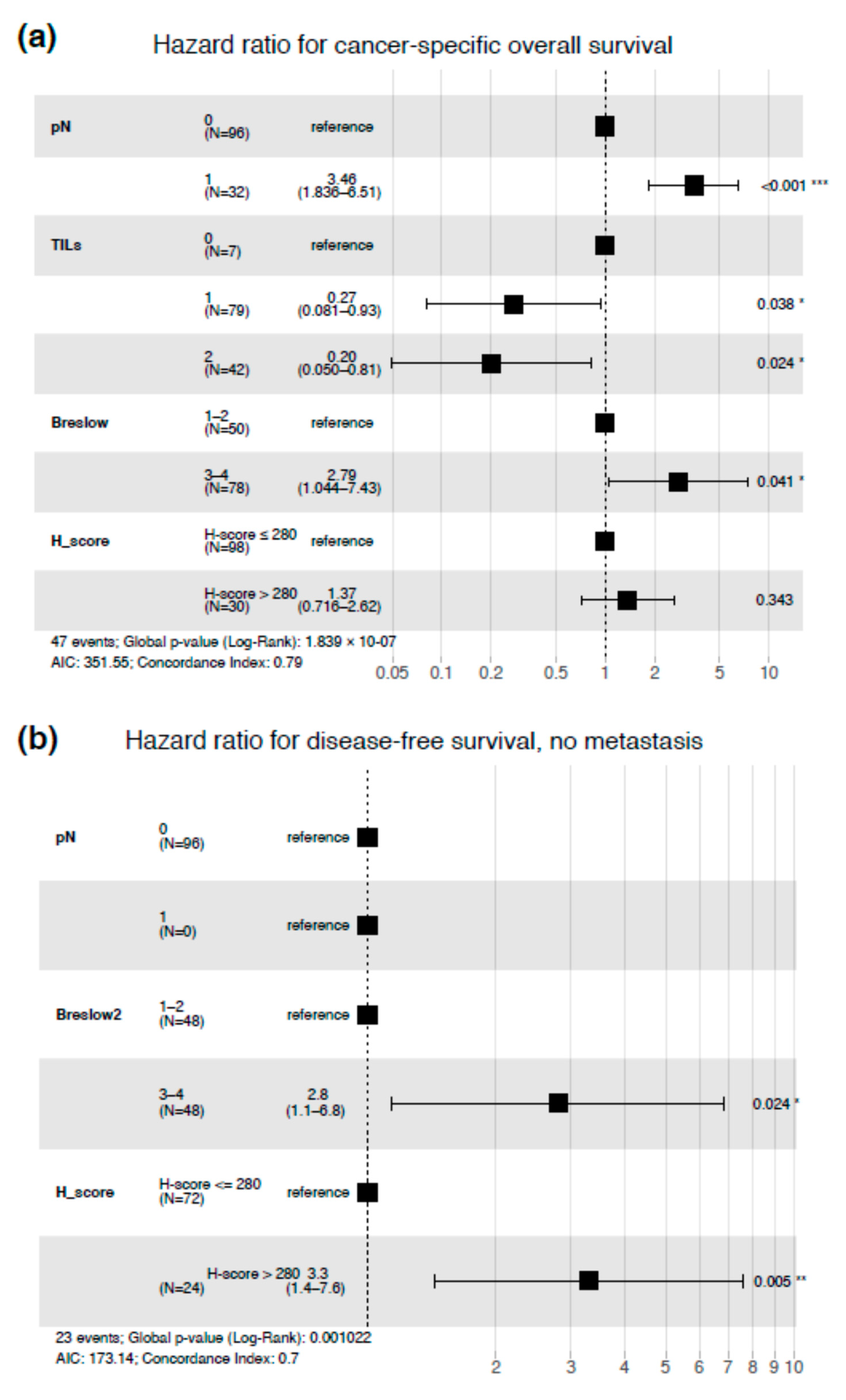
3.4. Impact of PARP1 Expression on Long-Term Survival in Cutaneous Melanoma Patients
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Matthews, N.H.; Li, W.-Q.; Qureshi, A.A.; Weinstock, M.A.; Cho, E. Epidemiology of Melanoma. In Cutaneous Melanoma: Etiology and Therapy; Wb Saunders: Filadelfia, PA, USA, 2017; pp. 3–22. [Google Scholar]
- Lugowska, I.; Teterycz, P.; Rutkowski, P. Immunotherapy of melanoma. Współczesna Onkol. 2018, 2018, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Moreira, A.; Heinzerling, L.; Bhardwaj, N. Current melanoma treatments: Where do we stand? Cancers 2021, 13, 221. [Google Scholar] [CrossRef] [PubMed]
- Simiczyjew, A.; Dratkiewicz, E.; Mazurkiewicz, J.; Ziętek, M.; Matkowski, R.; Nowak, D. The influence of tumor microenvironment on immune escape of melanoma. Int. J. Mol. Sci. 2020, 21, 8359. [Google Scholar] [CrossRef] [PubMed]
- Kwiatkowska, D.; Kluska, P.; Reich, A. Beyond PD-1 Immunotherapy in malignant melanoma. Dermatol. Ther. 2019, 9, 243–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Del Rivero, J.; Kohn, E.C. PARP Inhibitors: The cornerstone of DNA repair-targeted therapies. Oncology 2017, 31, 265–273. [Google Scholar] [PubMed]
- Domingues, B.; Lopes, J.M.; Soares, P.; Pópulo, H. Melanoma treatment in review. Immuno Targets Ther. 2018, 7, 35–49. [Google Scholar] [CrossRef] [Green Version]
- Mangerich, A.; Bürkle, A. How to kill tumor cells with inhibitors of poly(ADP-ribosyl)ation. Int. J. Cancer 2010, 128, 251–265. [Google Scholar] [CrossRef] [Green Version]
- Palazzo, L.; Ahel, I. PARPs in genome stability and signal transduction: Implications for cancer therapy. Biochem. Soc. Trans. 2018, 46, 1681–1695. [Google Scholar] [CrossRef] [Green Version]
- Cohen, M.S.; Chang, P. Insights into the biogenesis, function, and regulation of ADP-ribosylation. Nat. Chem. Biol. 2018, 14, 236–243. [Google Scholar] [CrossRef] [Green Version]
- Ray, C.A.; Nussenzweig, A. The multifaceted roles of PARP1 in DNA repair and chromatin remodelling. Nat. Rev. Mol. Cell Biol. 2017, 18, 610–621. [Google Scholar]
- Chung, H.T.; Joe, Y. Antagonistic crosstalk between SIRT1, PARP-1, and -2 in the regulation of chronic in-flammation associated with aging and metabolic diseases. Integr. Med. Res. 2014, 3, 198–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galluzzi, L.; Vitale, I.; Aaronson, S.A.; Abrams, J.M.; Adam, D.; Agostinis, P.; Turk, B.; Alnemri, E.S.; Altucci, L.; Amelio, I.; et al. Molecular mechanisms of cell death: Recommendations of the No-menclature committee on cell death 2018. Cell Death. Differ. 2018, 25, 486–541. [Google Scholar] [CrossRef] [PubMed]
- Hassa, P.O.; Hottiger, M.O. The functional role of poly(ADP-ribose)polymerase 1 as novel coactivator of NF-kappaB in inflammatory disorders. Cell Mol. Life Sci. 2002, 59, 1534–1553. [Google Scholar] [CrossRef] [PubMed]
- Krishnakumar, R.; Kraus, W.L. The PARP side of the nucleus: Molecular actions, physiological outcomes, and clinical targets. Mol. Cell 2010, 39, 8–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCann, K.E.; Hurvitz, S.A. Advances in the use of PARP inhibitor therapy for breast cancer. Drugs Context 2018, 7, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Vargas, J.M.; Oliver-Pozo, F.J.; Dantzer, F. PARP1 and Poly(ADP-ribosyl)ation signaling during Au-tophagy in response to nutrient deprivation. Oxid. Med. Cell Longev. 2019, 2019, 2641712. [Google Scholar] [CrossRef] [PubMed]
- Schiewer, M.J.; Knudsen, K.E. Transcriptional roles of PARP1 in cancer. Mol. Cancer Res. 2014, 12, 1069–1080. [Google Scholar] [CrossRef] [Green Version]
- Swindall, A.F.; Stanley, J.A.; Yang, E.S. PARP-1: Friend or Foe of DNA damage and repair in tumorigenesis? Cancers 2013, 5, 943–958. [Google Scholar] [CrossRef] [Green Version]
- Toma, M.; Skorski, T.; Śliwiński, T. Synthetic lethality as a functional tool in basic research and in anticancer therapy. Postępy Higieny Medycyny Doświadczalnej 2014, 68, 1091–1103. [Google Scholar] [CrossRef]
- Csonka, T.; Murnyák, B.; Szepesi, R.; Kurucz, A.; Klekner, Á.; Hortobágyi, T. Poly(ADP-ribose) polymerase-1 (PARP1) and p53 labelling index cor-relates with tumour grade in meningiomas. Folia Neuropathol. 2014, 52, 111–120. [Google Scholar] [CrossRef] [Green Version]
- Murnyák, B.; Kouhsari, M.C.; Hershkovitch, R.; Kálmán, B.; Marko-Varga, G.; Klekner, Á.; Hortobágyi, T. PARP1 expression and its correlation with survival is tu-mour molecular subtype dependent in glioblastoma. Oncotarget 2017, 8, 46348–46362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Idogawa, M.; Yamada, T.; Honda, K.; Sato, S.; Imai, K.; Hirohashi, S. Poly(ADP-ribose) polymerase-1 is a component of the oncogenic T-cell factor-4/beta-catenin complex. Gastroenterology 2005, 128, 1919–1936. [Google Scholar] [CrossRef] [PubMed]
- Schiewer, M.J.; Mandigo, A.C.; Gordon, N.; Huang, F.; Gaur, S.; De Leeuw, R.; Zhao, S.G.; Evans, J.; Han, S.; Parsons, T.; et al. PARP-1 regulates DNA repair factor availability. EMBO Mol. Med. 2018, 10, e8816. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Li, Y.; Lv, S.; Zhang, C.; Tian, Y. PARP-1 may be involved in angiogenesis in epithelial ovarian cancer. Oncol. Lett. 2016, 12, 4561–4567. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, M.I.; Peralta-Leal, A.; O’Valle, F.; Rodriguez-Vargas, J.M.; Gonzalez-Flores, A.; Majuelos-Melguizo, J.; López, L.; Serrano, S.; De Herreros, A.G.; Rodríguez-Manzaneque, J.C.; et al. PARP-1 Regulates metastatic melanoma through modulation of vimentin-induced malignant transformation. PLoS Genet. 2013, 9, e1003531. [Google Scholar] [CrossRef] [Green Version]
- Chow, J.P.; Man, W.Y.; Mao, M.; Chen, H.; Cheung, F.; Nicholls, J.M.; Tsao, S.W.; Lung, M.L.; Poon, R.Y.C. PARP1 Is overexpressed in nasopharyngeal carcinoma and its inhibition enhances radiotherapy. Mol. Cancer Ther. 2013, 12, 2517–2528. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Liu, Y.; Pang, Y.; Pacak, K.; Yang, C. Double-barreled gun: Combination of PARP inhibitor with conventional chemo-therapy. Pharmacol. Ther. 2018, 188, 168–175. [Google Scholar] [CrossRef]
- Pulliam, N.; Tang, J.; Wang, W.; Fang, F.; Sood, R.; O’Hagan, H.M.; Miller, K.D.; Clarke, R.B.; Nephew, K.P. Poly-ADP-ribosylation of estrogen receptor-alpha by PARP1 mediates antiestrogen resistance in human breast cancer cells. Cancers 2019, 11, 43. [Google Scholar] [CrossRef] [Green Version]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utiliz-ing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Towbin, H.; Staehelin, T.; Gordon, J. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocel-lulose sheets: Procedure and some applications. Proc. Natl. Acad. Sci. USA 1979, 76, 4350–4354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kupczyk, P.; Reich, A.; Gajda, M.; Hołysz, M.; Wysokińska, E.; Paprocka, M.; Szepietowski, J.C.; Jagodziński, P.P.; Ziółkowski, P.; Szepietowski, J.C.; et al. UCHL1/PGP 9.5 dynamic in neuro-immune-cutaneous milieu: Focus-ing on axonal nerve terminals and epidermal keratinocytes in psoriatic itch. Biomed. Res. Int. 2018, 2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Donizy, P.; Wu, C.L.; Mull, J.; Fujimoto, M.; Chłopik, A.; Peng, Y.; Shalin, S.C.; Selim, M.A.; Puig, S.; Figueras, M.T.F.; et al. Up-regulation of PARP1 expression significantly correlated with poor sur-vival in mucosal melanomas. Cells 2020, 9, 1135. [Google Scholar]
- Kassambara, A.; Kosinski, M.; Biecek, P. Survminer: Drawing Survival Curves Using “ggplot”. Available online: https://cran.r-project.org/package=survminer (accessed on 12 March 2020).
- Simiczyjew, A.; Pietraszek-Gremplewicz, K.; Dratkiewicz, E.; Podgórska, M.; Matkowski, R.; Ziętek, M.; Nowak, D. Combination of selected MET and EGFR inhibitors decreases melanoma cells’ invasive abilities. Front. Pharmacol. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Makowiecka, A.; Simiczyjew, A.; Nowak, D.; Mazur, A.J. Varying effects of EGF, HGF and TGFbeta on formation of invadopodia and invasiveness of melanoma cell lines of different origin. Eur. J. Histochem. 2016, 60, 2728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Staibano, S.; Pepe, S.; Muzio, L.L.; Somma, P.; Mascolo, M.; Argenziano, G.; Scalvenzi, M.; Salvatore, G.; Fabbrocini, G.; Molea, G.; et al. Poly (adenosine diphosphate-ribose) polymerase 1 expression in malignant melanomas from photoexposed areas of the head and neck region. Hum. Pathol. 2005, 36, 724–731. [Google Scholar] [CrossRef]
- Mactier, S.; Kaufman, K.L.; Wang, P.; Crossett, B.; Pupo, G.M.; Kohnke, P.L.; Thompson, J.F.; Scolyer, R.A.; Yang, J.Y.; Mann, G.J.; et al. Protein signatures correspond to survival outcomes of AJCC stage III melanoma patients. Pigment. Cell Melanoma Res. 2014, 27, 1106–1116. [Google Scholar] [CrossRef]
- Davies, J.R.; Jewell, R.; Affleck, P.; Anic, G.M.; Randerson-Moor, J.; Ozola, A.; Egan, K.M.; Elliott, F.; García-Casado, Z.; Hansson, J.; et al. Inherited variation in the PARP1 gene and survival from melanoma. Int. J. Cancer 2014, 135, 1625–1633. [Google Scholar] [CrossRef]
- Amiri, K.I.; Ha, H.C.; Smulson, M.E.; Richmond, A. Differential regulation of CXC ligand 1 transcription in melanoma cell lines by poly (ADP-ribose) polymerase-1. Oncogene 2006, 25, 7714–7722. [Google Scholar] [CrossRef] [Green Version]
- Nirodi, C.; Nagdas, S.; Gygi, S.P.; Olson, G.; Aebersold, R.; Richmond, A. A role for Poly(ADP-ribose) polymerase in the transcriptional regulation of the melanoma growth stimulatory activity (CXCL1) gene expression. J. Biol. Chem. 2001, 276, 9366–9374. [Google Scholar] [CrossRef] [Green Version]
- Moreno-Lama, L.; Galindo-Campos, M.A.; Martínez, C.M.; Comerma, L.; Vazquez, I.; Vernet-Tomas, M.; Ampurdanés, C.; Lutfi, N.; Martin-Caballero, J.; Dantzer, F.; et al. Coordinated signals from PARP-1 and PARP-2 are required to establish a proper T cell immune response to breast tumors in mice. Oncogene 2020, 39, 2835–2843. [Google Scholar] [CrossRef] [PubMed]
- Maniotis, A.J.; Folberg, R.; Hess, A.; Seftor, E.A.; Gardner, L.M.; Pe’Er, J.; Trent, J.M.; Meltzer, P.S.; Hendrix, M.J.C. Vascular channel formation by human melanoma cells In Vivo and In Vitro: Vasculogenic mimicry. Am. J. Pathol. 1999, 155, 739–752. [Google Scholar] [CrossRef] [Green Version]
- Fratangelo, F.; Camerlingo, R.; Carriero, M.V.; Pirozzi, G.; Palmieri, G.; Gentilcore, G.; Ragone, C.; Minopoli, M.; Ascierto, P.A.; Motti, M.L.; et al. Effect of ABT-888 on the apoptosis, motility and invasive-ness of BRAFi-resistant melanoma cells. Int. J. Oncol. 2018, 53, 1149–1159. [Google Scholar] [PubMed] [Green Version]
- Raineri, A.; Fasoli, S.; Campagnari, R.; Gotte, G.; Menegazzi, M. Onconase restores cytotoxicity in dabrafenib-resistant A375 Hu-man melanoma cells and affects cell migration, invasion and colony formation capability. Int. J. Mol. Sci. 2019, 20, 5890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raineri, A.; Prodomini, S.; Fasoli, S.; Gotte, G.; Menegazzi, M. Influence of onconase in the therapeutic potential of PARP inhibitors in A375 malignant melanoma cells. Biochem. Pharmacol. 2019, 167, 173–181. [Google Scholar] [CrossRef] [PubMed]
- De Koning, L.; Decaudin, D.; el Botty, R.; Nicolas, A.; Carita, G.; Schuller, M.; Ouine, B.; Cartier, A.; Naguez, A.; Fleury, J.; et al. PARP inhibition increases the response to chemotherapy in uveal mela-noma. Cancers 2019, 11, 751. [Google Scholar] [CrossRef] [Green Version]
- Simard, V.M.; St-Laurent, J.F.; Vigneault, F.; Gaudreault, M.; Dargis, N.; Guérin, M.-C.; Leclerc, S.; Morcos, M.; Black, D.; Molgat, Y.; et al. Altered expression of the poly(ADP-ribosyl)ation en-zymes in uveal melanoma and regulation of PARG gene expression by the transcription factor ERM. Investig. Ophthalmol. Vis. Sci. 2012, 53, 6219–6231. [Google Scholar] [CrossRef] [Green Version]
- Kashima, L.; Idogawa, M.; Mita, H.; Shitashige, M.; Yamada, T.; Ogi, K.; Suzuki, H.; Toyota, M.; Ariga, H.; Sasaki, Y.; et al. CHFR protein regulates mitotic checkpoint by targeting PARP-1 pro-tein for ubiquitination and degradation. J. Biol. Chem. 2012, 287, 12975–12984. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Li, C.; Jin, J.; Wang, J.; Huang, J.; Ma, Z.; Huang, H.; He, X.; Zhou, Y.; Xu, Y.; et al. High PARP-1 expression predicts poor survival in acute myeloid leukemia and PARP-1 inhibitor and SAHA-bendamustine hybrid inhibitor combination treatment synergistically enhances an-ti-tumor effects. Ebiomedicine 2018, 38, 47–56. [Google Scholar] [CrossRef]
- Robu, M.; Shah, R.G.; Petitclerc, N.; Brind’Amour, J.; Kandan-Kulangara, F.; Shah, G.M. Role of poly(ADP-ribose) polymerase-1 in the removal of UV-induced DNA lesions by nucleotide excision repair. Proc. Natl. Acad. Sci. USA 2013, 110, 1658–1663. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Forward (F) and Reverse (R) Primers Sequences | UPL Probes | Gene Accession Number |
|---|---|---|---|
| Target Gene | |||
| PARP1 | F: TCTTTGATGTGGAAAGTATGAAGAA R: GGCATCTTCTGAAGGTCGAT | #22 | NM_001618.3 |
| Housekeeping Genes (HKGs): | |||
| POLR2A | F: TCCGTATTCGCATCATGAAC R: TCATCCATCTTGTCCACCAC | #69 | NM_000937 |
| PPIA | F: TTCATCTGCACTGCCAAGAC R: CACTTTGCCAAACACCACAT | #158 | NM_021130 |
| GAPDH | F: TGGTATCGTGGAAGGACTCA R: GCAGGGATGATGTTCTGGAG | #158 | NM_002046 |
| Clinical Parameters | Parp1 Expression | ||
|---|---|---|---|
| Low (H-Score ≤ 280) (n = 98) | High (H-Score > 280) (n = 30) | p Value | |
| Age (18–86 years) a | 61 (50–73) | 69 (58–74) | 0.090 |
| Gender b | 0.41 | ||
| Female | 52 (53%) | 13 (43%) | |
| Male | 46 (47%) | 17 (57%) | |
| Primary tumor location c | 0.015 | ||
| Head/neck | 4 (4%) | 7 (23%) | |
| Extremities | 44 (45%) | 10 (33%) | |
| Trunk | 46 (47%) | 13 (43%) | |
| Hand/foot | 4 (4%) | 0 (0%) | |
| Primary tumor (pT) a | 0.16 | ||
| pT1 | 27 (28%) | 3 (10%) | |
| pT2 | 15 (15%) | 5 (17%) | |
| pT3 | 27 (28%) | 8 (27%) | |
| pT4 | 29 (30%) | 14 (47%) | |
| Sentinel lymph node biopsy status (SNLB) b | 0.099 | ||
| No metastases (SNLB-) | 24 (54%) | 10 (83%) | |
| Metastases present (SNLB+) | 20 (46%) | 2 (17%) | |
| Regional lymph nodes status (pN) b | 0.63 | ||
| Metastases absent (pN-) | 72 (73%) | 24 (80%) | |
| Metastases present (pN+) | 26 (27%) | 6 (20%) | |
| Distant metastases (pM) b | 1.0 | ||
| No metastases (pM-) | 88 (90%) | 27 (90%) | |
| Metastases present (pM+) | 10 (10%) | 3 (10%) | |
| Recurrence b | 0.083 | ||
| No | 67 (68%) | 15 (50%) | |
| Yes | 31 (32%) | 15 (50%) | |
| Histopathological Parameters | PARP1 Expression | ||
|---|---|---|---|
| Low (H-Score ≤ 280) (n = 98) | High (H-Score > 280) (n = 30) | p Value | |
| Breslow thickness a | 0.16 | ||
| ≤1 mm | 27 (28%) | 3 (10%) | |
| 1.01–2.00 mm | 15 (15%) | 5 (17%) | |
| 2.01–4.00 mm | 27 (28%) | 8 (27%) | |
| >4 mm | 29 (30%) | 14 (47%) | |
| Clark level a | 0.37 | ||
| I | 0 (0%) | 0 (0%) | |
| II | 32 (33%) | 5 (17%) | |
| III | 30 (31%) | 12 (40%) | |
| IV | 26 (27%) | 10 (33%) | |
| V | 10 (10%) | 3 (10%) | |
| Histological type b | 0.30 | ||
| Superficial spreading melanoma | 47 (48%) | 11 (37%) | |
| Nodular melanoma | 47 (48%) | 19 (63%) | |
| Acral lentiginous melanoma | 4 (4%) | 0 (0%) | |
| Mitotic rate a | 0.0010 | ||
| 0 | 33 (34%) | 1 (4%) | |
| 1–3 | 19 (20%) | 7 (23%) | |
| ≥4 | 45 (46%) | 22 (73%) | |
| Ulceration c | 0.036 | ||
| No | 64 (65%) | 13 (43%) | |
| Yes | 34 (35%) | 17 (57%) | |
| Lymphangioinvasion c | 1.0 | ||
| No | 92 (94%) | 29 (97%) | |
| Yes | 6 (6%) | 1 (3%) | |
| Tumor-infiltrating lymphocytes c | 0.60 | ||
| No | 6 (6%) | 1 (3%) | |
| Non-brisk | 58 (59%) | 21 (70%) | |
| Brisk | 34 (35%) | 8 (27%) | |
| Microsatellitosis c | 0.084 | ||
| No | 96 (98%) | 27 (90%) | |
| Yes | 2 (2%) | 3 (10%) | |
| Regression c | 0.14 | ||
| No | 95 (97%) | 27 (93%) | |
| Yes | 3 (3%) | 2 (7%) | |
| Clinical Parameters | PARP1 Expression | ||
|---|---|---|---|
| Low (H-Score ≤ 280) (n = 72) | High (H-Score > 280) (n = 24) | p Value | |
| Age (24–86) a | 61 (50–73) | 70 (60–75) | 0.055 |
| Gender b | 1.0 | ||
| Female | 37 (51%) | 12 (50%) | |
| Male | 35 (49%) | 12 (50%) | |
| Primary tumor location c | 0.016 | ||
| Head/neck | 3 (4%) | 6 (25%) | |
| Extremities | 33 (46%) | 8 (33%) | |
| Trunk | 36 (50%) | 10 (42%) | |
| Hand/foot | 0 (0%) | 0 (0%) | |
| Primary tumor (pT) a | 0.0030 | ||
| pT1 | 26 (36%) | 2 (8%) | |
| pT2 | 15 (21%) | 5 (21%) | |
| pT3 | 20 (28%) | 5 (21%) | |
| pT4 | 11 (15%) | 12 (50%) | |
| Distant metastases (pM) b | 1.0 | ||
| No metastases (pM-) | 68 (94%) | 23 (96%) | |
| Metastases present (pM+) | 4 (6%) | 1 (4%) | |
| Recurrence b | 0.0061 | ||
| No | 60 (87%) | 13 (54%) | |
| Yes | 12 (17%) | 11 (46%) | |
| Histopathological Parameters | PARP1 Expression | ||
|---|---|---|---|
| Low (H-Score ≤ 280) (n = 72) | High (H-Score > 280) (n = 24) | p Value | |
| Breslow thickness a | 0.0030 | ||
| ≤1 mm | 26 (36%) | 2 (8%) | |
| 1.01–2.00 mm | 15 (21%) | 5 (21%) | |
| 2.01–4.00 mm | 20 (28%) | 5 (21%) | |
| >4 mm | 11 (15%) | 12 (50%) | |
| Clark level a | 0.13 | ||
| I | 0 (0%) | 0 (0%) | |
| II | 30 (42%) | 4 (17%) | |
| III | 22 (31%) | 10 (42%) | |
| IV | 16 (22%) | 8 (33%) | |
| V | 4 (6%) | 2 (8%) | |
| Histological type b | 0.16 | ||
| Superficial spreading melanoma | 43 (60%) | 10 (42%) | |
| Nodular melanoma | 29 (40%) | 14 (58%) | |
| Acral lentiginous melanoma | 0 (0%) | 0 (0%) | |
| Mitotic rate a | 0.00071 | ||
| 0 | 30 (42%) | 1 (4%) | |
| 1–3 | 17 (24%) | 7 (29%) | |
| ≥4 | 25 (35%) | 16 (67%) | |
| Ulceration c | 0.023 | ||
| No | 53 (74%) | 11 (46%) | |
| Yes | 19 (26%) | 13 (54%) | |
| Lymphangioinvasion c | 1.0 | ||
| No | 70 (97%) | 24 (100%) | |
| Yes | 2 (3%) | 0 (0%) | |
| Tumor-infiltrating lymphocytes c | 0.26 | ||
| No | 4 (6%) | 1 (4%) | |
| Non-brisk | 37 (51%) | 17 (71%) | |
| Brisk | 31 (43%) | 6 (25%) | |
| Microsatellitosis c | 1.0 | ||
| No | 72 (100%) | 24 (100%) | |
| Yes | 0 (0%) | 0 (0%) | |
| Regression c | 0.15 | ||
| No | 71 (99%) | 22 (96%) | |
| Yes | 1 (1%) | 1 (4%) | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kupczyk, P.; Simiczyjew, A.; Marczuk, J.; Dratkiewicz, E.; Beberok, A.; Rok, J.; Pieniazek, M.; Biecek, P.; Nevozhay, D.; Slowikowski, B.; et al. PARP1 as a Marker of an Aggressive Clinical Phenotype in Cutaneous Melanoma—A Clinical and an In Vitro Study. Cells 2021, 10, 286. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10020286
Kupczyk P, Simiczyjew A, Marczuk J, Dratkiewicz E, Beberok A, Rok J, Pieniazek M, Biecek P, Nevozhay D, Slowikowski B, et al. PARP1 as a Marker of an Aggressive Clinical Phenotype in Cutaneous Melanoma—A Clinical and an In Vitro Study. Cells. 2021; 10(2):286. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10020286
Chicago/Turabian StyleKupczyk, Piotr, Aleksandra Simiczyjew, Jakub Marczuk, Ewelina Dratkiewicz, Artur Beberok, Jakub Rok, Malgorzata Pieniazek, Przemyslaw Biecek, Dmitry Nevozhay, Bartosz Slowikowski, and et al. 2021. "PARP1 as a Marker of an Aggressive Clinical Phenotype in Cutaneous Melanoma—A Clinical and an In Vitro Study" Cells 10, no. 2: 286. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10020286