
CellMAPtracer: A User-Friendly Tracking Tool for Long-Term Migratory and Proliferating Cells Associated with FUCCI Systems

 ,
,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Cell Culture, Random Migration, and Live Cell Imaging of BT549 Cells
2.2. Two-Channel FUCCI Time-Lapse Data
2.3. Three-Channel FUCCI Time-Lapse Data
2.4. CellMAPtracer
2.4.1. Availability and Installation
- (A)
- Standalone executable programThis can be accessed at https://github.com/ocbe-uio/CellMAPtracer/releases/tag/v1.1 (accessed on 20 February 2021). Three assets of CellMAPtracer for Windows, Linux, and macOS versions can be found and downloaded. After downloading the version compatible with their operating system, users should decompress the file and follow the instructions in the corresponding “readme.txt”.
- (B)
- MATLAB AppTo be able to run CellMAPtracer App within the MATLAB environment, users should follow three simple steps: (1) download the “App” folder from the CellMAPtracer repository: https://github.com/ocbe-uio/CellMAPtracer (accessed on 20 February 2021); (2) in MATLAB, go to the APPS tab and click “Install App”, find the “CellMAPtracer.mlappinstall” file, and then install it; (3) open CellMAPtracer App from the Application list in MATLAB.
- (C)
- From the source MATLAB codeTo be able to run CellMAPtracer code, users should clone the CellMAPtracer repository from https://github.com/ocbe-uio/CellMAPtracer (accessed on 20 February 2021) and then run “CellMAPtracer_Main.m” after opening a project in MATLAB.
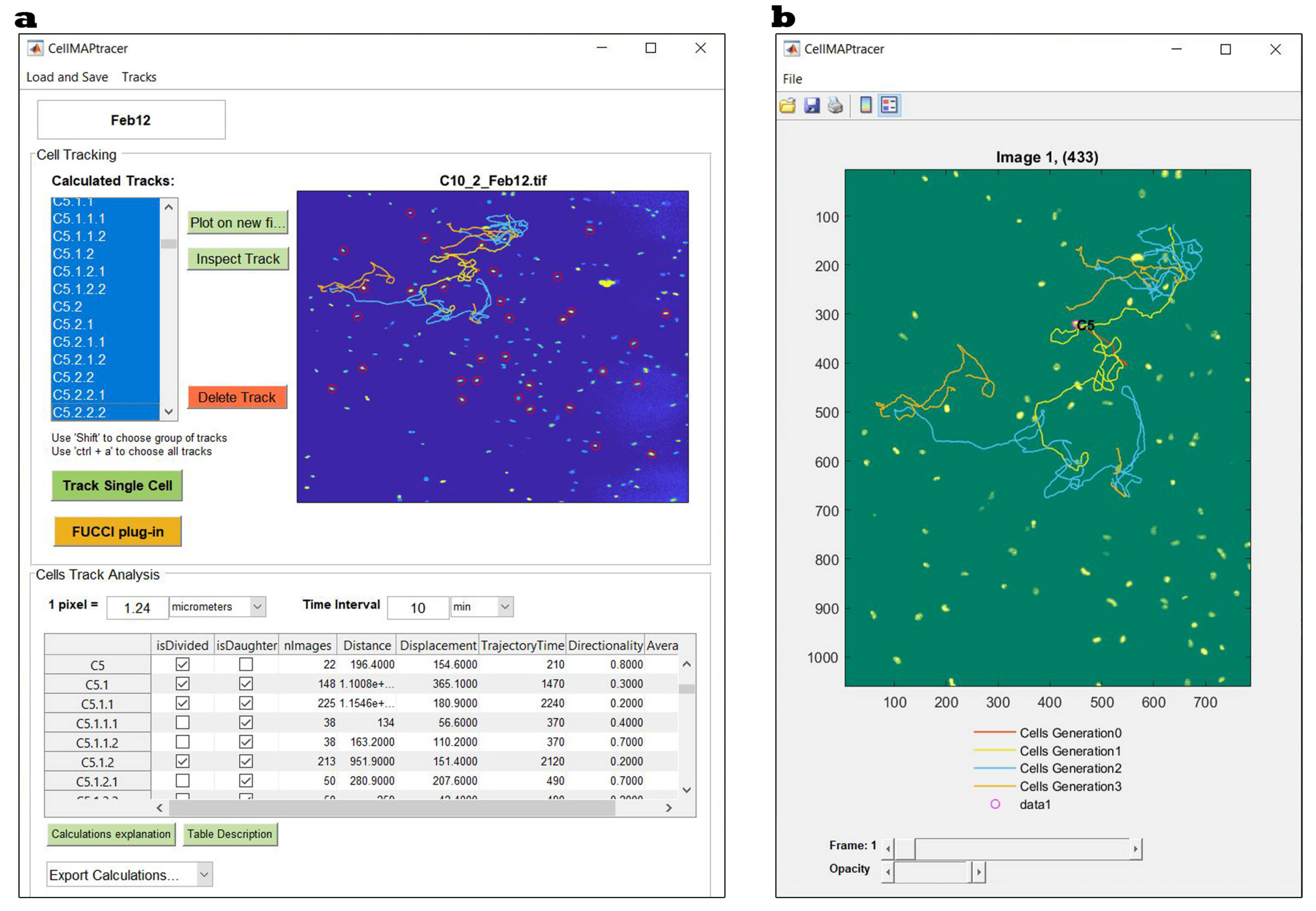
2.4.2. CellMAPtracer Description and Workflow
Tracking Single Cells
FUCCI Plug-In
- (A)
- Two channel systemThe RGB multi-TIFF stacks should be formatted with the red channel corresponding to Cdt1 and the green channel corresponding to Geminin (Figure 2a). Cdt1 and Geminin have opposing effects on DNA replication and show inverse oscillation patterns during the cell cycle. Cdt1 protein (red signal) peaks in G1 phase and then declines in S phase before becoming high again at the M/G1 transition [34]. In contrast, Geminin (green signal) is destabilized during G1 phase but accumulates during S and G2 phases [35]. The cell-cycle detection approach is based on three parameters: the minimum signal (MS), the minimum green signal in S phase (MGSS), and the minimum red signal in S phase (MRSS). The default values for MS, MGSS, and MRSS are set to 0.1 with the possibility of being adjusted by users. Figure 2b demonstrates the mechanism for determining the cell-cycle phase in a two-channel system.
- (B)
- Three-channel systemThe RGB multi-TIFF stacks should be formatted with the red channel corresponding to PCNA, the green channel corresponding to Geminin, and the blue channel corresponding to PIP (Figure 2a). PIP-FUCCI is a degradation-based biosensor of cell-cycle phases [28]. It includes two fluorescently tagged sensors—the PIP degron (Cdt1-PIP fragment) and Geminin1-110 fragment. PIP-mVenus is a direct indicator of replication as its dynamics is physically coupled to the PCNA presence at the replication forks. On the other hand, Gem1-110 (fused to mCherry fluorescent protein), as an antigen presenting cell (APC)-Cdh1 target, is an indirect sensor of the G1/S transition and is present in S and G2 phases of the cell cycle. The approach to detecting the cell-cycle phases is based on three parameters: the maximum green-blue ratio in G1 phase (MGBr-G1), the minimum green-blue ratio in G2 phase (MGBr-G2), and the minimum blue signal in G2 phase (MBS-G2). The default values for MGBr-G1, MGBr-G2, and MBS-G2 are set to 0.5, 0.7, and 0.1, respectively, with the possibility to adjust the values by the user. Figure 2c demonstrates the mechanism for determining the cell-cycle phase in a three-channel system.
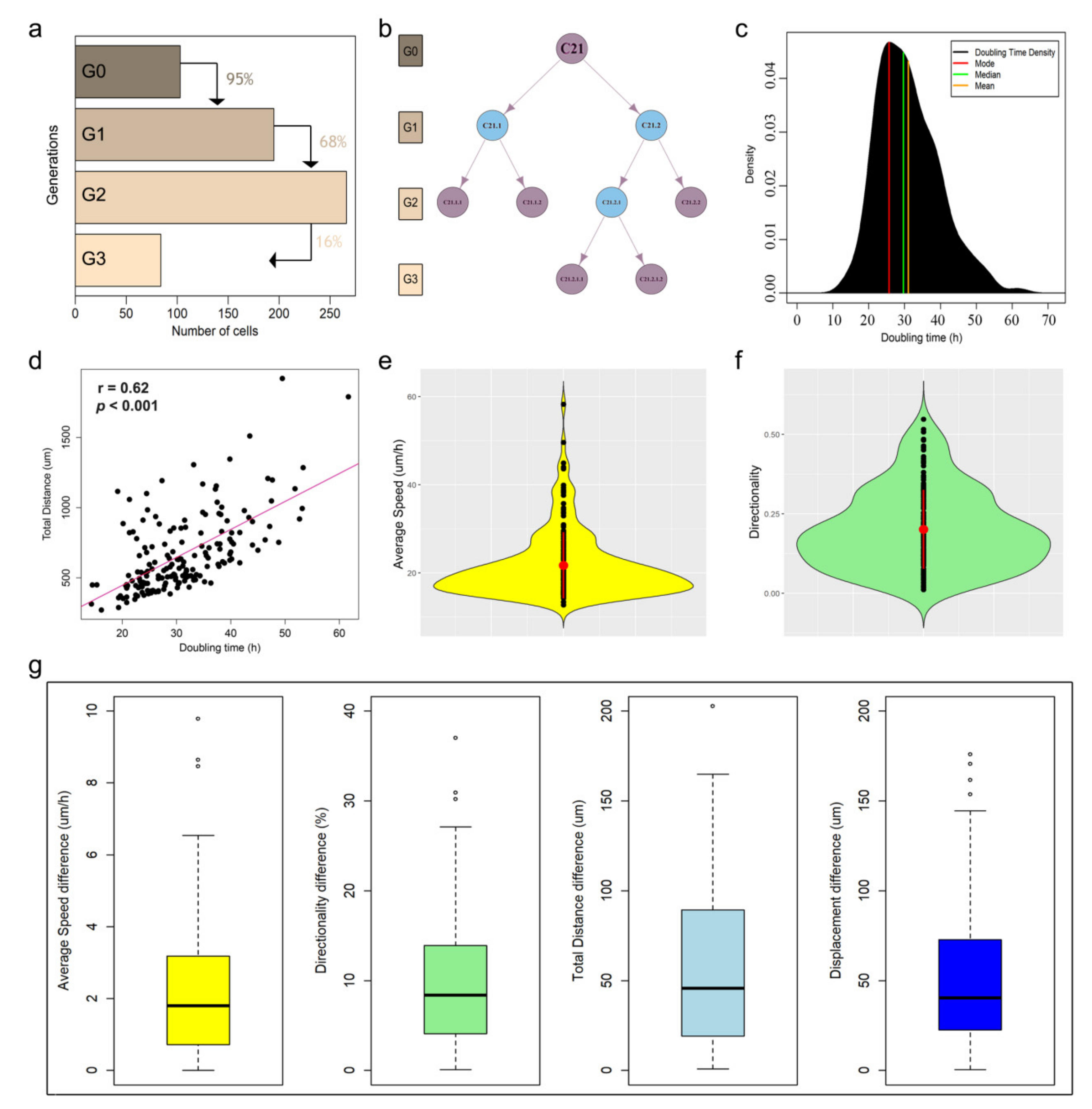
2.5. Trajectory Analysis
3. Results
3.1. Tracking and Analyzing Human Breast Cancer Cells
3.2. Tracking and Analyzing Human Epithelial Cells Expressing FUCCI
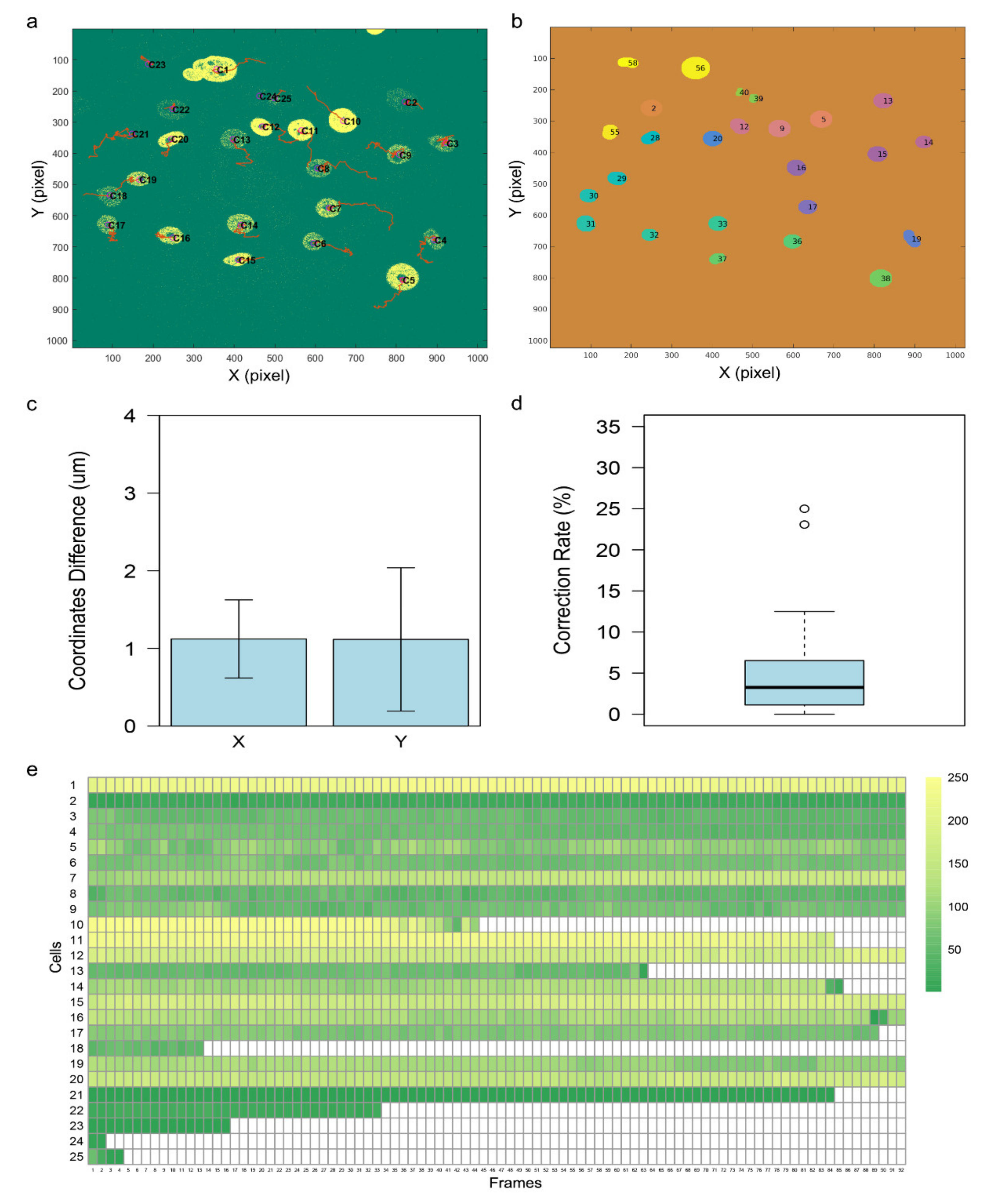
3.3. Evaluation of the Tracking Accuracy
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Friedl, P.; Gilmour, D. Collective cell migration in morphogenesis, regeneration and cancer. Nat. Rev. Mol. Cell Biol. 2009, 10, 445–457. [Google Scholar] [CrossRef] [PubMed]
- Masuzzo, P.; Van Troys, M.; Ampe, C.; Martens, L. Taking aim at moving targets in computational cell migration. Trends Cell Biol. 2016, 26, 88–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cordelières, F.P.; Petit, V.; Kumasaka, M.; Debeir, O.; Letort, V.; Gallagher, S.J.; Larue, L. Automated cell tracking and analysis in phase-contrast videos (iTrack4U): Development of Java software based on combined mean-shift processes. PLoS ONE 2013, 8, e81266. [Google Scholar] [CrossRef] [PubMed]
- Gorelik, R.; Gautreau, A. Quantitative and unbiased analysis of directional persistence in cell migration. Nat. Protoc. 2014, 9, 1931. [Google Scholar] [CrossRef]
- Mukherjee, D.P.; Ray, N.; Acton, S.T. Level set analysis for leukocyte detection and tracking. IEEE Trans. Image Process. 2004, 13, 562–572. [Google Scholar] [CrossRef] [PubMed]
- Emami, N.; Sedaei, Z.; Ferdousi, R. Computerized cell tracking: Current methods, tools and challenges. Vis. Inform. 2020. [Google Scholar] [CrossRef]
- Moen, E.; Bannon, D.; Kudo, T.; Graf, W.; Covert, M.; Van Valen, D. Deep learning for cellular image analysis. Nat. Methods 2019, 16, 1233–1246. [Google Scholar] [CrossRef] [PubMed]
- Boukari, F.; Makrogiannis, S. Automated cell tracking using motion prediction-based matching and event handling. IEEE/Acm Trans. Comput. Biol. Bioinform. 2018, 17, 959–971. [Google Scholar] [CrossRef] [PubMed]
- Sacan, A.; Ferhatosmanoglu, H.; Coskun, H. CellTrack: An open-source software for cell tracking and motility analysis. Bioinformatics 2008, 24, 1647–1649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, H.; Nelson, G.; Kennedy, S.; Nelson, D.; Johnson, J.; Spiller, D.; White, M.R.; Kell, D.B. Automatic tracking of biological cells and compartments using particle filters and active contours. Chemom. Intell. Lab. Syst. 2006, 82, 276–282. [Google Scholar] [CrossRef]
- Van Valen, D.A.; Kudo, T.; Lane, K.M.; Macklin, D.N.; Quach, N.T.; DeFelice, M.M.; Maayan, I.; Tanouchi, Y.; Ashley, E.A.; Covert, M.W. Deep learning automates the quantitative analysis of individual cells in live-cell imaging experiments. PLoS Comput. Biol. 2016, 12, e1005177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooper, S.; Barr, A.R.; Glen, R.; Bakal, C. NucliTrack: An integrated nuclei tracking application. Bioinformatics 2017, 33, 3320–3322. [Google Scholar] [CrossRef] [Green Version]
- DuChez, B.J. Automated tracking of cell migration with rapid data analysis. Curr. Protoc. Cell Biol. 2018, 76, 12.12.1–12.12.16. [Google Scholar] [CrossRef]
- Tinevez, J.-Y.; Perry, N.; Schindelin, J.; Hoopes, G.M.; Reynolds, G.D.; Laplantine, E.; Bednarek, S.Y.; Shorte, S.L.; Eliceiri, K.W. TrackMate: An open and extensible platform for single-particle tracking. Methods 2017, 115, 80–90. [Google Scholar] [CrossRef] [PubMed]
- Tsai, H.-F.; Gajda, J.; Sloan, T.F.; Rares, A.; Shen, A.Q. Usiigaci: Instance-aware cell tracking in stain-free phase contrast microscopy enabled by machine learning. SoftwareX 2019, 9, 230–237. [Google Scholar] [CrossRef]
- Fujimoto, K.; Seno, S.; Shigeta, H.; Mashita, T.; Ishii, M.; Matsuda, H. Tracking and Analysis of FUCCI-Labeled Cells Based on Particle Filters and Time-to-Event Analysis. IJBBB 2020. [Google Scholar] [CrossRef] [Green Version]
- Fazeli, E.; Roy, N.H.; Follain, G.; Laine, R.F.; von Chamier, L.; Hänninen, P.E.; Eriksson, J.E.; Tinevez, J.-Y.; Jacquemet, G. Automated cell tracking using StarDist and TrackMate. F1000Research 2020, 9, 1279. [Google Scholar] [CrossRef] [PubMed]
- Moen, E.; Borba, E.; Miller, G.; Schwartz, M.; Bannon, D.; Koe, N.; Camplisson, I.; Kyme, D.; Pavelchek, C.; Price, T. Accurate cell tracking and lineage construction in live-cell imaging experiments with deep learning. bioRxiv 2019. [Google Scholar] [CrossRef]
- Roccio, M.; Schmitter, D.; Knobloch, M.; Okawa, Y.; Sage, D.; Lutolf, M.P. Predicting stem cell fate changes by differential cell cycle progression patterns. Development 2013, 140, 459–470. [Google Scholar] [CrossRef] [Green Version]
- Koh, S.-B.; Mascalchi, P.; Rodriguez, E.; Lin, Y.; Jodrell, D.I.; Richards, F.M.; Lyons, S.K. A quantitative FastFUCCI assay defines cell cycle dynamics at a single-cell level. J. Cell Sci. 2017, 130, 512–520. [Google Scholar] [CrossRef] [Green Version]
- Hilsenbeck, O.; Schwarzfischer, M.; Skylaki, S.; Schauberger, B.; Hoppe, P.S.; Loeffler, D.; Kokkaliaris, K.D.; Hastreiter, S.; Skylaki, E.; Filipczyk, A. Software tools for single-cell tracking and quantification of cellular and molecular properties. Nat. Biotechnol. 2016, 34, 703–706. [Google Scholar] [CrossRef] [PubMed]
- Meijering, E.; Dzyubachyk, O.; Smal, I. Methods for cell and particle tracking. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 2012; Volume 504, pp. 183–200. [Google Scholar]
- Marcus, J.M.; Burke, R.T.; DeSisto, J.A.; Landesman, Y.; Orth, J.D. Longitudinal tracking of single live cancer cells to understand cell cycle effects of the nuclear export inhibitor, selinexor. Sci. Rep. 2015, 5, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Go, Y.-H.; Lee, H.-J.; Kong, H.-J.; Jeong, H.-C.; Lee, D.Y.; Hong, S.-K.; Sung, S.H.; Kwon, O.-S.; Cha, H.-J. Screening of cytotoxic or cytostatic flavonoids with quantitative Fluorescent Ubiquitination-based Cell Cycle Indicator-based cell cycle assay. R. Soc. Open Sci. 2018, 5, 181303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Svoboda, P.; Krizova, E.; Sestakova, S.; Vapenkova, K.; Knejzlik, Z.; Rimpelova, S.; Rayova, D.; Volfova, N.; Krizova, I.; Rumlova, M. Nuclear transport of nicotinamide phosphoribosyltransferase is cell cycle–dependent in mammalian cells, and its inhibition slows cell growth. J. Biol. Chem. 2019, 294, 8676–8689. [Google Scholar] [CrossRef] [PubMed]
- Yano, S.; Tazawa, H.; Kagawa, S.; Fujiwara, T.; Hoffman, R.M. FUCCI Real-Time Cell-Cycle Imaging as a Guide for Designing Improved Cancer Therapy: A Review of Innovative Strategies to Target Quiescent Chemo-Resistant Cancer Cells. Cancers 2020, 12, 2655. [Google Scholar] [CrossRef]
- Sakaue-Sawano, A.; Kurokawa, H.; Morimura, T.; Hanyu, A.; Hama, H.; Osawa, H.; Kashiwagi, S.; Fukami, K.; Miyata, T.; Miyoshi, H. Visualizing spatiotemporal dynamics of multicellular cell-cycle progression. Cell 2008, 132, 487–498. [Google Scholar] [CrossRef] [Green Version]
- Grant, G.D.; Kedziora, K.M.; Limas, J.C.; Cook, J.G.; Purvis, J.E. Accurate delineation of cell cycle phase transitions in living cells with PIP-FUCCI. Cell Cycle 2018, 17, 2496–2516. [Google Scholar] [CrossRef] [Green Version]
- Antos, K.; Ghannoum, S. CellMAPtracer 1.1 (Version v1.1). Zenodo 2020. [Google Scholar] [CrossRef]
- Breitenstein, M.D.; Reichlin, F.; Leibe, B.; Koller-Meier, E.; Van Gool, L. Online multiperson tracking-by-detection from a single, uncalibrated camera. Ieee Trans. Pattern Anal. Mach. Intell. 2010, 33, 1820–1833. [Google Scholar] [CrossRef]
- Meyer, F. Topographic distance and watershed lines. Signal Process. 1994, 38, 113–125. [Google Scholar] [CrossRef]
- Kornilov, A.S.; Safonov, I.V. An overview of watershed algorithm implementations in open source libraries. J. Imaging 2018, 4, 123. [Google Scholar] [CrossRef] [Green Version]
- Ciaparrone, G.; Sánchez, F.L.; Tabik, S.; Troiano, L.; Tagliaferri, R.; Herrera, F. Deep learning in video multi-object tracking: A survey. Neurocomputing 2020, 381, 61–88. [Google Scholar] [CrossRef] [Green Version]
- Nishitani, H.; Taraviras, S.; Lygerou, Z.; Nishimoto, T. The human licensing factor for DNA replication Cdt1 accumulates in G1 and is destabilized after initiation of S-phase. J. Biol. Chem. 2001, 276, 44905–44911. [Google Scholar] [CrossRef] [Green Version]
- Wohlschlegel, J.A.; Dwyer, B.T.; Dhar, S.K.; Cvetic, C.; Walter, J.C.; Dutta, A. Inhibition of eukaryotic DNA replication by geminin binding to Cdt1. Science 2000, 290, 2309–2312. [Google Scholar] [CrossRef]
- RC Team. R: A Language and Environment for Statistical Computing; RC Team: Vienna, Austria, 2013. [Google Scholar]
- Ghannoum S, A.K. CellMAPtracer TIFF files (Version 1.0.0) [Data set]. Zenodo 2020. [Google Scholar] [CrossRef]
- Grant, G.; Kedziora, K.M.; Limas, J.C.; Cook, J.G.; Purvis, J.E. PIP-FUCCI [Data set]. Zenodo 2020. [Google Scholar] [CrossRef]
- Chao, H.X.; Fakhreddin, R.I.; Shimerov, H.K.; Kedziora, K.M.; Kumar, R.J.; Perez, J.; Limas, J.C.; Grant, G.D.; Cook, J.G.; Gupta, G.P. Evidence that the human cell cycle is a series of uncoupled, memoryless phases. Mol. Syst. Biol. 2019, 15, e8604. [Google Scholar] [CrossRef]
- Rabouille, C.; Kondylis, V. Golgi ribbon unlinking: An organelle-based G2/M checkpoint. Cell Cycle 2007, 6, 2723–2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corda, D.; Barretta, M.L.; Cervigni, R.I.; Colanzi, A. Golgi complex fragmentation in G2/M transition: An organelle-based cell-cycle checkpoint. IUBMB Life 2012, 64, 661–670. [Google Scholar] [CrossRef]
- Bisel, B.; Wang, Y.; Wei, J.-H.; Xiang, Y.; Tang, D.; Miron-Mendoza, M.; Yoshimura, S.-i.; Nakamura, N.; Seemann, J. ERK regulates Golgi and centrosome orientation towards the leading edge through GRASP65. J. Cell Biol. 2008, 182, 837–843. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.-H.; Seemann, J. Golgi ribbon disassembly during mitosis, differentiation and disease progression. Curr. Opin. Cell Biol. 2017, 47, 43–51. [Google Scholar] [CrossRef]
- Millarte, V.; Farhan, H. The Golgi in cell migration: Regulation by signal transduction and its implications for cancer cell metastasis. Sci. World J. 2012, 2012, 498278. [Google Scholar] [CrossRef]
- Millarte, V.; Boncompain, G.; Tillmann, K.; Perez, F.; Sztul, E.; Farhan, H. Phospholipase C γ1 regulates early secretory trafficking and cell migration via interaction with p115. Mol. Biol. Cell 2015, 26, 2263–2278. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Huang, C.; Nevatia, R. Learning to associate: Hybridboosted multi-target tracker for crowded scene. In Proceedings of the 2009 IEEE Conference on Computer Vision and Pattern Recognition, Miami, FL, USA, 20–25 June 2009; pp. 2953–2960. [Google Scholar]
- Bártová, E.; Šustáčková, G.; Stixová, L.; Kozubek, S.; Legartová, S.; Foltánková, V. Recruitment of Oct4 protein to UV-damaged chromatin in embryonic stem cells. PLoS ONE 2011, 6, e27281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ulman, V.; Maška, M.; Magnusson, K.E.; Ronneberger, O.; Haubold, C.; Harder, N.; Matula, P.; Matula, P.; Svoboda, D.; Radojevic, M. An objective comparison of cell-tracking algorithms. Nat. Methods 2017, 14, 1141–1152. [Google Scholar] [CrossRef] [PubMed]
- Ghannoum S, A.K. CellMAPtracer-BT549-dense population TIFF file (Version 1.0.0) [Data set]. Zenodo 2020. [Google Scholar] [CrossRef]
- Heiser, L.M.; Sadanandam, A.; Kuo, W.-L.; Benz, S.C.; Goldstein, T.C.; Ng, S.; Gibb, W.J.; Wang, N.J.; Ziyad, S.; Tong, F. Subtype and pathway specific responses to anticancer compounds in breast cancer. Proc. Natl. Acad. Sci. USA 2012, 109, 2724–2729. [Google Scholar] [CrossRef] [Green Version]
- Cowley, G.S.; Weir, B.A.; Vazquez, F.; Tamayo, P.; Scott, J.A.; Rusin, S.; East-Seletsky, A.; Ali, L.D.; Gerath, W.F.; Pantel, S.E. Parallel genome-scale loss of function screens in 216 cancer cell lines for the identification of context-specific genetic dependencies. Sci. Data 2014, 1, 1–12. [Google Scholar]
- Sweeney, K.J.; Swarbrick, A.; Sutherland, R.L.; Musgrove, E.A. Lack of relationship between CDK activity and G 1 cyclin expression in breast cancer cells. Oncogene 1998, 16, 2865–2878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tse, H.T.K.; Weaver, W.M.; Di Carlo, D. Increased asymmetric and multi-daughter cell division in mechanically confined microenvironments. PLoS ONE 2012, 7, e38986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Telentschak, S.; Soliwoda, M.; Nohroudi, K.; Addicks, K.; Klinz, F.-J. Cytokinesis failure and successful multipolar mitoses drive aneuploidy in glioblastoma cells. Oncol. Rep. 2015, 33, 2001–2008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, K.; Takahashi, K. Reduced cell adhesion during mitosis by threonine phosphorylation of β1 integrin. J. Cell. Physiol. 2003, 197, 297–305. [Google Scholar] [CrossRef] [PubMed]
- Lancaster, O.M.; Le Berre, M.; Dimitracopoulos, A.; Bonazzi, D.; Zlotek-Zlotkiewicz, E.; Picone, R.; Duke, T.; Piel, M.; Baum, B. Mitotic rounding alters cell geometry to ensure efficient bipolar spindle formation. Dev. Cell 2013, 25, 270–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacquemet, G.; Ivaska, J. Mitosis-resistant adhesions provide molecular memory to dividing cells. Dev. Cell 2018, 45, 5–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bajar, B.T.; Lam, A.J.; Badiee, R.K.; Oh, Y.-H.; Chu, J.; Zhou, X.X.; Kim, N.; Kim, B.B.; Chung, M.; Yablonovitch, A.L. Fluorescent indicators for simultaneous reporting of all four cell cycle phases. Nat. Methods 2016, 13, 993–996. [Google Scholar] [CrossRef] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghannoum, S.; Antos, K.; Leoncio Netto, W.; Gomes, C.; Köhn-Luque, A.; Farhan, H. CellMAPtracer: A User-Friendly Tracking Tool for Long-Term Migratory and Proliferating Cells Associated with FUCCI Systems. Cells 2021, 10, 469. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10020469
Ghannoum S, Antos K, Leoncio Netto W, Gomes C, Köhn-Luque A, Farhan H. CellMAPtracer: A User-Friendly Tracking Tool for Long-Term Migratory and Proliferating Cells Associated with FUCCI Systems. Cells. 2021; 10(2):469. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10020469
Chicago/Turabian StyleGhannoum, Salim, Kamil Antos, Waldir Leoncio Netto, Cecil Gomes, Alvaro Köhn-Luque, and Hesso Farhan. 2021. "CellMAPtracer: A User-Friendly Tracking Tool for Long-Term Migratory and Proliferating Cells Associated with FUCCI Systems" Cells 10, no. 2: 469. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10020469