
Cytochrome c Oxidase at Full Thrust: Regulation and Biological Consequences to Flying Insects

 , ,
, , 
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
1.1. Flight as an Extreme Metabolic Process
1.2. Insect Flight Is Powered by Mitochondrial Metabolism
1.3. Regulation of Mitochondrial Energy and Redox Metabolism in Flying Insects
2. Cytochrome c Oxidase in Flying Insects
2.1. A Brief Historical Background
2.2. Biological and Physiological Roles of COX to Flying Insects
2.3. Regulation of COX Activity in Flying Insects
3. Phosphorylation as a Critical Mechanism of Post-Translational Modification and Regulation of COX
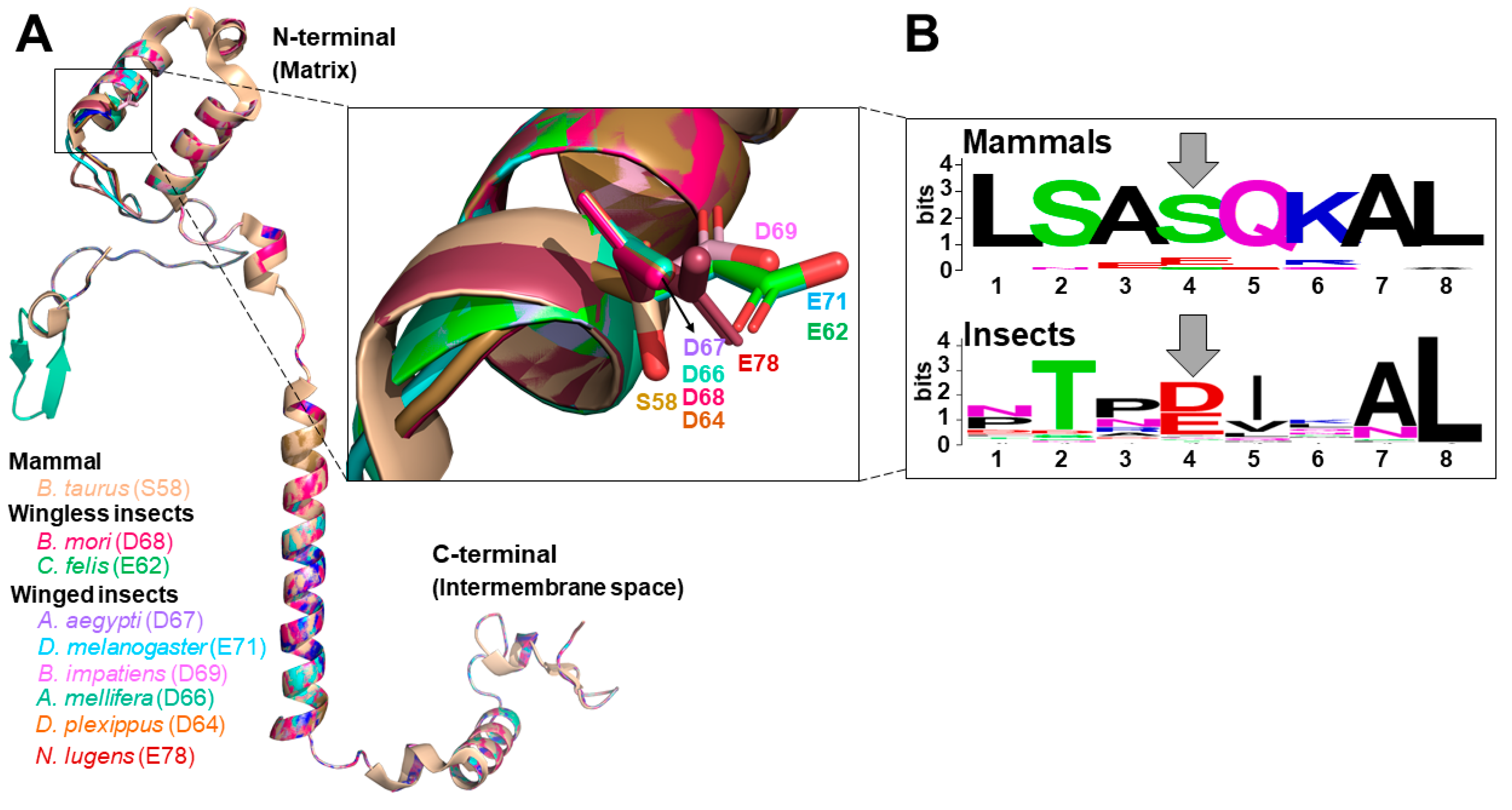
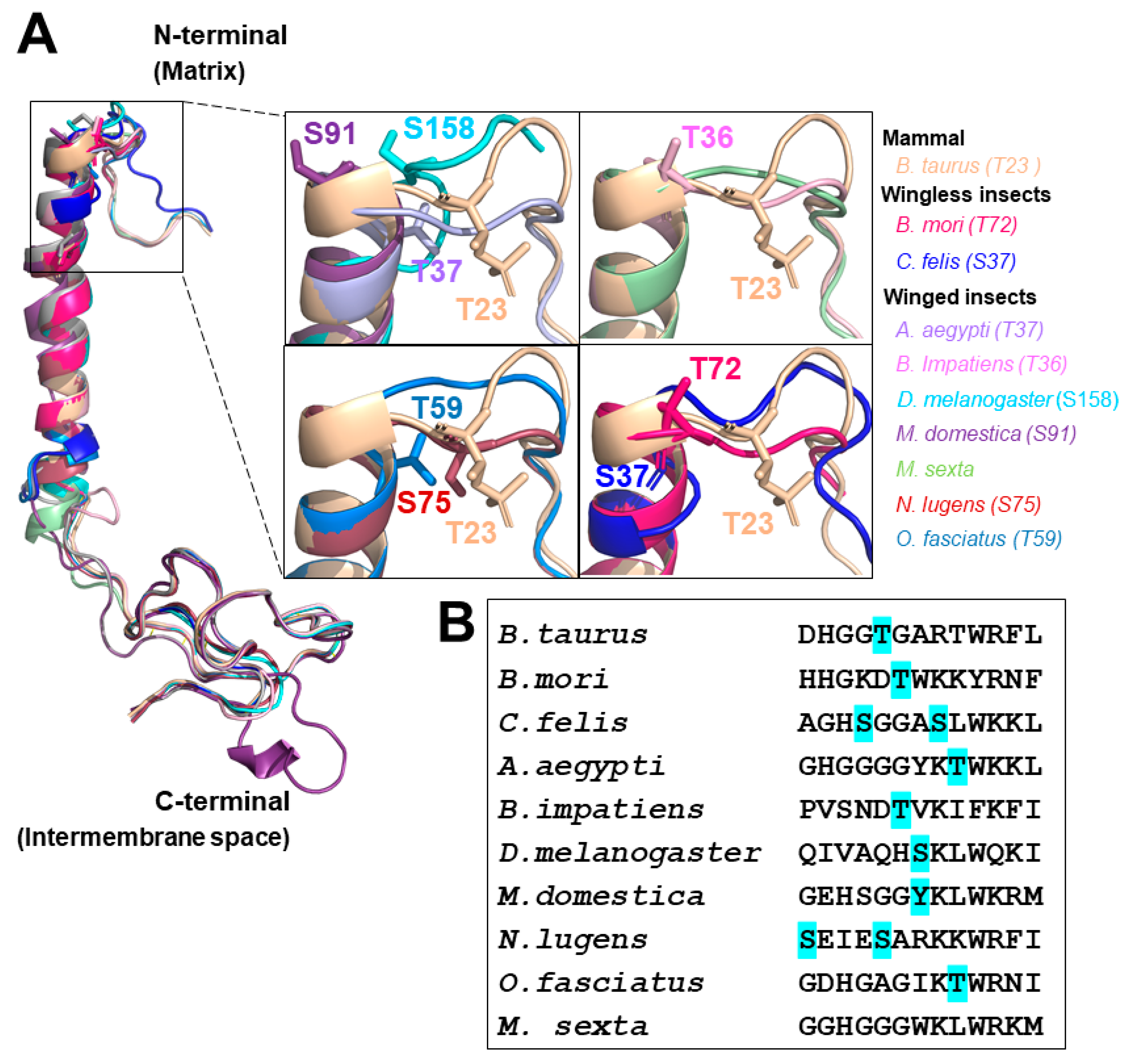
3.1. Structural Features of Insect COX Subunit IV
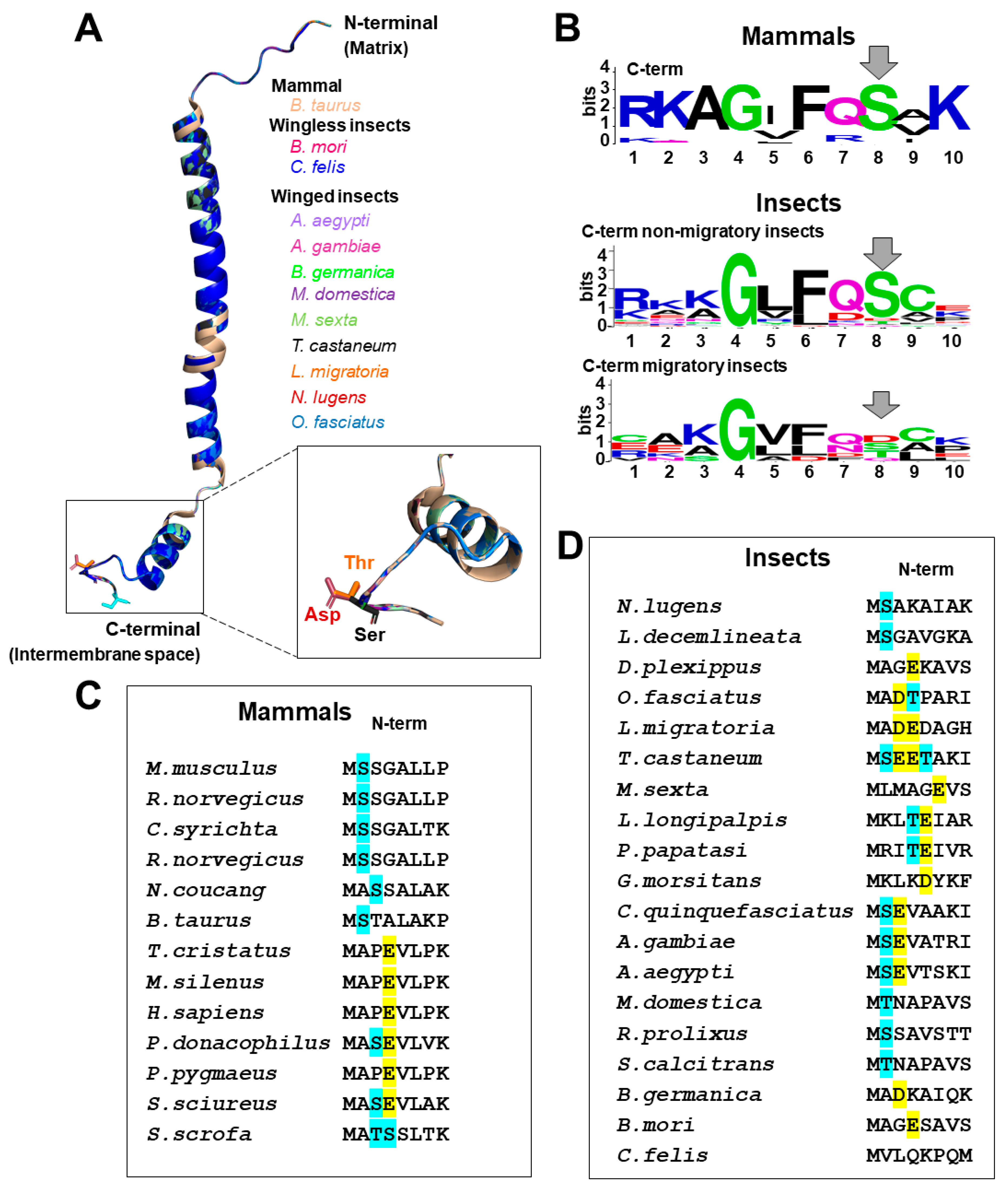
3.2. Structural Features of Insect COX Subunit VI
4. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stork, N.E.; McBroom, J.; Gely, C.; Hamilton, A.J. New approaches narrow global species estimates for beetles, insects, and terrestrial arthropods. Proc. Natl. Acad. Sci. USA 2015, 112, 7519–7523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hunter, P. The nature of flight. EMBO Rep. 2007, 8, 811–813. [Google Scholar] [CrossRef] [Green Version]
- Graham, J.B.; Aguilar, N.M.; Dudley, R.; Gans, C. Implications of the late Palaeozoic oxygen pulse for physiology and evolution. Nature 1995, 375, 117–120. [Google Scholar] [CrossRef]
- May, M.L. Heat exchange and endothermy in protodonata. Evolution 1982, 36, 1051–1058. [Google Scholar] [CrossRef]
- Dudley, R. Atmospheric oxygen, giant Paleozoic insects and the evolution of aerial locomotor performance. J. Exp. Biol. 1998, 201, 1043–1050. [Google Scholar]
- Prokop, J.; Nel, A.; Hoch, I. Discovery of the oldest known Pterygota in the Lower Carboniferous of the Upper Silesian Basin in the Czech Republic (Insecta: Archaeorthoptera). Geobios 2005, 38, 383–387. [Google Scholar] [CrossRef]
- Harrison, J.F.; Kaiser, A.; VandenBrooks, J.M. Atmospheric oxygen level and the evolution of insect body size. Proc. Biol. Sci. 2010, 277, 1937–1946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hetz, S.K.; Bradley, T.J. Insects breathe discontinuously to avoid oxygen toxicity. Nature 2005, 433, 516–519. [Google Scholar] [CrossRef]
- Kaiser, A.; Klok, C.J.; Socha, J.J.; Lee, W.K.; Quinlan, M.C.; Harrison, J.F. Increase in tracheal investment with beetle size supports hypothesis of oxygen limitation on insect gigantism. Proc. Natl. Acad. Sci. USA 2007, 104, 13198–13203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harrison, J.F.; Roberts, S.P. Flight respiration and energetics. Annu. Rev. Physiol. 2000, 62, 179–205. [Google Scholar] [CrossRef]
- Bomphrey, R.J.; Nakata, T.; Phillips, N.; Walker, S.M. Smart wing rotation and trailing-edge vortices enable high frequency mosquito flight. Nature 2017, 544, 92–95. [Google Scholar] [CrossRef] [PubMed]
- Sacktor, B. Cell structure and the metabolism of insect flight muscle. J. Biophys. Biochem. Cytol. 1955, 1, 29–46. [Google Scholar] [CrossRef] [Green Version]
- Smith, D.S. The structure of flight muscle sarcosomes in the blowfly Calliphora erythrocephala (diptera). J. Cell Biol. 1963, 19, 115–138. [Google Scholar] [CrossRef] [Green Version]
- Sacktor, B.; Cochran, D.G. The respiratory metabolism of insect flight muscle. I. Manometric studies of oxidation and concomitant phosphorylation with sarcosomes. Arch. Biochem. Biophys. 1958, 74, 266–276. [Google Scholar] [CrossRef]
- Lennie, R.W.; Birt, L.M. Aspects of the development of flight-muscle sarcosomes in the sheep blowfly, Luciliacuprina, in relation to changes in the distribution of protein and some respiratory enzymes during metamorphosis. Biochem. J. 1967, 102, 338–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, R.A.; Fraenkel, G. The oxygen consumption of flies during flight. J. Exp. Biol. 1940, 17, 402–407. [Google Scholar]
- Iwamoto, H. Structure, function and evolution of insect flight muscle. Biophysics 2011, 7, 21–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farmer, K.J.; Sohal, R.S. Relationship between superoxide anion radical generation and aging in the housefly, Musca domestica. Free Radic. Biol. Med. 1989, 7, 23–29. [Google Scholar] [CrossRef]
- Miwa, S.; St-Pierre, J.; Partridge, L.; Brand, M.D. Superoxide and hydrogen peroxide production by Drosophila mitochondria. Free Radic. Biol. Med. 2003, 35, 938–948. [Google Scholar] [CrossRef]
- Gonçalves, R.L.S.; Machado, A.C.L.; Paiva-Silva, G.O.; Sorgine, M.H.F.; Momoli, M.M.; Oliveira, J.H.M.; Vannier-Santos, M.A.; Galina, A.; Oliveira, P.L.; Oliveira, M.F. Blood-Feeding induces reversible functional changes in flight muscle mitochondria of Aedes aegypti mosquito. PLoS ONE 2009, 4, e7854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soares, J.B.R.C.; Gaviraghi, A.; Oliveira, M.F. Mitochondrial physiology in the major arbovirus vector Aedes aegypti: Substrate preferences and sexual differences define respiratory capacity and superoxide production. PLoS ONE 2015, 10, e0120600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindstedt, S.L.; McGlothlin, T.; Percy, E.; Pifer, J. Task-Specific design of skeletal muscle: Balancing muscle structural composition. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 1998, 120, 35–40. [Google Scholar] [CrossRef]
- Sohal, R.S.; McCarthy, J.L.; Allison, V.F. The formation of ‘giant’ mitochondria in the fibrillar flight muscles of the house fly, Musca domestica L. J. Ultrastruct. Res. 1972, 39, 484–495. [Google Scholar] [CrossRef]
- Taylor, C.R.; Heglund, N.C. Energetics and mechanics of terrestrial locomotion. Annu. Rev. Physiol. 1981, 44, 97–107. [Google Scholar] [CrossRef]
- Ellington, C.P. Power and efficiency of insect flight muscle. J. Exp. Biol. 1985, 115, 293–304. [Google Scholar]
- Masson, S.W.C.; Hedges, C.P.; Devaux, J.B.L.; James, C.S.; Hickey, A.J.R. Mitochondrial glycerol 3-phosphate facilitates bumblebee pre-flight thermogenesis. Sci. Rep. 2017, 7, 13107. [Google Scholar] [CrossRef] [Green Version]
- Gaviraghi, A.; Oliveira, M.F. A method for assessing mitochondrial physiology using mechanically permeabilized flight muscle of Aedes aegypti mosquitoes. Anal. Biochem. 2019, 576, 33–41. [Google Scholar] [CrossRef]
- Roeder, K.D. Movements of the thorax and potential changes in the thoracic muscles of insects during flight. Biol. Bull. 1951, 100, 95–106. [Google Scholar] [CrossRef] [PubMed]
- Pringle, J.W.S. The excitation and contraction of the flight muscles of insects. J. Physiol. 1949, 108, 226–232. [Google Scholar] [CrossRef]
- Josephson, R.K.; Malamud, J.G.; Stokes, D.R. Asynchronous muscle: A primer. J. Exp. Biol. 2000, 203, 2713–2722. [Google Scholar] [PubMed]
- Cullen, M.J. The distribution of asynchronous muscle in insects with particular reference to the Hemiptera: An electron microscope study. J. Entomol. Ser. Gen. Entomol. 1974, 49, 17–41. [Google Scholar] [CrossRef]
- Lund, J.; Webb, M.R.; White, D.C. Changes in the ATPase activity of insect fibrillar flight muscle during calcium and strain activation probed by phosphate-water oxygen exchange. J. Biol. Chem. 1987, 262, 8584–8590. [Google Scholar] [CrossRef]
- Josephson, R.K.; Young, D. Synchronous and asynchronous muscles in Cicadas. J. Exp. Biol. 1981, 91, 219–237. [Google Scholar]
- Josephson, R.K.; Malamud, J.G.; Stokes, D.R. The efficiency of an asynchronous flight muscle from a beetle. J. Exp. Biol. 2001, 204, 4125–4139. [Google Scholar]
- Linari, M.; Reedy, M.K.; Reedy, M.C.; Lombardi, V.; Piazzesi, G. Ca-Activation and stretch-activation in insect flight muscle. Biophys. J. 2004, 87, 1101–1111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makarieva, A.M.; Gorshkov, V.G.; Li, B. Energetics of the smallest: Do bacteria breathe at the same rate as whales? Proc. Biol. Sci. 2005, 272, 2219–2224. [Google Scholar] [CrossRef] [Green Version]
- Beenakkers, A.M.T.; Van der Horst, D.J.; Van Marrewijk, W.J.A. Insect flight muscle metabolism. Insect Biochem. 1984, 14, 243–260. [Google Scholar] [CrossRef]
- Krogh, A.; Weis-Fogh, T. The respiratory exchange of the desert locust (Schistocerca gregaria) before, during and after flight. J. Exp. Biol. 1951, 28, 344–357. [Google Scholar]
- Cao, T.; Jin, J.P. Evolution of flight muscle contractility and energetic efficiency. Front. Physiol. 2020, 11, 1038. [Google Scholar] [CrossRef]
- Ugur, B.; Chen, K.; Bellen, H.J. Drosophila tools and assays for the study of human diseases. Dis. Mod. Mech. 2016, 9, 235–244. [Google Scholar] [CrossRef] [Green Version]
- Tulp, A.; Stam, H.; van Dam, K. Permeability of housefly mitochondria towards dicarboxylate ions. Biochim. Biophys. Acta 1971, 234, 301–305. [Google Scholar] [CrossRef]
- van den Bergh, S.; Slater, E.C. The respiratory activity and permeability of housefly sarcosomes. Biochem. J. 1962, 82, 362–371. [Google Scholar] [CrossRef] [Green Version]
- Ferguson, M.; Mockett, R.J.; Shen, Y.; Orr, W.C.; Sohal, R.S. Age-Associated decline in mitochondrial respiration and electron transport in Drosophila melanogaster. Biochem. J. 2005, 390, 501–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansford, R.G.; Johnson, R.N. The nature and control of the tricarboxylate cycle in beetle flight muscle. Biochem. J. 1975, 148, 389–401. [Google Scholar] [CrossRef] [Green Version]
- Syromyatnikov, M.Y.; Gureev, A.P.; Vitkalova, I.Y.; Starkov, A.A.; Popov, V.N. Unique features of flight muscles mitochondria of honey bees (Apis mellifera L.). Arch. Insect Biochem. Physiol. 2019, 102, e21595. [Google Scholar] [CrossRef] [PubMed]
- Kashi, K.P.; Bond, E.J. Studies on isolation and respiratory activity of the mitochondria of Sitophilus granarius (L.). J. Stored Prod. Res. 1979, 15, 17–24. [Google Scholar] [CrossRef]
- Chamberlin, M.E. Changes in mitochondrial electron transport chain activity during insect metamorphosis. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 292, R1016–R1022. [Google Scholar] [CrossRef] [PubMed]
- Carafoli, E.; Lehninger, A.L. A survey of the interaction of calcium ions with mitochondria from different tissues and species. Biochem. J. 1971, 122, 681–690. [Google Scholar] [CrossRef] [Green Version]
- Carafoli, E.; Hansford, R.G.; Sacktor, B.; Lehninger, A.L. Interaction of Ca2+ with blowfly flight muscle mitochondria. J. Biol. Chem. 1971, 246, 964–972. [Google Scholar] [CrossRef]
- Zebe, E.C.; McShan, W.H. Lactic and α-glycerophosphate dehydrogenases in insects. J. Gen. Physiol. 1957, 40, 779–790. [Google Scholar] [CrossRef] [Green Version]
- Kubista, V. Accumulation of a stable phosphorus compound in glycolysing insect muscle. Nature 1957, 180, 549. [Google Scholar] [CrossRef] [PubMed]
- Estabrook, R.W.; Sacktor, B. α-glycerophosphate oxidase of flight muscle mitochondria. J. Biol. Chem. 1958, 233, 1014–1019. [Google Scholar] [CrossRef]
- Mráček, T.; Drahota, Z.; Houštěk, J. The function and the role of the mitochondrial glycerol-3-phosphate dehydrogenase in mammalian tissues. Biochim. Biophys. Acta 2013, 1827, 401–410. [Google Scholar] [CrossRef] [Green Version]
- Ohkawa, K.I.; Vogt, M.T.; Farber, E. Unusually high mitochondrial α glycerophosphate dehydrogenase activity in rat brown adipose tissue. J. Cell Biol. 1969, 41, 441–449. [Google Scholar] [CrossRef] [Green Version]
- Tribe, M.A.; Ashhurst, D.E. Biochemical and structural variations in the flight muscle mitochondria of ageing blowflies, Calliphoraerythrocephala. J. Cell Sci. 1972, 10, 443–469. [Google Scholar]
- Worm, R.A.A.; Beenakkers, A.M.T. Regulation of substrate utilization in the flight muscle of the locust, Locusta migratoria, during flight. Insect Biochem. 1980, 10, 53–59. [Google Scholar] [CrossRef]
- Van Handel, E.; Nayar, J.K. Turn-Over of diglycerides during flight and rest in the moth Spodoptera frugiperda. Insect Biochem. 1972, 2, 8–12. [Google Scholar] [CrossRef]
- Suarez, R.K.; Darveau, C.-A.; Welch, K.C.; O’Brien, D.M.; Roubik, D.W.; Hochachka, P.W. Energy metabolism in orchid bee flight muscles: Carbohydrate fuels all. J. Exp. Biol. 2005, 208, 3573–3579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bursell, E. Substrates of oxidative metabolism in dipteran flight muscle. Comp. Biochem. Physiol. B 1975, 52, 235–238. [Google Scholar] [CrossRef]
- Teulier, L.; Weber, J.-M.; Crevier, J.; Darveau, C.-A. Proline as a fuel for insect flight: Enhancing carbohydrate oxidation in hymenopterans. Proc. R. Soc. B Biol. Sci. 2016, 283, 20160333. [Google Scholar] [CrossRef] [Green Version]
- Bursell, E.; Slack, E. Oxidation of proline by sarcosomes of the tsetse fly, Glossina morsitans. Insect Biochem. 1976, 6, 159–167. [Google Scholar] [CrossRef]
- Gäde, G.; Auerswald, L. Beetles’ choice—Proline for energy output: Control by AKHs. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2002, 132, 117–129. [Google Scholar] [CrossRef]
- Auerswald, L.; Schneider, P.; Gäde, G. Utilisation of substrates during tethered flight with and without lift generation in the African fruit beetle Pachnodasinuata (Cetoniinae). J. Exp. Biol. 1998, 201, 2333–2342. [Google Scholar] [PubMed]
- Hansford, R.G.; Sacktor, B. The control of the oxidation of proline by isolated flight muscle mitochondria. J. Biol. Chem. 1970, 245, 991–994. [Google Scholar] [CrossRef]
- Sacktor, B.; Childress, C.C. Metabolism of proline in insect flight muscle and its significance in stimulating the oxidation of pyruvate. Arch. Biochem. Biophys. 1967, 120, 583–588. [Google Scholar] [CrossRef]
- Balboni, E. A proline shuttle in insect flight muscle. Biochem. Biophys. Res. Commun. 1978, 85, 1090–1096. [Google Scholar] [CrossRef]
- Jutsum, A.R.; Goldsworthy, G.J. Fuels for flight in Locusta. J. Insect Physiol. 1976, 22, 243–249. [Google Scholar] [CrossRef]
- van der Horst, D.J.; Baljet, A.M.C.; Beenakkers, A.M.T.; van Handel, E. Turnover of locust haemolymph diglycerides during flight and rest. Insect Biochem. 1978, 8, 369–373. [Google Scholar] [CrossRef]
- Scaraffia, P.Y.; Wells, M.A. Proline can be utilized as an energy substrate during flight of Aedes aegypti females. J. Insect Physiol. 2003, 49, 591–601. [Google Scholar] [CrossRef]
- Cao, T.; Sujkowski, A.; Cobb, T.; Wessells, R.J.; Jin, J.P. The glutamic acid-rich-long C-terminal extension of troponin T has a critical role in insect muscle functions. J. Biol. Chem. 2020, 295, 3794–3807. [Google Scholar] [CrossRef] [Green Version]
- Suarez, R.K.; Staples, J.F.; Lighton, J.R.; Mathieu-Costello, O. Mitochondrial function in flying honeybees (Apis mellifera): Respiratory chain enzymes and electron flow from complex III to oxygen. J. Exp. Biol. 2000, 203, 905–911. [Google Scholar] [PubMed]
- Suarez, R.K.; Lighton, J.R.; Joos, B.; Roberts, S.P.; Harrison, J.F. Energy metabolism, enzymatic flux capacities, and metabolic flux rates in flying honeybees. Proc. Natl. Acad. Sci. USA 1996, 93, 12616–12620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eanes, W.F.; Merritt, T.J.S.; Flowers, J.M.; Kumagai, S.; Sezgin, E.; Zhu, C.T. Flux control and excess capacity in the enzymes of glycolysis and their relationship to flight metabolism in Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 2006, 103, 19413–19418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merritt, T.J.S.; Sezgin, E.; Zhu, C.T.; Eanes, W.F. Triglyceride pools, flight and activity variation at the Gpdh locus in Drosophila melanogaster. Genetics 2006, 172, 293–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levin, E.; Lopez-Martinez, G.; Fane, B.; Davidowitz, G. Hawkmoths use nectar sugar to reduce oxidative damage from flight. Science 2017, 355, 733–735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gottsberger, G.; Schrauwen, J.; Linskens, H.F. Amino acids and sugars in nectar, and their putative evolutionary significance. Plant Syst. Evol. 1984, 145, 55–77. [Google Scholar] [CrossRef]
- Lehane, M. Biology of Blood-Sucking Insects; Springer Science & Business Media: London, UK, 1991. [Google Scholar]
- Sterkel, M.; Oliveira, J.H.M.; Bottino-Rojas, V.; Paiva-Silva, G.O.; Oliveira, P.L. The dose makes the poison: Nutritional overload determines the life traits of blood-feeding arthropods. Trends Parasitol. 2017, 33, 633–644. [Google Scholar] [CrossRef]
- Gaviraghi, A.; Oliveira, M.F. A simple and reliable method for longitudinal assessment of untethered mosquito induced flight activity. J. Insect Physiol. 2020, 126, 104098. [Google Scholar] [CrossRef]
- Nicholls, D.G.; Ferguson, S.J. Bioenergetics 4, 4th ed.; Academic Press: Boston, MA, USA, 2013; pp. 54–87. [Google Scholar]
- Wegener, G.; Bolas, N.M.; Thomas, A.A.G. Locust flight metabolism studied in vivo by 31P NMR spectroscopy. J. Comp. Physiol. B 1991, 161, 247–256. [Google Scholar] [CrossRef]
- Sacktor, B.; Hurlbut, E.C. Regulation of metabolism in working muscle in vivo. II. Concentrations of adenine nucleotides, arginine phosphate, and inorganic phosphate in insect flight muscle during flight. J. Biol. Chem. 1966, 241, 632–634. [Google Scholar] [CrossRef]
- Mitchell, P. Coupling of phosphorylation to electron and hydrogen transfer by a chemi-osmotic type of mechanism. Nature 1961, 191, 144–148. [Google Scholar] [CrossRef]
- Chance, B.; Williams, G.R. Respiratory enzymes in oxidative phosphorylation. III. The steady state. J. Biol. Chem. 1955, 217, 409–427. [Google Scholar] [CrossRef]
- Lardy, H.A.; Wellman, H. Oxidative phosphorylations; role of inorganic phosphate and acceptor systems in control of metabolic rates. J. Biol. Chem. 1952, 195, 215–224. [Google Scholar] [CrossRef]
- Hansford, R.G. Some properties of pyruvate and 2-oxoglutarate oxidation by blowfly flight-muscle mitochondria. Biochem. J. 1972, 127, 271–283. [Google Scholar] [CrossRef]
- Hansford, R.G.; Chappell, J.B. The energy-dependent accumulation of phosphate by blowfly mitochondria and its effect on the rate of pyruvate oxidation. Biochem. Biophys. Res. Commun. 1968, 30, 643–648. [Google Scholar] [CrossRef]
- Arnold, S.; Kadenbach, B. The intramitochondrial ATP/ADP-ratio controls cytochrome c oxidase activity allosterically. FEBS Lett. 1999, 443, 105–108. [Google Scholar] [CrossRef] [Green Version]
- Arnold, S.; Kadenbach, B. Cell respiration is controlled by ATP, an allosteric inhibitor of cytochrome-c oxidase. Eur. J. Biochem. 1997, 249, 350–354. [Google Scholar] [CrossRef] [PubMed]
- Gaviraghi, A.; Soares, J.B.R.C.; Mignaco, J.A.; Fontes, C.F.L.; Oliveira, M.F. Mitochondrial glycerol phosphate oxidation is modulated by adenylates through allosteric regulation of cytochrome c oxidase activity in mosquito flight muscle. Insect Biochem. Mol. Biol. 2019, 114, 103226. [Google Scholar] [CrossRef] [PubMed]
- Hochachka, P.W.; Nener, J.C.; Hoar, J.; Suarez, R.K.; Hand, S.C. Disconnecting metabolism from adenylate control during extreme oxygen limitation. Can. J. Zool. 1993, 71, 1267–1270. [Google Scholar] [CrossRef]
- Hochachka, P.W. Muscles as Molecular and Metabolic Machines, 1st ed.; CRC Press: Boca Raton, FL, USA, 1994; pp. 95–118. [Google Scholar]
- Crabtree, B.; Newsholme, E.A. The activities of phosphorylase, hexokinase, phosphofructokinase, lactate dehydrogenase and the glycerol 3-phosphate dehydrogenases in muscles from vertebrates and invertebrates. Biochem. J. 1972, 126, 49–58. [Google Scholar] [CrossRef]
- Mac Munn, C.A. Researches on Myohaematin and the Histohaematins. Philos. Trans. R. Soc. Lond. 1886, 177, 267–298. [Google Scholar] [CrossRef]
- Keilin, D. On cytochrome, a respiratory pigment, common to animals, yeast, and higher plants. Proc. R. Soc. Lond. B 1925, 98, 312–339. [Google Scholar]
- Watanabe, M.I.; Williams, C.M. Mitochondria in the flight muscles of insects. I. Chemical composition and enzymatic content. J. Gen. Physiol. 1951, 34, 675–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levenbook, L.; Williams, C.M. Mitochondria in the flight muscles of insects III. Mitochondrial cytochrome c in relation to the aging and wing beat frequency of flies. J. Gen. Physiol. 1956, 39, 497–512. [Google Scholar] [CrossRef] [Green Version]
- Herold, R.C.; Borei, H. Cytochrome changes during honey bee flight muscle development. Dev. Biol. 1963, 8, 67–79. [Google Scholar] [CrossRef]
- Sacktor, B. A comparison of the cytochrome oxidase activity of two strains of house flies. J. Econ. Entomol. 1950, 43, 832–838. [Google Scholar] [CrossRef]
- Allen, T.H. Enzymes in ontogenesis (Orthoptera) XI. Cytochrome oxidase in relation to respiratory activity and growth of the grasshopper egg. J. Cell. Comp. Physiol. 1940, 16, 149–163. [Google Scholar] [CrossRef]
- Ludwig, D. Cytochrome oxidase activity during diapause and metamorphosis of the Japanese beetle (Popillia japonica Newman). J. Gen. Physiol. 1953, 36, 751–757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mason, C.J.; Scully, E.D.; Geib, S.M.; Hoover, K. Contrasting diets reveal metabolic plasticity in the tree-killing beetle, Anoplophora glabripennis (Cerambycidae: Lamiinae). Sci. Rep. 2016, 6, 3813. [Google Scholar] [CrossRef] [Green Version]
- Tian, H.; Vinson, S.B.; Coates, C.J. Differential gene expression between alate and dealate queens in the red imported fire ant, Solenopsis invicta Buren (Hymenoptera: Formicidae). Insect Biochem. Mol. Biol. 2004, 34, 937–949. [Google Scholar] [CrossRef]
- Oliveira, P.L.; Oliveira, M.F. Vampires, Pasteur and reactive oxygen species. Is the switch from aerobic to anaerobic metabolism a preventive antioxidant defence in blood-feeding parasites? FEBS Lett. 2002, 525, 3–6. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, C.M.; Oliveira, M.P.; Paes, M.C.; Oliveira, M.F. Modulation of mitochondrial metabolism as a biochemical trait in blood feeding organisms: The redox vampire hypothesis redux. Cell Biol. Int. 2018, 42, 683–700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, C.G. Migration and Dispersal of Insects by Flight; Methuen & Co. Ltd.: London, UK, 1969; p. 763. [Google Scholar]
- Kim, K.; Yang, J.O.; Sung, J.Y.; Lee, J.Y.; Park, J.S.; Lee, H.S.; Lee, B.H.; Ren, Y.; Lee, D.W.; Lee, S.E. Minimization of energy transduction confers resistance to phosphine in the rice weevil, Sitophilus oryzae. Sci. Rep. 2019, 9, 14605. [Google Scholar] [CrossRef] [Green Version]
- Pridgeon, J.W.; Liu, N. Overexpression of the cytochrome c oxidase subunit I gene associated with a pyrethroid resistant strain of German cockroaches, Blattella germanica (L.). Insect Biochem. Mol. Biol. 2003, 33, 1043–1048. [Google Scholar] [CrossRef]
- Sacktor, B.; Shimada, Y. Degenerative changes in the mitochondria of flight muscle from aging blowflies. J. Cell Biol. 1972, 52, 465–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sohal, R.S. Aging, cytochrome oxidase activity, and hydrogen peroxide release by mitochondria. Free Radic. Biol. Med. 1993, 14, 583–588. [Google Scholar] [CrossRef]
- Ren, J.C.; Rebrin, I.; Klichko, V.; Orr, W.C.; Sohal, R.S. Cytochrome c oxidase loses catalytic activity and structural integrity during the aging process in Drosophila melanogaster. Biochem. Biophys. Res. Commun. 2010, 401, 64–68. [Google Scholar] [CrossRef] [Green Version]
- Sohal, R.S.; Toroser, D.; Brégère, C.; Mockett, R.J.; Orr, W.C. Age-Related decrease in expression of mitochondrial DNA encoded subunits of cytochrome c oxidase in Drosophila melanogaster. Mech. Ageing Dev. 2008, 129, 558–561. [Google Scholar] [CrossRef] [Green Version]
- Schwarze, S.R.; Weindruch, R.; Aiken, J.M. Oxidative stress and aging reduce COX I RNA and cytochrome oxidase activity in Drosophila. Free Radic. Biol. Med. 1998, 25, 740–747. [Google Scholar] [CrossRef]
- Walker, D.W.; Benzer, S. Mitochondrial “swirls” induced by oxygen stress and in the Drosophila mutant hyperswirl. Proc. Natl. Acad. Sci. USA 2004, 101, 10290–10295. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.F.; Lin, H.L.; Wang, L.J.; Hsu, T.; Fu, C.Y. Coordinated organization of mitochondrial lamellar cristae and gain of COX function during mitochondrial maturation in Drosophila. Mol. Biol. Cell 2020, 31, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Wolf, D.M.; Segawa, M.; Kondadi, A.K.; Anand, R.; Bailey, S.T.; Reichert, A.S.; van der Bliek, A.M.; Shackelford, D.B.; Liesa, M.; Shirihai, O.S. Individual cristae within the same mitochondrion display different membrane potentials and are functionally independent. EMBO J. 2019, 38, e101056. [Google Scholar] [CrossRef] [PubMed]
- Rauhamäki, V.; Wolfram, J.; Jokitalo, E.; Hanski, I.; Dahlhoff, E.P. Differences in the aerobic capacity of flight muscles between butterfly populations and species with dissimilar flight abilities. PLoS ONE 2014, 9, e78069. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.-Y.; Chen, B.; Zhao, D.-J.; Kang, L. Functional modulation of mitochondrial cytochrome c oxidase underlies adaptation to high-altitude hypoxia in a Tibetan migratory locust. Proc. Biol. Sci. 2013, 280, 20122758. [Google Scholar] [CrossRef] [Green Version]
- Singtripop, T.; Saeangsakda, M.; Tatun, N.; Kaneko, Y.; Sakurai, S. Correlation of oxygen consumption, cytochrome c oxidase, and cytochrome c oxidase subunit I gene expression in the termination of larval diapause in the bamboo borer, Omphisa fuscidentalis. J. Insect Physiol. 2007, 53, 933–939. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.; Ma, M.; Zhai, N.; Liu, Z.; Wang, H.; Guo, X.; Xu, B. Roles of a mitochondrial AccSCO2 gene from Apis cerana cerana in oxidative stress responses. J. Inorg. Biochem. 2017, 175, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Napiwotzki, J.; Shinzawa-Itoh, K.; Yoshikawa, S.; Kadenbach, B. ATP and ADP bind to cytochrome c oxidase and regulate its activity. Biol. Chem. 1997, 378, 1013–1021. [Google Scholar] [CrossRef] [PubMed]
- Napiwotzki, J.; Kadenbach, B. Extramitochondrial ATP/ADP-ratios regulate cytochrome c oxidase activity via binding to the cytosolic domain of subunit IV. Biol. Chem. 1998, 379, 335–339. [Google Scholar] [CrossRef]
- Kadenbach, B. Complex IV-the regulatory center of mitochondrial oxidative phosphorylation. Mitochondrion 2020, in press. [Google Scholar] [CrossRef]
- Balsa, E.; Marco, R.; Perales-Clemente, E.; Szklarczyk, R.; Calvo, E.; Landázuri, M.O.; Enríquez, J.A. NDUFA4 is a subunit of complex IV of the mammalian electron transport chain. Cell Metab. 2012, 16, 378–386. [Google Scholar] [CrossRef] [Green Version]
- Villani, G.; Greco, M.; Papa, S.; Attardi, G. Low reserve of cytochrome c oxidase capacity in vivo in the respiratory chain of a variety of human cell types. J. Biol. Chem. 1998, 273, 31829–31836. [Google Scholar] [CrossRef] [Green Version]
- Hwang, H.J.; Lynn, S.G.; Vengellur, A.; Saini, Y.; Grier, E.A.; Ferguson-Miller, S.M.; LaPres, J.J. Hypoxia inducible factors modulate mitochondrial oxygen consumption and transcriptional regulation of nuclear-encoded electron transport chain genes. Biochemistry 2015, 54, 3739–3748. [Google Scholar] [CrossRef] [Green Version]
- Ramzan, R.; Rhiel, A.; Weber, P.; Kadenbach, B.; Vogt, S. Reversible dimerization of cytochrome c oxidase regulates mitochondrial respiration. Mitochondrion 2019, 49, 149–155. [Google Scholar] [CrossRef]
- Lee, I.; Kadenbach, B. Palmitate decreases proton pumping of liver-type cytochrome c oxidase. Eur. J. Biochem. 2001, 268, 6329–6334. [Google Scholar] [CrossRef]
- Cooper, C.E.; Brown, G.C. The inhibition of mitochondrial cytochrome oxidase by the gases carbon monoxide, nitric oxide, hydrogen cyanide and hydrogen sulfide: Chemical mechanism and physiological significance. J. Bioenerg. Biomembr. 2008, 40, 533–539. [Google Scholar] [CrossRef] [PubMed]
- Vygodina, T.; Kirichenko, A.; Konstantinov, A.A. Direct regulation of cytochrome c oxidase by calcium ions. PLoS ONE 2013, 8, e74436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, L.; Mills, D.A.; Buhrow, L.; Hiser, C.; Ferguson-Miller, S. A conserved steroid binding site in cytochrome c oxidase. Biochemistry 2008, 47, 9931–9933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnold, S.; Goglia, F.; Kadenbach, B. 3,5-Diiodothyronine binds to subunit Va of cytochrome-c oxidase and abolishes the allosteric inhibition of respiration by ATP. Eur. J. Biochem. 1998, 252, 325–330. [Google Scholar] [CrossRef] [PubMed]
- Steenaart, N.A.; Shore, G.C. Mitochondrial cytochrome c oxidase subunit IV is phosphorylated by an endogenous kinase. FEBS Lett. 1997, 415, 294–298. [Google Scholar] [CrossRef] [Green Version]
- Miyazaki, T.; Neff, L.; Tanaka, S.; Horne, W.C.; Baron, R. Regulation of cytochrome c oxidase activity by c-Src in osteoclasts. J. Cell Biol. 2003, 160, 709–718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, I.; Salomon, A.R.; Ficarro, S.; Mathes, I.; Lottspeich, F.; Grossman, L.I.; Hüttemann, M. cAMP-Dependent tyrosine phosphorylation of subunit I inhibits cytochrome c oxidase activity. J. Biol. Chem. 2005, 280, 6094–6100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bender, E.; Kadenbach, B. The allosteric ATP-inhibition of cytochrome c oxidase activity is reversibly switched on by cAMP-dependent phosphorylation. FEBS Lett. 2000, 466, 130–134. [Google Scholar] [CrossRef] [Green Version]
- Guo, D.; Nguyen, T.; Ogbi, M.; Tawfik, H.; Ma, G.; Yu, Q.; Caldwell, R.W.; Johnson, J.A. Protein kinase C-epsilon co immunoprecipitates with cytochrome oxidase subunit IV and is associated with improved cytochrome-c oxidase activity and cardioprotection. Am. J. Physiol. Heart Circ. Physiol. 2007, 293, H2219–H2230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogbi, M.; Chew, C.S.; Pohl, J.; Stuchlik, O.; Ogbi, S.; Johnson, J.A. Cytochrome c oxidase subunit IV as a marker of protein kinase C epsilon function in neonatal cardiac myocytes: Implications for cytochrome c oxidase activity. Biochem. J. 2004, 382, 923–932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogbi, M.; Johnson, J.A. Protein kinase C epsilon interacts with cytochrome c oxidase subunit IV and enhances cytochrome c oxidase activity in neonatal cardiac myocyte preconditioning. Biochem. J. 2006, 393, 191–199. [Google Scholar] [CrossRef] [Green Version]
- Lee, I.; Bender, E.; Kadenbach, B. Control of mitochondrial membrane potential and ROS formation by reversible phosphorylation of cytochrome c oxidase. Mol. Cell. Biochem. 2002, 234–235, 63–70. [Google Scholar] [CrossRef]
- Helling, S.; Vogt, S.; Rhiel, A.; Ramzan, R.; Wen, L.; Marcus, K.; Kadenbach, B. Phosphorylation and kinetics of mammalian cytochrome c oxidase. Mol. Cell. Proteom. 2008, 7, 1714–1724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prabu, S.K.; Anandatheerthavarada, H.K.; Raza, H.; Srinivasan, S.; Spear, J.F.; Avadhani, N.G. Protein kinase A-mediated phosphorylation modulates cytochrome c oxidase function and augments hypoxia and myocardial ischemia-related injury. J. Biol. Chem. 2006, 281, 2061–2070. [Google Scholar] [CrossRef] [Green Version]
- Fang, J.-K.; Prabu, S.K.; Sepuri, N.B.; Raza, H.; Anandatheerthavarada, H.K.; Galati, D.; Spear, J.; Avadhani, N.G. Site specific phosphorylation of cytochrome c oxidase subunits I, IVi1 and Vb in rabbit hearts subjected to ischemia/reperfusion. FEBS Lett. 2007, 581, 1302–1310. [Google Scholar] [CrossRef] [Green Version]
- Srinivasan, S.; Spear, J.; Chandran, K.; Joseph, J.; Kalyanaraman, B.; Avadhani, N.G. Oxidative stress induced mitochondrial protein kinase A mediates cytochrome c oxidase dysfunction. PLoS ONE 2013, 8, e77129. [Google Scholar] [CrossRef] [Green Version]
- Samavati, L.; Lee, I.; Mathes, I.; Lottspeich, F.; Hüttemann, M. Tumor necrosis factor α inhibits oxidative phosphorylation through tyrosine phosphorylation at subunit I of cytochrome c oxidase. J. Biol. Chem. 2008, 283, 21134–21144. [Google Scholar] [CrossRef] [Green Version]
- Ramzan, R.; Staniek, K.; Kadenbach, B.; Vogt, S. Mitochondrial respiration and membrane potential are regulated by the allosteric ATP-inhibition of cytochrome c oxidase. Biochim. Biophys. Acta 2010, 1797, 1672–1680. [Google Scholar] [CrossRef] [Green Version]
- Lundby, A.; Secher, A.; Lage, K.; Nordsborg, N.B.; Dmytriyev, A.; Lundby, C.; Olsen, J.V. Quantitative maps of protein phosphorylation sites across 14 different rat organs and tissues. Nat. Commun. 2012, 3, 876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acin-Perez, R.; Salazar, E.; Kamenetsky, M.; Buck, J.; Levin, L.R.; Manfredi, G. Cyclic AMP produced inside mitochondria regulates oxidative phosphorylation. Cell Metab. 2009, 9, 265–276. [Google Scholar] [CrossRef] [Green Version]
- Hess, K.C.; Liu, J.; Manfredi, G.; Mühlschlegel, F.A.; Buck, J.; Levin, L.R.; Barrientos, A. A mitochondrial CO2-adenylyl cyclase-cAMP signalosome controls yeast normoxic cytochrome c oxidase activity. FASEB J. 2014, 28, 4369–4380. [Google Scholar] [CrossRef] [Green Version]
- Acin-Perez, R.; Gatti, D.L.; Bai, Y.; Manfredi, G. Protein phosphorylation and prevention of cytochrome oxidase inhibition by ATP: Coupled mechanisms of energy metabolism regulation. Cell Metab. 2011, 13, 712–719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Follmann, K.; Arnold, S.; Ferguson-Miller, S.; Kadenbach, B. Cytochrome c oxidase from eucaryotes but not from procaryotes is allosterically inhibited by ATP. Biochem. Mol. Biol. Int. 1998, 45, 1047–1055. [Google Scholar] [CrossRef] [PubMed]
- Alge, D.; Wastyn, M.; Mayer, C.; Jungwirth, C.; Zimmermann, U.; Zoder, R.; Fromwald, S.; Peschek, G.A. Allosteric properties of cyanobacterial cytochrome c oxidase: Inhibition of the coupled enzyme by ATP and stimulation by ADP. IUBMB Life 1999, 48, 187–197. [Google Scholar] [CrossRef]
- Ferguson-Miller, S.; Brautigan, D.L.; Margoliash, E. Correlation of the kinetics of electron transfer activity of various eukaryotic cytochromes c with binding to mitochondrial cytochrome c oxidase. J. Biol. Chem. 1976, 251, 1104–1115. [Google Scholar] [CrossRef]
- Bisson, R.; Schiavo, G.; Montecucco, C. ATP induces conformational changes in mitochondrial cytochrome c oxidase. Effect on the cytochrome c binding site. J. Biol. Chem. 1987, 262, 5992–5998. [Google Scholar] [CrossRef]
- Anthony, G.; Reimann, A.; Kadenbach, B. Tissue-Specific regulation of bovine heart cytochrome-c oxidase activity by ADP via interaction with subunit VIa. Proc. Natl. Acad. Sci. USA 1993, 90, 1652–1656. [Google Scholar] [CrossRef] [Green Version]
- Frank, V.; Kadenbach, B. Regulation of the H+/e- stoichiometry of cytochrome c oxidase from bovine heart by intramitochondrial ATP/ADP ratios. FEBS Lett. 1996, 382, 121–124. [Google Scholar] [CrossRef] [Green Version]
- Taanman, J.W.; Turina, P.; Capaldi, R.A. Regulation of cytochrome c oxidase by interaction of ATP at two binding sites, one on subunit VIa. Biochemistry 1994, 33, 11833–11841. [Google Scholar] [CrossRef]
- Hüther, F.J.; Kadenbach, B. ADP increases the affinity for cytochrome c by interaction with the matrix side of bovine heart cytochrome c oxidase. Biochem. Biophys. Res. Commun. 1987, 147, 1268–1275. [Google Scholar] [CrossRef]
- Kadenbach, B.; Hüttemann, M. The subunit composition and function of mammalian cytochrome c oxidase. Mitochondrion 2015, 24, 64–76. [Google Scholar] [CrossRef] [PubMed]
- Tsukihara, T.; Aoyama, H.; Yamashita, E.; Tomizaki, T.; Yamaguchi, H.; Shinzawa-Itoh, K.; Nakashima, R.; Yaono, R.; Yoshikawa, S. The whole structure of the 13-subunit oxidized cytochrome c oxidase at 2.8 A. Science 1996, 272, 1136–1144. [Google Scholar] [CrossRef]
- Nijtmans, L.G.; Taanman, J.W.; Muijsers, A.O.; Speijer, D.; Van den Bogert, C. Assembly of cytochrome-c oxidase in cultured human cells. Eur. J. Biochem. 1998, 254, 389–394. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Park, J.S.; Deng, J.H.; Bai, Y. Cytochrome c oxidase subunit IV is essential for assembly and respiratory function of the enzyme complex. J. Bioenerg. Biomembr. 2006, 38, 283–291. [Google Scholar] [CrossRef] [Green Version]
- Hüttemann, M.; Lee, I.; Gao, X.; Pecina, P.; Pecinova, A.; Liu, J.; Aras, S.; Sommer, N.; Sanderson, T.H.; Tost, M.; et al. Cytochrome c oxidase subunit 4 isoform 2-knockout mice show reduced enzyme activity, airway hyporeactivity, and lung pathology. FASEB J. 2012, 26, 3916–3930. [Google Scholar] [CrossRef] [Green Version]
- Hüttemann, M.; Kadenbach, B.; Grossman, L.I. Mammalian subunit IV isoforms of cytochrome c oxidase. Gene 2001, 267, 111–123. [Google Scholar] [CrossRef]
- Hüttemann, M.; Lee, I.; Liu, J.; Grossman, L.I. Transcription of mammalian cytochrome c oxidase subunit IV-2 is controlled by a novel conserved oxygen responsive element. FEBS J. 2007, 274, 5737–5748. [Google Scholar] [CrossRef] [PubMed]
- Reguera, D.P.; Čunátová, K.; Vrbacký, M.; Pecinová, A.; Houštěk, J.; Mráček, T.; Pecina, P. Cytochrome c oxidase subunit 4 isoform exchange results in modulation of oxygen affinity. Cells 2020, 9, 443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eddy, S.R. Accelerated Profile HMM Searches. PLoS Comput. Biol. 2011, 7, e1002195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armenteros, J.J.A.; Salvatore, M.; Emanuelsson, O.; Winther, O.; von Heijne, G.; Elofsson, A.; Nielsen, H. Detecting sequence signals in targeting peptides using deep learning. Life Sci. Alliance 2019, 2, e201900429. [Google Scholar] [CrossRef] [Green Version]
- Madeira, F.; Park, Y.M.; Lee, J.; Buso, N.; Gur, T.; Madhusoodanan, N.; Basutkar, P.; Tivey, A.R.N.; Potter, S.C.; Finn, R.D.; et al. The EMBL-EBI search and sequence analysis tools APIs in 2019. Nucleic Acids Res. 2019, 47, W636–W641. [Google Scholar] [CrossRef] [Green Version]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J.E. The Phyre2 web portal for protein modeling, prediction and analysis. Nat. Protoc. 2015, 10, 845–858. [Google Scholar] [CrossRef] [Green Version]
- Tsukihara, T.; Shimokata, K.; Katayama, Y.; Shimada, H.; Muramoto, K.; Aoyama, H.; Mochizuki, M.; Shinzawa-Itoh, K.; Yamashita, E.; Yao, M.; et al. The low-spin heme of cytochrome c oxidase as the driving element of the proton-pumping process. Proc. Natl. Acad. Sci. USA 2003, 100, 15304–15309. [Google Scholar] [CrossRef] [Green Version]
- Liko, I.; Degiacomi, M.T.; Mohammed, S.; Yoshikawa, S.; Schmidt, C.; Robinson, C.V. Dimer interface of bovine cytochrome c oxidase is influenced by local posttranslational modifications and lipid binding. Proc. Natl. Acad. Sci. USA 2016, 113, 8230–8235. [Google Scholar] [CrossRef] [Green Version]
- Schneyder, B.; Mell, O.; Anthony, G.; Kadenbach, B. Cross reactivity of monoclonal antibodies and cDNA hybridization suggest evolutionary relationships between cytochrome c oxidase subunits VIa and VIc and between VIIa and VIIb. Eur. J. Biochem. 1991, 198, 85–92. [Google Scholar] [CrossRef]
- Sampson, V.; Alleyne, T. Cytochrome c/cytochrome c oxidase interaction. Direct structural evidence for conformational changes during enzyme turnover. Eur. J. Biochem. 2001, 268, 6534–6544. [Google Scholar] [CrossRef]
- Linder, D.; Freund, R.; Kadenbach, B. Species-Specific expression of cytochrome c oxidase isozymes. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 1995, 112, 461–469. [Google Scholar] [CrossRef]
- Kadenbach, B.; Barth, J.; Akgün, R.; Freund, R.; Linder, D.; Possekel, S. Regulation of mitochondrial energy generation in health and disease. Biochim. Biophys. Acta 1995, 1271, 103–109. [Google Scholar] [CrossRef] [Green Version]
- Hüttemann, M.; Frank, V.; Kadenbach, B. The possible role of isoforms of cytochrome c oxidase subunit VIa in mammalian thermogenesis. Cell. Mol. Life Sci. 1999, 55, 1482–1490. [Google Scholar] [CrossRef] [PubMed]
- Kadenbach, B.; Frank, V.; Rieger, T.; Napiwotzki, J. Regulation of respiration and energy transduction in cytochrome c oxidase isozymes by allosteric effectors. Mol. Cell. Biochem. 1997, 174, 131–135. [Google Scholar] [CrossRef]
- Wong, H.S.; Dighe, P.A.; Mezera, V.; Monternier, P.A.; Brand, M.D. Production of superoxide and hydrogen peroxide from specific mitochondrial sites under different bioenergetic conditions. J. Biol. Chem. 2017, 292, 16804–16809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brand, M.D. Mitochondrial generation of superoxide and hydrogen peroxide as the source of mitochondrial redox signaling. Free Radic. Biol. Med. 2016, 100, 14–31. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mesquita, R.D.; Gaviraghi, A.; Gonçalves, R.L.S.; Vannier-Santos, M.A.; Mignaco, J.A.; Fontes, C.F.L.; Machado, L.E.S.F.; Oliveira, M.F. Cytochrome c Oxidase at Full Thrust: Regulation and Biological Consequences to Flying Insects. Cells 2021, 10, 470. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10020470
Mesquita RD, Gaviraghi A, Gonçalves RLS, Vannier-Santos MA, Mignaco JA, Fontes CFL, Machado LESF, Oliveira MF. Cytochrome c Oxidase at Full Thrust: Regulation and Biological Consequences to Flying Insects. Cells. 2021; 10(2):470. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10020470
Chicago/Turabian StyleMesquita, Rafael D., Alessandro Gaviraghi, Renata L.S. Gonçalves, Marcos A. Vannier-Santos, Julio A. Mignaco, Carlos Frederico L. Fontes, Luciana E.S.F. Machado, and Marcus F. Oliveira. 2021. "Cytochrome c Oxidase at Full Thrust: Regulation and Biological Consequences to Flying Insects" Cells 10, no. 2: 470. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10020470