
Preliminary Phytochemical Analysis and Evaluation of the Biological Activity of Leonotis nepetifolia (L.) R. Br Transformed Roots Extracts Obtained through Rhizobium rhizogenes-Mediated Transformation

 , ,
, ,  ,
,  ,
,  , , and
, , and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Leonotis nepetifolia Seeds Surface Sterilization Protocol
2.2. Leonotis nepetifolia Seedlings Inoculation with Rhizobium rhizogenes
2.3. DNA Isolation and Polymerase Chain Reaction
2.4. Plant Materials and Extract Preparation from LNNR and LNTR4 Roots of L. nepetifolia
2.5. Phytochemical Analysis
2.6. Cell Culture
2.7. Determination of Cytotoxic Effect of LNNR and LNTR4 Root Extracts by MTT Assay
2.8. Measurement of Cytoplasmic ROS Levels
2.9. 2,2-Diphenyl-1-Picrylhydrazyl (DPPH) Assay
2.10. DNA Repair and Protective Effect on H2O2-Induced HUVEC Cells after Treatment of LNNR and LNTR4 Root Extracts of L. nepetifolia Assessment by Comet Assay Method
2.11. Assessment of General Toxicity—Brine Shrimp Lethality Bioassay
2.12. Statistical Analysis
3. Results
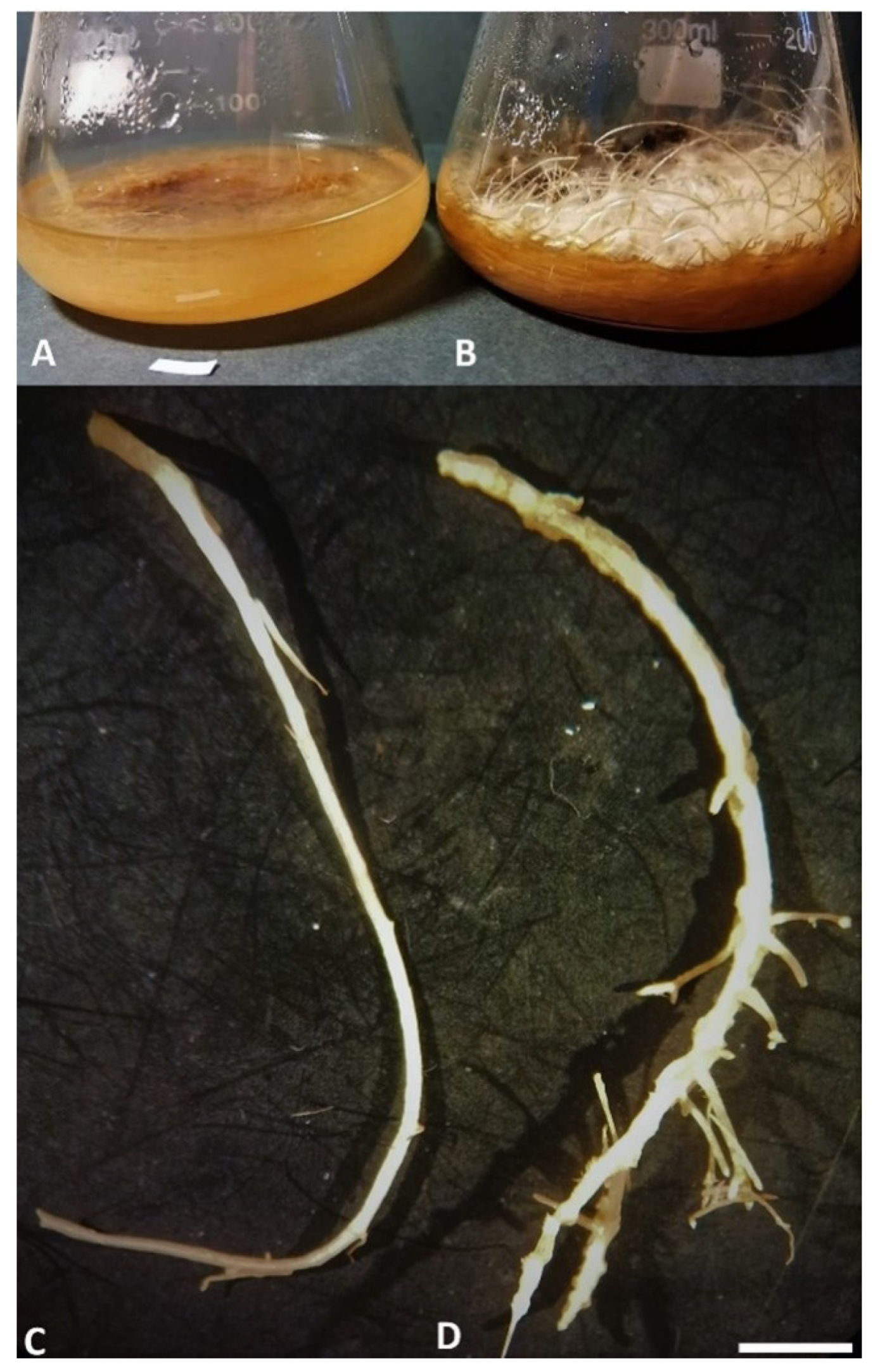
3.1. Hairy Roots Induction
3.2. Confirmation of Transformed Nature of Hairy Roots
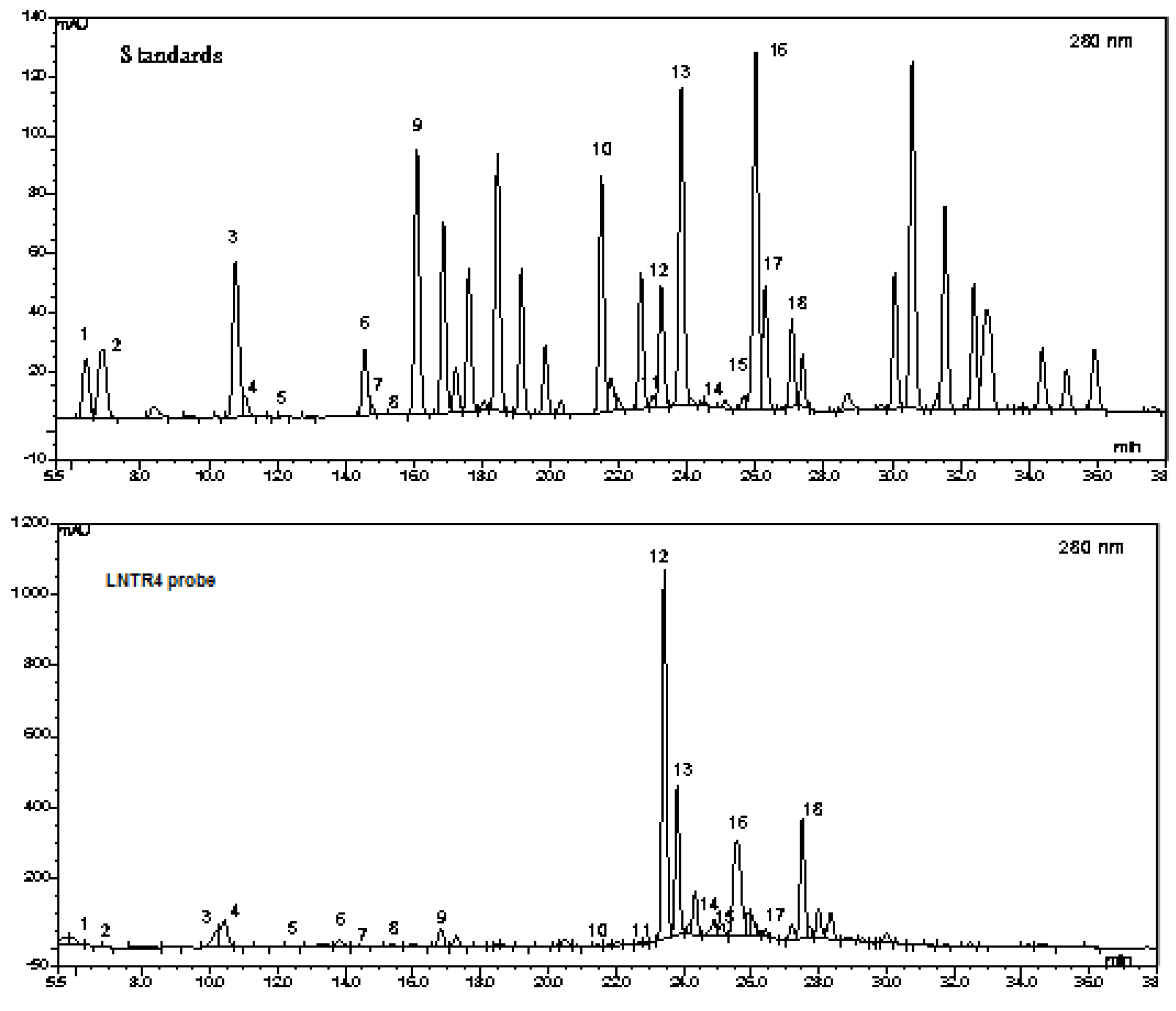
3.3. Phytochemical Analysis of Two Extracts of L. nepetifolia
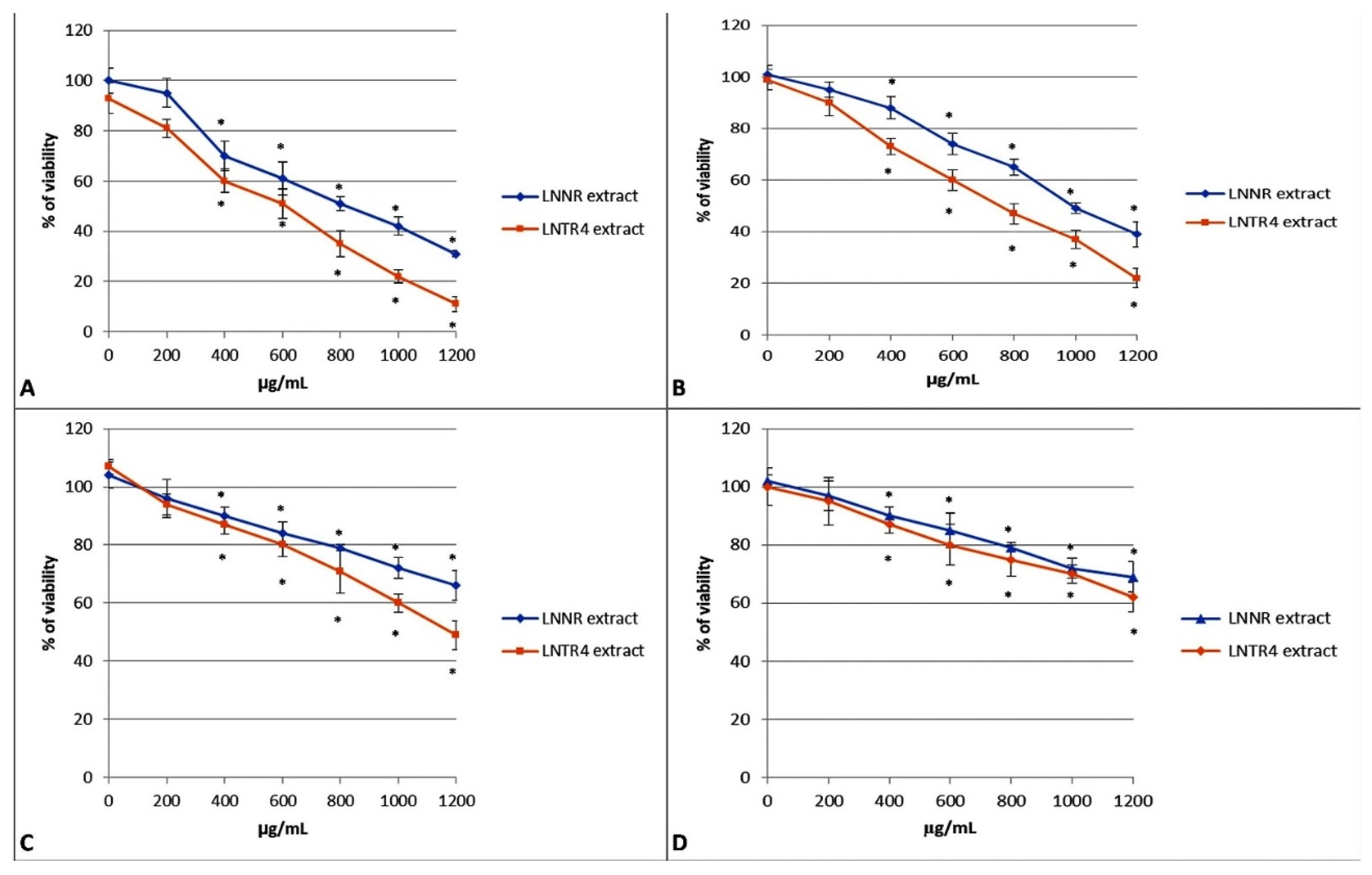
3.4. The Effect of LNNR and LNTR4 Root Extract from L. nepetifolia on Cell Viability
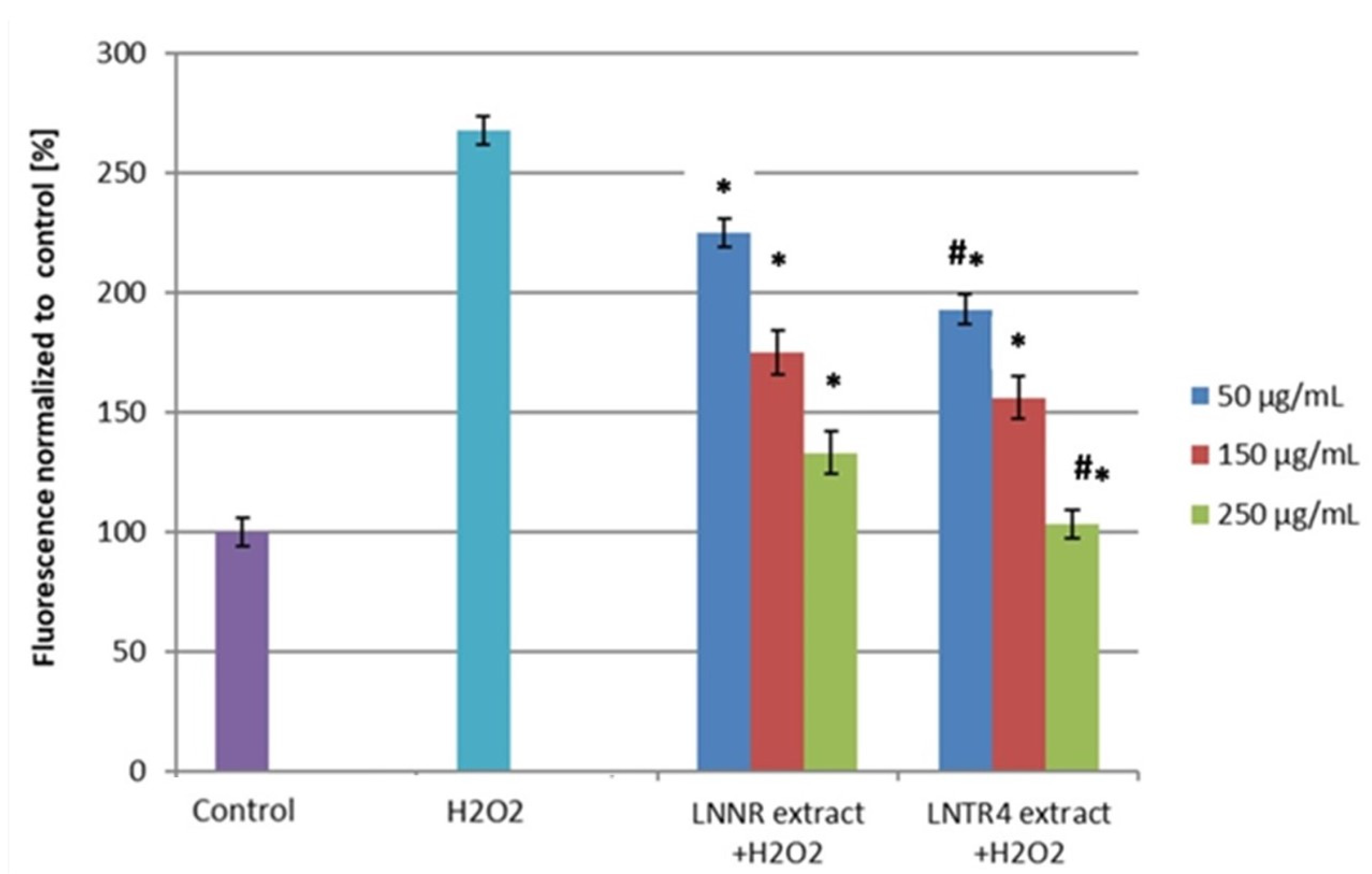
3.5. Effect of LNNR and LNTR4 Root Extracts on ROS Level
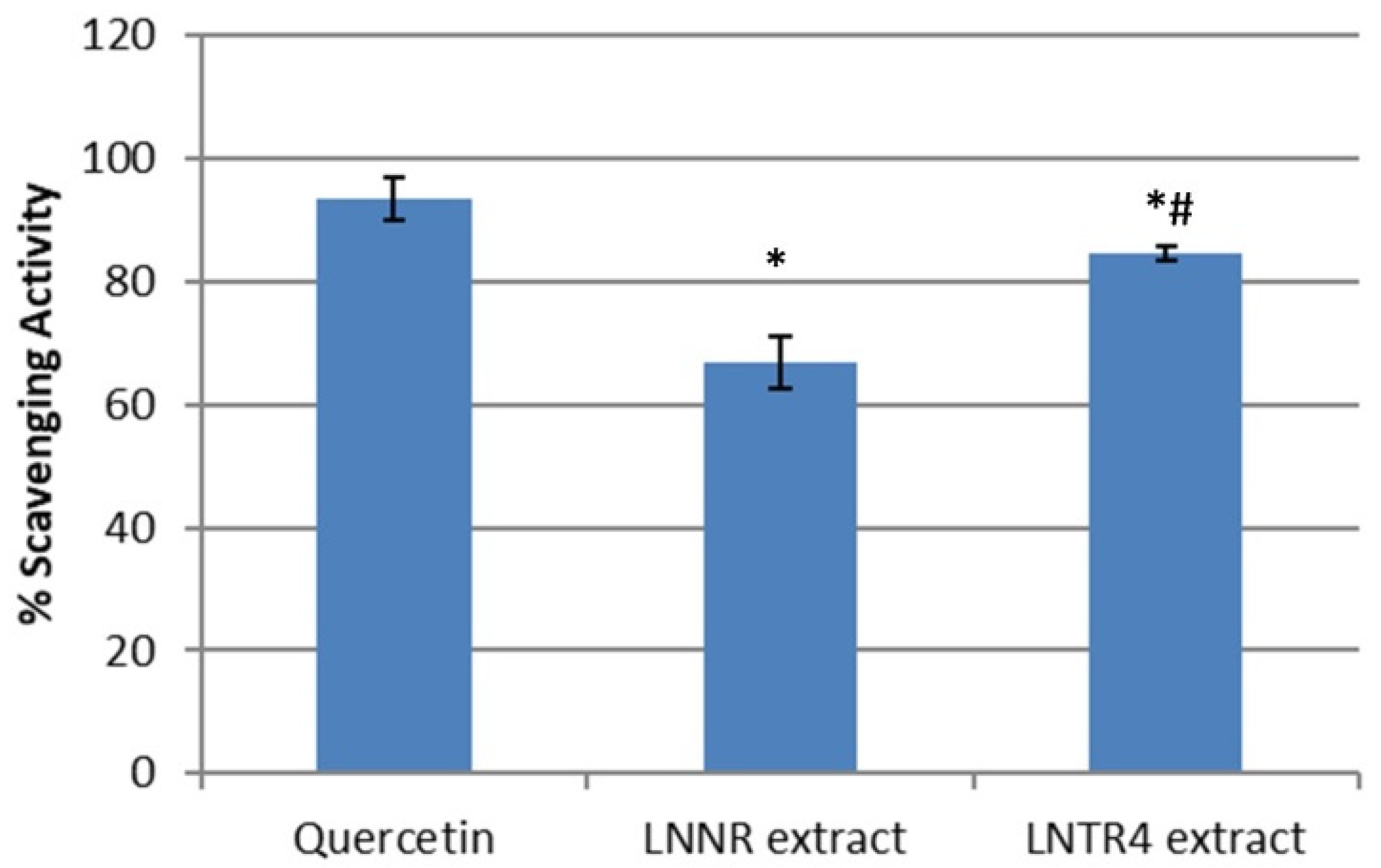
3.6. Antioxidant Activity (DPPH Assay)
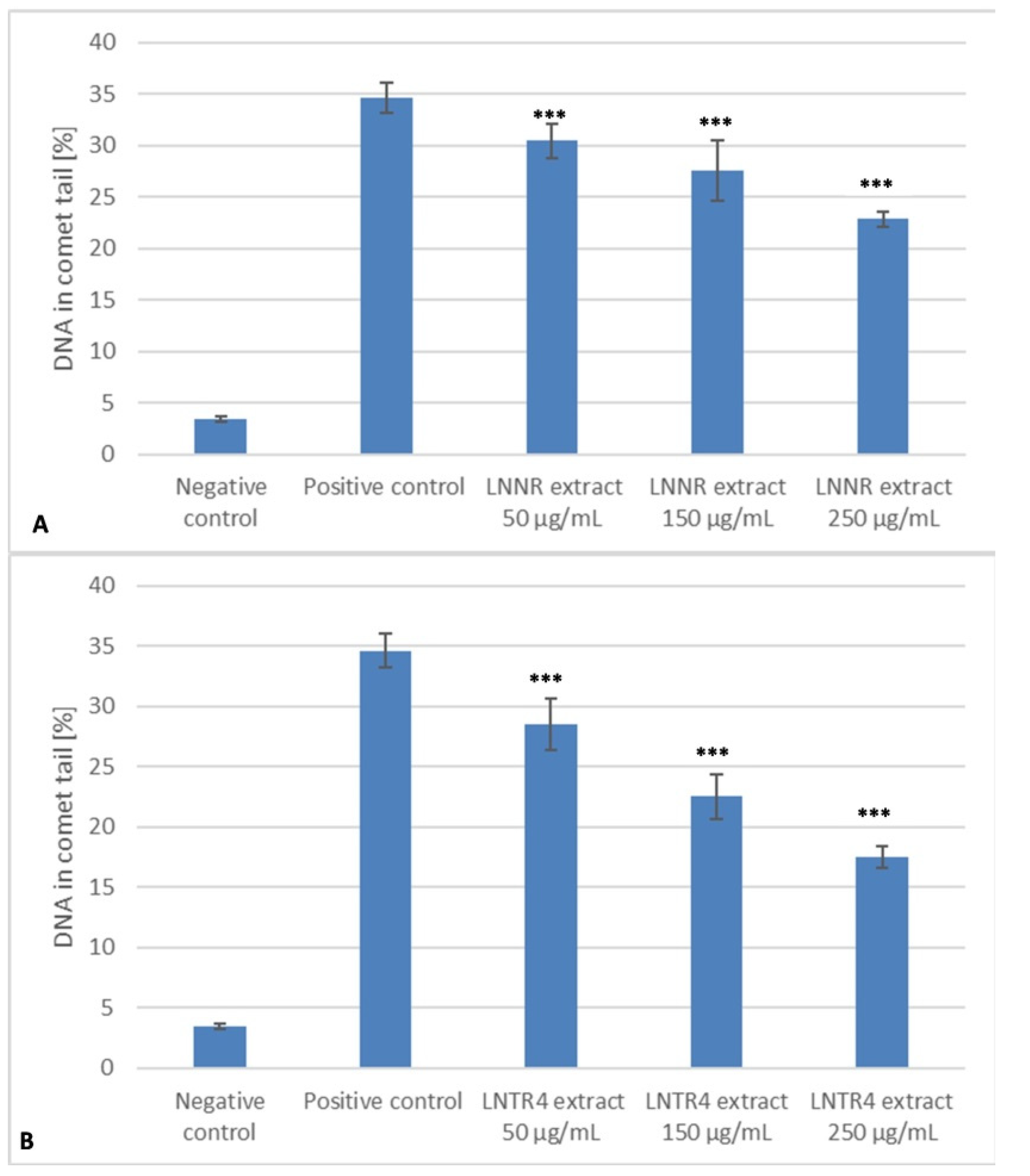
3.7. DNA Repair after Treatment of LNNR and LNTR4 Root Extracts from L. nepetifolia
3.8. DNA Protective Effect after Pretreatment with LNNR and LNTR4 Root Extracts
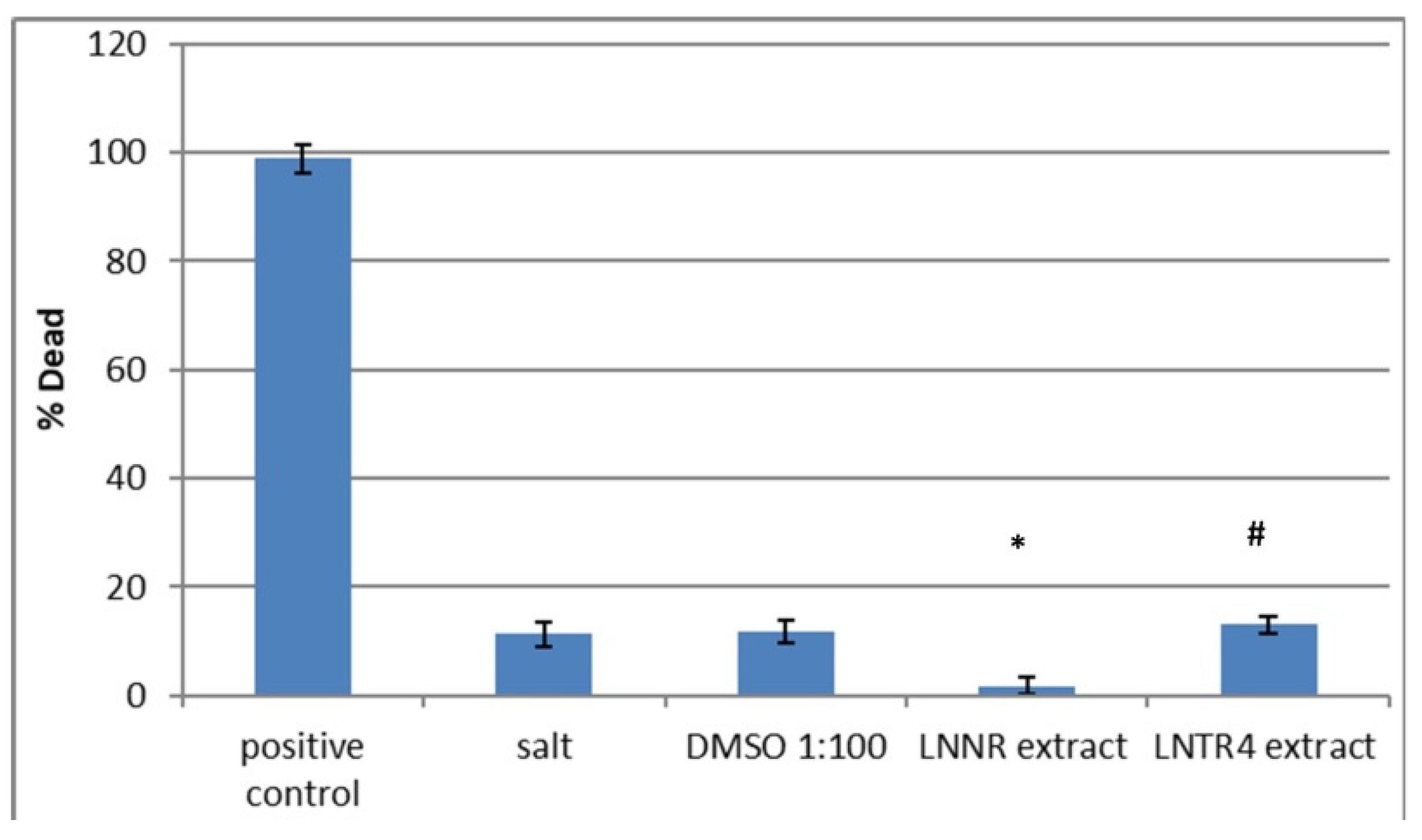
3.9. Brine Shrimp Lethality Bioassay after Treatment of LNNR and LNTR4 Root Extracts
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kowalczyk, T.; Łucka, M.; Szemraj, J.; Sakowicz, T. Hairy roots culture as a source of valuable biopharmaceuticals. Adv. Hyg. Exp. Med. 2016, 70, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Padh, H.; Shrivastava, N. Hairy root cultures: A suitable biological system for studying secondary metabolic pathways in plants. Eng. Life Sci. 2013, 13, 62–75. [Google Scholar] [CrossRef]
- Gutierrez-Valdes, N.; Häkkinen, S.T.; Lemasson, C.; Guillet, M.; Oksman-Caldentey, K.-M.; Ritala, A.; Cardon, F. Hairy Root Cultures—A Versatile Tool With Multiple Applications. Front. Plant Sci. 2020, 11, 33. [Google Scholar] [CrossRef] [PubMed]
- Tian, L. Using Hairy Roots for Production of Valuable Plant Secondary Metabolites. Adv. Biochem. Eng. Biotechnol. 2015, 149, 275–324. [Google Scholar] [CrossRef]
- Srivastava, S.; Srivastava, A.K. Hairy Root Culture for Mass-Production of High-Value Secondary Metabolites. Crit. Rev. Biotechnol. 2008, 27, 29–43. [Google Scholar] [CrossRef] [PubMed]
- Boobalan, S.; Kamalanathan, D. Tailoring enhanced production of aervine in Aerva lanata (L.) Juss. Ex Schult by Agrobacterium rhizogenes- mediated hairy root cultures. Indust. Crop. Prod. 2020, 155, 112814. [Google Scholar] [CrossRef]
- Fathi, R.; Mohebodini, M.; Chamani, E. High efficiency Agrobacterium rhizogenes-mediated genetic transformation in Cichorium intybus L. via removing macronutrients. Indust. Crop. Prod. 2019, 128, 572–580. [Google Scholar] [CrossRef]
- Skała, E.; Kicel, A.; Olszewska, M.A.; Kiss, A.K.; Wysokińska, H. Establishment of Hairy Root Cultures of Rhaponticum carthamoides (Willd.) Iljin for the Production of Biomass and Caffeic Acid Derivatives. BioMed Res. Int. 2015, 2015, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Huang, P.; Xia, L.; Liu, W.; Jiang, R.; Liu, X.; Tang, Q.; Xu, M.; Yu, L.; Tang, Z.; Zeng, J. Hairy root induction and benzylisoquinoline alkaloid production in Macleaya cordata. Sci. Rep. 2018, 8, 1–9. [Google Scholar] [CrossRef]
- Sitarek, P.; Skała, E.; Toma, M.; Wielanek, M.; Szemraj, J.; Nieborowska-Skorska, M.; Kolasa, M.; Skorski, T.; Wysokińska, H.; Śliwiński, T. A preliminary study of apoptosis induction in glioma cells via alteration of the Bax/Bcl-2-p53 axis by transformed and non-transformed root extracts of Leonurus sibiricus L. Tumor Biol. 2016, 37, 8753–8764. [Google Scholar] [CrossRef]
- Sitarek, P.; Skała, E.; Toma, M.; Wielanek, M.; Szemraj, J.; Skorski, T.; Białas, A.J.; Sakowicz, T.; Kowalczyk, T.; Radek, M.; et al. Transformed Root Extract of Leonurus sibiricus Induces Apoptosis through Intrinsic and Extrinsic Pathways in Various Grades of Human Glioma Cells. Pathol. Oncol. Res. 2017, 23, 679–687. [Google Scholar] [CrossRef] [PubMed]
- Pushpan, R.; Nishteswar, K.; Kumari, H. Ethno Medicinal Claims of Leonotis nepetifolia (L.) R. Br: A review. Int. J. Res. Ayurveda Pharm. 2012, 3, 783–785. [Google Scholar] [CrossRef]
- Dhawan, N.G.; Khan, A.S.; Srivastava, P. A General Appraisal of Leonotis nepetifolia (L) R. Br: An Essential Medicinal Plant. Bull. Environ. Pharmacol. Life Sci. 2013, 2, 118–121. [Google Scholar]
- Sobolewska, D.; Paśko, P.; Galanty, A.; Makowska-Wąs, J.; Padło, K.; Wasilak, W. Preliminary phytochemical and biological screening of methanolic and acetone extracts from Leonotis nepetifolia (L.) R. Br. J. Med. Plants Res. 2012, 6, 4582–4585. [Google Scholar] [CrossRef]
- Ashish, T.; Neeraj Sethiya, K.; Mishra, S.H. Preliminary pharmacognostic and phytochemical analysis of “Granthika” (Leonotis nepetaefolia): An Ayurvedic herb. Indian J. Tradit. Knowl. 2011, 10, 682–688. [Google Scholar]
- Makambila-Koubemba, M.C.; Mbatchi, B.; Ardid, D.; Gelot, A.; Henrion, C.; Janisson, R.; Abena, A.A.; Banzouzi, J.T. Pharmacological studies of ten medicinal plants used for analgesic purposes in Congo Brazzaville. Int. J. Pharmacol. 2011, 7, 608–615. [Google Scholar] [CrossRef]
- Gungurthy, J.; Sabbithi, S.; Chaitanya, K.B.; Ravella, A.; Ramesh, C. Antidiabetic activity of Leonotis nepetifolia in alloxan induced diabetic rats. Int. J. Preclin. Pharm. Res. 2013, 4, 5–9. [Google Scholar]
- Damasceno, L.M.O.; Silva, A.L.N.; Santos, R.F.; Feitosa, T.A.; Viana, L.G.F.; Oliveira-Júnior, R.G.; Silva, M.G.; Rolim, L.A.; Araújo, C.S.; Araújo, E.C.C.; et al. Cytotoxic Activity of Chemical Constituents and Essential Oil from the Leaves of Leonotis nepetifolia (Lamiaceae). Rev. Virtual Quim. 2019, 11, 517–528. [Google Scholar] [CrossRef]
- Kadalmani, B.; Niranjana, R.; Natarajan, U.A.; Chandiran, I.S. Antifertility and Antioxidant Activities of Ethanolic Extract of Leonotis nepetifolia in Male Albino Rats. J. Basic Clin. Appl. Health 2020, 3, 115–121. [Google Scholar] [CrossRef]
- Li, J.; Fronczek, F.R.; Ferreira, D.; Burandt, C.L.J.; Setola, V.; Roth, B.L.; Zjawiony, J.K. Bis-spirolabdane diterpenoids from Leonotis nepetaefolia. J. Nat. Prod. 2012, 75, 728–734. [Google Scholar] [CrossRef] [Green Version]
- Powder-George, Y. Secondary Metabolites from the leaves and stems of Leonotis nepetifolia (Lamiaceae). Trop. J. Nat. Prod. Res. 2018, 2, 293–296. [Google Scholar] [CrossRef]
- de Oliveira, A.P.; Borges, I.V.; Pereira, E.C.V.; Feitosa, T.A.; dos Santos, R.F.; de Oliveira-Junior, R.G.; Rolim, L.A.; Cordeiro Viana, L.G.F.; de ARaujo Ribeiro, L.A.; da Conceicao Santos, A.D.; et al. Influence of light intensity, fertilizing and season on the cirsiliol content, a chemical marker of Leonotis nepetifolia (Lamiaceae). Peer J. 2019, 7, e6187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sitarek, P.; Skała, E.; Wysokińska, H.; Wielanek, M.; Szemraj, J.; Toma, M.; Śliwiński, T. The Effect ofLeonurus sibiricusPlant Extracts on Stimulating Repair and Protective Activity against Oxidative DNA Damage in CHO Cells and Content of Phenolic Compounds. Oxidative Med. Cell. Longev. 2016, 2016, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Sitarek, P.; Kowalczyk, T.; Santangelo, S.; Białas, A.J.; Toma, M.; Wieczfinska, J.; Śliwiński, T.; Skała, E. The Extract of Leonurus sibiricus Transgenic Roots with AtPAP1 Transcriptional Factor Induces Apoptosis via DNA Damage and Down Regulation of Selected Epigenetic Factors in Human Cancer Cells. Neurochem. Res. 2018, 43, 1363–1370. [Google Scholar] [CrossRef] [Green Version]
- Rijo, P.D.D.M.; Falé, P.L.; Serralheiro, M.L.; Simões, M.; Gomes, A.; Reis, C. Optimization of medicinal plant extraction methods and their encapsulation through extrusion technology. Meas. J. Int. Meas. Confed. 2014, 58, 249–255. [Google Scholar] [CrossRef]
- Ntungwe, E.; Marçalo, J.; Garcia, C.; Reis, C.; Teodósio, C.; Oliveira, C.; Oliveira, C.; Roberto, A. Biological activity screening of seven Plectranthus species. J. Biomed. Biopharm. Res. 2017, 14, 95–108. [Google Scholar] [CrossRef]
- Singh, N.P.; McCoy, M.T.; Tice, R.R.; Schneider, E.L. A simpletechnique for quantification of low levels of DNA damage in individual cells. Exp. Cell Res. 1988, 175, 184–191. [Google Scholar] [CrossRef] [Green Version]
- Chandra, S.; Chandra, R. Engineering secondary metabolite production in hairy roots. Phytochem. Rev. 2011, 10, 371–395. [Google Scholar] [CrossRef]
- Kumar, V.; Sharma, A.; Prasad, B.C.N.; Gururaj, H.B.; Ravishankar, G.A. Agrobacterium rhizogenes mediated genetic transformation resulting in hairy root formation is enhanced by ultrasonication and acetosyringone treatment. Electron. J. Biotechnol. 2006, 9, 349–357. [Google Scholar] [CrossRef] [Green Version]
- Kowalczyk, T.; Sitarek, P.; Toma, M.; Picot, L.; Wielanek, M.; Skała, E.; Śliwiński, T. An Extract of Transgenic Senna obtusifolia L. Hairy Roots with Overexpression of PgSS1 Gene in Combination with Chemotherapeutic Agent Induces Apoptosis in the Leukemia Cell Line. Biomolecules 2020, 10, 510. [Google Scholar] [CrossRef] [Green Version]
- Gelvin, S.B. Agrobacteriumandplantgenesinvolved int-DNA transfer andintegration. Ann. Rev. Plant Phys. Plant Mol. Biol. 2000, 51, 223–256. [Google Scholar] [CrossRef]
- Brijwal, L.; Tamta, S. Agrobacterium rhizogenes mediated hairy root induction in endangered Berberis aristata DC. SpringerPlus 2015, 4, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saleh, N.M.; Thuc, L. Assessment of hairy roots induction in Solenostemon scutellarioides leaves by different strains of Agrobacterium rhizogenes. Afr. J. Biotechnol. 2009, 8, 3519–3523. [Google Scholar]
- Sitarek, P.; Kowalczyk, T.; Picot, L.; Michalska-Hejduk, D.; Bijak, M.; Białas, A.J.; Wielanek, M.; Śliwiński, T.; Skała, E. Growth of Leonurus sibiricus L. roots with over-expression of AtPAP1 transcriptional factor in closed bioreactor, production of bioactive phenolic compounds and evaluation of their biological activity. Ind. Crop. Prod. 2018, 122, 732–739. [Google Scholar] [CrossRef]
- El-Esawi, M.A.; Elkelish, A.; Elansary, H.O.; Ali, H.M.; Elshikh, M.; Witczak, J.; Ahmad, M. Genetic Transformation and Hairy Root Induction Enhance the Antioxidant Potential of Lactuca serriola L. Oxidative Med. Cell. Longev. 2017, 2017, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.-H.; Vishwakarma, R.K.; Lee, T.-T.; Chan, H.-S.; Tsay, H.-S. Establishment of hairy root lines and analysis of iridoids and secoiridoids in the medicinal plant Gentiana scabra. Bot. Stud. 2014, 55, 17. [Google Scholar] [CrossRef] [Green Version]
- Sitarek, P.; Synowiec, E.; Kowalczyk, T.; Śliwiński, T.; Skała, E. An In Vitro Estimation of the Cytotoxicity and Genotoxicity of Root Extract from Leonurus sibiricus L. Overexpressing AtPAP1 against Different Cancer Cell Lines. Molecules 2018, 23, 2049. [Google Scholar] [CrossRef] [Green Version]
- Veerabadran, U.; Venkatraman, A.; Souprayane, A.; Narayanasamy, M.; Perumal, D.; Elumalai, S.; Sivalingam, S.; Devaraj, V.; Perumal, A. Evaluation of antioxidant potential of leaves of Leonotis nepetifolia and its inhibitory effect on MCF7 and Hep2 cancer cell lines. Asian Pac. J. Trop. Dis. 2013, 3, 103–110. [Google Scholar] [CrossRef]
- Briguglio, G.; Costa, C.; Pollicino, M.; Giambò, F.; Catania, S.; Fenga, C. Polyphenols in cancer prevention: New insights (Review). Int. J. Funct. Nutr. 2020, 1, 9. [Google Scholar] [CrossRef]
- Sharma, A.; Kaur, M.; Katnoria, J.K.; Nagpal, A.K.; Sharma, M.K.A. Polyphenols in Food: Cancer Prevention and Apoptosis Induction. Curr. Med. Chem. 2018, 25, 4740–4757. [Google Scholar] [CrossRef]
- Zhou, Y.; Zheng, J.; Li, Y.; Xu, D.-P.; Li, S.; Chen, Y.-M.; Li, H.-B. Natural Polyphenols for Prevention and Treatment of Cancer. Nutrients 2016, 8, 515. [Google Scholar] [CrossRef]
- Miceli, N.; Cavò, E.; Ragusa, M.; Cacciola, F.; Mondello, L.; Dugo, L.; Acquaviva, R.; Malfa, G.A.; Marino, A.; D’Arrigo, M.; et al. Brassica incana Ten. (Brassicaceae): Phenolic Constituents, Antioxidant and Cytotoxic Properties of the Leaf and Flowering Top Extracts. Molecules 2020, 25, 1461. [Google Scholar] [CrossRef] [Green Version]
- Savikin, K.; Zdunic, G.; Jankovic, T.; Stanojkovic, T.; Juranic, Z.; Menkovic, N. In vitro cytotoxic and antioxidative activity of Cornus mas and Cotinus coggygria. Nat. Prod. Res. 2009, 23, 1731–1739. [Google Scholar] [CrossRef]
- Umamaheswari, M.; Chatterjee, T.K. In vitro antioxidant activities of the fractions of Coccinnia grandis L. leaf extract. Afr J Trad Compl. Altern. Med. 2008, 5, 61–73. [Google Scholar]
- Hunyadi, A. The mechanism(s) of action of antioxidants: From scavenging reactive oxygen/nitrogen species to redox signaling and the generation of bioactive secondary metabolites. Med. Res. Rev. 2019, 39, 2505–2533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, Y.; Ji, L.; Zhang, S.; Xu, L.; Yin, L.; Li, L.; Zhao, Y.; Peng, J. Total flavonoids from Rosa Laevigata Michx fruit attenuates hydrogen peroxide induced injury in human umbilical vein endothelial cells. Food Chem. Toxicol. 2012, 50, 3133–3141. [Google Scholar] [CrossRef] [PubMed]
- Behravan, J.; Mosafa, F.; Soudmand, N.; Taghiabadi, E.; Razavi, B.M.; Karimi, G. Protective Effects of Aqueous and Ethanolic Extracts of Portulaca oleracea L. Aerial Parts on H2O2- Induced DNA Damage in Lymphocytes by Comet Assay. J. Acupunct. Meridian Stud. 2011, 4, 193–197. [Google Scholar] [CrossRef] [Green Version]
- Shojaee, S.; Parhiz, H.; Eshaghi, A.; Emami, S.A.; Asili, J.; Behravan, J.; Mosaffa, F. In vitro protective effects of Scutellaria litwinowii root extract against H2O2-induced DNA damage and cytotoxicity. J. Complement Integr. Med. 2014, 11, 121–127. [Google Scholar] [CrossRef]
- Cheng, N.; Wang, Y.; Cao, W. The Protective Effect of Whole Honey and Phenolic Extract on Oxidative DNA Damage in Mice Lymphocytes Using Comet Assay. Plant Foods Hum. Nutr. 2017, 72, 388–395. [Google Scholar] [CrossRef]
- Grzesik, M.; Naparło, K.; Bartosz, G.; Sadowska-Bartosz, I. Antioxidant properties of catechins: Comparison with other antioxidants. Food Chem. 2018, 241, 480–492. [Google Scholar] [CrossRef] [PubMed]
- Bernatoniene, J.; Kopustinskiene, D.M. The Role of Catechins in Cellular Responses to Oxidative Stress. Molecules 2018, 23, 965. [Google Scholar] [CrossRef] [PubMed] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Acetosyringone | No Acetosyringone | ||||
|---|---|---|---|---|---|
| No. of Seedlings Per Treatment | No. of Seedlings With Hairy Roots | Hairy Roots Induction Efficiency (%) | No. of Seedlings Per Treatment | No. of Seedlings with Hairy Roots | Hairy Roots Induction Efficiency (%) |
| DM method | |||||
| 50 | 42 | 84 | 50 | 28 | 50 |
| IM method | |||||
| 50 | 31 | 62 | 50 | 17 | 34 |
| Compounds | LNNR Extract µg/g DW | LNTR4 Extract µg/g DW | |
|---|---|---|---|
| 1 | Hydroquinone | 126.9 ± 0.298 b | 107.7 ± 0.369 a |
| 2 | Gallic acid | 115.14 ± 0.994 a | 117.78 ± 0.825 a |
| 3 | α-Resorcylic acid | 168.52 ± 3.034 a | 161.62 ± 0.366 a |
| 4 | Catechol | 731.62 ± 0.192 a | 714.2 ± 1.854 a |
| 5 | Protocatechuic acid | 722.36 ± 0.834 b | 657.28 ± 0.259 a |
| 6 | (+)-Catechin | 5464 ± 13.31 a | 6808 ± 10.64 b |
| 7 | 4-Hydroxybenzoic acid | 325.4 ± 0.532 b | 209.2 ± 0.426 a |
| 8 | Gentisic acid | 118.44 ± 0.02 a | 118.78 ± 0.831 a |
| 9 | Chlorogenic acid | 152.22 ± 1.044 a | 264.6 ± 0.410 b |
| 10 | p-Coumaric acid | 2549 ± 3.393 a | 4907 ± 13.73 b |
| 11 | Sinapic acid | 2.382 ± 0.031 a | 14.39 ± 0.223 b |
| 12 | Coumarin | 981.6 ± 1.692 a | 1472.4 ± 2.563 b |
| 13 | m-Coumaric acid | 1508.5 ± 4.452 a | 2048 ± 2.759 b |
| 14 | Rutin | 230.9 ± 3.653 a | 261.6 ± 0.185 b |
| 15 | Ellagic acid | 232.00 ± 0.435 a | 455.4 ± 0.576 b |
| 16 | Hesperidin | 442.00 ± 1.192 a | 603.8 ± 0.419 b |
| 17 | o-Coumaric acid | 198.86 ± 0.673 a | 274.2 ± 0.934 b |
| 18 | Rosmarinic acid | 1844 ± 3.407 a | 2643 ± 2.401 b |
| Total sum of phenols and flavonoids | 15,913.842 ± 39.18 a | 21,838.95 ± 39.77 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kowalczyk, T.; Merecz-Sadowska, A.; Rijo, P.; Isca, V.M.S.; Picot, L.; Wielanek, M.; Śliwiński, T.; Sitarek, P. Preliminary Phytochemical Analysis and Evaluation of the Biological Activity of Leonotis nepetifolia (L.) R. Br Transformed Roots Extracts Obtained through Rhizobium rhizogenes-Mediated Transformation. Cells 2021, 10, 1242. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10051242
Kowalczyk T, Merecz-Sadowska A, Rijo P, Isca VMS, Picot L, Wielanek M, Śliwiński T, Sitarek P. Preliminary Phytochemical Analysis and Evaluation of the Biological Activity of Leonotis nepetifolia (L.) R. Br Transformed Roots Extracts Obtained through Rhizobium rhizogenes-Mediated Transformation. Cells. 2021; 10(5):1242. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10051242
Chicago/Turabian StyleKowalczyk, Tomasz, Anna Merecz-Sadowska, Patricia Rijo, Vera M. S. Isca, Laurent Picot, Marzena Wielanek, Tomasz Śliwiński, and Przemysław Sitarek. 2021. "Preliminary Phytochemical Analysis and Evaluation of the Biological Activity of Leonotis nepetifolia (L.) R. Br Transformed Roots Extracts Obtained through Rhizobium rhizogenes-Mediated Transformation" Cells 10, no. 5: 1242. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10051242