
The RHO Family GTPases: Mechanisms of Regulation and Signaling

Institute of Biochemistry and Molecular Biology II, Medical Faculty of the Heinrich Heine University, Universitätsstrasse 1, Building 22.03.05, 40225 Düsseldorf, Germany
*
Author to whom correspondence should be addressed.
Cells 2021, 10(7), 1831; https://0-doi-org.brum.beds.ac.uk/10.3390/cells10071831
Submission received: 9 April 2021
/
Revised: 13 July 2021
/
Accepted: 13 July 2021
/
Published: 20 July 2021
(This article belongs to the Special Issue Regulation and Function of Small GTPases 2.0)
Abstract
:Much progress has been made toward deciphering RHO GTPase functions, and many studies have convincingly demonstrated that altered signal transduction through RHO GTPases is a recurring theme in the progression of human malignancies. It seems that 20 canonical RHO GTPases are likely regulated by three GDIs, 85 GEFs, and 66 GAPs, and eventually interact with >70 downstream effectors. A recurring theme is the challenge in understanding the molecular determinants of the specificity of these four classes of interacting proteins that, irrespective of their functions, bind to common sites on the surface of RHO GTPases. Identified and structurally verified hotspots as functional determinants specific to RHO GTPase regulation by GDIs, GEFs, and GAPs as well as signaling through effectors are presented, and challenges and future perspectives are discussed.
1. Introduction
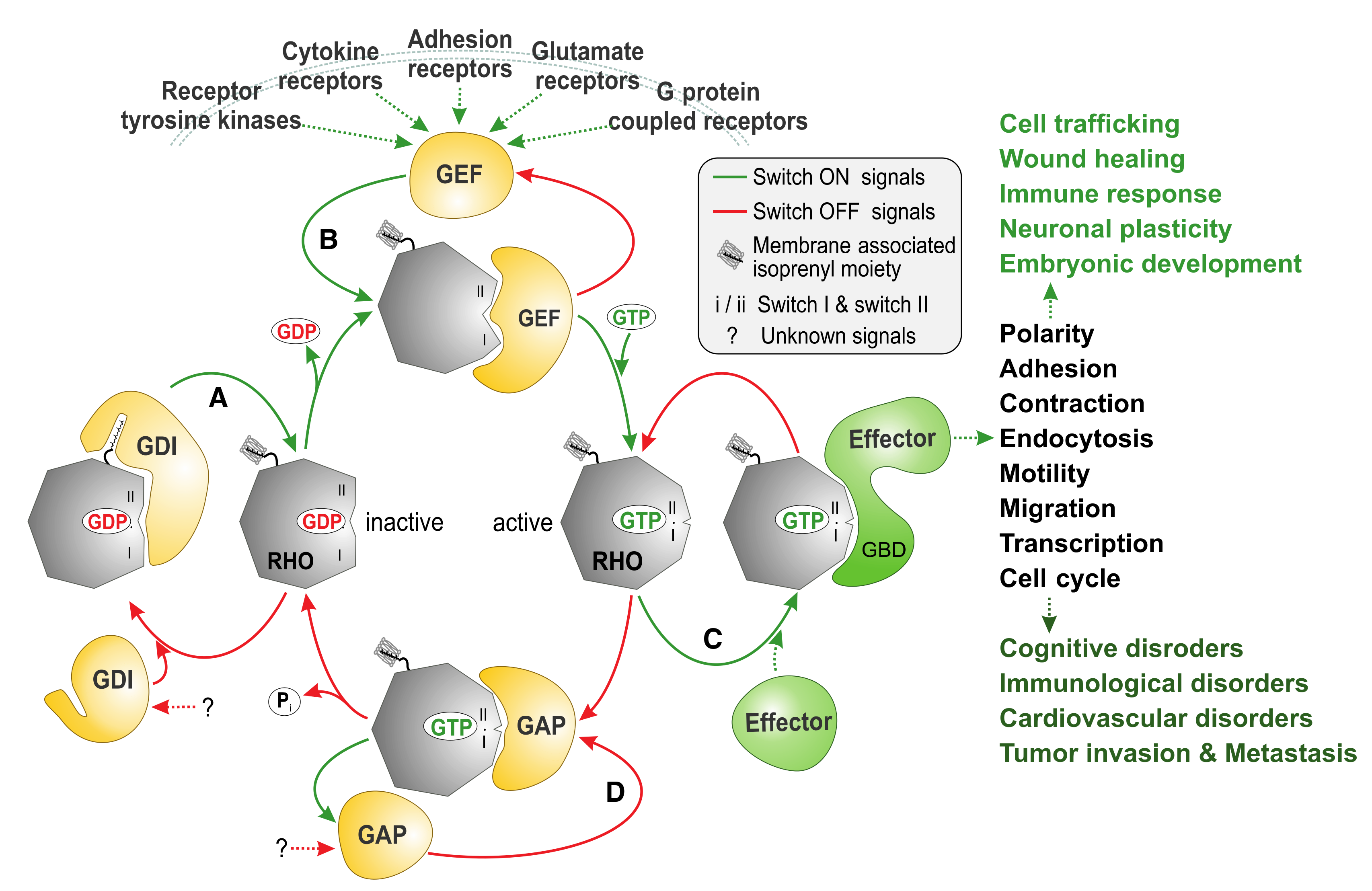
The RHO (RAS homolog) family is an integral part of the RAS superfamily of guanine nucleotide-binding proteins. RHO family proteins are crucial for several reasons: (i) approximately 1% of the human genome encodes proteins that either regulate or are regulated by direct interaction with RHO proteins; (ii) they control almost all fundamental cellular processes in eukaryotes including morphogenesis, polarity, movement, cell division, gene expression, and cytoskeleton reorganization [1]; and (iii) they are associated with a series of human diseases (Figure 1) [2].
The RHO family of proteins functions as molecular switches in the cell and cycle between being in a GDP-bound, inactive state and a GTP-bound, active state [3]. Invaluable insights have been gained by structural and biochemical studies of RHO GTPases and their complexes with interacting partners thus far, increasing our understanding of both how the switch mechanism of the RHO GTPases is regulated and how a RHO GTPase can interact with four classes of structurally and functionally unrelated protein families (Figure 1) [3]. The cellular regulation of this cycle involves guanine nucleotide exchange factors (GEFs), which accelerate intrinsic GDP/GTP exchange, and GTPase-activating proteins (GAPs), which stimulate intrinsic GTP hydrolysis activity [4]. The formation of the active GTP-bound state of the GTPase is accompanied by a conformational change in two regions (known as switch I and II), which provides a platform for the selective interaction with structurally and functionally diverse proteins (the so-called downstream effectors; Table 1) that initiate a network of cytoplasmic and nuclear signaling cascades [5,6]. A prerequisite of RHO protein function is membrane association, which is achieved by isoprenylation, a posttranslational modification. In this respect, RHO proteins are regulated by a third control mechanism that directs their membrane targeting to specific subcellular sites. Specifically, guanine nucleotide dissociation inhibitors (GDIs) bind selectively to prenylated RHO proteins and control their cycle between the cytosol and membrane. Activation of RHO proteins results in their association with effector molecules that subsequently activate a wide variety of downstream signaling cascades, thereby regulating many important physiological and pathophysiological processes in eukaryotic cells [7].
The molecular mechanisms of RHO GTPase regulation have been well characterized, but our understanding of the signal transduction to downstream targets and, most notably, the autoinhibitory mechanisms of GEFs, GAPs, and effectors remain unclear. Very important and challenging, the elucidation of these critical control mechanisms will open new directions for the design of additional therapeutic interventions.
Signaling by these GTPases is controlled by other mechanisms including post-translational modifications such as phosphorylation, ubiquitylation, sumoylation, and acetylation (see for more details [8,9]).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
RHO GTPases, potential effectors and their functions in mammalian cells.
| RHO GTPases | Effector Proteins | Function | Functions and Effects | References |
|---|---|---|---|---|
| RHOA | ROCK I/II | Ser /Thr kinase | Actin myosin contraction, Stress fiber formation | [10,11] |
| Citron kinase | Ser /Thr kinase | Cytokinesis | [12] | |
| MBS | Phosphatase subunit | MLC inactivation | [13] | |
| DIA 1/2 | Formin-like proteins | Actin polymerization | [10] | |
| RHOB | Integrin β1 | Cell surface receptor | Cell adhesion and migration | [14] |
| RHOC | FMNL3 | Formin like proteins | Migration, Invasion | [15] |
| RHOH | Kaiso | Transcription factor | TCR activation | [16] |
| RAC1 | PAK1/2/3 * | Ser /Thr kinase | JNK activation, Actin filament stabilization | [17] |
| MLK 2/3 * | Ser /Thr kinase | JNK activation | [18,19] | |
| WAVE | Scaffold | Actin organization | [20] | |
| p70 S6 kinase * | Ser /Thr kinase | Translation regulation | [21] | |
| IQGAP1/2 * | Scaffold | Actin/cell-cell contacts | [22,23] | |
| MEKK1/4 * | Ser /Thr kinase | JNK activation | [24] | |
| POR1 | Scaffold | Actin organization | [25] | |
| p67phox* | Scaffold | ROS generation | [26] | |
| PI3 kinase | Lipid kinase | PIP3 levels | [27] | |
| DAG kinase | Lipid kinase | PA levels | [28,29] | |
| PLCβ2 * | Lipase | DAG and IP3 levels | [30] | |
| RAC1B | p120ctn | Catenin | Cellular transformation | [31] |
| RAC2 | LFA-1 | Cell surface receptor | B cell adhesion | [32] |
| RAC3 | GIT1 | ARF GAP and scaffold | Regulation of cell adhesion and differentiation | [33] |
| RHOG | Kinectin | Kinesin receptor | Microtubule dependent transport | [34] |
| CDC42 | N-WASP | Scaffold | Actin organization | [35] |
| PAK4 | Ser/Thr kinase | Actin organization | [36] | |
| MRCKα/β | Ser/Thr kinase | Actin organization | [37] | |
| TCL | GIT-PIX complex | Scaffold | Stabilization of focal adhesion | [38,39] |
| RHOD | Plexin A1/B1 | Semaphorin co-receptor | Growth cone formation | [40] |
| RIF | DIA 1/2 | Formin-like proteins | Actin organization | [40,41] |
| RND1 | Stathmin2 | Neuronal growth associated proteins | Microtubule depolymerization, Neurite extension | [42] |
| RND2 | Rapostlin | Formin-binding protein | Neurite branching | [43] |
| RND3 | Socius | Scaffold | Loss of stress fibers | [44] |
| ROCKI | Ser/Thr kinase | Actomyosin contractility | [45,46] |
* Proteins shown with an asterisk are shared effectors for both RAC1 and CDC42.
2. The RHO Family and the Molecular Switch Mechanism
Members of the RHO family have emerged as key regulatory molecules that couple changes in the extracellular environment to intracellular signal transduction pathways. To date, 20 canonical members of the RHO family have been identified in humans and can be categorized into distinct subfamilies based on their sequence homology: RHO (RHOA, RHOB, and RHOC); RAC (RAC1, RAC1B, RAC2, RAC3, and RHOG); CDC42 (CDC42, G25K, TC10, TCL, WRCH1, and WRCH2); RHOD (RHOD, RIF); RND (RND1, RND2, and RND3); and RHOH [47].
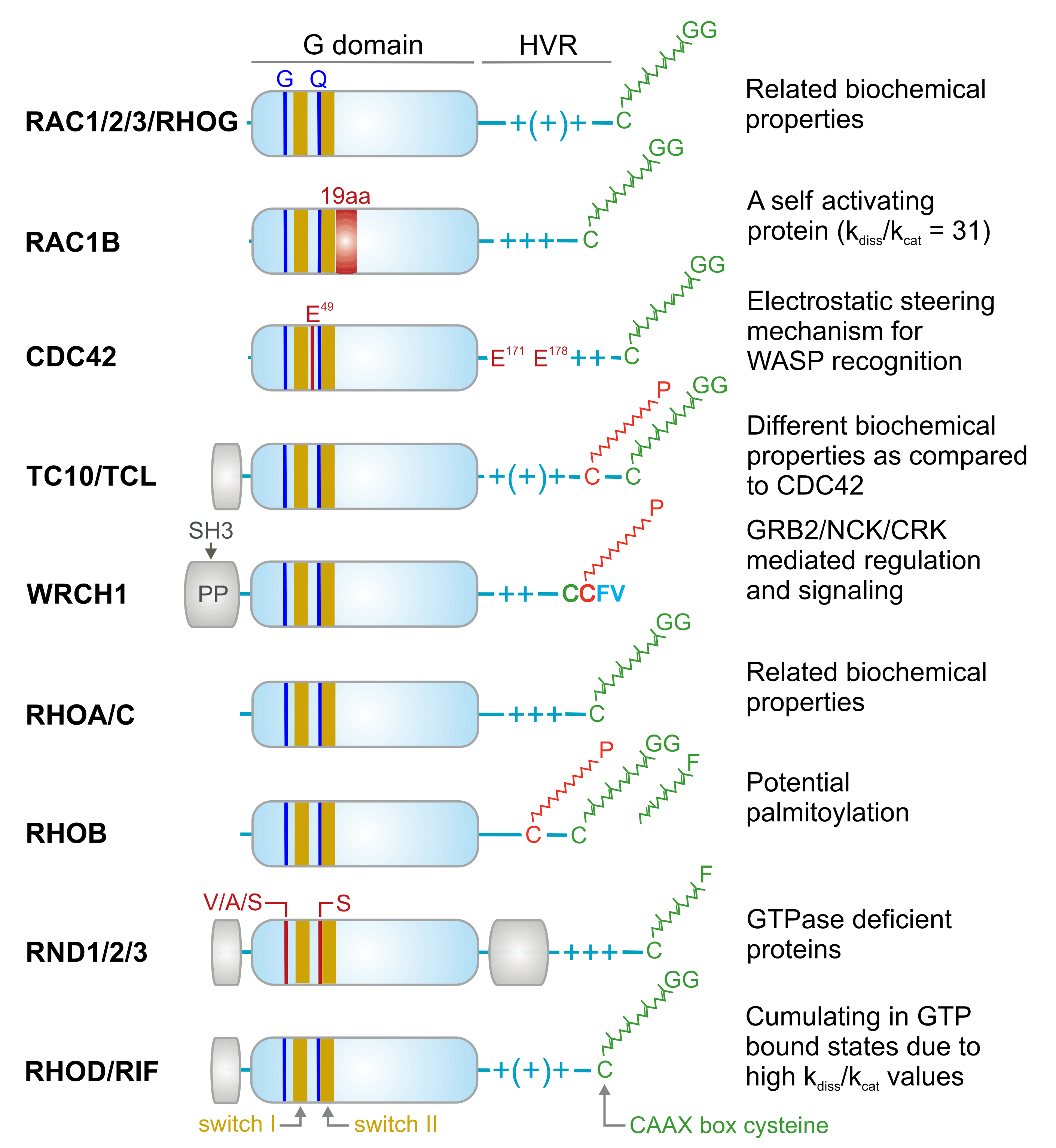
RHO family proteins are approximately 21–25 kDa in size. They typically contain a conserved GDP-/GTP-binding domain, called the G domain, and a C-terminal hypervariable region (HVR) ending with a consensus sequence known as CAAX (C is cysteine, A is any aliphatic amino acid, and X is any amino acid) (Figure 2). The G domain consists of five conserved sequence motifs (G1 to G5) that are involved in nucleotide binding and hydrolysis [48]. In the cycle between the inactive and active states, at least two regions of the protein, switch I (G2) and switch II (G3), undergo structural rearrangement and transmit an “OFF” to “ON” signal [3]. Subcellular localization, which is known to be critical for the biological activity of RHO proteins, is achieved through a series of posttranslational modifications at a cysteine residue in the CAAX motif including isoprenylation (geranylgeranyl or farnesyl), endoproteolysis, and carboxyl methylation [49].
Figure 2.
Domains, signature motifs, and post-translational modification of RHO GTPases. RHO GTPases contain a highly conserved G domain, which is responsible for GDP/GTP binding and GTP hydrolysis. Switch I and switch II regions are the consensus binding sites for GEFs, GAPs, GDIs, and effectors, and undergo conformational changes upon the nucleotide exchange and hydrolysis [3]. All members of the RHO family contain conserved glycine 12 (G) and glutamine 61 (Q; RAC1 numbering), except for the RND proteins, which contain, among other deviations, other residues at these positions. This is why RND proteins constantly remain in the GTP bound state [50]. Other signatures are, for example, a 19-amino acid insertion next to the switch II region in RAC1B with drastic biochemical consequences [51], and glutamic acids (E) in CDC42 crucial for a selective WASP interaction [52]. Some members have amino acid insertion outside the G domain (yellow boxes) with yet unknown properties. The N-terminal insertion in WRCH1 contains proline-rich motifs responsible for interaction with SH3-contining adaptor proteins [53]. Most members have comparable biochemical properties such as nucleotide binding, exchange, and hydrolysis. In contrast to most members, which end up under resting conditions in an inactive GDP-bound state, RAC1B, RHOD, and RIF cumulate in the GTP-bound state due to a faster intrinsic nucleotide exchange reaction (kdis) compared to the intrinsic GTP hydrolysis reaction (kcat) [54]. The C-terminal hypervariable region (HVR) contains the terminal CAAX box, which undergoes posttranslational modification by geranylgeranylation (GG) or alternatively farnesylation (F) in the case of RHOB and the RND proteins at the conserved cysteine (green). Additional modification by a palmitoyl (P) moiety has been reported for RHOB, and the CDC42-related proteins TC10, TCL, and WRCH1. These modifications lead to the membrane anchorage of the members, a process that is stabilized and potentiated through variable numbers of positively charged arginine and lysine residues (+).
Figure 2.
Domains, signature motifs, and post-translational modification of RHO GTPases. RHO GTPases contain a highly conserved G domain, which is responsible for GDP/GTP binding and GTP hydrolysis. Switch I and switch II regions are the consensus binding sites for GEFs, GAPs, GDIs, and effectors, and undergo conformational changes upon the nucleotide exchange and hydrolysis [3]. All members of the RHO family contain conserved glycine 12 (G) and glutamine 61 (Q; RAC1 numbering), except for the RND proteins, which contain, among other deviations, other residues at these positions. This is why RND proteins constantly remain in the GTP bound state [50]. Other signatures are, for example, a 19-amino acid insertion next to the switch II region in RAC1B with drastic biochemical consequences [51], and glutamic acids (E) in CDC42 crucial for a selective WASP interaction [52]. Some members have amino acid insertion outside the G domain (yellow boxes) with yet unknown properties. The N-terminal insertion in WRCH1 contains proline-rich motifs responsible for interaction with SH3-contining adaptor proteins [53]. Most members have comparable biochemical properties such as nucleotide binding, exchange, and hydrolysis. In contrast to most members, which end up under resting conditions in an inactive GDP-bound state, RAC1B, RHOD, and RIF cumulate in the GTP-bound state due to a faster intrinsic nucleotide exchange reaction (kdis) compared to the intrinsic GTP hydrolysis reaction (kcat) [54]. The C-terminal hypervariable region (HVR) contains the terminal CAAX box, which undergoes posttranslational modification by geranylgeranylation (GG) or alternatively farnesylation (F) in the case of RHOB and the RND proteins at the conserved cysteine (green). Additional modification by a palmitoyl (P) moiety has been reported for RHOB, and the CDC42-related proteins TC10, TCL, and WRCH1. These modifications lead to the membrane anchorage of the members, a process that is stabilized and potentiated through variable numbers of positively charged arginine and lysine residues (+).

Once an isoprenoid moiety is added to CAAX, a RHO protein is translocated to the endoplasmic reticulum, where RCE1 cleaves the AAX tripeptide tail, and then, RHO undergoes carboxymethylation by ICMT [55] RHO proteins can also be phosphorylated, which can affect their association with their regulators or effectors or influence their membrane stability [56,57,58].
A characteristic region of RHO family GTPases is the insert helix (amino acids 124–136, RHOA numbering), which may play a role in effector activation and downstream processes [59].
Although the majority of the RHO family proteins are remarkably inefficient GTP-hydrolyzing enzymes, in quiescent cells, they accumulate in an inactive state because GTP hydrolysis by RHO proteins is, on average, two orders of magnitude faster than GDP/GTP exchange [47]. These different intrinsic activities provide the basis for a two-state molecular switch mechanism, which greatly depends on the regulatory functions of GEFs and GAPs. Eleven of the 20 RHO family members possess classical molecular switches, namely, RHOA, RHOB, RHOC, RAC1, RAC2, RAC3, RHOG, CDC42, G25K, TC10, and TCL [47].
Atypical RHO family members including RND1, RND2, RND3, RAC1B, RHOH, WRCH1, RHOD, and RIF have been proposed to accumulate in the GTP-bound form in cells [47]. RND1, RND2, RND3, and RHOH constitute a completely distinct group of proteins within the RHO family (Figure 2) [60], as they do not share several essential amino acids including Gly-12 (RAC1 numbering) in the G1 motif (a phosphate-binding loop or P-loop) and Gln-61 in the G3 motif or switch II region, which are critical in GTP hydrolysis. Thus, they can be considered GTPase-deficient RHO-related GTP-binding proteins [61]. RHOD and RIF are involved in the regulation of actin dynamics [41] and exhibit much faster nucleotide exchange than GTP hydrolysis. WRCH1, a CDC42-like protein that has been reported to be a fast-cycling protein, resembles RAC1B, RHOD, and RIF in this regard (Figure 2) [47]. These atypical members do not possess the classical switch mechanism and, therefore, may be regulated through other mechanisms.
3. Regulation of RHO Family GTPases
3.1. Guanine Nucleotide Dissociation Inhibitors (GDIs)
Despite the vast number of RHOGEFs and RHOGAPs, only three GDIs exist in the human genome. The RHOGDI family includes ubiquitously expressed GDI1 (or RHOGDIα) [62]; GDI2 (GDIβ, LY-GDI or D4-GDI), mainly in hematopoietic tissue [63]; and GDI3 (or GDIγ), which is usually expressed in human cerebral, lung, and pancreatic tissue [64]. An N-terminal extension that anchors GDI3 to the membrane of Golgi vesicles distinguishes this isoform from the others [65].
Several studies in recent decades have provided information about the structure and function of GDIs and proposed that they act as shuttles for RHO GTPase [8,66,67,68]. The shuttling process is initiated by the release of RHO GTPases from donor membranes, the formation of inhibitory cytosolic GDI-RHO GTPase complexes, and the delivery of RHO GTPases to the membranes of subcellular compartments [66,67].
It has been demonstrated that the isoprenylation process in cells can be regulated by GDIs [69]. GDI mediates the release of RHO GTPases from the membrane, maintains them in an inactivated state, and safeguards them against degradation or nonspecific activation by RHOGEFs [25,29,30]. Different structural studies have revealed two sites of GDI and RHO GTPase interaction [70,71,72,73,74]. First, an N-terminal regulatory arm of GDI binds to the switch region of RHO GTPases and inhibits GDP dissociation and GTP hydrolysis. Second, the N-terminus of GDI attracts the positively charged RHO hypervariable region, which is engaged with negatively charged phospholipids of the membrane and initiates the insertion of the geranylgeranyl moiety on the RHO GTPases into a hydrophobic pocket in the GDI molecule, leading to membrane release [75].
3.2. Guanine Nucleotide Exchange Factors (GEFs)
GEFs are able to selectively bind to their respective RHO proteins and accelerate the exchange of tightly bound GDP for GTP [8]. Typically, GEFs profoundly reduce the affinity of RHO proteins for GDP, leading to its displacement from GDP and subsequent association with GTP [76,77]. This reaction involves several stages including an intermediate state in which the GEF is in the complex with the nucleotide-free RHO protein. This intermediate does not accumulate in the cell and rapidly dissociates because of the high intracellular GTP concentration, leading to the formation of the active RHO-GTP complex. The main principle driving this mechanism is based on the binding affinity of nucleotide-free RHO protein being significantly greater for GTP than for GEF proteins [76,78]. Cellular activation of RHO proteins and their cellular signaling can be selectively uncoupled from GEFs through the overexpression of dominant-negative mutants of RHO proteins (e.g., threonine 19 in RHOA is replaced with asparagine) [79]. Dominant-negative mutants form a tight complex with their cognate GEFs, preventing them from activating endogenous RHO proteins. RHOGEFs are classified into two distinct families: DBL homology (DH) domain-containing proteins, and dedicator of cytokinesis (DOCK) proteins [80,81].
3.2.1. DBL Family GEFs
RHOGEFs of the diffuse B-cell lymphoma (DBL) family directly activate the proteins of the RHO family [82]. The prototype of this GEF family is the DBL protein, which was isolated as an oncogenic product from diffuse B-cell lymphoma cells in an oncogene screen [83] and was later reported to act on CDC42 [84]. Human DBL family proteins have recently been grouped into functionally distinct categories based on both their catalytic efficiencies and their sequence–structure relationship [47]. Members of the DBL family are characterized by a unique DBL homology (DH) domain [85,86,87,88].
The DH domain is a highly efficient catalytic machine [80] that is able to accelerate the nucleotide exchange of RHO proteins by as much as 107-fold. The DH domain is often followed by a pleckstrin homology (PH) domain, indicating its essential and conserved function. A model for PH domain-assisted nucleotide exchange has been proposed for some GEFs such as DBL, DBS, and TRIO [80]. Thus, the PH domain serves multiple roles in signaling events by anchoring GEFs to the membrane (via phosphoinositides) and directing them toward their respective GTPase partners, which are on the membrane [80].
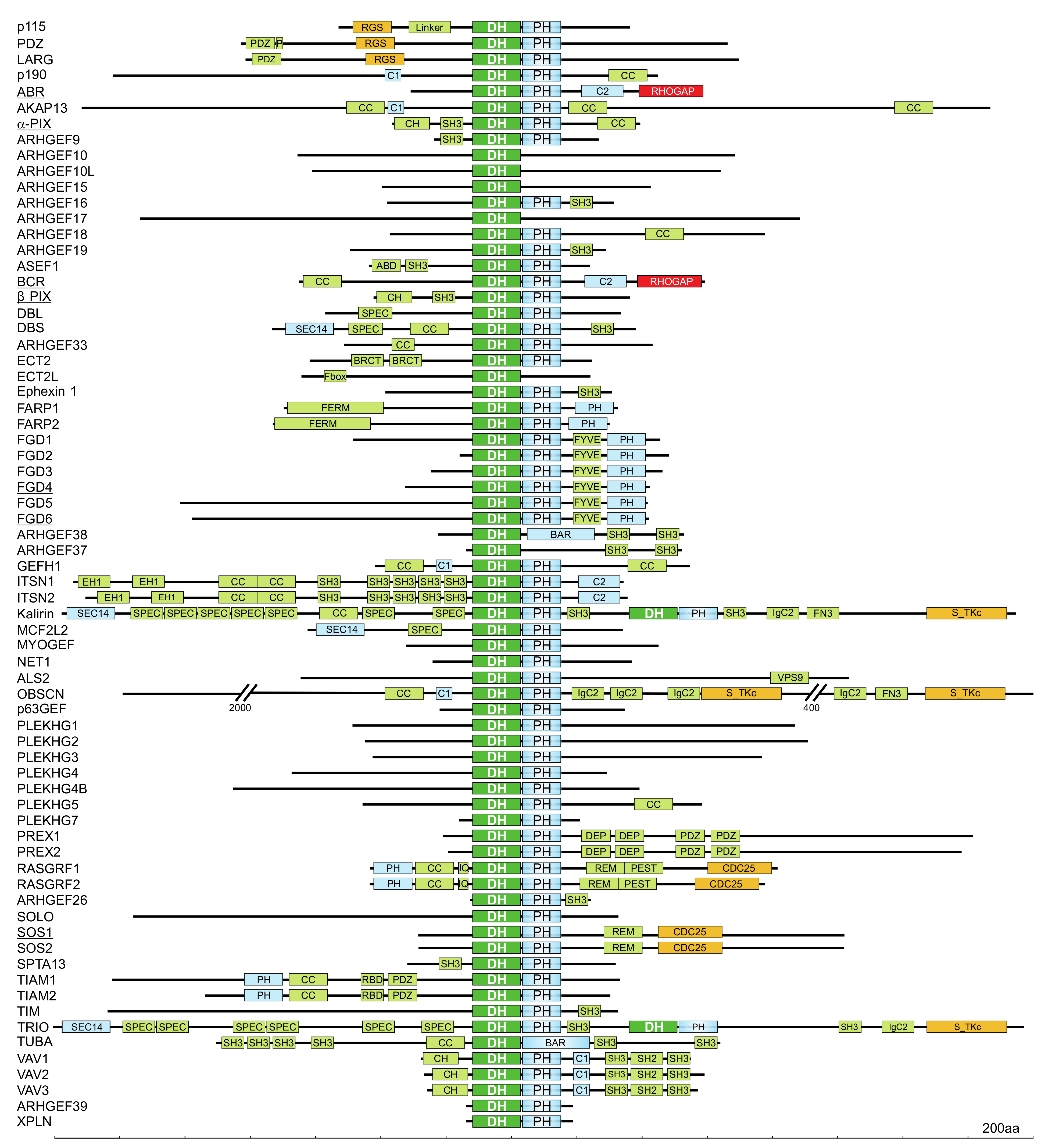
Through a search for DH domain-containing proteins in the human genome, 74 DBL proteins have been identified (Figure 3) [47]. Interestingly, nine of these DBL proteins lack the C-terminal tandem PH domain, and three of these proteins contain a membrane bending and tubulating BAR (BIN/amphiphysin/RVS) domain, and seven of 20 investigated DBL proteins do not exhibit any GEF activity (Figure 3) [47]. In addition to the DH-PH tandem motif, DBL family proteins are highly diverse and contain additional domains with different functions (Figure 3) including SH2, SH3, CH, RGS, PDZ, and/or IQ domains, which enable their interaction with other proteins; BAR, PH FYVE, C1, and C2 domains, which enable their interaction with membrane lipids; and other functional domains such as Ser/Thr kinase, RASGEF, RHOGAP, and RANGEF [82]. These additional domains have been implicated in autoregulation, subcellular localization, and connection to upstream signaling molecules [40,49,50]. Spatiotemporal regulation of DBL proteins has been suggested as a mechanism that specifically initiates the activation of substrate RHO proteins and controls a broad spectrum of normal and pathological cellular functions [89]. Thus, it is evident that members of the DBL protein family are attractive therapeutic targets for a variety of diseases [90,91].
3.2.2. Structural and Functional Characteristics of the DH domain
The DH domain is the signature of DBL family proteins. The catalytic guanine nucleotide exchange activity of DBL family proteins is realized entirely within the DH domain, which is not only sufficient for catalytic activity but also critical for substrate specificity [47,92]. The catalytic DH domain consists of approximately 200 residues, and as determined by x-ray and NMR analyses of the DH domain in several DBL proteins, it is composed of a unique extended bundle of 10–15 alpha helices [93]. This helical fold is mainly composed of three conserved regions, CR1, CR2, and CR3, each of which is 10–30 residues long and forms separate alpha helices that are packed together [45,53]. The CR1 and CR3 regions are solvent exposed until complexed with RHO proteins [47]. Except for these three conserved regions (CR1, CR2, and CR3) in DH domains, DBL family members share little homology with each other [87].
3.2.3. The Tandem PH Domain in DBL Proteins
In the majority of DBL family proteins, the catalytic DH domain is followed by a PH domain consisting of approximately 100 residues (Figure 3), and even though the identity of the PH domain among members of the DBL family is less than 20%, the PH-domain containing DBL proteins share a similar three-dimensional structure with two orthogonal antiparallel β-sheets and a folded C-terminal α-helix that cover one end [94,95]. The PH domain was originally identified in a number of cytoplasmic signaling proteins that displayed homology with a region repeated in pleckstrin [96,97]. The DH-PH tandem is a signature motif of the DBL family, indicating that the PH domain has an essential and conserved function [85,88]. The tandem PH domain can act as a “membrane-targeting device” due to its ability to bind phosphoinositides [98]. It can also bind directly to RHO proteins and potentiate the DH-catalyzed nucleotide exchange reaction [92,93]. In contrast, the PH domains have been shown to bind and inhibit the activity of the DH domain [99,100]. In addition to its membrane-targeting properties, emerging evidence suggests that the PH domain may also play important regulatory roles by serving as a protein–protein interaction module [101].
3.2.4. A Plethora of DBL Family Proteins
It is evident that DBL family proteins are more abundant and varied in cells than RHO family proteins. To date, 74 DBL proteins have been reported in humans, and they are classified into different subfamilies: 46 DBL proteins are monospecific for RHO-, RAC-, and CDC42-selective proteins, five are bispecific for RHO- and CDC42-selective proteins, and six are oligospecific for all three RHO protein subgroups [47]. Since there are many more DBL proteins and many of them can activate more than one RHO protein, the activation of RHO proteins catalyzed by DBL family proteins constitutes a level of regulation in which the signaling pathways can converge or diverge toward one or more RHO proteins [7]. This multifunctionality suggests that at least one representative of each DBL subfamily is expressed in all mammalian cells, but they may act at distinct subcellular sites.
3.3. DOCK Family of RHOGEFs
The 11 members of the DOCK family can be categorized into four subfamilies: DOCK-A, DOCK-B, DOCK-C, and DOCK-D [81,102]. DOCK GEFs have two conserved domains: lipid-binding DOCK homology region 1 (DHR-1), which facilitates DOCK localization to membrane compartments, and catalytic DOCK homology region 2 (DHR-2), which induces the GDP-GTP exchange reaction [81,102,103,104]. It has been proposed that DOCK GEFs activate RAC1 and CDC42 proteins, but not other RHO proteins [105,106].
DOCK proteins orchestrate important processes in brain development including neuron, microglial, and Schwann cell development and functions [102,107]. DOCK2 and DOCK8 play significant roles in immune responses such as the chemotactic responses of T cells and B cells, ROS production in neutrophils, and migration of mature dendritic cells [81]. Li et al. demonstrated that DOCK1 forms a complex with ELMO1, RAC1, RAC2, and Gαi2, which initiates actin polymerization in breast cancer cells [108].
DOCK2 has been indicated to increase amyloid beta plaque formation, which makes this protein a potential Alzheimer’s therapeutic target [109,110]. Janssen et al. showed that in T cells, DOCK8 can form a complex with WASP and ARP2/3 and link TCR to the actin cytoskeleton to form a synapse for T cell responses [111].
Overall, DOCK GEFs play pivotal roles in different biological processes that can be dependent or independent of their GEF activity.
3.4. GTPase-Activating Proteins (GAPs)
Hydrolysis of bound GTP is the timing mechanism that terminates signal transduction of RHO family proteins and enables their return to an inactive, GDP-bound state [87]. The intrinsic GTPase reaction is usually slow but can be stimulated to accelerate by several orders of magnitude through interaction with RHO-specific GAPs [112,113]. The RHOGAP family is identified by the presence of a conserved catalytic GAP domain that is sufficient for engaging RHO proteins and mediating accelerated catalysis [114,115]. The GAP domain inserts a conserved arginine residue, termed an “arginine finger”, into the GTP-binding site of the cognate RHO protein to stabilize the transition state and catalyze the GTPase reaction [74,76,77]. This mechanism is similar to that of other small GTP-binding proteins including RAS, RAB, and ARF, although the sequence and folding of the respective GAP families differ from other GTP-binding proteins [115,116]. Masking the catalytic arginine finger is an elegant mechanism for the inhibition of GAP activity. This action has also been recently discovered in the tumor suppressor protein DLC1, an RHOGAP, which is competitively and selectively inhibited by the SH3 domain in p120RASGAP [117,118].
The first RHOGAP discovered, p50RHOGAP, was identified through a biochemical analysis of human spleen cell extracts in the presence of recombinant RHOA [119]. The majority of RHOGAP family members typically harbor several other functional domains and motifs that are implicated in tight regulation and membrane targeting (Figure 4) [74,82,83]. Numerous mechanisms have been shown to affect the specificity and catalytic activity of RHOGAPs (e.g., intramolecular autoinhibition [120], posttranslational modification [121], and regulation by interaction with lipid membranes [122] and proteins [118]).
RHOGAP insensitivity has been frequently analyzed through the substitution of either amino acid that is critical for GTP hydrolysis by RHO proteins (e.g., Gly14 or Gln63 in RHOA), and these mutations generate proteins known as constitutively active mutants [123,124]. In other mutants, the catalytic arginine residue of the GAP domain is replaced with an alanine residue [113,124]. The latter approach is, in principle, very useful under cell-free conditions but not optimal in cells because an Arg-to-Ala mutant may provide a readout similar to that of the wild-type protein as it interferes with downstream signaling by competing with effector(s) for binding to RHO proteins. These RHOGAP mutants are able to bind persistently to their target protein, sequestering the target, which most likely leads to a readout similar to that of activated wild-type RHOGAP. Therefore, it has recently been suggested that mutating critical “binding determinants”, particularly Lys319 and Arg323 (p50 numbering), may be a better strategy than substituting the catalytic arginine [114]. Charge reversal of these residues most likely leads to loss of RHOGAP association with its substrate RHO protein and thus abrogates the activity of the GAP domain. This outcome renders mutagenesis not only a tool for determining the specificity of RHOGAPs, but also for investigating GAP domain-independent function(s) of the RHOGAPs.
3.4.1. RHOGAP Family Proteins
The GTPase reaction is of great medical significance, since any disruption of this reaction such as that caused by inhibitory mutations in genes encoding GAP proteins results in persistent downstream signaling. The discovery that GAPs are required for GTPase downregulation was made on the basis that microinjection of recombinant GTP-bound RAS into living cells results in faster GTP hydrolysis than is realized in vitro [125]. The first discovered RHOGAP, p50RHOGAP, was identified by biochemical analysis of human spleen cell extracts with recombinant RHOA [119], and this discovery led to the identification of other RHOGAP-containing proteins such as chimaerin and BCR, whose amino acid sequences are related to p50RHOGAP [126]. Since then, more than 66 RHOGAP-containing proteins have been identified in humans [114,127] The RHOGAP family is identified by the presence of a conserved catalytic GAP domain that is sufficient for RHOGAP interaction with RHO proteins and, in most cases, stimulation of the intrinsic GTP hydrolysis reaction of RHO GTPases [115]. In addition to their signature RHOGAP domain, most RHOGAP family members frequently harbor several other functional domains (Figure 4). The majority of these domains can be classified into the following three major groups: (i) lipid- and membrane-binding domains; (ii) peptide- and protein-interacting domains; and (iii) catalytic domains with enzymatic activities. The most widespread domains are PH, CC, P, SRC homology 3, and BAR/F-BAR (Figure 4). These domains are implicated in regulation, membrane targeting, localization, and potential phosphorylation sites and indicate the complexity of the regulation of GTPase activity. Thirteen GAPs lack any additional putative domains but contain highly variable regions in their N- and C-termini (Figure 4). It is possible that these regions consist of motifs that have not yet been identified, and these regions may contribute to their specific function in the cell.
3.4.2. Structural and Functional Characteristics of the RHOGAP Domain
The GAP domain of the RHOGAP family consists of approximately 190 amino acids and shares high sequence homology within the family. Although the RHOGAP domain shares no similarities to RASGAP family members at the amino acid level, RHOGAPs and RASGAPs resemble each other in their tertiary structure [128,129]. Comparative structural analysis of the RHOGAP domain with other GAPs of RAS subfamilies has suggested that GAP domains in RAS and RHO family proteins are evolutionarily related [128,130] and that the catalytic domains of RHOGAPs share a core structural fold. The RHOGAP domain is made up of seven α-helices. The functional characteristic of the RHOGAP domain is a pair of conserved basic residues: catalytic arginine (the arginine finger) and lysine (Arg282 and Lys319 in p50RHOGAP numbering) [114,131].
3.4.3. The Mechanism by Which the GAP Domain Mediates GTP Hydrolysis
Crystallographic studies of RHOGAP domains in complex with CDC42 bound to GppNHp, RHOA/CDC42 bound to GDP·AlF4 [76,77,95] and RHOA bound to GDP·MgF3 [132] have provided insights into the catalytic mechanism of GTP hydrolysis upon stimulation. The GTPase reaction, as part of the switch mechanism, leads to changes in the conformation of the GTPase, especially in flexible and mobile loops known as switch regions. RHOGAP interacts with the switch I and II regions [3,133] and the P-loop of the RHO protein. The GAP domain accelerates the intrinsic GTP hydrolysis by RHO proteins in two ways. First, it directly contributes to catalysis by inserting catalytic arginine in the GAP domain into the active site of the RHO protein. This establishes contacts with the main-chain carbonyl of Gly12 (RAC1 numbering) and helps stabilize the GTP-hydrolysis transition state [134]. Second, this interaction stabilizes the negative charges formed during the transition state of GTP hydrolysis and positions the catalytic glutamine residue (Gln61 RAC1 numbering) of the RHO protein to enable its coordination with nucleophilic water molecules [129,135]. RHOGAP also stabilizes the switch regions of the RHO protein by interacting with residues associated with its intrinsic GTPase activity [113]. ARHGAP36, CNTD1, DEP1, DEP2, FAM13B, INPP5P [136], and OCRL1 lack an arginine finger, which makes them catalytically inactive (Figure 4) [114]. ARHGAP36 is involved in GLI transcription factor activation, but this function proceeds independent of its GAP domain. CNTD1 lacks RHOGAP activity and acts as an ARF6 GAP. DEP1 and DEP2 coordinate cell cycle progression and interfere with RHOA action and signaling even though they lack RHOGAP activity. OCRL1 has been shown to interact with GTP-bound RAC1 without stimulating hydrolysis. p85α and p85β (85-kDa regulatory subunits of phosphoinositide 3-kinases) are also RHOGAP-like proteins (Figure 4), as they do not show any detectable GAP activity toward different RHO proteins [28]. A prerequisite of GAP function is that the GAP domain position its catalytic residue Arg282 (p50 numbering); therefore, GAPs include a number of amino acids that are critical for binding and stabilizing the protein complex. Both p85 isoforms lack most of binding determinants (e.g., Arg323, Asn391, Val394, and Pro398) as well as the conserved amino acids around the arginine finger [114].
3.4.4. Overabundance and Diversity
Using database searches, 66 distinct RHOGAP domain-containing proteins were found to be encoded in the human genome, whereas the number of RHO family proteins that need to be regulated by GAPs was 18 (excluding constitutively active RHO proteins). The overabundance of RHOGAPs implies that they must be tightly regulated in the cell to prevent RHO proteins from being accidentally turned off. Of the 66 RHOGAPs, 57 proteins have a common catalytic domain capable of terminating RHO protein signaling by stimulating the slow intrinsic GTP hydrolysis (GTPase) reaction (Figure 4). Investigation of the sequence-structure-function relationship between RHOGAPs and RHO proteins by combining in vitro data with in silico data has revealed that the RHOGAP domain itself is nonselective, and in some cases, it is rather inefficient under cell-free conditions. This finding suggests that other domains in RHOGAPs confer substrate specificity and fine-tunes their catalytic efficiency in cells [114].
3.4.5. Regulation and GAP Proteins Functions
RHOGAPs are widely expressed, which makes their apparent redundancy questionable. Therefore, cells must regulate RHOGAPs very tightly to prevent unwanted events that switch off signaling. To ensure stringent regulatory control, RHOGAPs are modulated at different levels, indicating that regions outside the RHOGAP domain most likely determine the specificity of RHOGAPs (Figure 4). Numerous mechanisms have been shown to affect the catalytic activity and substrate specificity of RHOGAPs (e.g., autoinhibition (GRAF and OPHN1) [120]); posttranslational regulation such as phosphorylation (p190GAP and Mgc-RACGAP) [121]; lipid binding via PH or C2 domains [122]; protein–protein interactions (DLC1/p120RASGAP) [117,118] and subcellular distribution through specific colocalization of RHOGAPs with RHO proteins at the membrane, for example, with a scaffolding protein (Figure 4) [137].
4. Downstream Effectors of RHO GTPases
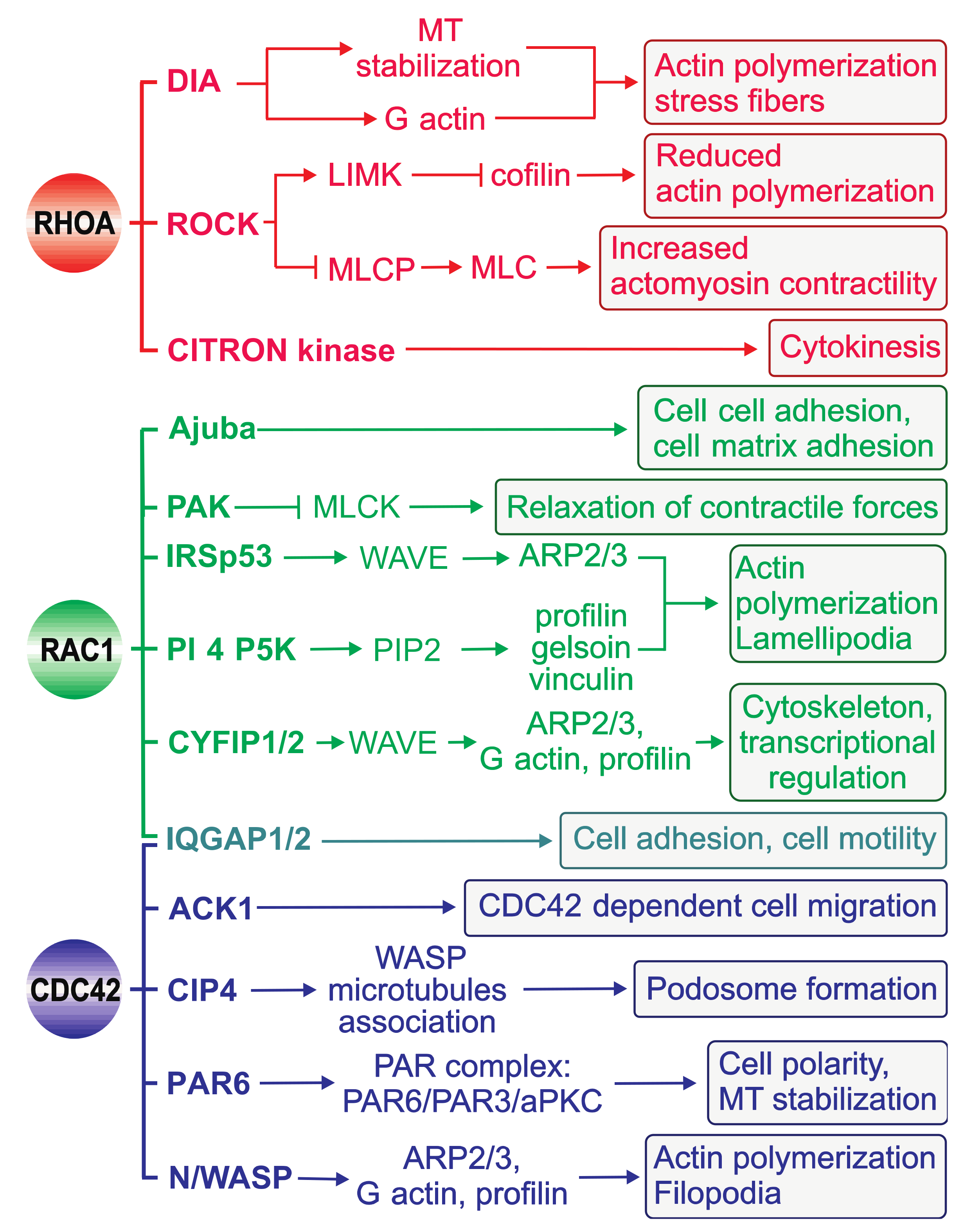
The ability of RHO GTPases to control a wide range of intracellular signaling pathways is attributed to their association with their cellular targets: effector proteins (Figure 5, Table 1). In contrast to regulators that interact with RHO GTPases to modulate their switch function, effectors require GTPases to be in a specific conformation to realize their own intrinsic function. To date, more than 70 potential effectors have been identified for RHOA, RAC1, and CDC42 [58].
The effector proteins are either kinases or scaffolding proteins (Figure 5, Table 1). Kinases form an important class of RHO effectors and result in downstream phosphorylation cascades. Different RHO-associated serine/threonine kinases such as PAK (p21-activated kinase), ROCK (RHO-associated coiled-coil kinase), CRIK (citron kinase), and PKN (protein kinase novel) interact with and are regulated by their partner GTPases [138,139,140]. Another group of effectors comprise scaffolding proteins, which probably form a framework for signaling cascades, especially through filamentous actin dynamics. IQ motif-containing GTPase-activating protein 1 (IQGAP1) [141], mammalian homolog of Drosophila diaphanous 1 (DIA1), Wiskott-Aldrich syndrome protein (WASP), and Rhotekin (RTKN) are the most extensively investigated effectors in this regard and facilitate complex formation in cells [142].
4.1. Structural Characteristics of RHO GTPase-effector Interactions
The crystal structures of the GTPase-binding domains (GBDs) of PKN and RHO kinase (ROCK) in complex with RHOA revealed that the domains, as predicted from their primary structure, form α-helical coiled coils that are arranged in an antiparallel and parallel fashion, respectively [143,144]. A 13-residue left-handed coiled coil in the C-terminal portion of the ROCK-GBD, which is considered the minimal sequence required for RHO-interacting motif activity, binds exclusively to the switch and α2 regions of RHOA. In contrast, the RHOA-PKN complex has two possible contact sites on RHOA [143]: contact site 1 consists of the α1, β2/β3, and α5 regions of RHOA, whereas contact site 2 overlaps remarkably well with the ROCK-binding site. The structures of CDC42 in complex with effector proteins containing a CDC42/RAC-interactive binding (CRIB) motif such as PAK1 and WASP, which have been determined mostly by NMR spectroscopy due to their high flexibility [145,146,147,148,149], have shown that the GBD in this class of effectors makes extensive contact with the surface of RHO GTPases. Specifically, GBD binds through its β-hairpin and C-terminal α-helix to the α1, switch I, and II regions and wraps around the α5 and β2 regions of the GTPase with its extended N-terminus, which encompasses the CRIB motif. The basic region of WASP, immediately upstream of the CRIB motif, has been shown to generate favorable electrostatic steering forces to unique glutamate residues in CDC42 (Glu49, Glu171, and Glu178) that control the accelerated WASP-CDC42 association reaction (Figure 2) [52,150]. This process is a prerequisite for WASP activation and a critical step in the temporal regulation and integration of WASP-mediated cellular responses (Figure 5).
Two other effectors, arfaptin and p67phox, have novel structures and contact sites on the GTPase [151,152]. Arfaptin forms an elongated crescent-shaped dimer with three helix coiled-coils that makes contact with the switches I and II and α2 regions of RAC1, regardless of its nucleotide-bound state [152], and structurally mimics the DH domain of Tiam1 [153]. p67phox has an α-helical domain that consists of four tetratricopeptide repeat (TPR) motifs, which bind α1, the N-terminal residues of switch I, and the G3 and G5 loops, but not the switch II region or the principal parts of switch I [151]. It has been proposed that the switch regions might be the contact sites for a third protein that is associated with the Rac1-GTP-p67phox complex [154,155].
The mechanism of effector activation of the GTPase–effector complex structures mentioned thus far have not been clarified; however, intramolecular autoinhibition and exposure of their functional domains are known to be required. A common feature of effector complexes is that, with the exception of p67phox, they all make intensive contact with the switch/α2 regions of RHO GTPases, which indicates that this region probably serves as the platform for the GTP-dependent recognition of effectors. Two invariant leucine residues (Leu69 and Leu72), which form crucial hydrophobic contacts with almost all effector domains, have been proposed as essential elements for the CDC42/RAC-mediated activation of CRIB-containing effectors [148]. A different activation mechanism has been implicated for the RHO-specific effectors PKN and ROCK, with other domains that bind cooperatively to sites outside the switch regions of RHOA [156].
4.2. RHO GTPase-Mediated Effector Signaling
The fact that effectors commonly contact distinct residues within the highly conserved switch I and II regions of RHO GTPases [3,5] strongly suggests that other domains bind cooperatively to sites outside the switch regions [3,156]. This possibility might explain the cellular specificity of RHO GTPase–effector interactions. Pioneering experiments by Alan Hall and colleagues showed that the reorganization of the actin cytoskeleton is regulated by proteins in the RHO family including CDC42, RAC1, and RHOA (Figure 5) [157]. CDC42 and RAC1 activation, in turn, activates the ARP2/3 complex indirectly via WASP and WAVE to induce branched actin filament networks and the formation of tight bundles of parallel filaments that form the core in filopodia and the formation of a network of diagonally oriented actin filaments that give rise to thin sheets of lamellipodia. RHOA activation leads to the activation of ROCK and DIA and the organization of actomyosin bundles into stress fibers as well as the formation of focal adhesions [1,5]. Coordination of the distinct roles of these GTPases is crucial for regulating cell migration, as demonstrated by wound closure in a fibroblast monolayer: CDC42 regulates cell polarity, RAC1 regulates the protrusion of lamellipodia at the leading edge, and RHO regulates the turnover of highly organized structures termed focal adhesions (reviewed in [7,158,159,160,161,162,163,164,165]).
Moreover, RHO GTPases control signal transduction pathways that influence gene expression including the serum response factor (SRF), nuclear factor κB (NFκB) transcription factor, c-JUN N-terminal kinase (JNK), and p38 mitogen-activated protein kinase pathways [19,166]. It has been reported that several enzyme activities can be altered by RHO GTPases. RAC1 can bind directly to p67phox, a component of the NADPH oxidase complex, and activate NADPH oxidase activity to generate reactive oxygen species (ROS) (Table 1) [167]. The BCR gene produces a 160 kDa product called p160bcr, which encompasses several distinct domains. p160 exhibited GAP activity toward RAC1, RAC2, and CDC42 GTPases. An early study has shown that BCR regulates RAC-mediated superoxide production by the NADPH-oxidase system of leukocytes [168].
5. Conclusions
Abnormal activation of RHO proteins has been shown to play a crucial role in cancer, infectious and cognitive disorders, and cardiovascular diseases. However, several studies must be performed to gain understanding into the complexity of RHO protein signaling. (i) The RHO family comprises 20 signaling proteins, of which only RHOA, RAC1, and CDC42 have been comprehensively studied thus far. The functions of the less-characterized members of this protein family await detailed investigation. (ii) Despite intensive research over the past two decades, the mechanisms by which RHOGDIs associate and extract RHO proteins from the membrane and the factors displacing the RHO protein from the complex with RHOGDI remain to be elucidated. (iii) A tremendous number of 20 RHO-regulating proteins (85 GEFs and 66 GAPs) exist in the human genome. How these regulators selectively recognize their RHO protein targets is not well understood, and the majority of GEFs and GAPs in humans remain uncharacterized. (iv) GDIs, GEFs, GAPs, and effectors, despite their structural diversity, share consensus binding sites within the switch I and II regions [3]. However, all these RHO-binding partners require contact with other regions, not their shared binding region, to realize their specificity for different RHO proteins. (v) A major challenge ahead, which has not been fully addressed thus far, will be gaining an understanding of the spatial temporal regulation of RHO GTPase activity and the interaction of RHO proteins with distinct downstream effectors. (vi) Most GEFs and GAPs need to be regulated and their activation is generally achieved through the release of autoinhibitory elements [92,120]. With a few exceptions [4], the operating principles of these autoregulatory mechanisms remain obscure. (vii) A better understanding of the specificity and the mode of action of the regulatory proteins as well as the selective recruitment and activation of effectors to specific subcellular sites is not only fundamentally important for understanding many aspects of RHO biology, but is also the master key to unlocking the identity of key targets useful in developing drugs against a variety of diseases caused by aberrant RHO protein functions. This regards the spatiotemporal features whose understanding is afflicted with major conceptual shortcomings. Future models should consider both the emerging principle of biomolecular condensates (or non-membrane-bound organelles) that are assembled in liquid-liquid phase separation [169,170] and the modulating principle of accessory proteins [171,172,173], which appears to safeguard the strength, efficiency, and specificity of signal transduction.
Author Contributions
N.M. and M.R.A. contributed to the conception and design of the article, prepared the figures and Table 1, and provided critical revision of the article and final approval of the accepted version for publication. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the German Research Foundation (Deutsche Forschungsgemeinschaft or DFG; AH 92/8-1); the German Research Foundation (Deutsche Forschungsgemeinschaft or DFG) through the International Research Training Group “Intra and interorgan communication of the cardiovascular system” (grant number: IRTG 1902-p6); the European Network on Noonan Syndrome and Related Disorders (NSEuroNet, grant number: 01GM1621B); and the German Federal Ministry of Education and Research (BMBF) German Network of RASopathy Research (GeNeRARe, grant numbers: 01GM1902C).
Acknowledgments
We are grateful to our colleagues Mamta Jaiswal and Ehsan Amin for the stimulating discussion.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| ABD | APC binding domain |
| ACK1 | Activated CDC42 kinase 1 |
| AlF4 | Tetrafluoroaluminate |
| ANK | Ankyrin-repeat |
| ARP2/3 | Actin related protein 2/3 |
| ASH | ASPM-SPD-2-Hydin |
| BAR | BIN–Amphiphysin–RVS |
| BCR | Breakpoint cluster region |
| BRCT | BRCA1 C-terminal domain |
| C1 | DAG-binding domain |
| C2 | Calcium/lipid-binding domain |
| CC | Coiled-coil |
| CDC25 | Catalytic RASGEF domain |
| CDC42 | Cell division control protein 42 |
| CH | Calponin homology |
| CIP4 | CDC42 interacting protein |
| CNTD1 | Cyclin N-terminal domain-containing protein |
| CYFIP1/2 | Cytoplasmic FMR-1 interacting protein 1/2 |
| DAG kinase | Diacyglycerol kinase |
| DAG | Diacylglycerol |
| DBL | Diffuse B-cell lymphoma |
| DEP | Dishevelled, Egl-10 and Pleckstrin domain |
| DH | DBL homology |
| DIA | Diaphanous-related formin |
| DLC1 | Deleted in liver Cancer |
| EH1 | EPS15 homology |
| F-BAR | Extended Fes-CIP4 homology (FCH)-BAR |
| Far | Farnesyl moiety |
| FERM | 4.1 protein/ezrin/radixin/moesin |
| FF | Conserved phenylalanines |
| FMNL3 | Formin-like protein 3 |
| FYVE | FAB1/YOTB/VAC1/EEA1 |
| GAPs | GTPase activating proteins |
| GDIs | Guanine nucleotide dissociation inhibitors |
| GDP | Guanosine-5′-diphosphate |
| GEFs | Guanine nucleotide exchange factors |
| GG | Geranylgeranyl moiety |
| GIT | G protein-coupled receptor kinase-interactor 1 |
| GppNHp | Guanosine-5′- [(β, γ)-imido] triphosphate, Trisodium salt |
| GTP | Guanosine-5′-triphosphate |
| GTPase | GTP-binding and hydrolyzing domain |
| GTPase | Guanine triphosphatase |
| HVR | Hypervariable region |
| ICMT | Isoprenylcystein carboxyl methyltransferase |
| IP3 | Inositol triphosphate |
| IPP5 | Inositol polyphosphate 5-phosphatase |
| IQ | Isoleucine-glutamine-rich |
| IQGAP | IQ motif-containing GTPase activating proteins |
| IRSp53 | Insulin receptor substrate 53 kDa |
| kDa | Kilodalton |
| LFA-1 | Leukocyte function associated molecule 1 alpha |
| lgC2 | Immunoglobulin C-2 Type |
| LIMK | LIM domain kinase |
| MBS | Myosin binding subunit |
| MEKK | Mitogen Activated Protein kinase kinase kinase |
| MgF3 | Magnesium fluoride |
| MLC | Myosin light chain |
| MLCK | Myosin light chain kinase |
| MLCP | Myosin light chain phosphatase |
| MLK | Mixed lineage kinase |
| MRCK | Myotonic dystrophy kinase related- CDC42 binding Kinase |
| MT | Microtubule |
| MYSc | Myosin head domain |
| MyTH4 | Myosin tail homology 4 |
| OCRL1 | Lowe oculocerebrorenal syndrome protein |
| OPHN1 | Oligophrenin |
| P | Proline-rich |
| PA | Phosphatidic acid |
| PAK | p21 activated kinase |
| P | Palmitoyl moiety |
| PAR6 | Partitioning defective 6 homolog |
| PBR | Polybasic region |
| PDZ | PSD95/DLG/ZO-1 |
| PEST | Proline-glutamic acid-serine-threonine-rich |
| PH | Pleckstrin homology |
| PI3 kinase | Phosphatidylinositol 3-kinase |
| PI4P5K | Phosphatidylinositol 4, phosphate 5 kinase |
| PIP2 | Phosphatidylinositol 4,5 biphosphate |
| PIP3 | Phosphatidylinosiltol-3,4,5-triphosphate |
| PIX | p21-activated protein kinase exchange factor |
| PLC | Phospholipase C |
| POR1 | Partner of RAC1 |
| RA | RAS association domain |
| RAC | RAS-related C3 botulinum toxin substrate |
| RALBD | RAL-binding domain |
| RBD | RAS-binding domain |
| Rce1 | RAS converting enzyme |
| REM | RAS exchanger motif |
| RGS | Regulator of G protein signaling |
| RHO | RAS homolog |
| ROCK | RHO-associated protein kinase |
| S_TKc | Serine/Threonine protein kinase catalytic domain |
| SAM | Sterile alpha motif |
| SEC14 | Phosphoinositide binding |
| SH | SRC homology |
| SH2 | SRC homology 2 |
| SH3 | SRC homology 3 |
| SPEC | Spectrin repeat |
| StART | Steroidogenic acute regulatory protein-related lipid transfer |
| TCR | T cell receptor |
| VPS9 | Vacuolar protein sorting-associated 9 |
| WASP | Wiskott-Aldrich Syndrome Protein |
| WAVE | WASP family verprolin homologous protein |
| WW | Conserved tryptophans |
References
- Jaffe, A.B.; Hall, A. Rho GTPases: Biochemistry and biology. Annu. Rev. Cell Dev. Biol. 2005, 21, 247–269. [Google Scholar] [CrossRef] [Green Version]
- Ellenbroek, S.I.J.; Collard, J.G. Rho GTPases: Functions and association with cancer. Clin. Exp. Metastasis 2007, 24, 657–672. [Google Scholar] [CrossRef]
- Dvorsky, R.; Ahmadian, M.R. Always look on the bright site of Rho: Structural implications for a conserved intermolecular interface. EMBO Rep. 2004, 5, 1130–1136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cherfils, J.; Zeghouf, M. Regulation of small GTPases by GEFs, GAPs, and GDIs. Physiol. Rev. 2013, 93, 269–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bishop, A.L.; Hall, A. Rho GTPases and their effector proteins. Biochem. J. 2000, 348, 241–255. [Google Scholar] [CrossRef]
- Burridge, K.; Wennerberg, K. Rho and Rac Take Center Stage. Cell 2004, 116, 167–179. [Google Scholar] [CrossRef] [Green Version]
- Etienne-Manneville, S.; Hall, A. Rho GTPases in cell biology. Nature 2002, 420, 629–635. [Google Scholar] [CrossRef]
- Hodge, R.G.; Ridley, A.J. Regulating Rho GTPases and their regulators. Nat. Rev. Mol. Cell Biol. 2016, 17, 496–510. [Google Scholar] [CrossRef] [PubMed]
- Knyphausen, P.; Kuhlmann, N.; de Boor, S.; Lammers, M. Lysine-acetylation as a fundamental regulator of ran function: Implications for signaling of proteins of the ras-superfamily. Small GTPases 2015, 6, 189–195. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, N.; Kato, T.; Fujita, A.; Ishizaki, T.; Narumiya, S. Cooperation between mDia1 and ROCK in Rho-induced actin reorganization. Nat. Cell Biol. 1999, 1, 136–143. [Google Scholar] [CrossRef]
- Maekawa, M.; Ishizaki, T.; Boku, S.; Watanabe, N.; Fujita, A.; Iwamatsu, A.; Obinata, T.; Ohashi, K.; Mizuno, K.; Narumiya, S. Signaling from Rho to the actin cytoskeleton through protein kinases ROCK and LIM-kinase. Science 1999, 285, 895–898. [Google Scholar] [CrossRef] [PubMed]
- Madaule, P.; Eda, M.; Watanabe, N.; Fujisawa, K.; Matsuoka, T.; Bito, H.; Ishizaki, T.; Narumiya, S. Role of citron kinase as a target of the small GTPase Rho in cytokinesis. Nature 1998, 394, 491–494. [Google Scholar] [CrossRef]
- Kimura, K.; Ito, M.; Amano, M.; Chihara, K.; Fukata, Y.; Nakafuku, M.; Yamamori, B.; Feng, J.; Nakano, T.; Okawa, K.; et al. Regulation of myosin phosphatase by Rho and Rho-associated kinase (Rho- kinase). Science 1996, 273, 245–248. [Google Scholar] [CrossRef] [PubMed]
- Vega, F.M.; Colomba, A.; Reymond, N.; Thomas, M.; Ridley, A.J. RhoB regulates cell migration through altered focal adhesion dynamics. Open Biol. 2012, 2. [Google Scholar] [CrossRef] [Green Version]
- Vega, F.M.; Fruhwirth, G.; Ng, T.; Ridley, A.J. RhoA and RhoC have distinct roles in migration and invasion by acting through different targets. J. Cell Biol. 2011, 193, 655–665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mino, A.; Troeger, A.; Brendel, C.; Cantor, A.; Harris, C.; Ciuculescu, M.F.; Williams, D.A. Small GTPases RhoH participates in a multi-protein complex with the zinc finger protein kaiso that regulates both cytoskeletal structures and chemokine-induced T cells. Small GTPases 2018. [Google Scholar] [CrossRef] [Green Version]
- Bagrodia, S.; Cerione, R.A. PAK to the future. Trends Cell Biol. 1999, 9, 350–355. [Google Scholar] [CrossRef]
- Nagata, K.I.; Puls, A.; Futter, C.; Aspenstrom, P.; Schaefer, E.; Nakata, T.; Hirokawa, N.; Hall, A. The MAP kinase kinase kinase MLK2 co-localizes with activated JNK along microtubules and associates with kinesin superfamily motor KIF3. EMBO J. 1998, 17, 149–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teramoto, H.; Coso, O.A.; Miyata, H.; Igishi, T.; Miki, T.; Silvio Gutkind, J. Signaling from the small GTP-binding proteins Rac1 and Cdc42 to the c- Jun N-terminal kinase/stress-activated protein kinase pathway: A role for mixed lineage kinase 3/protein-tyrosine kinase 1, a novel member of the mixed lineage kinase family. J. Biol. Chem. 1996, 271, 27225–27228. [Google Scholar] [CrossRef] [Green Version]
- Miki, H.; Suetsugu, S.; Takenawa, T. WAVE, a novel WASP-family protein involved in actin reorganization induced by Rac. EMBO J. 1998, 17, 6932–6941. [Google Scholar] [CrossRef] [Green Version]
- Chou, M.M.; Blenis, J. The 70 kDa S6 kinase complexes with and is activated by the Rho family G proteins Cdc42 and Rac1. Cell 1996, 85, 573–583. [Google Scholar] [CrossRef] [Green Version]
- Fukata, M.; Watanabe, T.; Noritake, J.; Nakagawa, M.; Yamaga, M.; Kuroda, S.; Matsuura, Y.; Iwamatsu, A.; Perez, F.; Kaibuchi, K. Rac1 and Cdc42 capture microtubules through IQGAP1 and CLIP-170. Cell 2002, 109, 873–885. [Google Scholar] [CrossRef] [Green Version]
- Fukataf, M.; Kuroda, S.; Fujii, K.; Nakamura, T.; Shoji, I.; Matsuura, Y.; Okawa, K.; Iwamatsu, A.; Kikuchi, A.; Kaibuchi, K. Regulation of cross-linking of actin filament by IQGAP1, a target for Cdc42. J. Biol. Chem. 1997, 272, 29579–29583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fanger, G.R.; Johnson, N.L.; Johnson, G.L. MEK kinases are regulated by EGF and selectively interact with Rac/Cdc42. EMBO J. 1997, 16, 4961–4972. [Google Scholar] [CrossRef] [Green Version]
- Van Aelst, L.; Joneson, T.; Bar-Sagi, D. Identification of a novel Rac1-interacting protein involved in membrane ruffling. EMBO J. 1996, 15, 3778–3786. [Google Scholar] [CrossRef]
- Diekmann, D.; Abo, A.; Johnston, C.; Segal, A.W.; Hall, A. Interaction of Rac with p67phox and regulation of phagocytic NADPH oxidase activity. Science 1994, 265, 531–533. [Google Scholar] [CrossRef] [PubMed]
- Bokoch, G.M.; Vlahos, C.J.; Wang, Y.; Knaus, U.G.; Traynor-Kaplan, A.E. Rac GTPase interacts specifically with phosphatidylinositol 3-kinase. Biochem. J. 1996, 315, 775–779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tolias, K.F.; Cantley, L.C.; Carpenter, C.L. Rho family GTPases bind to phosphoinositide kinases. J. Biol. Chem. 1995, 270, 17656–17659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Houssa, B.; De Widt, J.; Kranenburg, O.; Moolenaar, W.H.; Van Blitterswijk, W.J. Diacylglycerol kinase θ binds to and is negatively regulated by active RhoA. J. Biol. Chem. 1999, 274, 6820–6822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Illenberger, D.; Schwald, F.; Pimmer, D.; Binder, W.; Maier, G.; Dietrich, A.; Gierschik, P. Stimulation of phospholipase C-β2 by the Rho GTPases Cdc42Hs and Rac1. EMBO J. 1998, 17, 6241–6249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, A.; Karnoub, A.E.; Palmby, T.R.; Lengyel, E.; Sondek, J.; Der, C.J. Rac1b, a tumor associated, constitutively active Rac1 splice variant, promotes cellular transformation. Oncogene 2004, 23, 9369–9380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arana, E.; Vehlow, A.; Harwood, N.E.; Vigorito, E.; Henderson, R.; Turner, M.; Tybulewicz, V.L.L.J.; Batista, F.D. Activation of the Small GTPase Rac2 via the B Cell Receptor Regulates B Cell Adhesion and Immunological-Synapse Formation. Immunity 2008, 28, 88–99. [Google Scholar] [CrossRef] [Green Version]
- Hajdo-Milasinovic, A.; van der Kammen, R.A.; Moneva, Z.; Collard, J.G. Rac3 inhibits adhesion and differentiation of neuronal cells by modifying GIT1 downstream signaling. J. Cell Sci. 2009, 122, 2127–2136. [Google Scholar] [CrossRef] [Green Version]
- Vignal, E.; Blangy, A.; Martin, M.; Gauthier-Rouvière, C.; Fort, P. Kinectin Is a Key Effector of RhoG Microtubule-Dependent Cellular Activity. Mol. Cell. Biol. 2001, 21, 8022–8034. [Google Scholar] [CrossRef] [Green Version]
- Rohatgi, R.; Ma, L.; Miki, H.; Lopez, M.; Kirchhausen, T.; Takenawa, T.; Kirschner, M.W. The interaction between N-WASP and the Arp2/3 complex links Cdc42-dependent signals to actin assembly. Cell 1999, 97, 221–231. [Google Scholar] [CrossRef] [Green Version]
- Abo, A.; Qu, J.; Cammarano, M.S.; Dan, C.; Fritsch, A.; Baud, V.; Belisle, B.; Minden, A. PAK4, a novel effector for Cdc42Hs, is implicated in the reorganization of the actin cytoskeleton and in the formation of filopodia. EMBO J. 1998, 17, 6527–6540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leung, T.; Chen, X.-Q.; Tan, I.; Manser, E.; Lim, L. Myotonic Dystrophy Kinase-Related Cdc42-Binding Kinase Acts as a Cdc42 Effector in Promoting Cytoskeletal Reorganization. Mol. Cell. Biol. 1998, 18, 130–140. [Google Scholar] [CrossRef] [Green Version]
- Wilson, E.; Leszczynska, K.; Poulter, N.S.; Edelmann, F.; Salisbury, V.A.; Noy, P.J.; Bacon, A.; Rappoport, J.Z.; Heath, J.K.; Bicknell, R.; et al. RhoJ interacts with the GIT-PIX complex and regulates focal adhesion disassembly. J. Cell Sci. 2014, 127, 3039–3051. [Google Scholar] [CrossRef] [Green Version]
- Ackermann, K.L.; Florke, R.R.; Reyes, S.S.; Tader, B.R.; Hamann, M.J. TCL/RhoJ plasma membrane localization and nucleotide exchange is coordinately regulated by amino acids within the N terminus and a distal loop region. J. Biol. Chem. 2016, 291, 23604–23617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aspenström, P. Atypical Rho GTPases RhoD and Rif integrate cytoskeletal dynamics and membrane trafficking. Biol. Chem. 2014, 395, 477–484. [Google Scholar] [CrossRef]
- Gad, A.K.B.; Aspenström, P. Rif proteins take to the RhoD: Rho GTPases at the crossroads of actin dynamics and membrane trafficking. Cell. Signal. 2010, 22, 183–189. [Google Scholar] [CrossRef]
- Li, Y.H.; Ghavampur, S.; Bondallaz, P.; Will, L.; Grenningloh, G.; Puschel, A.W. Rndl regulates axon extension by enhancing the microtubule destabilizing activity of SCG10. J. Biol. Chem. 2009, 284, 363–371. [Google Scholar] [CrossRef] [Green Version]
- Fujita, H.; Katoh, H.; Ishikawa, Y.; Mori, K.; Negishi, M. Rapostlin is a novel effector of Rnd2 GTPase inducing neurite branching. J. Biol. Chem. 2002, 277, 45428–45434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, H.; Harada, A.; Mori, K.; Negishi, M. Socius Is a Novel Rnd GTPase-Interacting Protein Involved in Disassembly of Actin Stress Fibers. Mol. Cell. Biol. 2002, 22, 2952–2964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riento, K.; Totty, N.; Villalonga, P.; Garg, R.; Guasch, R.; Ridley, A.J. RhoE function is regulated by ROCK I-mediated phosphorylation. EMBO J. 2005, 24, 1170–1180. [Google Scholar] [CrossRef] [Green Version]
- Riou, P.; Villalonga, P.; Ridley, A.J. Rnd proteins: Multifunctional regulators of the cytoskeleton and cell cycle progression. BioEssays 2010, 32, 986–992. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, M.; Dvorsky, R.; Ahmadian, M.R. Deciphering the molecular and functional basis of Dbl family proteins: A novel systematic approach toward classification of selective activation of the Rho family proteins. J. Biol. Chem. 2013, 288, 4486–4500. [Google Scholar] [CrossRef] [Green Version]
- Wittinghofer, A.; Vetter, I.R. Structure-Function Relationships of the G Domain, a Canonical Switch Motif. Annu. Rev. Biochem. 2011, 80, 943–971. [Google Scholar] [CrossRef] [PubMed]
- Roberts, P.J.; Mitin, N.; Keller, P.J.; Chenette, E.J.; Madigan, J.P.; Currin, R.O.; Cox, A.D.; Wilson, O.; Kirschmeier, P.; Der, C.J. Rho family GTPase modification and dependence on CAAX motif-signaled posttranslational modification. J. Biol. Chem. 2008, 283, 25150–25163. [Google Scholar] [CrossRef] [Green Version]
- Fiegen, D.; Blumenstein, L.; Stege, P.; Vetter, I.R.; Ahmadian, M.R. Crystal structure of Rnd3/RhoE: Functional implications. FEBS Lett. 2002, 525, 100–104. [Google Scholar] [CrossRef] [Green Version]
- Fiegen, D.; Haeusler, L.C.; Blumenstein, L.; Herbrand, U.; Dvorsky, R.; Vetter, I.R.; Ahmadian, M.R. Alternative Splicing of Rac1 Generates Rac1b, a Self-activating GTPase. J. Biol. Chem. 2004, 279, 4743–4749. [Google Scholar] [CrossRef] [Green Version]
- Hemsath, L.; Dvorsky, R.; Fiegen, D.; Carlier, M.F.; Ahmadian, M.R. An electrostatic steering mechanism of Cdc42 recognition by Wiskott-Aldrich syndrome proteins. Mol. Cell 2005, 20, 313–324. [Google Scholar] [CrossRef]
- Kazemein Jasemi, N.; Herrmann, C.; Estirado, E.M.; Gremer, L.; Willbold, D.; Brunsveld, L.; Dvorsky, R.; Ahmadian, M. The intramolecular allostery of GRB2 governing its interaction with SOS1 is modulated by phosphotyrosine ligands. Biochem. J. 2021. [Google Scholar] [CrossRef]
- Ahmadian, M.R.; Jaiswal, M.; Fansa, E.K.; Dvorsky, R. New insight into the molecular switch mechanism of human Rho family proteins: Shifting a paradigm. Biol. Chem. 2013, 394, 89–95. [Google Scholar] [CrossRef]
- Winter-Vann, A.M.; Casey, P.J. Post-prenylation-processing enzymes as new targets in oncogenesis. Nat. Rev. Cancer 2005, 5, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Lang, P.; Gesbert, F.; Delespine-Carmagnat, M.; Stancou, R.; Pouchelet, M.; Bertoglio, J. Protein kinase A phosphorylation of RhoA mediates the morphological and functional effects of cyclic AMP in cytotoxic lymphocytes. EMBO J. 1996, 15, 510–519. [Google Scholar] [CrossRef]
- Forget, M.A.; Desrosiers, R.R.; Gingras, D.; Béliveau, R. Phosphorylation states of Cdc42 and RhoA regulate their interactions with Rho GDP dissociation inhibitor and their extraction from biological membranes. Biochem. J. 2002, 361, 243–254. [Google Scholar] [CrossRef] [PubMed]
- Bustelo, X.R.; Sauzeau, V.; Berenjeno, I.M. GTP-binding proteins of the Rho/Rac family: Regulation, effectors and functions in vivo. BioEssays 2007, 29, 356–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rose, R.; Weyand, M.; Lammers, M.; Ishizaki, T.; Ahmadian, M.R.; Wittinghofer, A. Structural and mechanistic insights into the interaction between Rho and mammalian Dia. Nature 2005, 435, 513–518. [Google Scholar] [CrossRef] [PubMed]
- Troeger, A.; Chae, H.D.; Senturk, M.; Wood, J.; Williams, D.A. A unique carboxyl-Terminal insert domain in the hematopoietic-Specific, gtpase-Deficient rho gtpase rhoh regulates post-Translational processing. J. Biol. Chem. 2013, 288, 36451–36462. [Google Scholar] [CrossRef] [Green Version]
- Garavini, H.; Riento, K.; Phelan, J.P.; McAlister, M.S.B.; Ridley, A.J.; Keep, N.H. Crystal structure of the core domain of RhoE/Rnd3: A constitutively activated small G protein. Biochemistry 2002, 41, 6303–6310. [Google Scholar] [CrossRef] [PubMed]
- Xie, F.; Shao, S.; Zhang, B.; Wang, H.; Liu, B. Role of Rho-specific guanine nucleotide dissociation inhibitor α regulation in cell migration. Acta Histochem. 2017, 119, 183–189. [Google Scholar] [CrossRef]
- Griner, E.M.; Theodorescu, D. The faces and friends of RhoGDI2. Cancer Metastasis Rev. 2012, 31, 519–528. [Google Scholar] [CrossRef] [PubMed]
- De León-Bautista, M.P.; Del Carmen Cardenas-Aguayo, M.; Casique-Aguirre, D.; Almaraz-Salinas, M.; Parraguirre-Martinez, S.; Olivo-Diaz, A.; Del Rocío Thompson-Bonilla, M.; Vargas, M. Immunological and functional characterization of RhoGDI3 and its molecular targets RhoG and RhoB in human pancreatic cancerous and normal cells. PLoS One 2016, 11, e0166370. [Google Scholar] [CrossRef]
- Brunet, N.; Morin, A.; Olofsson, B. RhoGDI-3 regulates RhoG and targets this protein to the Golgi complex through its unique N-terminal domain. Traffic 2002, 3, 342–358. [Google Scholar] [CrossRef]
- Garcia-Mata, R.; Boulter, E.; Burridge, K. The “invisible hand”: Regulation of RHO GTPases by RHOGDIs. Nat. Rev. Mol. Cell Biol. 2011, 12, 493–504. [Google Scholar] [CrossRef] [Green Version]
- DerMardirossian, C.; Bokoch, G.M. GDIs: Central regulatory molecules in Rho GTPase activation. Trends Cell Biol. 2005, 15, 356–363. [Google Scholar] [CrossRef] [PubMed]
- Moissoglu, K.; Schwartz, M.A. Spatial and temporal control of Rho GTPase functions. Cell. Logist. 2014, 4, e943618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tnimov, Z.; Abankwa, D.; Alexandrov, K. RhoGDI facilitates geranylgeranyltransferase-I-mediated RhoA prenylation. Biochem. Biophys. Res. Commun. 2014, 452, 967–973. [Google Scholar] [CrossRef]
- Grizot, S.; Fauré, J.; Fieschi, F.; Vignais, P.V.; Dagher, M.C.; Pebay-Peyroula, E. Crystal structure of the Rac1 - RhoGDI complex involved in NADPH oxidase activation. Biochemistry 2001, 40, 10007–10013. [Google Scholar] [CrossRef] [PubMed]
- Scheffzek, K.; Stephan, I.; Jensen, O.N.; Illenberger, D.; Gierschik, P. The Rac-RhoGDI complex and the structural basis for the regulation of Rho proteins by RhoGDI. Nat. Struct. Biol. 2000, 7, 122–126. [Google Scholar] [CrossRef]
- Dransart, E.; Olofsson, B.; Cherfils, J. RhoGDIs revisited: Novel roles in Rho regulation. Traffic 2005, 6, 957–966. [Google Scholar] [CrossRef]
- Hoffman, G.R.; Nassar, N.; Cerione, R.A. Structure of the Rho family GTP-binding protein Cdc42 in complex with the multifunctional regulator RhoGDI. Cell 2000, 100, 345–356. [Google Scholar] [CrossRef] [Green Version]
- Longenecker, K.; Read, P.; Derewenda, U.; Dauter, Z.; Liu, X.; Garrard, S.; Walker, L.; Somlyo, A.V.; Nakamoto, R.K.; Somlyo, A.P.; et al. How RhoGDI binds Rho. Acta Crystallogr. Sect. D Biol. Crystallogr. 1999, 55, 1503–1515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mosaddeghzadeh, N.; Kazemein Jasemi, N.S.; Majoleé, J.; Zhang, S.C.; Hordijk, P.L.; Dvorsky, R.; Ahmadian, M.R. New insights into the specificity of the RHO GTPase-GDI interaction: An electrostatic force mechanism. J. Biol. Chem. 2021. Under revision. [Google Scholar]
- Cherfils, J.; Chardin, P. GEFs: Structural basis for their activation of small GTP-binding proteins. Trends Biochem. Sci. 1999, 24, 306–311. [Google Scholar] [CrossRef]
- Guo, Z.; Ahmadian, M.R.; Goody, R.S. Guanine nucleotide exchange factors operate by a simple allosteric competitive mechanism. Biochemistry 2005, 44, 15423–15429. [Google Scholar] [CrossRef] [PubMed]
- Hutchinson, J.P.; Eccleston, J.F. Mechanism of nucleotide release from Rho by the GDP dissociation stimulator protein. Biochemistry 2000, 39, 11348–11359. [Google Scholar] [CrossRef]
- Heasman, S.J.; Ridley, A.J. Mammalian Rho GTPases: New insights into their functions from in vivo studies. Nat. Rev. Mol. Cell Biol. 2008, 9, 690–701. [Google Scholar] [CrossRef]
- Rossman, K.L.; Der, C.J.; Sondek, J. GEF means go: Turning on Rho GTPases with guanine nucleotide-exchange factors. Nat. Rev. Mol. Cell Biol. 2005, 6, 167–180. [Google Scholar] [CrossRef]
- Kunimura, K.; Uruno, T.; Fukui, Y. DOCK family proteins: Key players in immune surveillance mechanisms. Int. Immunol. 2020, 32, 5–15. [Google Scholar] [CrossRef]
- Cook, D.R.; Rossman, K.L.; Der, C.J. Rho guanine nucleotide exchange factors: Regulators of Rho GTPase activity in development and disease. Oncogene 2014, 33, 4021–4035. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, S.K.; Wheelock, R.H.P.; Aaronson, S.A.; Eva, A. Identification of the protein encoded by the human diffuse B-cell lymphoma (dbl) oncogene. Proc. Natl. Acad. Sci. USA 1986, 83, 8868–8872. [Google Scholar] [CrossRef] [Green Version]
- Hart, M.J.; Eva, A.; Evans, T.; Aaronson, S.A.; Cerione, R.A. Catalysis of guanine nucleotide exchange on the CDC42Hs protein by the dbloncogene product. Nature 1991, 354, 311–314. [Google Scholar] [CrossRef] [PubMed]
- Aittaleb, M.; Boguth, C.A.; Tesmer, J.J.G. Structure and function of heterotrimeric G protein-regulated Rho guanine nucleotide exchange factors. Mol. Pharmacol. 2010, 77, 111–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffman, G.R.; Cerione, R.A. Signaling to the Rho GTPases: Networking with the DH domain. FEBS Lett. 2002, 513, 85–91. [Google Scholar] [CrossRef] [Green Version]
- Jaiswal, M.; Dubey, B.N.; Koessmeier, K.T.; Gremer, L.; Ahmadian, M.R. Biochemical assays to characterize rho GTPases. Methods Mol. Biol. 2012, 827, 37–58. [Google Scholar] [CrossRef] [PubMed]
- Viaud, J.; Gaits-Iacovoni, F.; Payrastre, B. Regulation of the DH-PH tandem of guanine nucleotide exchange factor for Rho GTPases by phosphoinositides. Adv. Biol. Regul. 2012, 52, 303–314. [Google Scholar] [CrossRef]
- Dubash, A.D.; Wennerberg, K.; García-Mata, R.; Menold, M.M.; Arthur, W.T.; Burridge, K. A novel role for Lsc/p115 RhoGEF and LARG in regulating RhoA activity downstream of adhesion to fibronectin. J. Cell Sci. 2007, 120, 3989–3998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bos, J.L.; Rehmann, H.; Wittinghofer, A. GEFs and GAPs: Critical Elements in the Control of Small G Proteins. Cell 2007, 129, 865–877. [Google Scholar] [CrossRef] [Green Version]
- Vigil, D.; Cherfils, J.; Rossman, K.L.; Der, C.J. Ras superfamily GEFs and GAPs: Validated and tractable targets for cancer therapy? Nat. Rev. Cancer 2010, 10, 842–857. [Google Scholar] [CrossRef] [Green Version]
- Jaiswal, M.; Gremer, L.; Dvorsky, R.; Haeusler, L.C.; Cirstea, I.C.; Uhlenbrock, K.; Ahmadian, M.R. Mechanistic insights into specificity, activity, and regulatory elements of the regulator of G-protein signaling (RGS)-containing Rho-specific guanine nucleotide exchange factors (GEFs) p115, PDZ-RhoGEF (PRG), and leukemia-associated RhoGEF (LARG). J. Biol. Chem. 2011, 286, 18202–18212. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Wang, H.; Eberstadt, M.; Schnuchel, A.; Olejniczak, E.T.; Meadows, R.P.; Schkeryantz, J.M.; Janowick, D.A.; Harlan, J.E.; Harris, E.A.S.; et al. NMR structure and mutagenesis of the N-terminal Dbl homology domain of the nucleotide exchange factor Trio. Cell 1998, 95, 269–277. [Google Scholar] [CrossRef] [Green Version]
- Erickson, J.W.; Cerione, R.A. Structural Elements, Mechanism, and Evolutionary Convergence of Rho Protein-Guanine Nucleotide Exchange Factor Complexes. Biochemistry 2004, 43, 837–842. [Google Scholar] [CrossRef] [PubMed]
- Lemmon, M.A.; Ferguson, K.M.; Abrams, C.S. Pleckstrin homology domains and the cytoskeleton. FEBS Lett. 2002, 513, 71–76. [Google Scholar] [CrossRef] [Green Version]
- Haslam, R.J.; Koide, H.B.; Hemmings, B.A. Pleckstrin domain homology. Nature 1993, 363, 309–310. [Google Scholar] [CrossRef] [PubMed]
- Tyers, M.; Haslam, R.J.; Rachubinski, R.A.; Harley, C.B. Molecular analysis of pleckstrin: The major protein kinase c substrate of platelets. J. Cell. Biochem. 1989, 40, 133–145. [Google Scholar] [CrossRef] [PubMed]
- Dinitto, J.; Lambright, D. Membrane and juxtamembrane targeting by PH and PTB domains. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2006, 1761, 850–867. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Luby-Phelps, K.; Das, B.; Shu, X.; Xia, Y.; Mosteller, R.D.; Krishna, U.M.; Falck, J.R.; White, M.A.; Broek, D. Role of substrates and products of PI 3-kinase in regulating activation of Rac-related guanosine triphosphatases by Vav. Science 1998, 279, 558–560. [Google Scholar] [CrossRef]
- Nimnual, A.S. Coupling of Ras and Rac Guanosine Triphosphatases Through the Ras Exchanger Sos. Science 1998, 279, 560–563. [Google Scholar] [CrossRef]
- Lemmon, M.A. Pleckstrin homology domains: Not just for phosphoinositides. Biochem. Soc. Trans. 2004, 32, 707–711. [Google Scholar] [CrossRef]
- Shi, L. Dock protein family in brain development and neurological disease. Commun. Integr. Biol. 2013, 6, e26839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goicoechea, S.M.; Awadia, S.; Garcia-Mata, R. I’m coming to GEF you: Regulation of RhoGEFs during cell migration. Cell Adhes. Migr. 2014, 8, 535–549. [Google Scholar] [CrossRef]
- Chang, L.; Yang, J.; Jo, C.H.; Boland, A.; Zhang, Z.; McLaughlin, S.H.; Abu-Thuraia, A.; Killoran, R.C.; Smith, M.J.; Côté, J.F.; et al. Structure of the DOCK2−ELMO1 complex provides insights into regulation of the auto-inhibited state. Nat. Commun. 2020, 11, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Laurin, M.; Côté, J.F. Insights into the biological functions of Dock family guanine nucleotide exchange factors. Genes Dev. 2014, 28, 533–547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Côté, J.F.; Vuori, K. Identification of an evolutionary conserved superfamily of DOCK180-related proteins with guanine nucleotide exchange activity. J. Cell Sci. 2002, 115, 4901–4913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyamoto, Y.; Yamauchi, J. Cellular signaling of Dock family proteins in neural function. Cell. Signal. 2010, 22, 175–182. [Google Scholar] [CrossRef]
- Li, H.; Yang, L.; Fu, H.; Yan, J.; Wang, Y.; Guo, H.; Hao, X.; Xu, X.; Jin, T.; Zhang, N. ARTICLE Association between Gai2 and ELMO1/Dock180 connects chemokine signalling with Rac activation and metastasis. Nat. Commun. 2013. [Google Scholar] [CrossRef] [Green Version]
- Cimino, P.J.; Sokal, I.; Leverenz, J.; Fukui, Y.; Montine, T.J. DOCK2 is a microglial specific regulator of central nervous system innate immunity found in normal and Alzheimer’s disease brain. Am. J. Pathol. 2009, 175, 1622–1630. [Google Scholar] [CrossRef] [Green Version]
- Cimino, P.J.; Yang, Y.; Li, X.; Hemingway, J.F.; Cherne, M.K.; Khademi, S.B.; Fukui, Y.; Montine, K.S.; Montine, T.J.; Keene, C.D. Ablation of the microglial protein DOCK2 reduces amyloid burden in a mouse model of Alzheimer’s disease. Exp. Mol. Pathol. 2013, 94, 366–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janssen, E.; Tohme, M.; Hedayat, M.; Leick, M.; Kumari, S.; Ramesh, N.; Massaad, M.J.; Ullas, S.; Azcutia, V.; Goodnow, C.C.; et al. A DOCK8-WIP-WASp complex links T cell receptors to the actin cytoskeleton. J. Clin. Investig. 2016, 126, 3837–3851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eberth, A.; Dvorsky, R.; Becker, C.F.W.; Beste, A.; Goody, R.S.; Ahmadian, M.R. Monitoring the real-time kinetics of the hydrolysis reaction of guanine nucleotide-binding proteins. Biol. Chem. 2005, 386, 1105–1114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fidyk, N.J.; Cerione, R.A. Understanding the catalytic mechanism of GTPase-activating proteins: Demonstration of the importance of switch domain stabilization in the stimulation of GTP hydrolysis. Biochemistry 2002, 41, 15644–15653. [Google Scholar] [CrossRef] [PubMed]
- Amin, E.; Jaiswal, M.; Derewenda, U.; Reis, K.; Nouri, K.; Koessmeier, K.T.; Aspenström, P.; Somlyo, A.V.; Dvorsky, R.; Ahmadian, M.R. Deciphering the molecular and functional basis of RHOGAP family proteins: A systematic approach toward selective inactivation of RHO family proteins. J. Biol. Chem. 2016, 291, 20353–20371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheffzek, K.; Ahmadian, M.R. GTPase activating proteins: Structural and functional insights 18 years after discovery. Cell. Mol. Life Sci. 2005, 62, 3014–3038. [Google Scholar] [CrossRef] [PubMed]
- Scheffzek, K.; Ahmadian, M.R.; Kabsch, W.; Wiesmüller, L.; Lautwein, A.; Schmitz, F.; Wittinghofer, A. The Ras-RasGAP complex: Structural basis for GTPase activation and its loss in oncogenic ras mutants. Science 1997, 277, 333–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaiswal, M.; Dvorsky, R.; Amin, E.; Risse, S.L.; Fansa, E.K.; Zhang, S.C.; Taha, M.S.; Gauhar, A.R.; Nakhaei-Rad, S.; Kordes, C.; et al. Functional cross-talk between ras and rho pathways: A ras-specific gtpase-activating protein (p120RasGAP) competitively inhibits the rhogap activity of deleted in liver cancer (DLC) tumor suppressor by masking the catalytic arginine finger. J. Biol. Chem. 2014, 289, 6839–6849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.Y.; Guan, M.; Vigil, D.; Der, C.J.; Lowy, D.R.; Popescu, N.C. p120Ras-GAP binds the DLC1 Rho-GAP tumor suppressor protein and inhibits its RhoA GTPase and growth-suppressing activities. Oncogene 2009, 28, 1401–1409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garrett, M.D.; Self, A.J.; Van Oers, C.; Hall, A. Identification of distinct cytoplasmic targets for ras/R-ras and rho regulatory proteins. J. Biol. Chem. 1989, 264, 10–13. [Google Scholar] [CrossRef]
- Eberth, A.; Lundmark, R.; Gremer, L.; Dvorsky, R.; Koessmeier, K.T.; McMahon, H.T.; Ahmadian, M.R. A BAR domain-mediated autoinhibitory mechanism for RhoGAPs of the GRAF family. Biochem. J. 2009, 417, 371–377. [Google Scholar] [CrossRef] [Green Version]
- Minoshima, Y.; Kawashima, T.; Hirose, K.; Tonozuka, Y.; Kawajiri, A.; Bao, Y.C.; Deng, X.; Tatsuka, M.; Narumiya, S.; May, W.S.; et al. Phosphorylation by Aurora B converts MgcRacGAP to a RhoGAP during cytokinesis. Dev. Cell 2003, 4, 549–560. [Google Scholar] [CrossRef] [Green Version]
- Ligeti, E.; Dagher, M.C.; Hernandez, S.E.; Koleske, A.J.; Settleman, J. Phospholipids Can Switch the GTPase Substrate Preference of a GTPase-activating Protein. J. Biol. Chem. 2004, 279, 5055–5058. [Google Scholar] [CrossRef] [Green Version]
- Ahmadian, M.R.; Mittal, R.; Hall, A.; Wittinghofer, A. Aluminium fluoride associates with the small guanine nucleotide binding proteins. FEBS Lett. 1997, 408, 315–318. [Google Scholar] [CrossRef] [Green Version]
- Graham, D.L.; Eccleston, J.F.; Lowe, P.N. The conserved arginine in Rho-GTPase-activating protein is essential for efficient catalysis but not for complex formation with Rho·GDP and aluminum fluoride. Biochemistry 1999, 38, 985–991. [Google Scholar] [CrossRef] [PubMed]
- Trahey, M.; Mccormick, F. A cytoplasmic protein stimulates normal N-ras p21 GTPase, but does not affect oncogenic mutants. Science 1987, 238, 542–545. [Google Scholar] [CrossRef]
- Diekmann, D.; Brill, S.; Garrett, M.D.; Totty, N.; Hsuan, J.; Monfries, C.; Hall, C.; Lim, L.; Hall, A. Bcr encodes a GTPase-activating protein for p21rac. Nature 1991, 351, 400–402. [Google Scholar] [CrossRef]
- Lancaster, C.A.; Taylor-Harris, P.M.; Self, A.J.; Brill, S.; Van Erp, H.E.; Hall, A. Characterization of rhoGAP. A GTPase-activating protein for rho-related small GTPases. J. Biol. Chem. 1994, 269, 1137–1142. [Google Scholar] [CrossRef]
- Rittinger, K.; Taylor, W.R.; Smerdon, S.J.; Gamblin, S.J. Support for shared ancestry of GAPs. Nature 1998, 392, 448–449. [Google Scholar] [CrossRef]
- Scheffzek, K.; Ahmadian, M.R.; Wittinghofer, A. GTPase-activating proteins: Helping hands to complement an active site. Trends Biochem. Sci. 1998, 23, 257–262. [Google Scholar] [CrossRef]
- Scheffzek, K.; Lautwein, A.; Kabscht, W.; Ahmadian, M.R.; Wittinghofer, A. Crystal structure of the GTPase-activating domain of human p120GAP and implications for the interaction with Ras. Nature 1996, 384, 591–596. [Google Scholar] [CrossRef]
- Barrett, T.; Xiao, B.; Dodson, E.J.; Dodson, G.; Ludbrook, S.B.; Nurmahomed, K.; Gamblin, S.J.; Musacchio, A.; Smerdon, S.J.; Eccleston, J.F. The structure of the GTPase-activating domain from p50rhoGAP. Nature 1997, 385, 458–461. [Google Scholar] [CrossRef]
- Graham, D.L.; Lowe, P.N.; Grime, G.W.; Marsh, M.; Rittinger, K.; Smerdon, S.J.; Gamblin, S.J.; Eccleston, J.F. MgF3- as a Transition State Analog of Phosphoryl Transfer. Chem. Biol. 2002, 9, 375–381. [Google Scholar] [CrossRef]
- Vetter, I.R.; Wittinghofer, A. The guanine nucleotide-binding switch in three dimensions. Science 2001, 294, 1299–1304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rittinger, K.; Walker, P.A.; Eccleston, J.F.; Nurmahomed, K.; Owen, D.; Laue, E.; Gamblin, S.J.; Smerdon, S.J. Crystal structure of a small G protein in complex with the GTPase- activating protein rhoGAP. Nature 1997, 388, 693–697. [Google Scholar] [CrossRef]
- Rittinger, K.; Walker, P.A.; Eccleston, J.F.; Smerdon, S.J.; Gamblin, S.J. Structure at 1.65 Å of RhoA and its GTPase-activating protein in complex with a transition-state analogue. Nature 1997, 389, 758–762. [Google Scholar] [CrossRef] [PubMed]
- Jefferson, A.B.; Majerus, P.W. Properties of type II inositol polyphosphate 5-phosphatase. J. Biol. Chem. 1995, 270, 9370–9377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernards, A. GAPs galore! A survey of putative Ras superfamily GTPase activating proteins in man and Drosophila. Biochim. Biophys. Acta Rev. Cancer 2003, 1603, 47–82. [Google Scholar] [CrossRef]
- Amin, E.; Dubey, B.N.; Zhang, S.C.; Gremer, L.; Dvorsky, R.; Moll, J.M.; Taha, M.S.; Nagel-Steger, L.; Piekorz, R.P.; Somlyo, A.V.; et al. Rho-kinase: Regulation, (dys)function, and inhibition. Biol. Chem. 2013, 394, 1399–1410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narumiya, S.; Tanji, M.; Ishizaki, T. Rho signaling, ROCK and mDia1, in transformation, metastasis and invasion. Cancer Metastasis Rev. 2009, 28, 65–76. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.S.; Manser, E. PAK and other Rho-associated kinases - Effectors with surprisingly diverse mechanisms of regulation. Biochem. J. 2005, 386, 201–214. [Google Scholar] [CrossRef]
- Hedman, A.C.; Smith, J.M.; Sacks, D.B. The biology of IQGAP proteins: Beyond the cytoskeleton. EMBO Rep. 2015, 16, 427–446. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.A.; Wang, M.J.; Chi, C.W.; Wu, C.W.; Chen, J.Y. Rho/Rhotekin-mediated NF-κB activation confers resistance to apoptosis. Oncogene 2004, 23, 8731–8742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maesaki, R.; Ihara, K.; Shimizu, T.; Kuroda, S.; Kaibuchi, K.; Hakoshima, T. The structural basis of Rho effector recognition revealed by the crystal structure of human RhoA complexed with the effector domain of PKN/PRK1. Mol. Cell 1999, 4, 793–803. [Google Scholar] [CrossRef]
- Dvorsky, R.; Blumenstein, L.; Vetter, I.R.; Ahmadian, M.R. Structural Insights into the Interaction of ROCKI with the Switch Regions of RhoA. J. Biol. Chem. 2004, 279, 7098–7104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdul-Manan, N.; Aghazadeh, B.; Liu, G.A.; Majumdar, A.; Ouerfelli, O.; Simlnovitch, K.A.; Rosen, M.K. Structure of Cdc42 in complex with the GTPase-binding domain of the “Wiskott-Aldrich syndrome” protein. Nature 1999, 399, 379–383. [Google Scholar] [CrossRef]
- Mott, H.R.; Owen, D.; Nietlispach, D.; Lowe, P.N.; Manser, E.; Lim, L.; Laue, E.D. Structure of the small G protein Cdc42 bound to the GTpasebinding domain of ACK. Nature 1999, 399, 384–388. [Google Scholar] [CrossRef] [PubMed]
- Gizachew, D.; Guo, W.; Chohan, K.K.; Sutcliffe, M.J.; Oswald, R.E. Structure of the complex of Cdc42Hs with a peptide derived from P-21 activated kinase. Biochemistry 2000, 39, 3963–3971. [Google Scholar] [CrossRef]
- Morreale, A.; Venkatesan, M.; Mott, H.R.; Owen, D.; Nietlispach, D.; Lowe, P.N.; Laue, E.D. Structure of Cdc42 bound to the GTPase binding domain of PAK. Nat. Struct. Biol. 2000, 7, 384–388. [Google Scholar] [CrossRef]
- Garrard, S.M.; Capaldo, C.T.; Gao, L.; Rosen, M.K.; Macara, I.G.; Tomchick, D.R. Structure of Cdc42 in a complex with the GTPase-binding domain of the cell polarity protein, Par6. EMBO J. 2003, 22, 1125–1133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinelli, S.; Krumbach, O.H.F.; Pantaleoni, F.; Coppola, S.; Amin, E.; Pannone, L.; Nouri, K.; Farina, L.; Dvorsky, R.; Lepri, F.; et al. Functional Dysregulation of CDC42 Causes Diverse Developmental Phenotypes. Am. J. Hum. Genet. 2018, 102, 309–320. [Google Scholar] [CrossRef] [Green Version]
- Lapouge, K.; Smith, S.J.M.; Walker, P.A.; Gamblin, S.J.; Smerdon, S.J.; Rittinger, K. Structure of the TPR domain of p67(phox) in complex with Rac·GTP. Mol. Cell 2000, 6, 899–907. [Google Scholar] [CrossRef]
- Tarricone, C.; Xiao, B.; Justin, N.; Walker, P.A.; Rittinger, K.; Gamblin, S.J.; Smerdon, S.J. The structural basis of Arfaptin-mediated cross-talk between Rac and Arf signalling pathways. Nature 2001, 411, 215–219. [Google Scholar] [CrossRef]
- Cherfils, J. Structural mimicry of DH domains by Arfaptin suggests a model for the recognition of Rac-GDP by its guanine nucleotide exchange factors. FEBS Lett. 2001, 507, 280–284. [Google Scholar] [CrossRef] [Green Version]
- Diebold, B.A.; Bokoch, G.M. Molecular basis for Rac2 regulation of phagocyte NADPH oxidase. Nat. Immunol. 2001, 2, 211–215. [Google Scholar] [CrossRef]
- Hoffman, G.R.; Cerione, R.A. Rac inserts its way into the immune response. Nat. Immunol. 2001, 2, 194–196. [Google Scholar] [CrossRef] [PubMed]
- Blumenstein, L.; Ahmadian, M.R. Models of the cooperative mechanism for Rho effector recognition: Implications for RhoA-mediated effector activation. J. Biol. Chem. 2004, 279, 53419–53426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, A. Rho GTpases and the actin cytoskeleton. Science 1998, 279, 509–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawson, C.D.; Ridley, A.J. Rho GTPase signaling complexes in cell migration and invasion. J. Cell Biol. 2018, 217, 447–457. [Google Scholar] [CrossRef] [PubMed]
- Guan, X.; Guan, X.; Dong, C.; Jiao, Z. Rho GTPases and related signaling complexes in cell migration and invasion. Exp. Cell Res. 2020, 388. [Google Scholar] [CrossRef]
- Hall, A. Rho family GTPases. Biochem. Soc. Trans. 2012, 40, 1378–1382. [Google Scholar] [CrossRef] [Green Version]
- Schaks, M.; Giannone, G.; Rottner, K. Actin dynamics in cell migration. Essays Biochem. 2019, 63, 483–495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devreotes, P.; Horwitz, A.R. Signaling networks that regulate cell migration. Cold Spring Harb. Perspect. Biol. 2015, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zegers, M.M.; Friedl, P. Rho GTPases in collective cell migration. Small GTPases 2014, 5. [Google Scholar] [CrossRef]
- Sadok, A.; Marshall, C.J. Rho gtpases masters of cell migration. Small GTPases 2014, 5. [Google Scholar] [CrossRef] [Green Version]
- Raftopoulou, M.; Hall, A. Cell migration: Rho GTPases lead the way. Dev. Biol. 2004, 265, 23–32. [Google Scholar] [CrossRef] [Green Version]
- Miralles, F.; Posern, G.; Zaromytidou, A.I.; Treisman, R. Actin dynamics control SRF activity by regulation of its coactivator MAL. Cell 2003, 113, 329–342. [Google Scholar] [CrossRef] [Green Version]
- Hordijk, P.L. Regulation of NADPH oxidases: The role of Rac proteins. Circ. Res. 2006, 98, 453–462. [Google Scholar] [CrossRef]
- Voncken, J.W.; van Schaick, H.; Kaartinen, V.; Deemer, K.; Coates, T.; Landing, B.; Pattengale, P.; Dorseuil, O.; Bokoch, G.M.; Groffen, J.; et al. Increased neutrophil respiratory burst in bcr-null mutants. Cell 1995, 80, 719–728. [Google Scholar] [CrossRef] [Green Version]
- Lyon, A.S.; Peeples, W.B.; Rosen, M.K. A framework for understanding the functions of biomolecular condensates across scales. Nat. Rev. Mol. Cell Biol. 2021, 22, 215–235. [Google Scholar] [CrossRef] [PubMed]
- Araki, Y.; Hong, I.; Gamache, T.R.; Ju, S.; Collado-Torres, L.; Shin, J.H.; Huganir, R.L. Syngap isoforms differentially regulate synaptic plasticity and dendritic development. Elife 2020, 9, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Pudewell, S.; Wittich, C.; Kazemein Jasemi, N.S.; Bazgir, F.; Ahmadian, M.R. Accessory proteins of the RAS-MAPK pathway: Moving from the side line to the front line. Commun. Biol. 2021, 4, 696. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Banjade, S.; Cheng, H.C.; Kim, S.; Chen, B.; Guo, L.; Llaguno, M.; Hollingsworth, J.V.; King, D.S.; Banani, S.F.; et al. Phase transitions in the assembly of multivalent signalling proteins. Nature 2012, 483, 336–340. [Google Scholar] [CrossRef] [PubMed]
- Case, L.B.; Zhang, X.; Ditlev, J.A.; Rosen, M.K. Stoichiometry controls activity of phase-separated clusters of actin signaling proteins. Science 2019, 363, 1093–1097. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Molecular principles of RHO GTPase regulation and signaling. Most RHO GTPases (20 canonical human members) act as molecular switches by cycling between a GDP-bound, inactive state and a GTP-bound, active state. They interact specifically with four structurally and functionally unrelated classes of proteins: (A) In resting cells, guanine nucleotide dissociation inhibitors (GDIs; 4 human members) sequester RHO in the cytoplasm, away from the membrane, by binding to the lipid anchor and thus creating an inactivated cytosolic pool; (B) in stimulated cells, different classes of membrane receptors activate guanine nucleotide exchange factors (GEFs; 85 human members: 74 DBL and 11 DOCK family proteins), which in turn activate RHO by accelerating the intrinsic exchange of GDP for GTP and switch ON signal transduction; (C) active GTP-bound RHO interacts through the GTPase-binding domain (GBD) with and activates downstream targets (effectors; >70 human members) to cause a variety of intracellular pathways, which control a multitude of biochemical processes involved in the regulation of different biological (dys)functions; (D) GTPase-activating proteins (GAPs; 66 human members) negatively regulate RHO by stimulating its slow intrinsic GTP hydrolysis activity and switch OFF signal transduction. Notably, all RHO-interacting proteins recognize and bind RHO at consensus-binding sites called switch I and II.
Figure 1.
Molecular principles of RHO GTPase regulation and signaling. Most RHO GTPases (20 canonical human members) act as molecular switches by cycling between a GDP-bound, inactive state and a GTP-bound, active state. They interact specifically with four structurally and functionally unrelated classes of proteins: (A) In resting cells, guanine nucleotide dissociation inhibitors (GDIs; 4 human members) sequester RHO in the cytoplasm, away from the membrane, by binding to the lipid anchor and thus creating an inactivated cytosolic pool; (B) in stimulated cells, different classes of membrane receptors activate guanine nucleotide exchange factors (GEFs; 85 human members: 74 DBL and 11 DOCK family proteins), which in turn activate RHO by accelerating the intrinsic exchange of GDP for GTP and switch ON signal transduction; (C) active GTP-bound RHO interacts through the GTPase-binding domain (GBD) with and activates downstream targets (effectors; >70 human members) to cause a variety of intracellular pathways, which control a multitude of biochemical processes involved in the regulation of different biological (dys)functions; (D) GTPase-activating proteins (GAPs; 66 human members) negatively regulate RHO by stimulating its slow intrinsic GTP hydrolysis activity and switch OFF signal transduction. Notably, all RHO-interacting proteins recognize and bind RHO at consensus-binding sites called switch I and II.

Figure 3.
Domain organization of DBL family proteins. The DBL family RHOGEFs are mostly multimodular proteins and have a number of functional domains that may mediate cross talk between RHO proteins and other signaling pathways. DH domains are almost always found with a PH domain in the C-terminus. Some DBL proteins contain two DH–PH cassettes, while some DBL proteins lack tandem PH domains. Functional domains, in addition to the catalytic DH domain (green), are probably involved in lipid and membrane binding (blue), protein interactions (bright green), and enzymatic activities (red and orange). A scale of amino acid numbers in increments of 200 is shown at the bottom. Underlined proteins do not exhibit activity under cell-free conditions [47].
Figure 3.
Domain organization of DBL family proteins. The DBL family RHOGEFs are mostly multimodular proteins and have a number of functional domains that may mediate cross talk between RHO proteins and other signaling pathways. DH domains are almost always found with a PH domain in the C-terminus. Some DBL proteins contain two DH–PH cassettes, while some DBL proteins lack tandem PH domains. Functional domains, in addition to the catalytic DH domain (green), are probably involved in lipid and membrane binding (blue), protein interactions (bright green), and enzymatic activities (red and orange). A scale of amino acid numbers in increments of 200 is shown at the bottom. Underlined proteins do not exhibit activity under cell-free conditions [47].

Figure 4.
Domain organization of the RHOGAP family proteins (adapted from Amin et al., 2016 [114]. RHOGAPs are mostly multimodular proteins and have a number of functional domains that may mediate cross talk between RHO proteins and other signaling pathways. Functional domains, in addition to the catalytic GAP domain (red), are probably involved in lipid and membrane binding (blue), protein interactions (bright green), and enzymatic activities (red and orange). A scale of amino acid numbers in increments of 200 is shown at the bottom. Underlined proteins are GAP-like proteins with no RHOGAP activity [114].
Figure 4.
Domain organization of the RHOGAP family proteins (adapted from Amin et al., 2016 [114]. RHOGAPs are mostly multimodular proteins and have a number of functional domains that may mediate cross talk between RHO proteins and other signaling pathways. Functional domains, in addition to the catalytic GAP domain (red), are probably involved in lipid and membrane binding (blue), protein interactions (bright green), and enzymatic activities (red and orange). A scale of amino acid numbers in increments of 200 is shown at the bottom. Underlined proteins are GAP-like proteins with no RHOGAP activity [114].

Figure 5.
Regulation of actin-based motility by RHOA, RAC1, and CDC42. Activated CDC42, RAC, and RHO bind to and specifically activate their downstream effectors, which are either kinases (e.g., ROCK, PAK, and PI5K) or scaffolding proteins (e.g., DIA, WASP, IRSp53, and IQGAP). These effector proteins activate diverse signaling pathways with distinct effects on the actin cytoskeleton and cellular morphology. An important aspect of cell motility is the equilibrium between the myosin light chain (MLC) and phosphorylated MLC, which is tightly regulated.
Figure 5.
Regulation of actin-based motility by RHOA, RAC1, and CDC42. Activated CDC42, RAC, and RHO bind to and specifically activate their downstream effectors, which are either kinases (e.g., ROCK, PAK, and PI5K) or scaffolding proteins (e.g., DIA, WASP, IRSp53, and IQGAP). These effector proteins activate diverse signaling pathways with distinct effects on the actin cytoskeleton and cellular morphology. An important aspect of cell motility is the equilibrium between the myosin light chain (MLC) and phosphorylated MLC, which is tightly regulated.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Mosaddeghzadeh, N.; Ahmadian, M.R. The RHO Family GTPases: Mechanisms of Regulation and Signaling. Cells 2021, 10, 1831. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10071831
AMA Style
Mosaddeghzadeh N, Ahmadian MR. The RHO Family GTPases: Mechanisms of Regulation and Signaling. Cells. 2021; 10(7):1831. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10071831
Chicago/Turabian StyleMosaddeghzadeh, Niloufar, and Mohammad Reza Ahmadian. 2021. "The RHO Family GTPases: Mechanisms of Regulation and Signaling" Cells 10, no. 7: 1831. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10071831
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.