
The Parkinson’s Disease-Associated Protein DJ-1 Protects Dictyostelium Cells from AMPK-Dependent Outcomes of Oxidative Stress


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plasmid Constructs
2.2. D. discoideum Strains and Culture Conditions
2.3. Molecular Manipulation of DNA and RNA
2.3.1. Quantitative PCR (qPCR)
2.3.2. One-Time Reverse Transcriptase-PCR (RT-PCR)
2.4. Cotransformation of Dictyostelium
2.5. Phenotypic Characterization
2.5.1. Seahorse Respirometry
2.5.2. Phototaxis
2.5.3. Growth in Axenic Liquid Medium
2.5.4. Morphogenesis
2.5.5. Phagocytosis and Pinocytosis
2.6. Statistical Analysis
3. Results
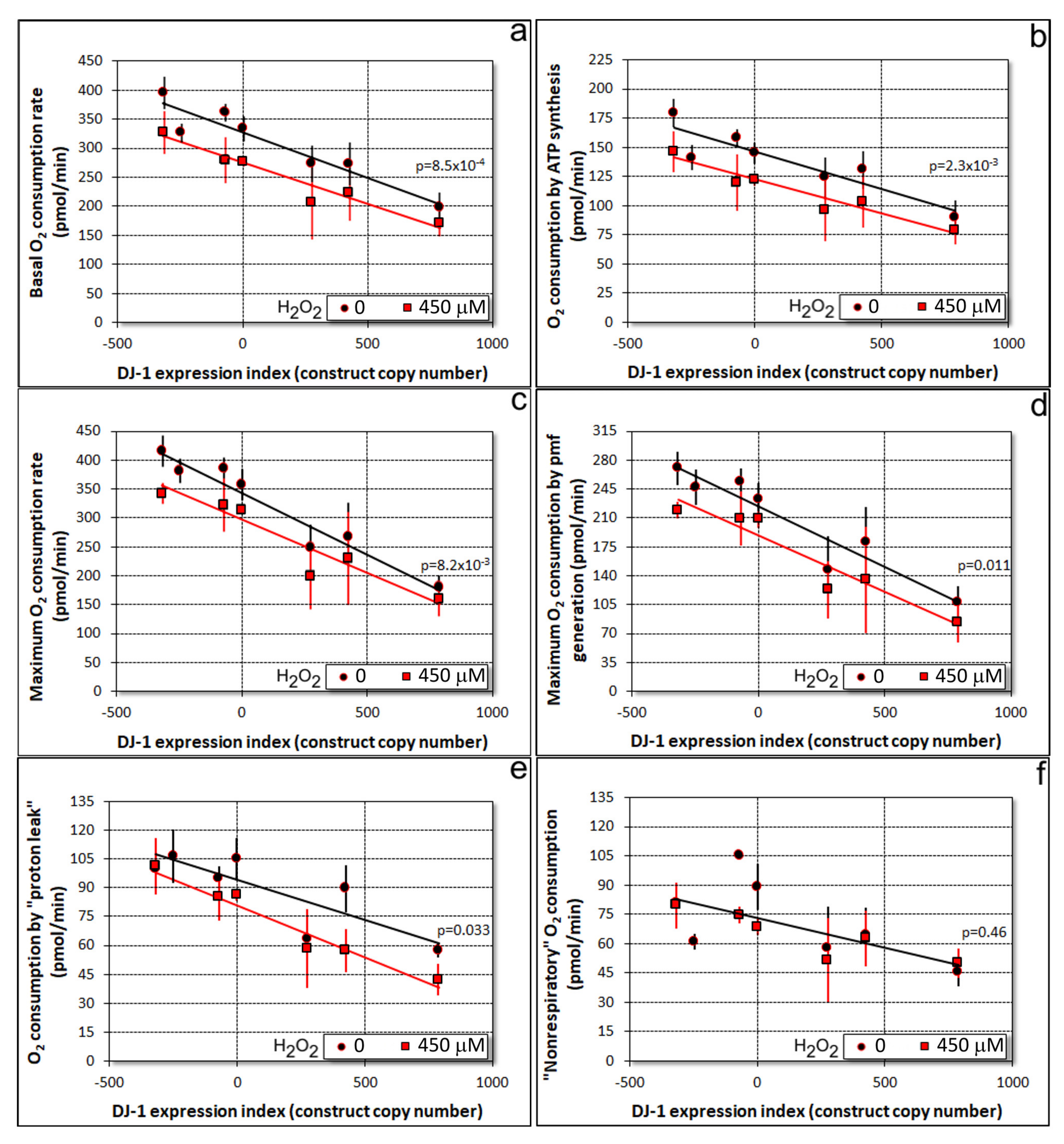
3.1. Oxidative Stress Impairs Mitochondrial Respiration Independently of DJ-1
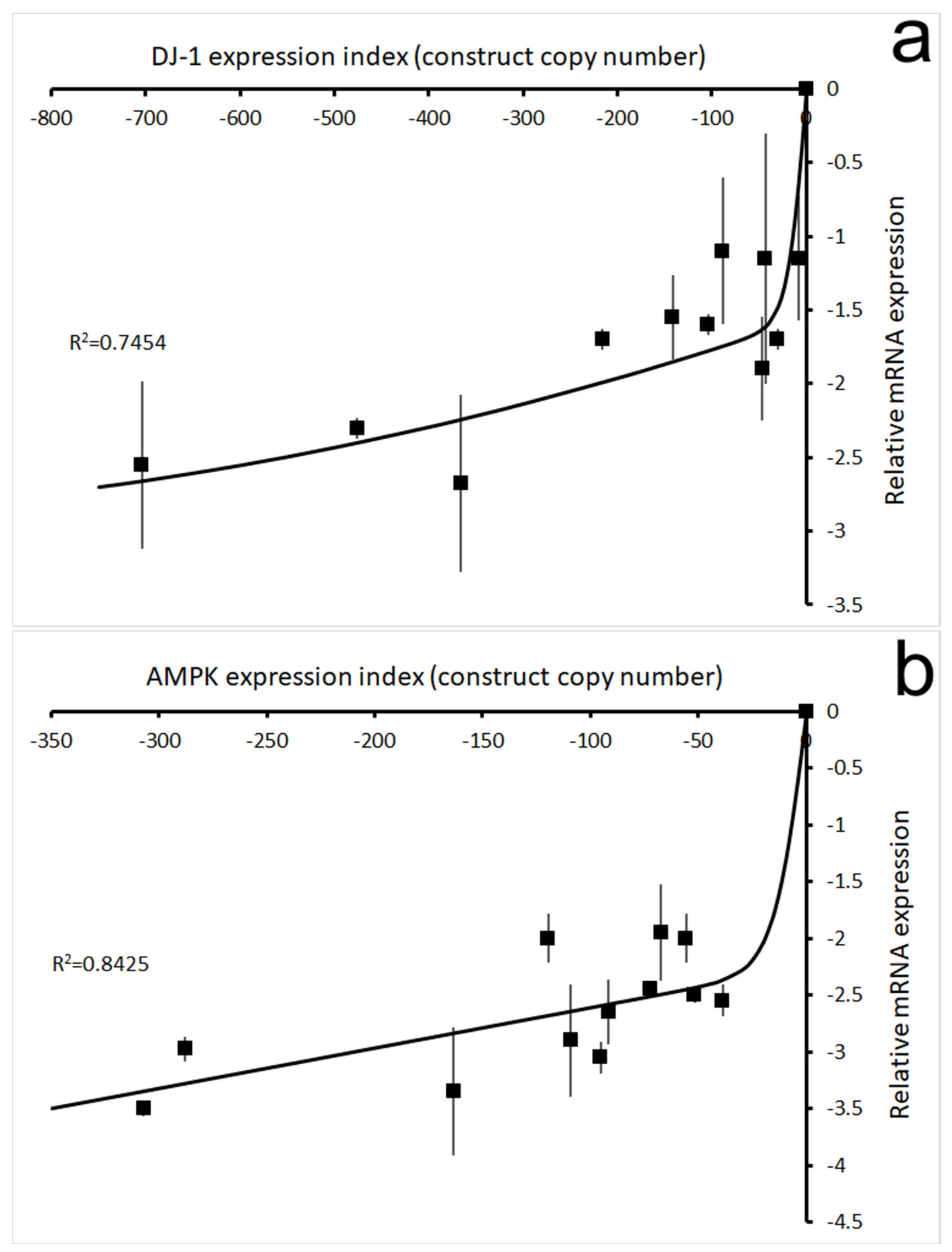
3.2. Under Oxidative Stress, Knockdown of DJ-1 Produced AMPK-Mediated Phenotypic Defects in D. discoideum
3.3. The Combination of Oxidative Stress and Reduced Levels of DJ-1 Causes a Phototaxis Defect That Is AMPK-Mediated
3.4. Oxidative Stress Causes an AMPK-Dependent Exacerbation of the Phagocytosis Defect Produced by DJ-1 Loss
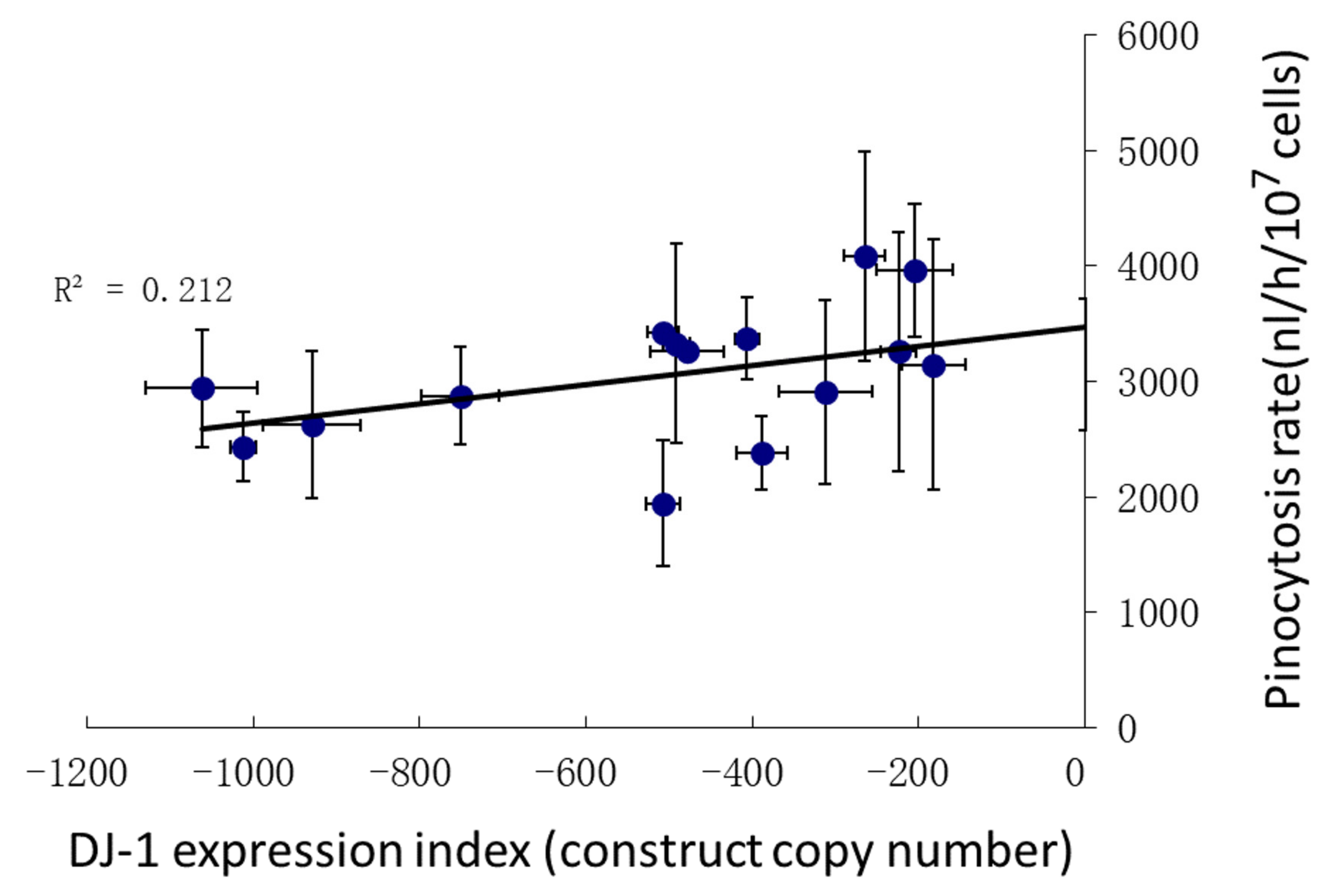
3.5. Pinocytosis Is Not Affected Significantly by DJ-1 Knockdown under Conditions of Oxidative Stress
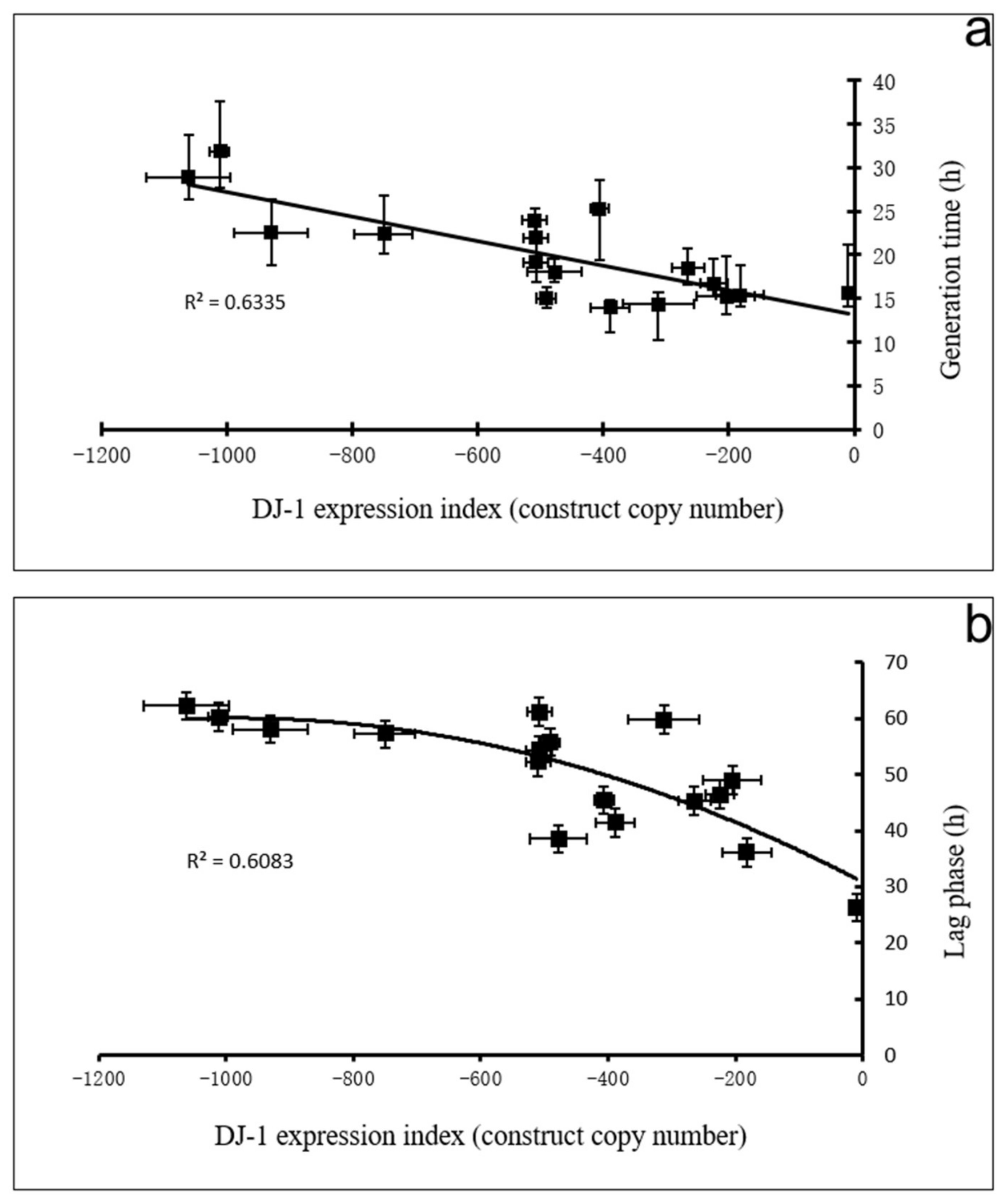
3.6. DJ-1 Protects Cells from AMPK-Dependent Inhibition of Growth Caused by Oxidative Stress
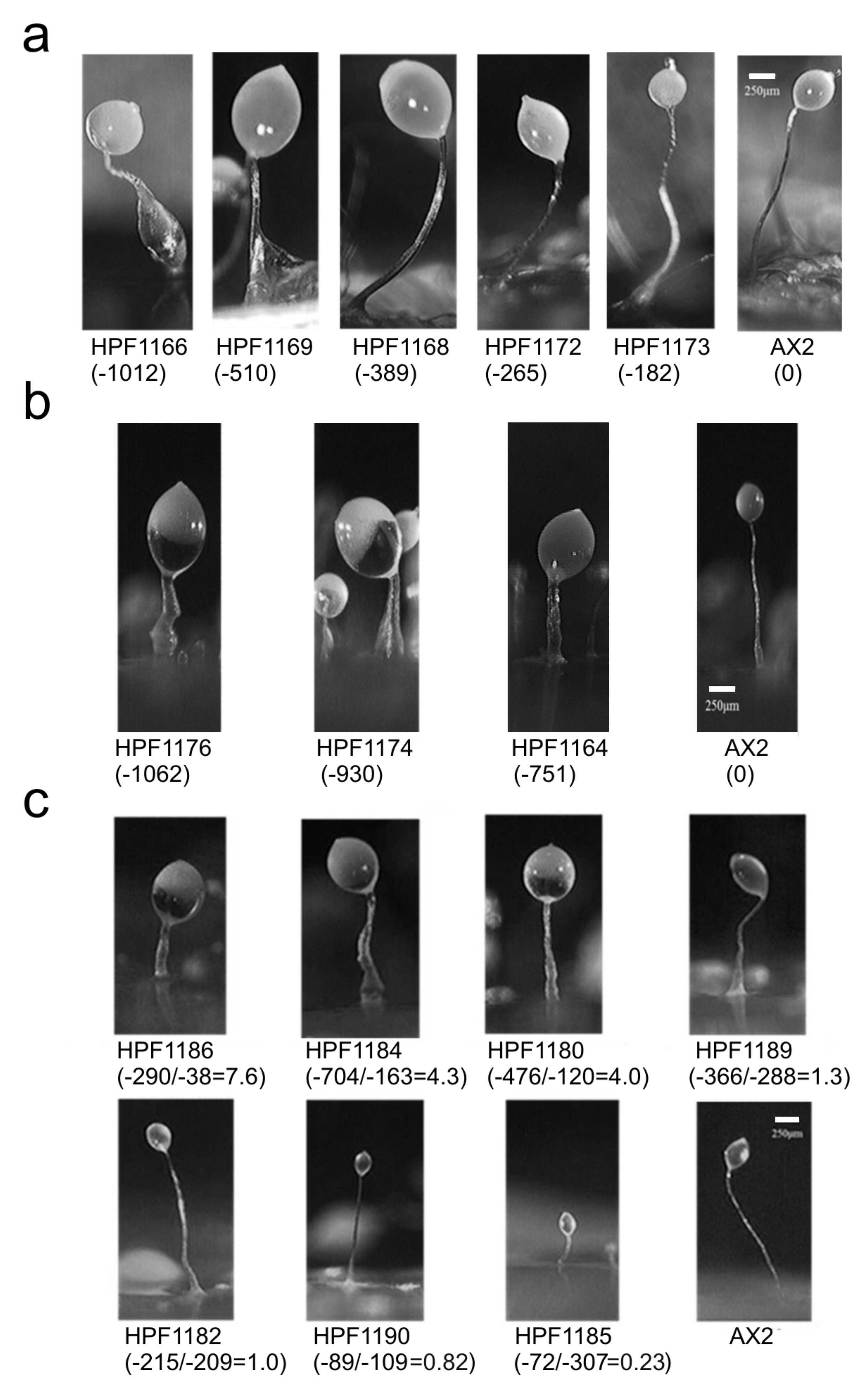
3.7. DJ-1, AMPK and Oxidative Stress Interact in Regulating Fruiting Body Morphology
4. Discussion
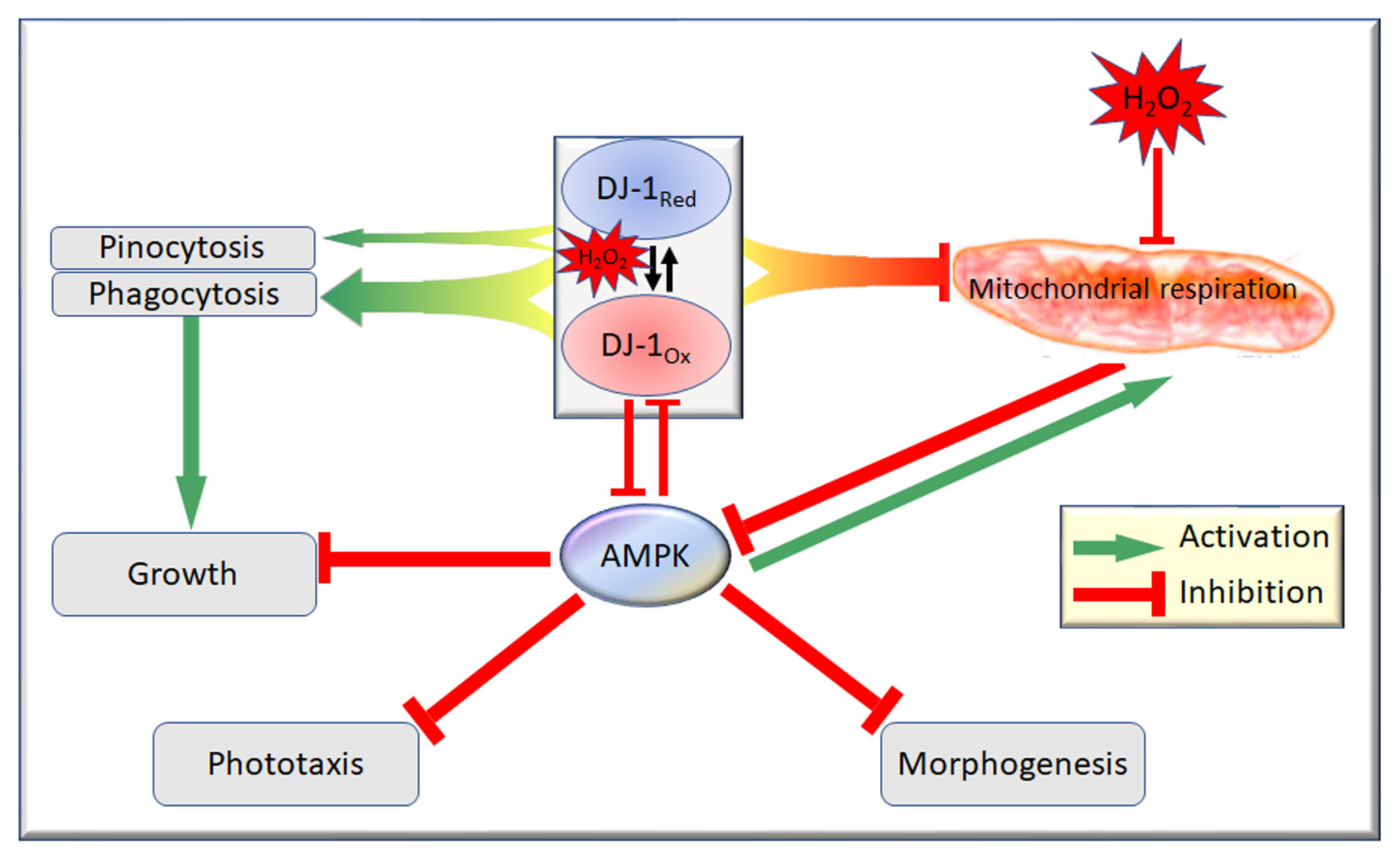
- Regardless of its oxidation state, DJ-1 is an activator of phagocytosis [18] (Figure 4, Supplementary Figure S4) and also slightly activates pinocytosis [18] (Figure 5, Supplementary Figure S5). This elicits corresponding effects on the growth on bacterial lawns and in liquid medium [18] (Figure 6 and Figure 7; Supplementary Figures S4 and S5).
- In its reduced state, DJ-1 has little or no interaction with AMPK. In unstressed cells most DJ-1 is in the reduced state; therefore, altering its expression exerts no detectable effect on phototaxis (Supplementary Figure S3) and relatively small effects on morphogenesis (Supplementary Figure S6) [18]. For the same reason, in the absence of oxidative stress, AMPK activation by mitochondrial disease has no detectable effect on phagocytosis or pinocytosis [30].
- In its oxidized state, DJ-1 becomes sensitive to inhibition by AMPK. As a consequence of this, oxidative stress contributes to an AMPK-mediated impairment of phagocytosis (Figure 4), additional to that caused by DJ-1 loss. AMPK antisense-inhibition reverses the effect of oxidative stress on phagocytosis without significantly altering the inhibitory effect of DJ-1 knockdown (Figure 4). The same probably occurs in relation to pinocytosis, which was already shown to be slightly inhibited by DJ-1 loss [18], although this could not be detected statistically in the present experiments (Figure 5). The magnitude of this AMPK-dependent effect is in proportion to the relative magnitude of DJ-1′s influence on these two types of endocytosis [18].
- In its oxidized state, DJ-1 becomes capable of inhibiting AMPK. As a consequence of this, it opposes and limits the activation of AMPK caused by oxidative stress. For this reason, wild type DJ-1 at normal levels prevents oxidative stress from causing dramatic defects in phototaxis, morphogenesis or growth in liquid. However, the loss of DJ-1 allows for the unrestrained activation of AMPK by oxidative stress; therefore, the combination produces a dramatic AMPK-dependent impairment of growth (Figure 6 and Figure 7), phototaxis (Figure 3) and morphogenesis (Figure 8).
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- van Duijn, C.M.; Dekker, M.C.; Bonifati, V.; Galjaard, R.J.; Houwing-Duistermaat, J.J.; Snijders, P.J.; Testers, L.; Breedveld, G.J.; Horstink, M.; Sandkuijl, L.A.; et al. Park7, a novel locus for autosomal recessive early-onset parkinsonism, on chromosome 1p36. Am. J. Hum. Genet. 2001, 69, 629–634. [Google Scholar] [CrossRef] [Green Version]
- Bonifati, V.; Rizzu, P.; van Baren, M.J.; Schaap, O.; Breedveld, G.J.; Krieger, E.; Dekker, M.C.; Squitieri, F.; Ibanez, P.; Joosse, M.; et al. Mutations in the DJ-1 gene associated with autosomal recessive early-onset Parkinsonism. Science 2003, 299, 256–259. [Google Scholar] [CrossRef] [Green Version]
- Hedrich, K.; Djarmati, A.; Schafer, N.; Hering, R.; Wellenbrock, C.; Weiss, P.H.; Hilker, R.; Vieregge, P.; Ozelius, L.J.; Heutink, P.; et al. DJ-1 (PARK7) mutations are less frequent than Parkin (PARK2) mutations in early-onset Parkinson disease. Neurology 2004, 62, 389–394. [Google Scholar] [CrossRef]
- Shendelman, S.; Jonason, A.; Martinat, C.; Leete, T.; Abeliovich, A. DJ-1 is a redox-dependent molecular chaperone that inhibits alpha-synuclein aggregate formation. PLoS Biol. 2004, 2, e362. [Google Scholar] [CrossRef] [Green Version]
- Zhou, W.; Zhu, M.; Wilson, M.A.; Petsko, G.A.; Fink, A.L. The oxidation state of DJ-1 regulates its chaperone activity toward alpha-synuclein. J. Mol. Biol. 2006, 356, 1036–1048. [Google Scholar] [CrossRef]
- Xu, J.; Zhong, N.; Wang, H.; Elias, J.E.; Kim, C.Y.; Woldman, I.; Pifl, C.; Gygi, S.P.; Geula, C.; Yankner, B.A. The Parkinson’s disease-associated DJ-1 protein is a transcriptional co-activator that protects against neuronal apoptosis. Hum. Mol. Genet. 2005, 14, 1231–1241. [Google Scholar] [CrossRef] [Green Version]
- Clements, C.M.; McNally, R.S.; Conti, B.J.; Mak, T.W.; Ting, J.P. DJ-1, a cancer- and Parkinson’s disease-associated protein, stabilizes the antioxidant transcriptional master regulator Nrf2. Proc. Natl. Acad. Sci. USA 2006, 103, 15091–15096. [Google Scholar] [CrossRef] [Green Version]
- Olzmann, J.A.; Brown, K.; Wilkinson, K.D.; Rees, H.D.; Huai, Q.; Ke, H.; Levev, A.I.; Li, L.; Chin, L.S. Familial Parkinson’s disease-associated L166P mutation disrupts DJ-1 protein folding and function. J. Biol. Chem. 2004, 279, 8506–8515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Li, L.; Chin, L.S. Parkinson disease protein DJ-1 converts from a zymogen to a protease by carboxyl-terminal cleavage. Hum. Mol. Genet. 2010, 19, 2395–2408. [Google Scholar] [CrossRef] [Green Version]
- Canet-Avilés, R.M.; Wilson, M.A.; Miller, D.W.; Ahmad, R.; McLendon, C.; Bandyopadhyay, S.; Baptista, M.J.; Ringe, D.; Petsko, G.A.; Cookson, M.R. The Parkinson’s disease protein DJ-1 is neuroprotective due to cysteine-sulfinic acid-driven mitochondrial localization. Proc. Natl. Acad. Sci. USA 2004, 101, 9103–9108. [Google Scholar] [CrossRef] [Green Version]
- Taira, T.; Saito, Y.; Niki, T.; Iguchi-Ariga, S.M.; Takahashi, K.; Ariga, H. DJ-1 has a role in antioxidative stress to prevent cell death. EMBO Rep. 2004, 5, 213–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paterna, J.C.; Leng, A.; Weber, E.; Feldon, J.; Bueler, H. DJ-1and Parkin modulate dopamine-dependent behavior and inhibit MPTP-induced nigral dopamine neuron loss in mice. Mol. Ther. 2007, 15, 698–704. [Google Scholar] [CrossRef]
- Inden, M.; Taira, T.; Kitamura, Y.; Yanagida, T.; Tsuchiya, D.; Takata, K.; Yanagisawa, D.; Nishimura, K.; Taniguchi, T.; Kiso, Y.; et al. PARK7 DJ-1 protects against degeneration of nigral dopaminergic neurons in Parkinson’s disease rat model. Neurobiol. Dis. 2006, 24, 144–158. [Google Scholar] [CrossRef] [PubMed]
- Blackinton, J.; Lakshminarasimhan, M.; Thomas, K.J.; Ahmad, R.; Greggio, E.; Raza, A.S.; Cookson, M.R.; Wilson, M.A. Formation of a stabilized cysteine sulfinic acid is critical for the mitochondrial function of the parkinsonism protein DJ-1. J. Biol. Chem. 2009, 284, 6476–6485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larsen, N.J.; Ambrosi, G.; Mullett, S.J.; Berman, S.B.; Hinkle, D.A. DJ-1 knockdown impairs astrocyte mitochondrial function. Neuroscience 2011, 196, 251–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beilina, A.; Van Der Brug, M.; Ahmad, R.; Kesavapany, S.; Miller, D.W.; Petsko, G.A.; Cookson, M.R. Mutations in PTEN-induced putative kinase 1 associated with recessive parkinsonism have differential effects on protein stability. Proc. Natl. Acad. Sci. USA 2005, 102, 5703–5708. [Google Scholar] [CrossRef] [Green Version]
- Dev, K.K.; Hofele, K.; Barbieri, S.; Buchman, V.L.; van der Putten, H. Part II: Alpha-synuclein and its molecular pathophysiological role in neurodegenerative disease. Neuropharmacology 2003, 45, 14–44. [Google Scholar] [CrossRef]
- Chen, S.; Annesley, S.J.; Jasim, R.A.F.; Musco, V.J.; Sanislav, O.; Fisher, P.R. The Parkinson’s disease-associated protein DJ-1 plays a positive nonmitochondrial role in endocytosis in Dictyostelium cells. Dis. Models Mech. 2017, 10, 1261–1271. [Google Scholar]
- Zhang, L.; Shimoji, M.; Thomas, B.; Moore, D.J.; Yu, S.W.; Marupudi, N.I.; Torp, R.; Torgner, I.A.; Ottersen, O.P.; Dawson, T.M.; et al. Mitochondrial localization of the Parkinson’s disease related protein DJ-1: Implications for pathogenesis. Hum. Mol. Genet. 2005, 14, 2063–2073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kojima, W.; Kujuro, Y.; Okatsu, K.; Bruno, Q.; Koyano, F.; Kimura, M.; Yamano, K.; Tanaka, K.; Matsuda, N. Unexpected mitochondrial matrix localization of Parkinson’s disease-related DJ-1 mutants but not wild-type DJ-1. Genes Cells 2016, 21, 772–788. [Google Scholar] [CrossRef]
- Björkblom, B.; Maple-Grødem, J.; Puno, M.R.; Odell, M.; Larsen, J.P.; Møller, S.G. Reactive Oxygen Species-mediated DJ-1 monomerization modulates intracellular trafficking involving Karyopherin β2. Mol. Cell. Biol. 2014, 34, 3024–3040. [Google Scholar] [CrossRef] [Green Version]
- Annesley, S.J.; Fisher, P.R. Dictyostelium discoideum—A model for many reasons. Mol. Cell. Biochem. 2009, 329, 73–91. [Google Scholar] [CrossRef]
- Williams, R.S.; Boeckeler, K.; Gräf, R.; Müller-Taubenberger, A.; Li, Z.; Isberg, R.R.; Wessels, D.; Soll, D.R.; Alexander, H.; Alexander, S. Towards a molecular understanding of human diseases using Dictyostelium discoideum. Trends Mol. Med. 2006, 12, 415–424. [Google Scholar] [CrossRef]
- Francione, L.M.; Annesley, S.J.; Carilla-Latorre, S.; Escalante, R.; Fisher, P.R. The Dictyostelium model for mitochondrial disease. Semin. Cell Dev. Biol. 2011, 22, 120–130. [Google Scholar] [CrossRef] [Green Version]
- Annesley, S.J.; Chen, S.; Francione, L.M.; Sanislav, O.; Chavan, A.J.; Farah, C.; De Piazza, S.W.; Storey, C.L.; Ilievska, J.; Fernando, S.G.; et al. Dictyostelium, a microbial model for brain disease. Biochim. Biophys. Acta. 2014, 1840, 1413–1432. [Google Scholar] [CrossRef]
- Fey, P.; Dodson, R.; Basu, S.; Chisholm, R.L. One Stop Shop for Everything Dictyostelium: DictyBase and the Dicty Stock Center. Methods Mol. Biol. 2013, 983, 59–92. [Google Scholar]
- Lay, S.T.; Sanislav, O.; Annesley, S.J.; Fisher, P.R. Mitochondrial stress tests using Seahorse respirometry on intact Dictyostelium discoideum cells. Methods Mol. Biol. 2016, 1407, 41–62. [Google Scholar]
- Annesley, S.J.; Lay, S.T.; De Piazza, S.W.; Sanislav, O.; Hammersley, E.; Allan, C.Y.; Francione, L.M.; Bui, M.Q.; Chen, Z.-P.; Ngoei, K.R.W.; et al. Immortalized Parkinson’s Disease lymphocytes have enhanced mitochondrial respiratory activity. Dis. Models Mech. 2016, 9, 1295–1305. [Google Scholar] [CrossRef] [Green Version]
- Haylett, W.; Swart, C.; van der Westhuizen, F.; van Dyk, H.; van der Merwe, L.; van der Merwe, C.; Loos, B.; Carr, J.; Kinnear, C.; Bardien, S. Altered mitochondrial respiration and other features of mitochondrial function in Parkin-mutant fibroblasts from Parkinson’s disease patients. Parkinson Dis. 2016, 2016, 1819209. [Google Scholar]
- Bokko, P.B.; Francione, L.; Bandala-Sanchez, E.; Ahmed, A.U.; Annesley, S.J.; Huang, X.; Khurana, T.; Kimmel, A.R.; Fisher, P.R. Diverse cytopathologies in mitochondrial disease are caused by AMP-activated protein kinase signaling. Mol. Biol. Cell. 2007, 18, 1874–1886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Witke, W.; Nellen, W.; Noegel, A. Homologous recombination in the Dictyostelium alpha-actinin gene leads to an altered mRNA and lack of the protein. EMBO J. 1987, 6, 4143–4148. [Google Scholar] [CrossRef]
- Garcia, M.X.U.; Foote, C.; van Es, S.; Devreotes, P.N.; Alexander, S.; Alexander, H. Differential developmental expression and cell type specificity of Dictyostelium catalases and their response to oxidative stress and UV-light. Biochim. Biophys. Acta 2000, 1492, 295–310. [Google Scholar] [CrossRef]
- Rodogiannis, K.; Duong, J.T.; Kovarik, M.L. Microfluidic single-cell analysis of oxidative stress in Dictyostelium discoideum. Analyst 2018, 143, 3643. [Google Scholar] [CrossRef] [PubMed]
- Wilczynska, Z.; Barth, C.; Fisher, P.R. Mitochondrial mutations impair signal transduction in Dictyostelium discoideum slugs. Biochem. Biophys. Res. Commun. 1997, 234, 39–43. [Google Scholar] [CrossRef] [PubMed]
- Nellen, W.; Silan, C.; Firtel, R.A. DNA-mediated transformation in Dictyostelium discoideum: Regulated expression of an actin gene fusion. Mol. Cell. Biol. 1984, 4, 2890–2898. [Google Scholar] [CrossRef] [Green Version]
- Wilczynska, Z.; Fisher, P.R. Analysis of a complex plasmid insertion in a phototaxis-deficient transformant of Dictyostelium discoideum selected on a Micrococcus luteus lawn. Plasmid 1994, 32, 182–194. [Google Scholar] [CrossRef]
- Fisher, P.R.; Annesley, S.J. Slug phototaxis, thermotaxis, and spontaneous turning behavior. Methods Mol. Biol. 2006, 346, 137–170. [Google Scholar]
- Kotsifas, M.; Barth, C.; de Lozanne, A.; Lay, S.T.; Fisher, P.R. Chaperonin 60 and mitochondrial disease in Dictyostelium. J. Muscle Res. Cell Motil. 2002, 23, 839–852. [Google Scholar] [CrossRef]
- Maselli, A.; Laevsky, G.; Knecht, D.A. Kinetics of binding, uptake and degradation of live fluorescent (DsRed) bacteria by Dictyostelium discoideum. Microbiology 2002, 148, 413–420. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.B.; Wei, Y.H. AMPK-mediated increase of glycolysis as an adaptive response to oxidative stress in human cells: Implication of the cell survival in mitochondrial diseases. Biochim. Biophys. Acta 2012, 1822, 233–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghosh, A.S.; Dutta, S.; Raha, S. Hydrogen peroxide-induced apoptosis-like cell death in Entamoeba histolytica. Parasitol. Int. 2010, 59, 166–172. [Google Scholar] [CrossRef] [PubMed]
- Barth, C.; Fraser, D.J.; Fisher, P.R. Coinsertional replication is responsible for tandem multimer formation during plasmid integration into the Dictyostelium genome. Plasmid 1998, 39, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Trempe, J.-F.; Fon, E.A. Structure and function of Parkin, PINK1, and DJ-1, the three musketeers of neuroprotection. Front. Neurol. 2013, 4, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, E.; Fisher, P.R.; Grant, W.N.; Williams, K.L. Sensory behaviour in Dictyostelium discoideum slugs: Phototaxis and thermotaxis are not mediated by a change in slug speed. J. Cell Sci. 1982, 54, 329–339. [Google Scholar] [CrossRef]
- Chen, Z.; Shen, X.; Shen, F.; Zhong, W.; Wu, H.; Liu, S.; Lai, J. TAK1 activates AMPK-dependent cell death pathway in hydrogen peroxide-treated cardiomyocytes, inhibited by heat shock protein-70. Mol. Cell. Biochem. 2013, 377, 35–44. [Google Scholar] [CrossRef]
- Cosson, P.; Soldati, T. Eat, kill or die: When amoeba meets bacteria. Curr. Opin. Microbiol. 2008, 11, 271–276. [Google Scholar] [CrossRef]
- Maurya, R.; Kumar, R.; Saran, S. Dictyostelium AMPKα regulates aggregate size and cell-type patterning. Open Biol. 2017, 7, 170055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciccetti, F.; Brownell, A.; Williams, K.; Chen, Y.; Livni, E.; Isacson, O. Neuroinflammation of the nigrostriatal pathway during progressive 6-OHDA dopamine degeneration in rats monitored by immunohistochemistry and PET imaging. Eur. J. Neurosci. 2002, 15, 991–998. [Google Scholar] [CrossRef]
- Sherer, T.B.; Kim, J.H.; Betarbet, R.; Greenamyre, J.T. Subcutaneous rotenone exposure causes highly selective dopaminergic degeneration and alpha-synuclein aggregation. Exp. Neurol. 2003, 179, 9–16. [Google Scholar] [CrossRef]
- McCormack, A.L.; Atienza, J.G.; Johnston, L.C.; Andersen, J.K.; Vu, S.; Di Monte, D.A. Role of oxidative stress in paraquat-induced dopaminergic cell degeneration. J. Neurochem. 2005, 93, 1030–1037. [Google Scholar] [CrossRef]
- Cappelletti, G.; Surrey, T.; Maci, R. The parkinsonism producing neurotoxin MPP+ affects microtubule dynamics by acting as a destabilising factor. FEBS Lett. 2005, 579, 4781–4786. [Google Scholar] [CrossRef] [Green Version]
- Francione, L.M.; Smith, P.K.; Accari, S.L.; Taylor, P.E.; Bokko, P.B.; Bozzarro, S.; Beech, P.L.; Fisher, P.R. Legionella pneumophila multiplication is enhanced by chronic AMPK signalling in mitochondrially diseased Dictyostelium cells. Dis. Model. Mech. 2009, 2, 479–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.J.; Kim, S.J.; Kim, I.K.; Ko, J.; Jeong, C.S.; Kim, G.H.; Park, C.; Kang, S.O.; Suh, P.G.; Lee, H.S.; et al. Crystal structures of human DJ-1 and Escherichia coli Hsp31, which share an evolutionarily conserved domain. J. Biol. Chem. 2003, 278, 44552–44559. [Google Scholar] [CrossRef] [Green Version]
- Honbou, K.; Suzuki, N.N.; Horiuchi, M.; Niki, T.; Taira, T.; Ariga, H.; Inagaki, F. The crystal structure of DJ-1, a protein related to male fertility and Parkinson’s disease. J. Biol. Chem. 2003, 278, 31380–31384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, X.; Tong, L. Crystal structure of human DJ-1, a protein associated with early onset Parkinson’s disease. J. Biol. Chem. 2003, 278, 31372–31379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, X.; Choi, I.G.; Kim, R.; Wang, W.; Jancarik, J.; Yokota, H.; Kim, S.H. Crystal structure of an intracellular protease from Pyrococcus horikoshii at 2-A resolution. Proc. Natl. Acad. Sci. USA 2000, 97, 14079–14084. [Google Scholar] [CrossRef] [Green Version]
- Mitsumoto, A.; Nakagawa, Y. DJ-1 is an indicator for endogenous reactive oxygen species elicited by endotoxin. Free. Radic. Res. 2001, 35, 885–893. [Google Scholar] [CrossRef]
- Fernando, S.; Allan, C.Y.; Mroczek, K.; Pearce, X.; Sanislav, O.; Fisher, P.R.; Annesley, S.J. Cytotoxicity and Mitochondrial Dysregulation Caused by α-Synuclein in Dictyostelium discoideum. Cells 2020, 9, 2289. [Google Scholar] [CrossRef]
- Ugalde, C.L.; Annesley, S.J.; Gordon, S.E.; Mroczek, K.; Perugini, M.A.; Lawson, V.A.; Fisher, P.R.; Finkelstein, D.I.; Hill, A.F. Misfolded α-synuclein causes hyperactive respiration without functional deficit in live neuroblastoma cells. Dis. Models Mech. 2020, 13, 13. [Google Scholar] [CrossRef] [Green Version]
- Hwang, S.; Song, S.; Hong, Y.K.; Choi, G.; Suh, Y.S.; Han, S.Y.; Lee, M.; Park, S.H.; Lee, J.H.; Lee, S.; et al. Drosophila DJ-1 decreases neural sensitivity to stress by negatively regulating Daxx-like protein through dFOXO. PLoS Genet. 2013, 9, e1003412. [Google Scholar] [CrossRef] [Green Version]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, S.; Annesley, S.J.; Jasim, R.A.F.; Fisher, P.R. The Parkinson’s Disease-Associated Protein DJ-1 Protects Dictyostelium Cells from AMPK-Dependent Outcomes of Oxidative Stress. Cells 2021, 10, 1874. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10081874
Chen S, Annesley SJ, Jasim RAF, Fisher PR. The Parkinson’s Disease-Associated Protein DJ-1 Protects Dictyostelium Cells from AMPK-Dependent Outcomes of Oxidative Stress. Cells. 2021; 10(8):1874. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10081874
Chicago/Turabian StyleChen, Suwei, Sarah J. Annesley, Rasha A. F. Jasim, and Paul R. Fisher. 2021. "The Parkinson’s Disease-Associated Protein DJ-1 Protects Dictyostelium Cells from AMPK-Dependent Outcomes of Oxidative Stress" Cells 10, no. 8: 1874. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10081874