
Role of yUbp8 in Mitochondria and Hypoxia Entangles the Finding of Human Ortholog Usp22 in the Glioblastoma Pseudo-Palisade Microlayer

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast Strains, Growth
2.2. O2 Consumption Measurement
2.3. Total Protein Extraction, SDS-PAGE, Western Blot and Immunohistochemistry
2.4. Rosella Experiment
2.5. Fluorescent Imaging
2.6. Real Time qRT-PCR and End-Point PCR
- ACT1 Fw: 5′-GCTGAAAGAGAAATTGTCCG-3′;
- ACT1 Rev: 5′-ACACTTCATGATGGAGTTGTA-3′;
- OXI1 Fw: 5′-GTACCAACACCTTATGCAT-3′;
- OXI1 Rev: 5′-CATTCAAGATACTAAACCTAA-3′;
- POR1 Fw: 5′-CAAGGATTTCTATCATGCTACC-3′;
- POR1 Rev: 5′GCTTGTCATTCAACTTTGCTTC-3′.
2.7. Statistical Analysis
2.8. Glioma Samples
2.9. Immunohistochemistry
3. Results
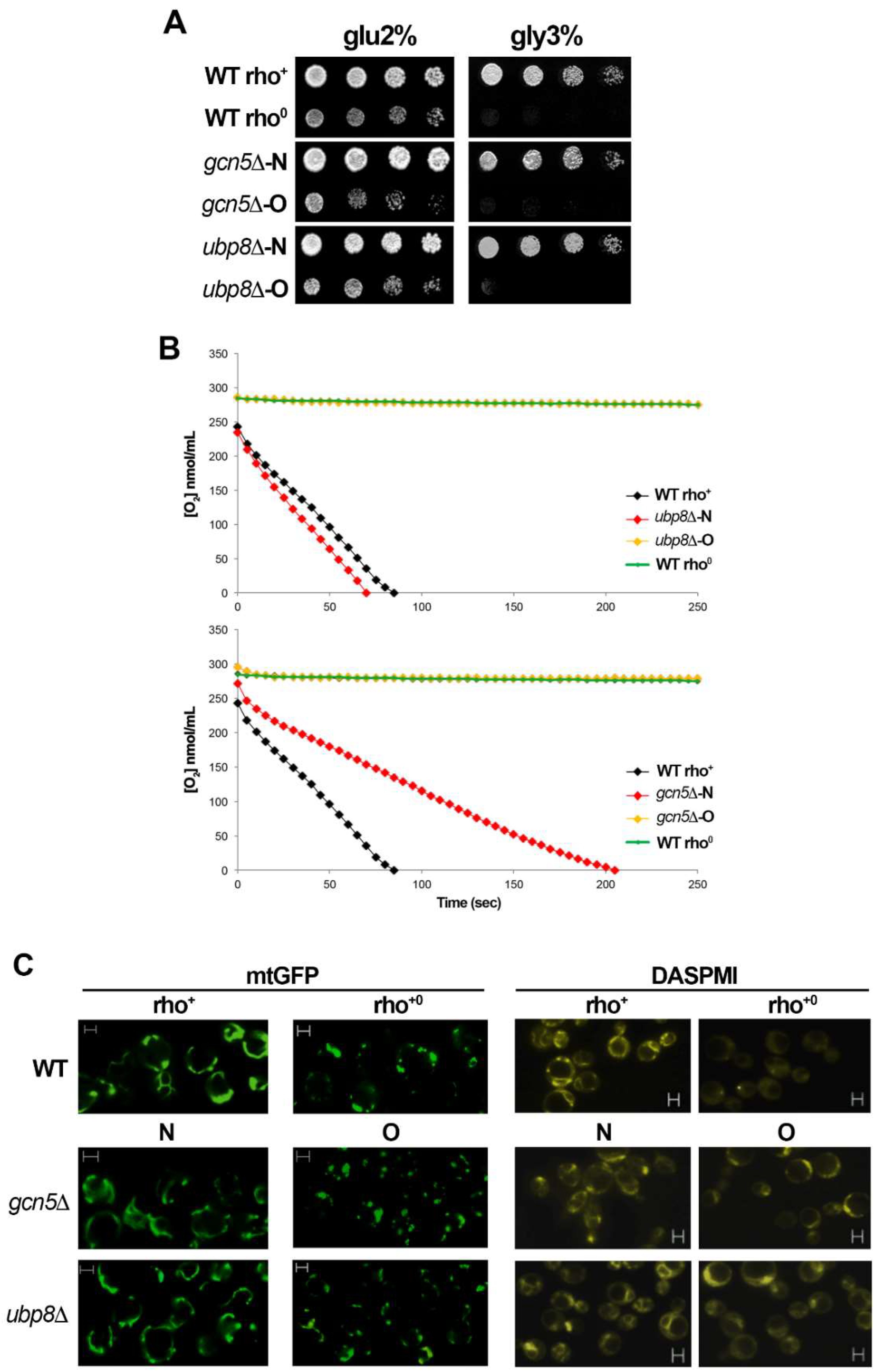
3.1. Lack of Gcn5 and Ubp8 Induces a Progressive Loss of Respiration in Time
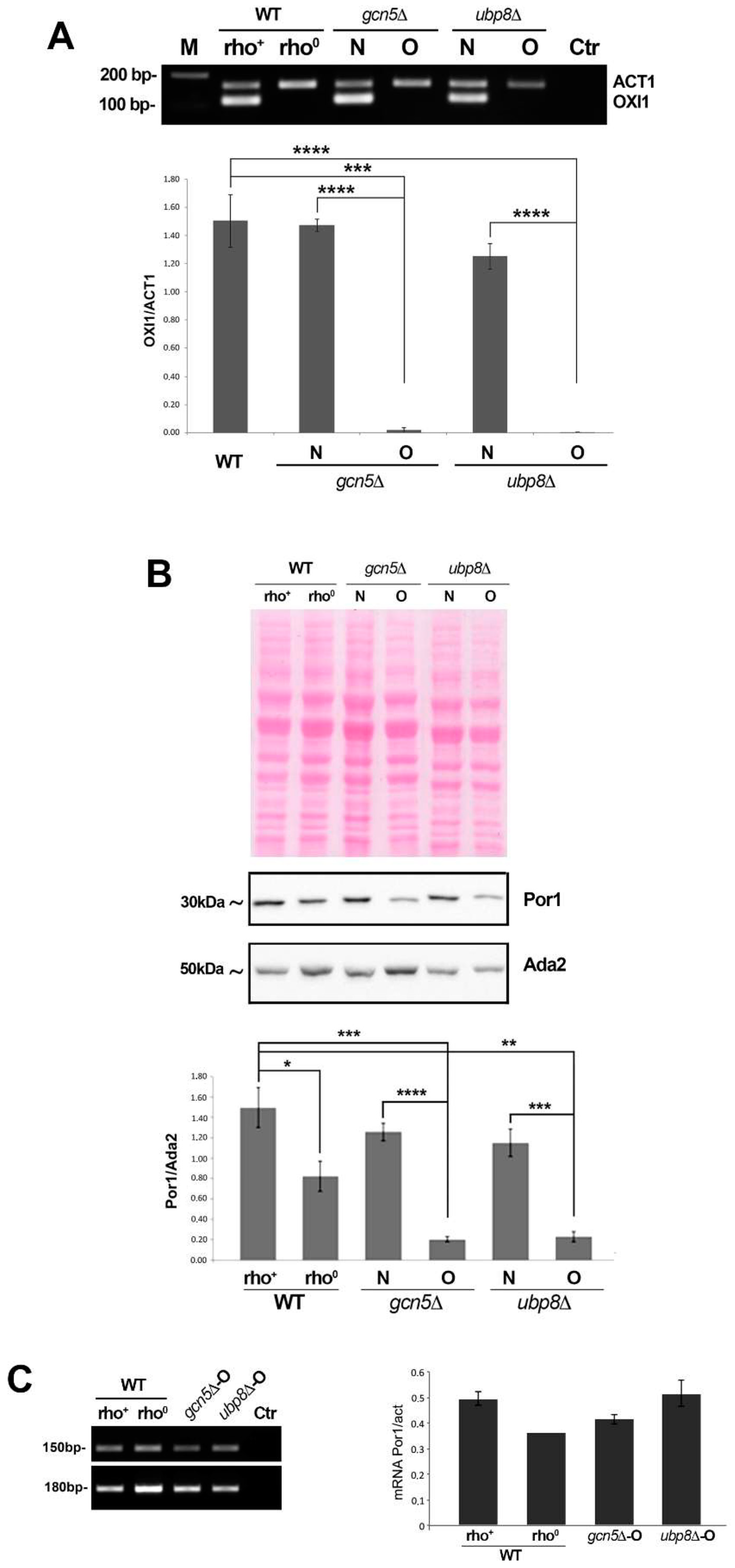
3.2. Mitochondria Components Are Lost in gcn5Δ and ubp8Δ Old Strains
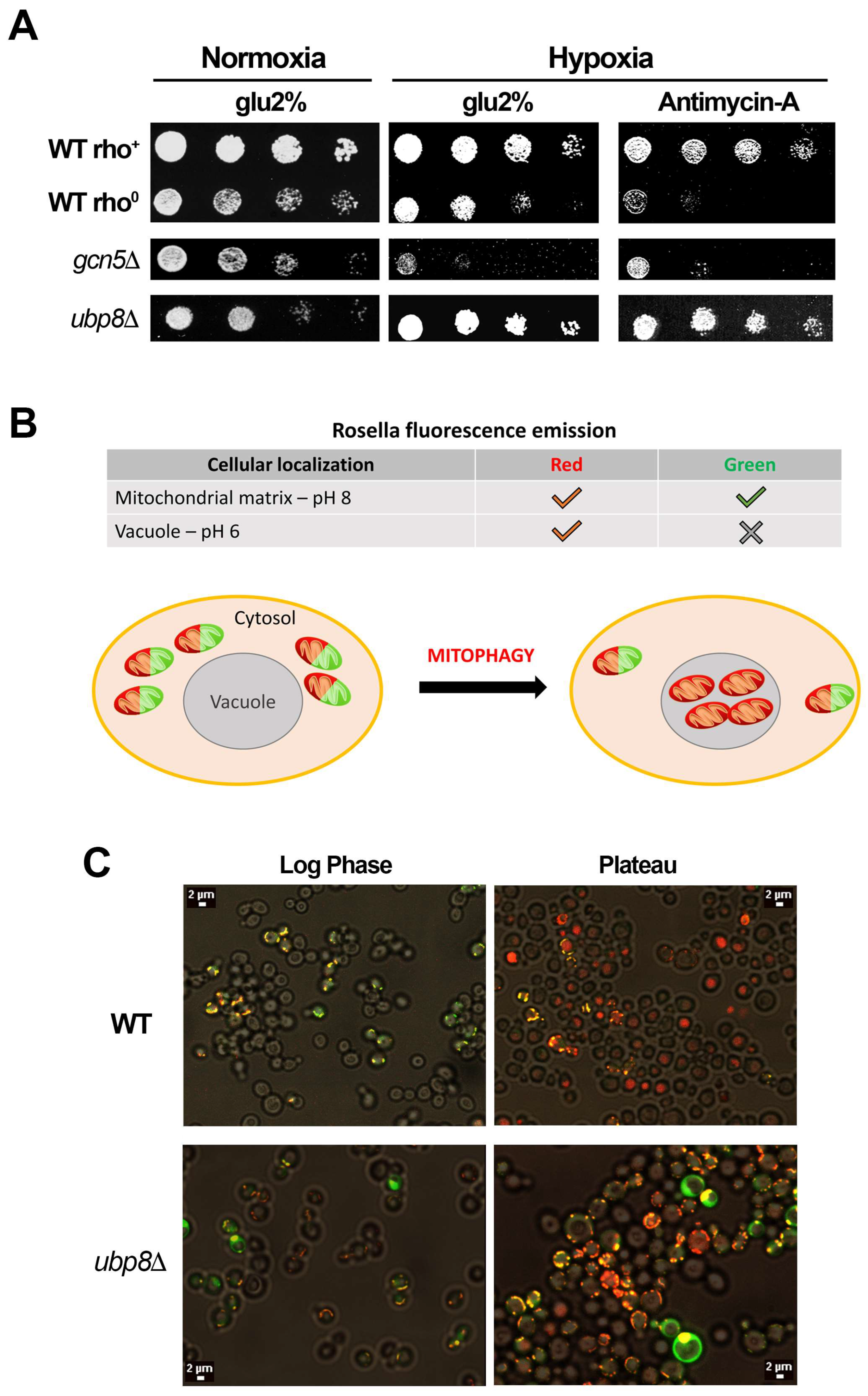
3.3. Yeast Cells Missing Ubp8 Are Able to Grow in Hypoxic Conditions and Show Abnormal Mitophagy
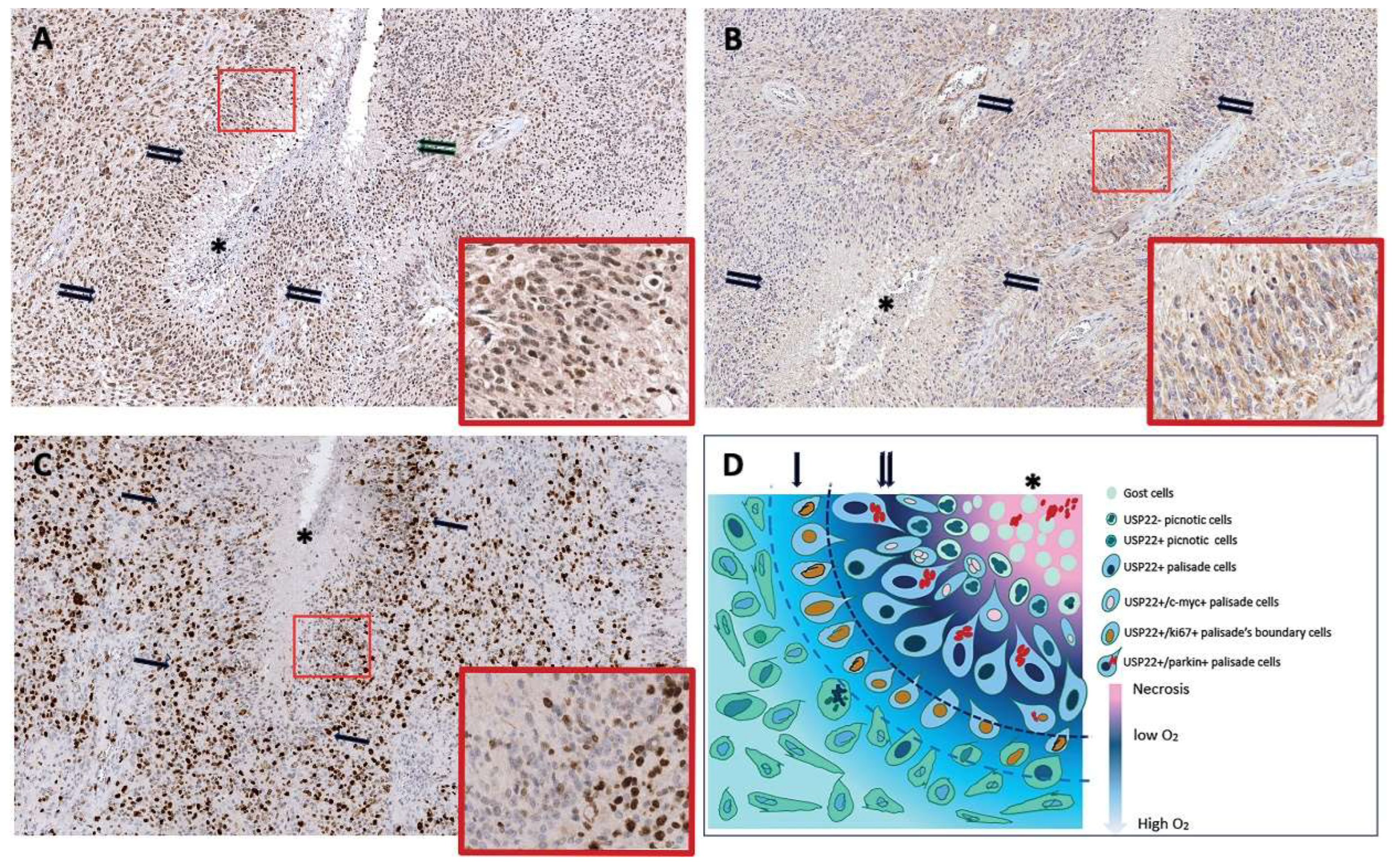
3.4. The Human Ortholog of Ubp8 Usp22 Is Overexpressed in the Pseudo-Palisade Tissue of Glioblastoma Multiforme Tumor Specimen
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schaefer, A.M.; Taylor, R.W.; Turnbull, D.M.; Chinnery, P.F. The epidemiology of mitochondrial disorders—Past, present and future. Biochim. Biophys. Acta 2004, 1659, 115–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foury, F.; Roganti, T.; Lecrenier, N.; Purnelle, B. The complete sequence of the mitochondrial genome of Saccharomyces cerevisiae. FEBS Lett. 1998, 440, 325–331. [Google Scholar] [CrossRef] [Green Version]
- Taylor, R.C.; Dillin, A. Aging as an event of proteostasis collapse. Cold Spring Harb. Perspect. Biol. 2011, 3, a004440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.J.; Clark-Walker, G.D. Alpha and beta subunits of F1-ATPase are required for survival of petite mutants in Saccharomyces cerevisiae. Mol. Gen. Genet. 1999, 262, 898–908. [Google Scholar] [CrossRef] [PubMed]
- Bendich, A.J. Structural analysis of mitochondrial DNA molecules from fungi and plants using moving pictures and pulsed-field gel electrophoresis. J. Mol. Biol. 1996, 255, 564–588. [Google Scholar] [CrossRef]
- Drose, S.; Brandt, U. The mechanism of mitochondrial superoxide production by the cytochrome bc1 complex. J. Biol. Chem. 2008, 283, 21649–21654. [Google Scholar] [CrossRef] [Green Version]
- Eisenberg-Bord, M.; Schuldiner, M. Ground control to major TOM: Mitochondria-nucleus communication. FEBS J. 2017, 284, 196–210. [Google Scholar] [CrossRef] [Green Version]
- Farre, J.C.; Burkenroad, A.; Burnett, S.F.; Subramani, S. Phosphorylation of mitophagy and pexophagy receptors coordinates their interaction with Atg8 and Atg11. EMBO Rep. 2013, 14, 441–449. [Google Scholar] [CrossRef] [Green Version]
- Grant, P.A.; Duggan, L.; Cote, J.; Roberts, S.M.; Brownell, J.E.; Candau, R.; Ohba, R.; Owen-Hughes, T.; Allis, C.D.; Winston, F.; et al. Yeast Gcn5 functions in two multisubunit complexes to acetylate nucleosomal histones: Characterization of an Ada complex and the SAGA (Spt/Ada) complex. Genes Dev. 1997, 11, 1640–1650. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Ohkuni, K.; Couplan, E.; Jazwinski, S.M. The histone acetyltransferase GCN5 modulates the retrograde response and genome stability determining yeast longevity. Biogerontology 2004, 5, 305–316. [Google Scholar] [CrossRef]
- Lemasters, J.J. Selective mitochondrial autophagy, or mitophagy, as a targeted defense against oxidative stress, mitochondrial dysfunction, and aging. Rejuvenation Res. 2005, 8, 3–5. [Google Scholar] [CrossRef] [PubMed]
- Pickles, S.; Vigie, P.; Youle, R.J. Mitophagy and Quality Control Mechanisms in Mitochondrial Maintenance. Curr. Biol. 2018, 28, R170–R185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tatsuta, T.; Langer, T. Quality control of mitochondria: Protection against neurodegeneration and ageing. EMBO J. 2008, 27, 306–314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhatia-Kissova, I.; Camougrand, N. Mitophagy in yeast: Actors and physiological roles. FEMS Yeast Res. 2010, 10, 1023–1034. [Google Scholar] [CrossRef] [Green Version]
- Galluzzi, L.; Baehrecke, E.H.; Ballabio, A.; Boya, P.; Bravo-San Pedro, J.M.; Cecconi, F.; Choi, A.M.; Chu, C.T.; Codogno, P.; Colombo, M.I.; et al. Molecular definitions of autophagy and related processes. EMBO J. 2017, 36, 1811–1836. [Google Scholar] [CrossRef]
- Kanki, T.; Wang, K.; Cao, Y.; Baba, M.; Klionsky, D.J. Atg32 is a mitochondrial protein that confers selectivity during mitophagy. Dev. Cell 2009, 17, 98–109. [Google Scholar] [CrossRef] [Green Version]
- Narendra, D.; Tanaka, A.; Suen, D.F.; Youle, R.J. Parkin is recruited selectively to impaired mitochondria and promotes their autophagy. J. Cell Biol. 2008, 183, 795–803. [Google Scholar] [CrossRef] [Green Version]
- Graef, M.; Nunnari, J. Mitochondria regulate autophagy by conserved signalling pathways. EMBO J. 2011, 30, 2101–2114. [Google Scholar] [CrossRef] [Green Version]
- Priault, M.; Salin, B.; Schaeffer, J.; Vallette, F.M.; di Rago, J.P.; Martinou, J.C. Impairing the bioenergetic status and the biogenesis of mitochondria triggers mitophagy in yeast. Cell Death Differ. 2005, 12, 1613–1621. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Qi, H.; Tang, Y.; Shen, H.M. Post-translational Modifications of Key Machinery in the Control of Mitophagy. Trends Biochem. Sci. 2020, 45, 58–75. [Google Scholar] [CrossRef]
- Kinner, A.; Kolling, R. The yeast deubiquitinating enzyme Ubp16 is anchored to the outer mitochondrial membrane. FEBS Lett. 2003, 549, 135–140. [Google Scholar] [CrossRef] [Green Version]
- Muller, M.; Kotter, P.; Behrendt, C.; Walter, E.; Scheckhuber, C.Q.; Entian, K.D.; Reichert, A.S. Synthetic quantitative array technology identifies the Ubp3-Bre5 deubiquitinase complex as a negative regulator of mitophagy. Cell Rep. 2015, 10, 1215–1225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inoue, M.; Isomura, M.; Ikegawa, S.; Fujiwara, T.; Shin, S.; Moriya, H.; Nakamura, Y. Isolation and characterization of a human cDNA clone (GCN5L1) homologous to GCN5, a yeast transcription activator. Cytogenet Cell Genet. 1996, 73, 134–136. [Google Scholar] [CrossRef] [PubMed]
- Scott, I.; Wang, L.; Wu, K.; Thapa, D.; Sack, M.N. GCN5L1/BLOS1 Links Acetylation, Organelle Remodeling, and Metabolism. Trends Cell Biol. 2018, 28, 346–355. [Google Scholar] [CrossRef] [PubMed]
- Scott, I.; Webster, B.R.; Chan, C.K.; Okonkwo, J.U.; Han, K.; Sack, M.N. GCN5-like protein 1 (GCN5L1) controls mitochondrial content through coordinated regulation of mitochondrial biogenesis and mitophagy. J. Biol. Chem. 2014, 289, 2864–2872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scott, I.; Webster, B.R.; Li, J.H.; Sack, M.N. Identification of a molecular component of the mitochondrial acetyltransferase programme: A novel role for GCN5L1. Biochem. J. 2012, 443, 655–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thapa, D.; Wu, K.; Stoner, M.W.; Xie, B.; Zhang, M.; Manning, J.R.; Lu, Z.; Li, J.H.; Chen, Y.; Gucek, M.; et al. The protein acetylase GCN5L1 modulates hepatic fatty acid oxidation activity via acetylation of the mitochondrial beta-oxidation enzyme HADHA. J. Biol. Chem. 2018, 293, 17676–17684. [Google Scholar] [CrossRef] [Green Version]
- Canzonetta, C.; Leo, M.; Guarino, S.R.; Montanari, A.; Francisci, S.; Filetici, P. SAGA complex and Gcn5 are necessary for respiration in budding yeast. Biochim. Biophys. Acta 2016, 1863, 3160–3168. [Google Scholar] [CrossRef]
- Leo, M.; Fanelli, G.; Di Vito, S.; Traversetti, B.; La Greca, M.; Palladino, R.A.; Montanari, A.; Francisci, S.; Filetici, P. Ubiquitin protease Ubp8 is necessary for S. cerevisiae respiration. Biochim. Biophys. Acta Mol. Cell Res. 2018, 1865, 1491–1500. [Google Scholar] [CrossRef]
- Montanari, A.; Leo, M.; De Luca, V.; Filetici, P.; Francisci, S. Gcn5 histone acetyltransferase is present in the mitoplasts. Biol. Open 2019, 8, bio041244. [Google Scholar] [CrossRef] [Green Version]
- Hambardzumyan, D.; Bergers, G. Glioblastoma: Defining Tumor Niches. Trends Cancer 2015, 1, 252–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boso, D.; Rampazzo, E.; Zanon, C.; Bresolin, S.; Maule, F.; Porcu, E.; Cani, A.; Della Puppa, A.; Trentin, L.; Basso, G.; et al. HIF-1alpha/Wnt signaling-dependent control of gene transcription regulates neuronal differentiation of glioblastoma stem cells. Theranostics 2019, 9, 4860–4877. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Bao, S.; Wu, Q.; Wang, H.; Eyler, C.; Sathornsumetee, S.; Shi, Q.; Cao, Y.; Lathia, J.; McLendon, R.E.; et al. Hypoxia-inducible factors regulate tumorigenic capacity of glioma stem cells. Cancer Cell 2009, 15, 501–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; He, Z.; Huang, M.; Liu, T.; Wang, Y.; Xu, H.; Duan, H.; Ma, P.; Zhang, L.; Zamvil, S.S.; et al. Vascular niche IL-6 induces alternative macrophage activation in glioblastoma through HIF-2alpha. Nat. Commun. 2018, 9, 559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brat, D.J.; Mapstone, T.B. Malignant glioma physiology: Cellular response to hypoxia and its role in tumor progression. Ann. Intern. Med. 2003, 138, 659–668. [Google Scholar] [CrossRef]
- Qiu, G.Z.; Mao, X.Y.; Ma, Y.; Gao, X.C.; Wang, Z.; Jin, M.Z.; Sun, W.; Zou, Y.X.; Lin, J.; Fu, H.L.; et al. Ubiquitin-specific protease 22 acts as an oncoprotein to maintain glioma malignancy through deubiquitinating B cell-specific Moloney murine leukemia virus integration site 1 for stabilization. Cancer Sci. 2018, 109, 2199–2210. [Google Scholar] [CrossRef] [Green Version]
- Zhou, A.; Lin, K.; Zhang, S.; Chen, Y.; Zhang, N.; Xue, J.; Wang, Z.; Aldape, K.D.; Xie, K.; Woodgett, J.R.; et al. Nuclear GSK3beta promotes tumorigenesis by phosphorylating KDM1A and inducing its deubiquitylation by USP22. Nat. Cell Biol. 2016, 18, 954–966. [Google Scholar] [CrossRef] [Green Version]
- De Luca, C.; Zhou, Y.; Montanari, A.; Morea, V.; Oliva, R.; Besagni, C.; Bolotin-Fukuhara, M.; Frontali, L.; Francisci, S. Can yeast be used to study mitochondrial diseases? Biolistic tRNA mutants for the analysis of mechanisms and suppressors. Mitochondrion 2009, 9, 408–417. [Google Scholar] [CrossRef]
- Rosado, C.J.; Mijaljica, D.; Hatzinisiriou, I.; Prescott, M.; Devenish, R.J. Rosella: A fluorescent pH-biosensor for reporting vacuolar turnover of cytosol and organelles in yeast. Autophagy 2008, 4, 205–213. [Google Scholar] [CrossRef] [Green Version]
- Louis, D.N.O.H.; Wiestler, O.D.; Cavenee, W.K. (Eds.) WHO Classification of Tumours of the Central Nervous System, 4th ed.; International Agency for Research on Cancer: Lyon, France, 2016. [Google Scholar]
- Chen, Y.C.; Cheng, T.H.; Lin, W.L.; Chen, C.L.; Yang, W.Y.; Blackstone, C.; Chang, C.R. Srv2 Is a Pro-fission Factor that Modulates Yeast Mitochondrial Morphology and Respiration by Regulating Actin Assembly. iScience 2019, 11, 305–317. [Google Scholar] [CrossRef] [Green Version]
- Westermann, B.; Neupert, W. Mitochondria-targeted green fluorescent proteins: Convenient tools for the study of organelle biogenesis in Saccharomyces cerevisiae. Yeast 2000, 16, 1421–1427. [Google Scholar] [CrossRef]
- Weiss-Brummer, B.; Guba, R.; Haid, A.; Schweyen, R.J. Fine structure of OXI1, the mitochondrial gene coding for subunit II of yeast cytochrome c oxidase. Curr. Genet. 1979, 1, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Torggler, R.; Papinski, D.; Kraft, C. Assays to Monitor Autophagy in Saccharomyces cerevisiae. Cells 2017, 6, 23. [Google Scholar] [CrossRef] [Green Version]
- Miyata, N.; Fujii, S.; Kuge, O. Porin proteins have critical functions in mitochondrial phospholipid metabolism in yeast. J. Biol. Chem. 2018, 293, 17593–17605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyata, N.; Watanabe, Y.; Tamura, Y.; Endo, T.; Kuge, O. Phosphatidylserine transport by Ups2-Mdm35 in respiration-active mitochondria. J. Cell Biol. 2016, 214, 77–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Palma, A.; Fanelli, G.; Cretella, E.; De Luca, V.; Palladino, R.A.; Panzeri, V.; Roffia, V.; Saliola, M.; Mauri, P.; Filetici, P. Gcn5p and Ubp8p Affect Protein Ubiquitylation and Cell Proliferation by Altering the Fermentative/Respiratory Flux Balance in Saccharomyces cerevisiae. mBio 2020, 11, e01504-20. [Google Scholar] [CrossRef] [PubMed]
- Ge, P.; Dawson, V.L.; Dawson, T.M. PINK1 and Parkin mitochondrial quality control: A source of regional vulnerability in Parkinson’s disease. Mol. Neuro. 2020, 15, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, S.A.; Alcalay, R.N. Neuropathology of genetic synucleinopathies with parkinsonism: Review of the literature. Mov. Disord. 2017, 32, 1504–1523. [Google Scholar] [CrossRef]
- Koutelou, E.; Hirsch, C.L.; Dent, S.Y. Multiple faces of the SAGA complex. Curr. Opin. Cell Biol. 2010, 22, 374–382. [Google Scholar] [CrossRef] [Green Version]
- Harris, A.L. Hypoxia—A key regulatory factor in tumour growth. Nat. Rev. Cancer 2002, 2, 38–47. [Google Scholar] [CrossRef]
- Stoppacciaro, A.; Di Vito, S.; Filetici, P. Epigenetic Factors and Mitochondrial Biology in Yeast: A New Paradigm for the Study of Cancer Metabolism? Front. Pharm. 2018, 9, 1349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glinsky, G.V. Death-from-cancer signatures and stem cell contribution to metastatic cancer. Cell Cycle 2005, 4, 1171–1175. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Genotype | Source |
|---|---|---|
| W303 | MATa ade2–1 trp1–1 leu2–3112 his3–11,15 ura3 can1–100 | [29] |
| YVDL09 | MATa ade2–1 trp1–1 leu2–3112 his3–11,15 ura3 can1–100 + pVT100U-mtGFP | This study |
| W303-rho0 | MATa ade2–1 trp1–1 leu2–3112 his3–11,15 ura3 can1–100. rho0 | This study |
| YVDL11 | MATa ade2–1 trp1–1 leu2–3112 his3–11,15 ura3 can1–100. rho0 + pVT100U-mtGFP | This study |
| YPO4 | MATa ade2–1 trp1–1 leu2–3112 his3–11,15 ura3 can1–100 gcn5::KanMX4 | [29] |
| YVDL12 | MATa ade2–1 trp1–1 leu2–3112 his3–11,15 ura3 can1–100 gcn5::KanMX4 + pVT100U-mtGFP | This study |
| YV01 | MATa ade2–1 trp1–1 leu2–3112 his3–11,15 ura3 can1–100 gcn5::KanMX4 | This study |
| YVDL13 | MATa ade2–1 trp1–1 leu2–3112 his3–11,15 ura3 can1–100 gcn5::KanMX4 + pVT100U-mtGFP | This study |
| YFT21 | MATa ade2–1 trp1–1 leu2–3112 his3–11,15 ura3 can1–100 ubp8::His3MX6 | [29] |
| YVDL14 | MATa ade2–1 trp1–1 leu2–3112 his3–11,15 ura3 can1–100 ubp8::His3MX6 + pVT100U-mtGFP | This study |
| YV02 | MATa ade2–1 trp1–1 leu2–3112 his3–11,15 ura3 can1–100 ubp8::His3MX6 | This study |
| YVDL15 | MATa ade2–1 trp1–1 leu2–3112 his3–11,15 ura3 can1–100 ubp8::His3MX6 + pVT100U-mtGFP | This study |
| Ros01 | MATa ade2–1 trp1–1 leu2–3112 his3–11,15 ura3 can1–100 + pAS1NBmRosellaI | This study |
| Ros02 | MATa ade2–1 trp1–1 leu2–3112 his3–11,15 ura3 can1–100 gcn5::KanMX4 + pAS1NBmRosellaI | This study |
| Ros03 | MATa ade2–1 trp1–1 leu2–3112 his3–11,15 ura3 can1–100 ubp8::His3MX6 + pAS1NBmRosellaI | This study |
| Total No. of Cases | Cases of Glioblastomas Expressing USP22 | Cases with Palisade Necrosis Cells Over-Expressing USP22 | Cases with USP22 Comparable Expression in Vital Tumor Cells and Palisade Necrosis Cells | Cases with Palisade Necrosis Cells Ipo-Expressing USP22 |
|---|---|---|---|---|
| No. of cases | 30/30 | 19/30 | 9/30 | 1/30 |
| USP22 Mean Expression + SD | 2 ± 0.643 | 2.947 ± 0.229 | 2.363 ± 0.809 | |
| Test T | 6.94706 × 10−13 p < 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Luca, V.; Leo, M.; Cretella, E.; Montanari, A.; Saliola, M.; Ciaffi, G.; Vecchione, A.; Stoppacciaro, A.; Filetici, P. Role of yUbp8 in Mitochondria and Hypoxia Entangles the Finding of Human Ortholog Usp22 in the Glioblastoma Pseudo-Palisade Microlayer. Cells 2022, 11, 1682. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11101682
De Luca V, Leo M, Cretella E, Montanari A, Saliola M, Ciaffi G, Vecchione A, Stoppacciaro A, Filetici P. Role of yUbp8 in Mitochondria and Hypoxia Entangles the Finding of Human Ortholog Usp22 in the Glioblastoma Pseudo-Palisade Microlayer. Cells. 2022; 11(10):1682. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11101682
Chicago/Turabian StyleDe Luca, Veronica, Manuela Leo, Elisabetta Cretella, Arianna Montanari, Michele Saliola, Gabriele Ciaffi, Andrea Vecchione, Antonella Stoppacciaro, and Patrizia Filetici. 2022. "Role of yUbp8 in Mitochondria and Hypoxia Entangles the Finding of Human Ortholog Usp22 in the Glioblastoma Pseudo-Palisade Microlayer" Cells 11, no. 10: 1682. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11101682