
An In-Vitro Study of the Expansion and Transcriptomics of CD4+ and CD8+ Naïve and Memory T Cells Stimulated by IL-2, IL-7 and IL-15

 , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture, Cytokine Stimulation, Quantification
2.2. Growth Plate
2.3. Transcriptomics Analysis
3. Results
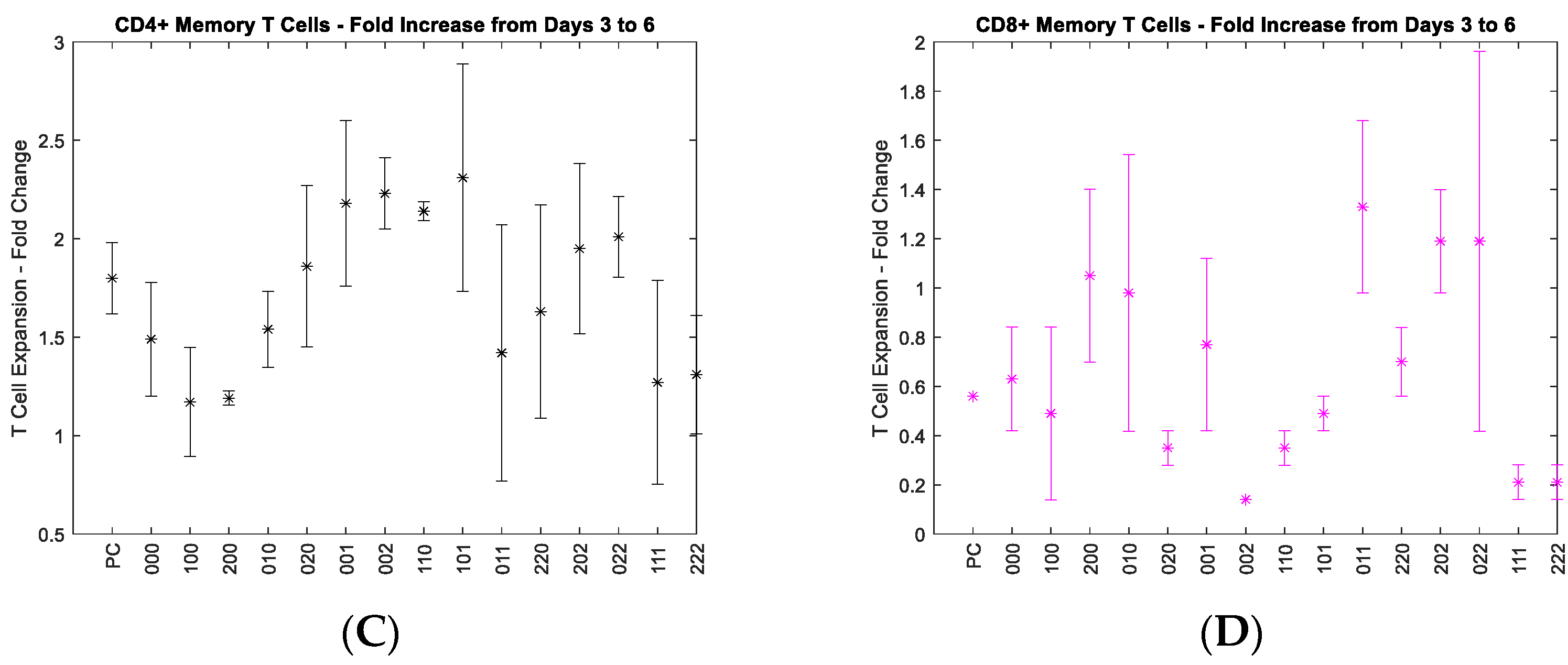
3.1. Identify Optimal Cytokine Combinations from the Statistical Analysis of Cell Growth Rates


3.2. Investigate the Interaction of the Three Cytokines in Promoting the Growth of CD4+ and CD8+ T Cells
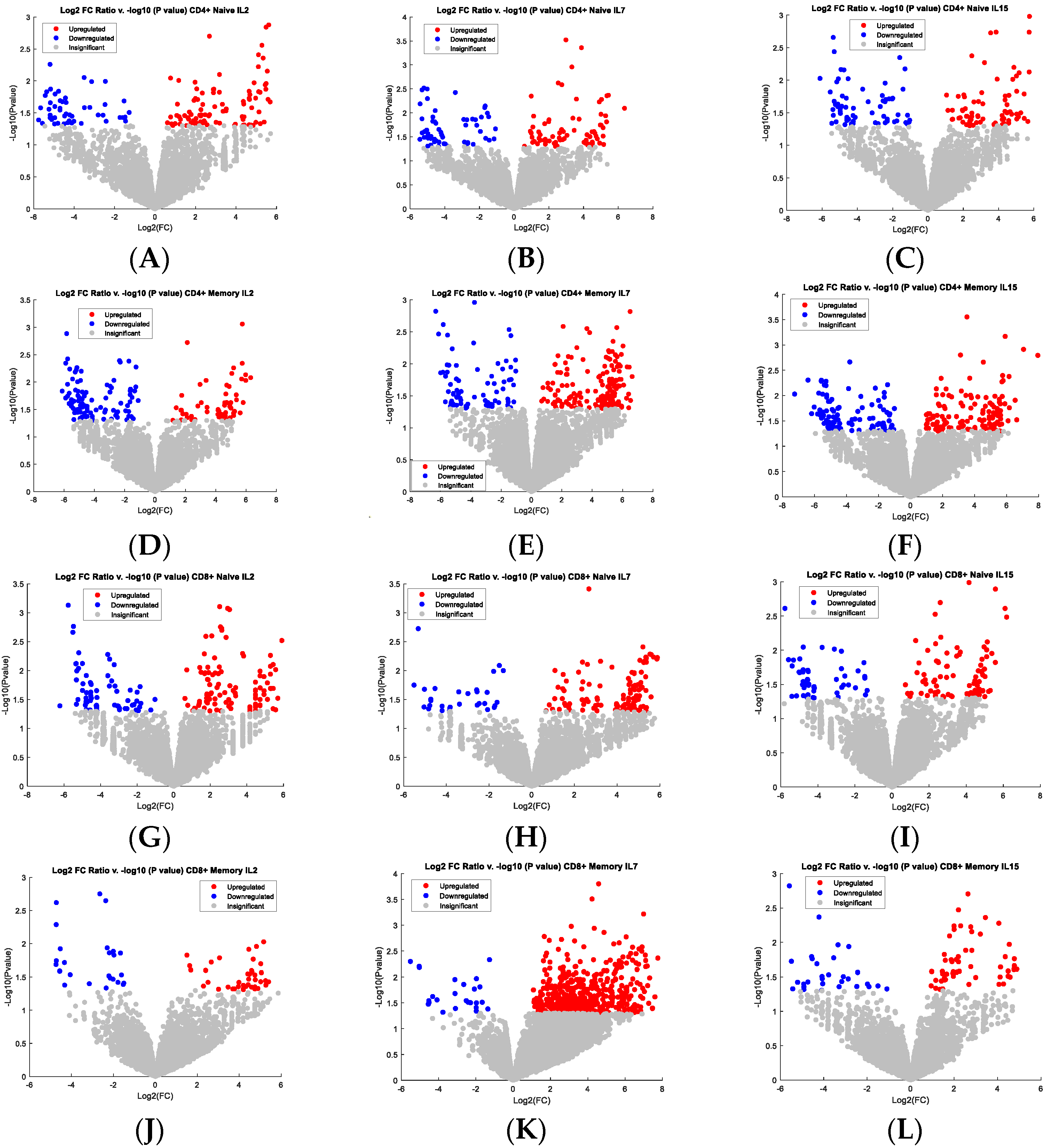
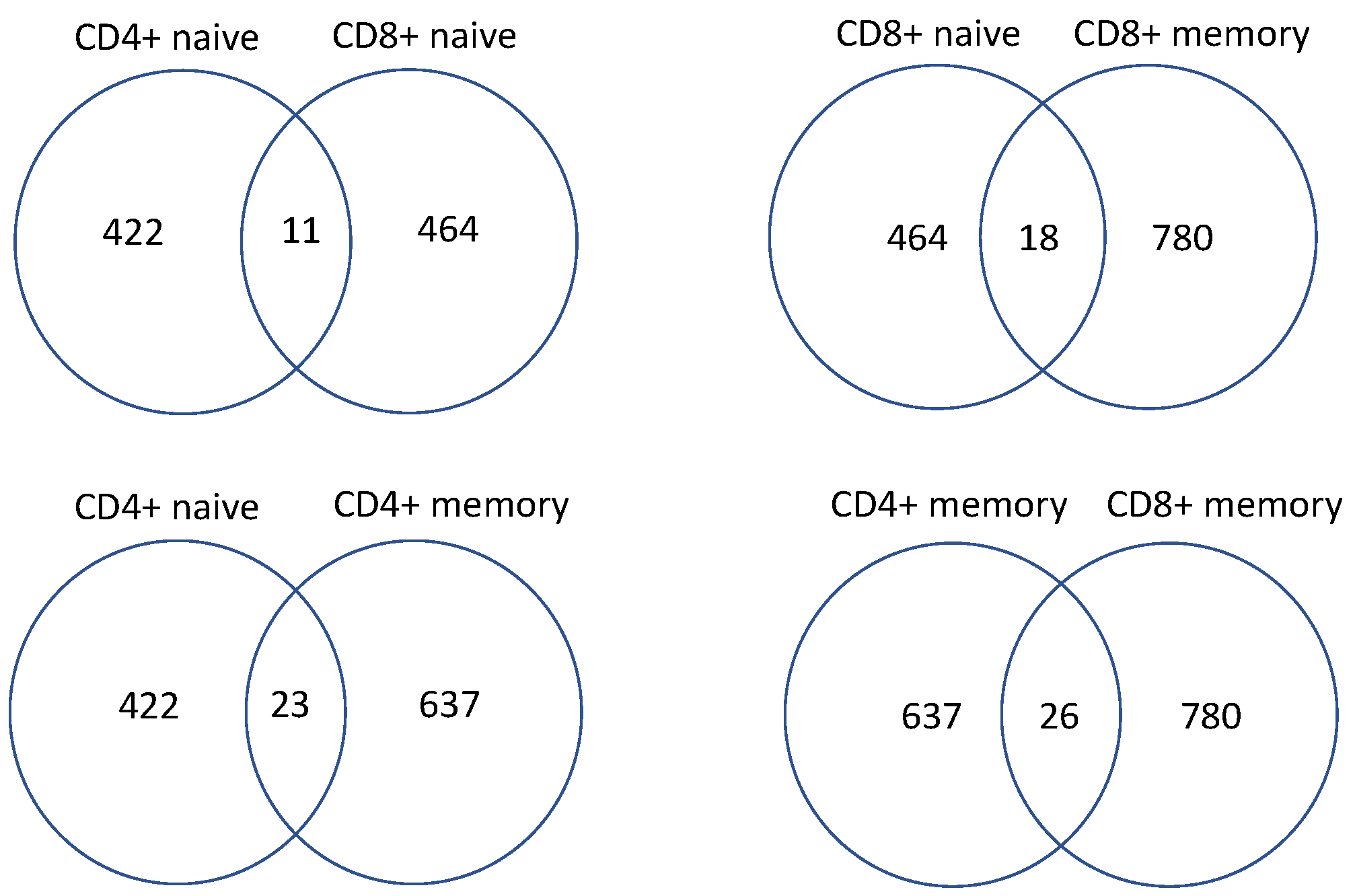
3.3. Study Genes Indicating the Purtabation in the Signal Transduction of Activated CD4+ and CD8+ T Cells Stimulated by Individual Cytokines Using RNA-Sequencing
4. Discussion
4.1. The Impact of Each Cytokine on T Cell Growth
4.1.1. Use of Fold Change over Growth Rate
4.1.2. Impact of Cytokines on the Growth of CD4+ and CD8+ T Cells
4.2. Discussion of Genes Regulated by Cytokines in CD4+ and CD8+ T Cells
4.3. Functionality Assessment
4.4. Limitation and Future Work
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Das, K.R.; Vernau, L.; Grupp, S.A.; Barrett, D.M. Naive T-cell Deficits at Diagnosis and after Chemotherapy Impair Cell Therapy Potential in Pediatric Cancers. Cancer Discov. 2019, 9, 492–499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fraietta, J.A.; Lacey, S.F.; Orlando, E.J.; Pruteanu-Malinici, I.; Gohil, M.; Lundh, S.; Boesteanu, A.C.; Wang, Y.; O’Connor, R.S.; Hwang, W.-T.; et al. Determinants of response and resistance to CD19 chimeric antigen receptor (CAR) T cell therapy of chronic lymphocytic leukemia. Nat. Med. 2019, 24, 563–571. [Google Scholar] [CrossRef]
- Sommermeyer, D.; Hudacek, M.; Kosasih, P.L.; Gogishvili, T.; Maloney, D.G.; Turtle, C.; Riddell, S.R. Chimeric antigen receptor-modified T cells derived from defined CD8+ and CD4+ subsets confer superior antitumor reactivity in vivo. Leukemia 2016, 30, 492–500. [Google Scholar] [CrossRef] [Green Version]
- Tay, R.E.; Richardson, E.K.; Toh, H.C. Revisting the role of CD4+ T cells in cancer immunotherapy—New insights into old paradigms. Cancer Gene Ther. 2021, 28, 5–17. [Google Scholar] [CrossRef]
- Geginat, J.; Sallusto, F.; Lanzavecchia, A. Cytokine-driven Proliferation and Differentiation of Human Naive, Central Memory, and Effector Memory CD4+ T Cells. J. Exp. Med. 2001, 194, 1711–1720. [Google Scholar] [CrossRef]
- Alves, L.N.; Hooibrink, B.; Arosa, F.A.; van Lier, R.A.W. IL-15 induces antigen-independent expansion and differentiation of human naive CD8+ T cells in vitro. Blood 2003, 102, 2541–2546. [Google Scholar] [CrossRef] [Green Version]
- Jaleco, S.; Swainson, L.; Dardalhon, V.; Burjanadze, M.; Kinet, S.; Taylor, N. Homeostasis of Naive and Memory CD4+ T Cells: IL-2 and IL-7 Differentially Regulate the Balance Between Proliferation and Fas-Mediated Apoptosis. J. Immunol. 2003, 171, 61–68. [Google Scholar] [CrossRef] [Green Version]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast Universal Rna-Seq Aligner. Bioinformatics 2012, 29, 15–21. [Google Scholar] [CrossRef]
- Breese, M.R.; Liu, Y. NGSUtils: A Software Suite for Analyzing and Manipulating Next-Generation Sequencing Datasets. Bioinformatics 2013, 29, 494–496. [Google Scholar] [CrossRef] [Green Version]
- Liao, Y.; Smyth, G.K.; Shi, W. FeatureCounts: An Efficient General Purpose Program for Assigning Sequence Reads to Genomic Features. Bioinformatics 2013, 30, 923–930. [Google Scholar] [CrossRef] [Green Version]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warde-Farley, D.; Donaldson, S.L.; Comes, O.; Zuberi, K.; Badrawi, R.; Chao, P.; Franz, M.; Grouios, C.; Kazi, F.; Lopes, C.T.; et al. The GeneMANIA Prediction server: Biological network integration for gene prioritization and predicting gene function. Nucleic Acids Res. 2010, 38, W214–W220. [Google Scholar] [CrossRef] [PubMed]
- Kleiner, G.; Marcuzzi, A.; Zanin, V.; Monasta, L.; Zauli, G. Cytokine Levels in the Serum of Healthy Subjects. Mediat. Inflamm. 2013, 2013, 434010. [Google Scholar] [CrossRef] [PubMed]
- Coppolla, C.; Hopkins, B.M.; Huhn, S.; Du, Z.; Huang, Z.; Kelly, W.J. Investigation of the Impact from IL-2, IL-7, and IL-15 on the Growth and Signaling of Activated CD4+ T Cells. Int. J. Mol. Sci. 2020, 21, 7814. [Google Scholar] [CrossRef]
- Reynolds, J.; Coles, M.; Lythe, G.; Molina-Paris, C. Mathematical Model of Naive T Cell Division and Survival IL-7 Thresholds. Front. Immunol. 2013, 4, 434. [Google Scholar] [CrossRef] [Green Version]
- Kanegane, H.; Tosato, G. Activation of Naive and Memory T Cells by IL-15. Blood 1996, 88, 230–235. [Google Scholar]
- Wold, S.; Esbensen, K.; Geladi, P. Principal Component Analysis. Chemom. Intell. Lab. Syst. 1987, 2, 37–52. [Google Scholar]
- Sassone-Corsi, P. Coupling gene expression to cAMP signalling: Role of CREB and CREM. Biochem. Cell Biol. 1998, 30, 27–38. [Google Scholar] [CrossRef]
- Tan, J.T.; Dudl, E.; LeRoy, E.; Murray, R.; Sprent, J.; Weinberg, K.I.; Surh, C.D. IL-7 is critical for homeostatic proliferation and survival of naı¨ve T cells. PNAS 2001, 98, 15. [Google Scholar] [CrossRef] [Green Version]
- Schluns, K.S.; Kieper, W.C.; Jameson, S.C.; Lefrancois, L. IL-7 mediates the homeostasis of naïve and memory CD8 T cells in vivo. Nat. Immunol. 2000, 1, 5. [Google Scholar] [CrossRef]
- Jankauskas, S.S.; Wong, D.W.L.; Bucala, R.; Djudjaj, S.; Boor, P. Evolving Complexity of MIF Signaling. Cell. Signal. 2019, 57, 76–88. [Google Scholar] [CrossRef] [PubMed]
- Altan-Bonnet, G.; Mukherjee, R. Cytokine-mediated communication: A quantitative appraisal of immune complexity. Nat. Rev. Immunol. 2019, 19, 205–217. [Google Scholar] [CrossRef] [PubMed]
- Briscoe, J.; Therond, P.P. The mechanisms of Hedgehog signalling and its roles in development and disease. Nat. Rev. Mol. Cell. Biol. 2013, 14, 416–429. [Google Scholar] [CrossRef] [PubMed]
- Cooke, P.S.; Mesa, A.M.; Sirohi, V.K.; Levin, E.R. Role of nuclear and membrane estrogen signaling pathways in the male and female reproductive tract. Differentiation 2021, 118, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Artavanis-Tsakonas, S.; Rand, M.D.; Lake, R.J. Notch Signaling: Cell Fate Control and Signal Integration in Development. Science 1999, 284, 770–776. [Google Scholar] [CrossRef] [Green Version]
- Adachi, T.; Alam, R. The mechanism of IL-5 signal transduction. Am. J. Physiol. 1998, 275, C623–C633. [Google Scholar] [CrossRef]
- Moreau, J.; Pradeu, T.; Grignolio, A.; Nardini, C.; Castiglione, F.; Tieri, P.; Capri, M.; Salvioli, S.; Taupin, J.-L.; Garagnani, P.; et al. The emerging role and ECM crosslinking in T cell mobility as a hallmark of immunosenescence in humans. Ageing Res. Rev. 2017, 35, 322–335. [Google Scholar] [CrossRef]
- Arts, R.J.W.; Joosten, L.A.B.; van der Meer, J.W.M.; Netea, M.G. TREM-1: Intracellular signaling pathways and interaction with pattern recognition receptors. J. Leukoc. Biol. 2012, 93, 209–215. [Google Scholar] [CrossRef]
- Amelio, A.L.; Miraglia, L.J.; Conkright, J.J.; Mercer, B.A.; Batalov, S.; Cavett, V.; Orth, A.P.; Busby, J.; Hogenesch, J.B.; Conkright, M.D. A coactivator trap identifies NONO (p54nrb) as a component of the cAMP-signaling pathway. PNAS 2007, 104, 20314–20319. [Google Scholar] [CrossRef] [Green Version]
- Honda, K.; Yanai, H.; Negishi, H.; Asagiri, M.; Sato, M.; Mizutani, T.; Shimada, N.; Ohba, Y.; Takaoka, A.; Yoshida, N.; et al. IRF-7 is the master regulator of type-I interferon-dependent immune responses. Nature 2005, 434, 772–777. [Google Scholar] [CrossRef]
- Gattinoni, L.; Ji, Y.; Restifo, N.P. Wnt/β-catenin signaling in T cell immunity and cancer immunotherapy. Clin. Cancer Res. 2010, 16, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Condition | Key | IL-2 (ng/mL) | IL-7 (ng/mL) | IL-15 (ng/mL) |
|---|---|---|---|---|
| 1 | no cytokines | 0 | 0 | 0 |
| 2 | 000 | 10 | 1.5 | 10 |
| 3 | 100 | 33 | 1.5 | 10 |
| 4 | 200 | 60 | 1.5 | 10 |
| 5 | 010 | 10 | 36 | 10 |
| 6 | 020 | 10 | 80 | 10 |
| 7 | 001 | 10 | 1.5 | 36 |
| 8 | 002 | 10 | 1.5 | 80 |
| 9 | 110 | 33 | 36 | 10 |
| 10 | 101 | 33 | 1.5 | 36 |
| 11 | 011 | 10 | 36 | 36 |
| 12 | 220 | 60 | 80 | 10 |
| 13 | 202 | 60 | 1.5 | 80 |
| 14 | 022 | 10 | 80 | 80 |
| 15 | 111 | 33 | 36 | 36 |
| 16 | 222 | 60 | 80 | 80 |
| Stimulus | CD4+ Naïve | p Value | CD4+ Memory | p Value | CD8+ Naïve | p Value | CD8+ Memory | p Value |
|---|---|---|---|---|---|---|---|---|
| IL-2 | X | 0.000457 | 0.231 | 0.2338 | 0.9533 | |||
| IL-7 | X | 0.003395 | 0.828 | 0.5216 | 0.6237 | |||
| IL-15 | 0.829209 | 0.221 | 0.9788 | 0.4289 | ||||
| IL-2/IL-7 | 0.076981 | 0.944 | 0.1731 | 0.0654 | ||||
| IL-2/IL-15 | 0.347061 | 0.772 | 0.8352 | 0.7763 | ||||
| IL-7/IL-15 | X | 0.008359 | 0.196 | X | 0.0141 | 0.6194 | ||
| IL-2/IL-7/IL-15 | X | 0.012759 | 0.444 | X | 0.0186 | X | 0.0126 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hopkins, B.; Fisher, J.; Chang, M.; Tang, X.; Du, Z.; Kelly, W.J.; Huang, Z. An In-Vitro Study of the Expansion and Transcriptomics of CD4+ and CD8+ Naïve and Memory T Cells Stimulated by IL-2, IL-7 and IL-15. Cells 2022, 11, 1701. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11101701
Hopkins B, Fisher J, Chang M, Tang X, Du Z, Kelly WJ, Huang Z. An In-Vitro Study of the Expansion and Transcriptomics of CD4+ and CD8+ Naïve and Memory T Cells Stimulated by IL-2, IL-7 and IL-15. Cells. 2022; 11(10):1701. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11101701
Chicago/Turabian StyleHopkins, Brooks, Justin Fisher, Meiping Chang, Xiaoyan Tang, Zhimei Du, William J. Kelly, and Zuyi Huang. 2022. "An In-Vitro Study of the Expansion and Transcriptomics of CD4+ and CD8+ Naïve and Memory T Cells Stimulated by IL-2, IL-7 and IL-15" Cells 11, no. 10: 1701. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11101701