
Structural Variability of Lipoarabinomannan Modulates Innate Immune Responses within Infected Alveolar Epithelial Cells

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of M. smegmatis EmbC Gene Knockout Strain
2.2. Extraction and Analysis of LAM/LM Mixture
2.3. Identification of Biological Characteristics
2.4. Uptake and Survival of Intracellular Pathogens
2.5. Transmission Electron Microscopy (TEM)
2.6. Detection of ROS Release in A549 Cells
2.7. Assessing LC3 Recruitments with a Confocal Fluorescence Microscope
2.8. The Expression Changes of Genes Related to Pathogen Recognition and Immune Responses
2.9. Statistical Analysis
3. Results
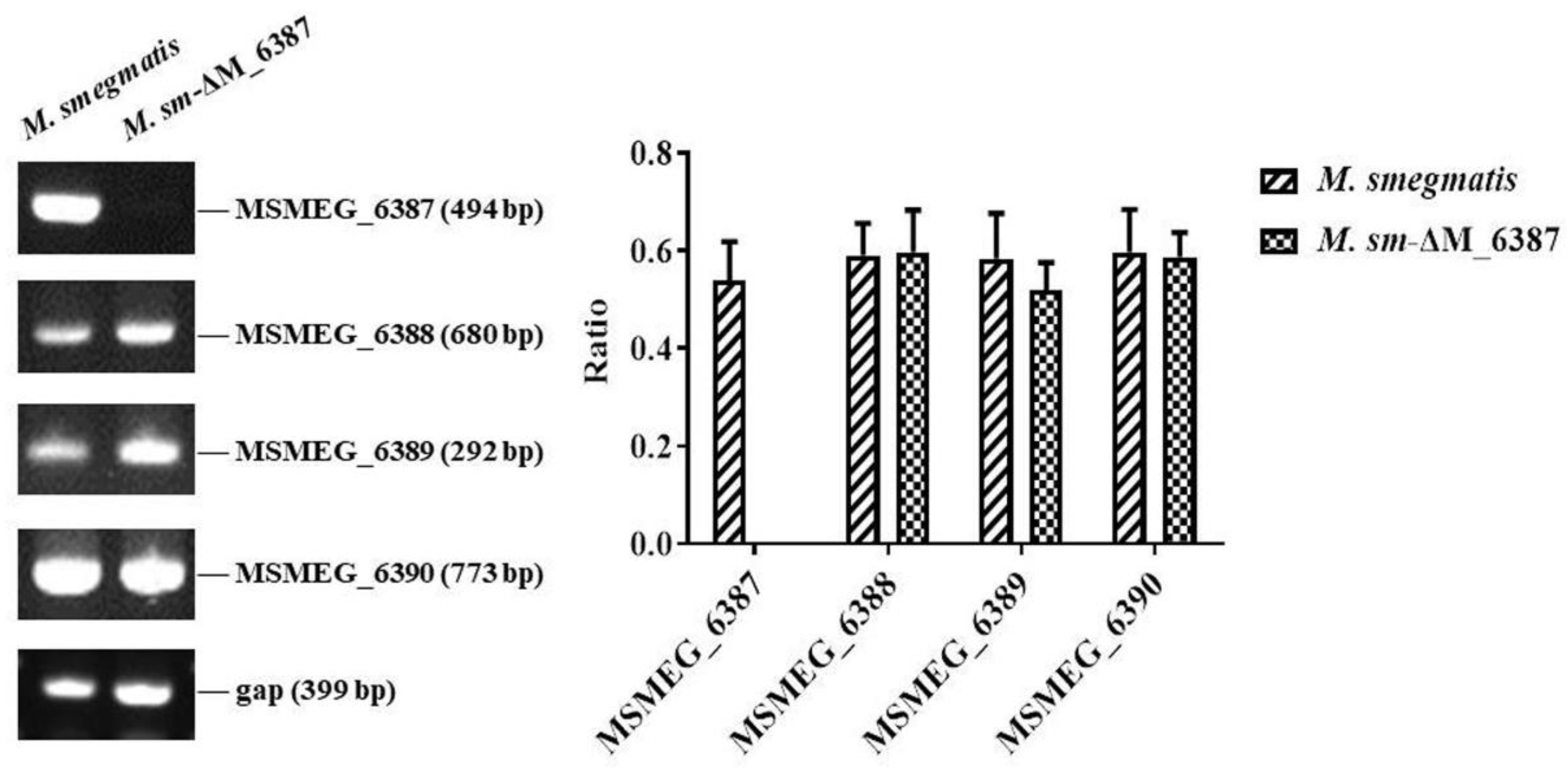
3.1. Construction of an EmbC Gene Knockout Strain
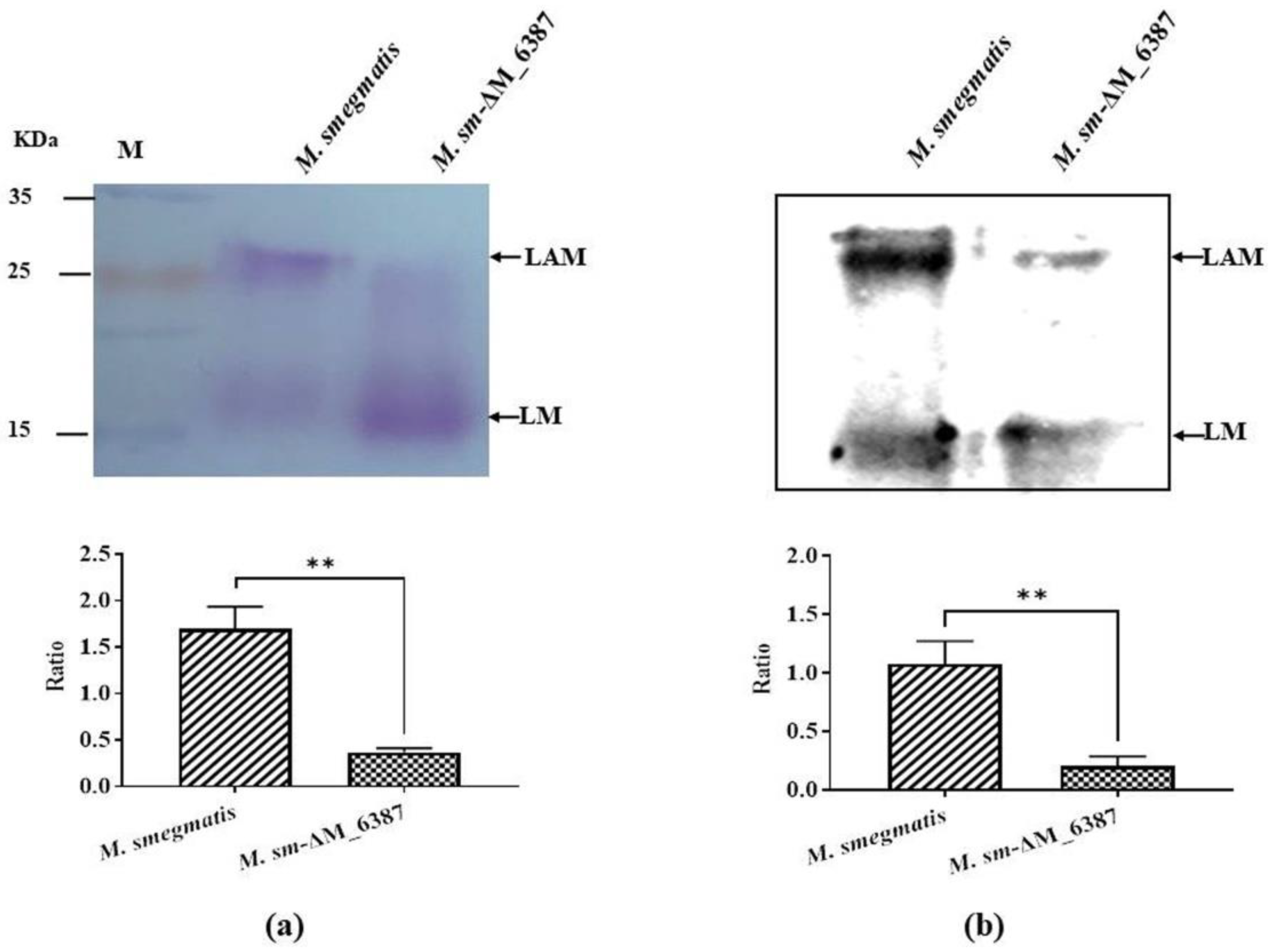
3.2. EmbC Inactivation Induced the Change in LAM:LM in M. smegmatis
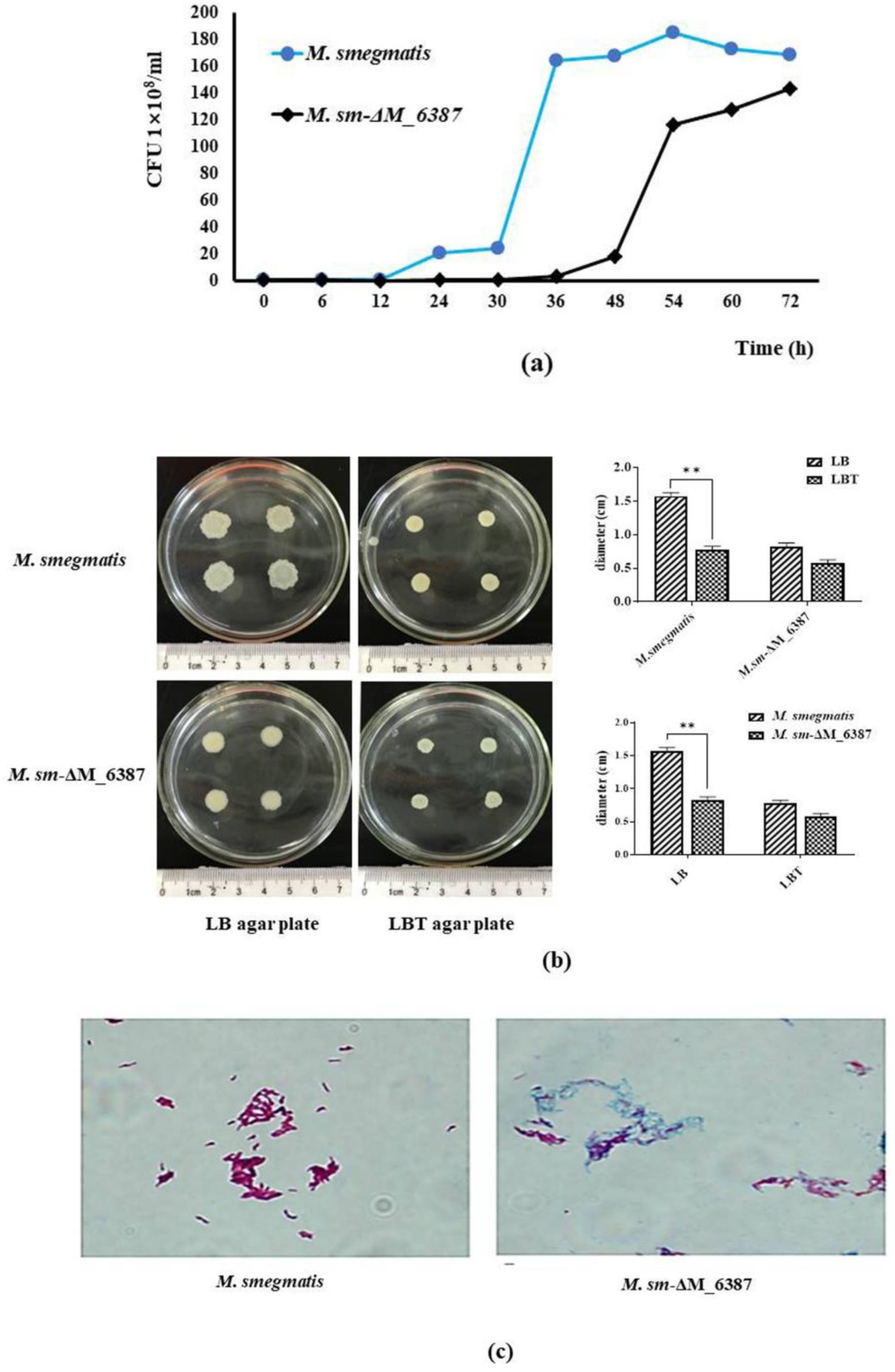
3.3. EmbC in M. smegmatis Is Critical for Mycobacterial Cell Wall Integrity
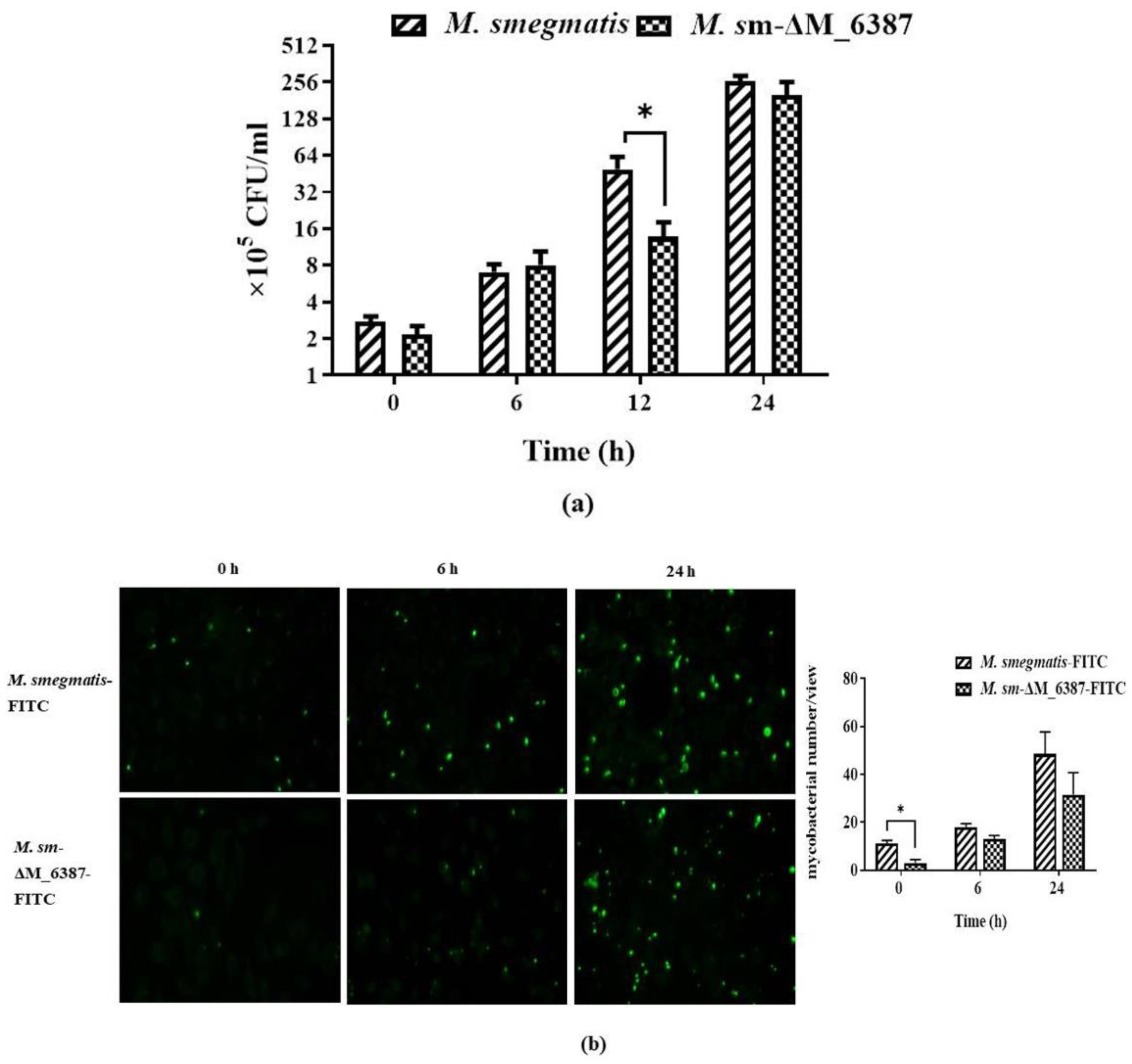
3.4. Intracellular Uptake of M. smegmatis and M. sm-ΔM_6387
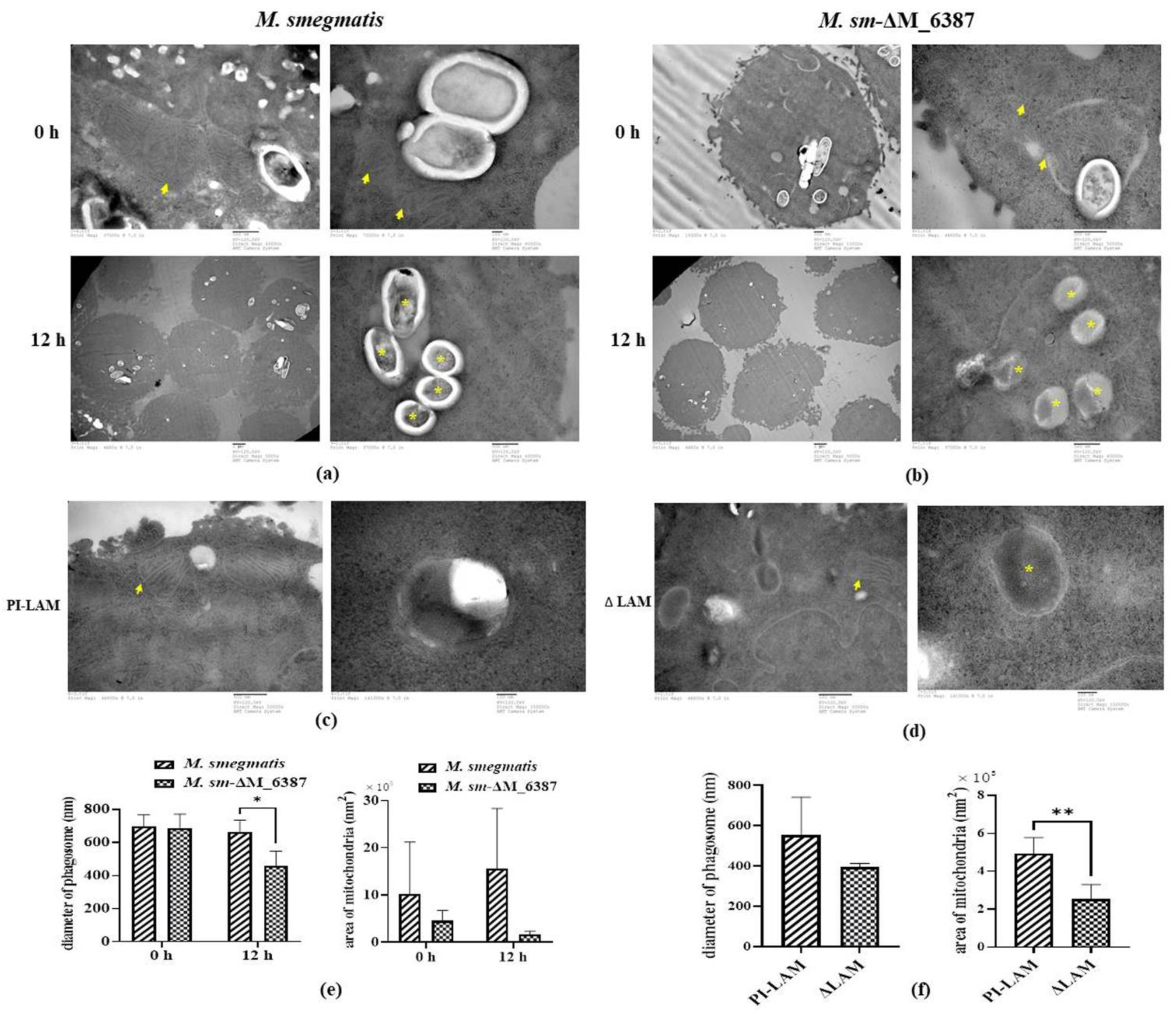
3.5. Changes in the Phagosome and Mitochondrial Size within Infected Alveolar Epithelial Cells
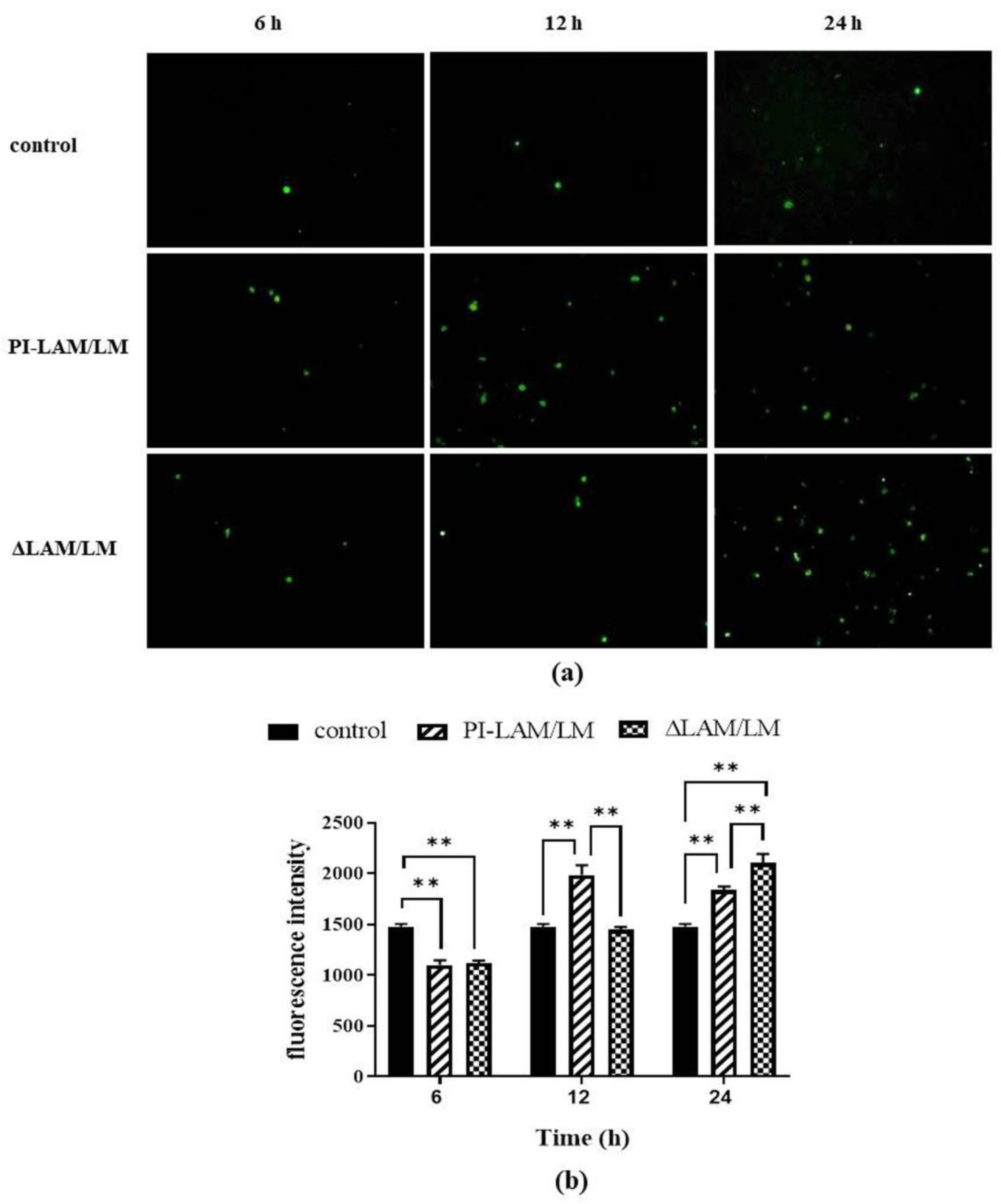
3.6. LAM/LM Molecules Affected the Release of ROS
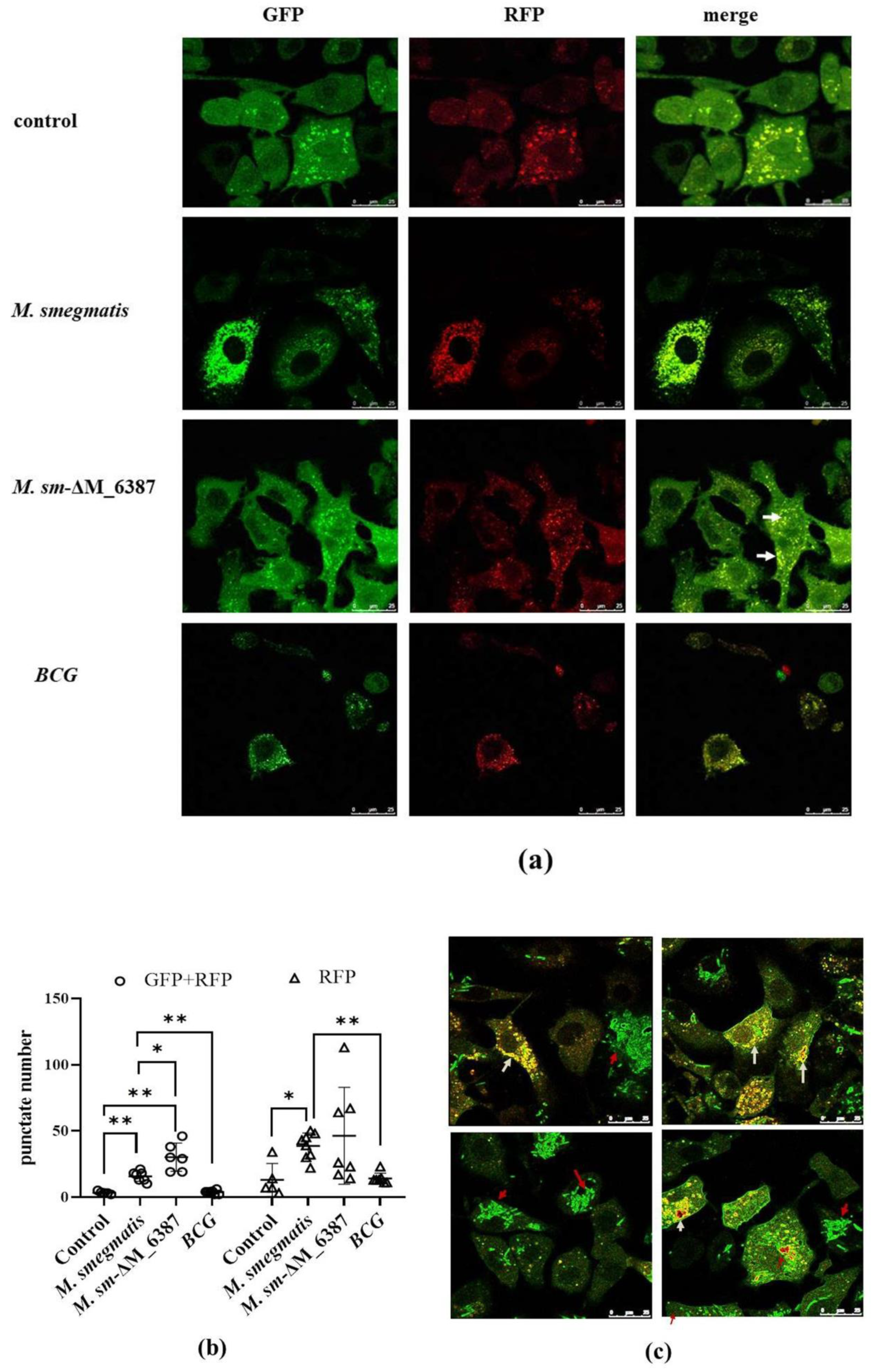
3.7. Recruitment and Localization of LC3 Molecules within Infected Alveolar Epithelial Cells
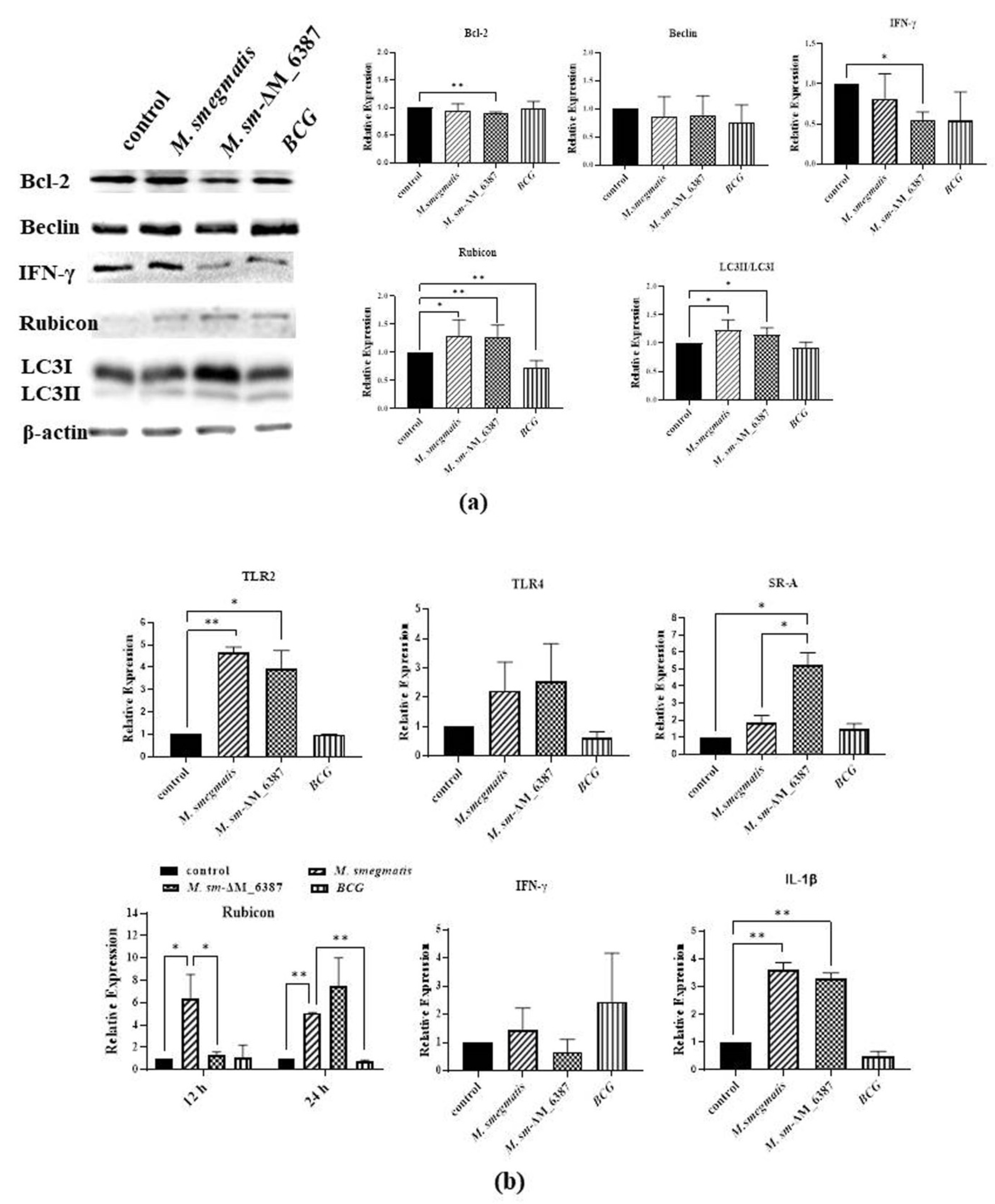
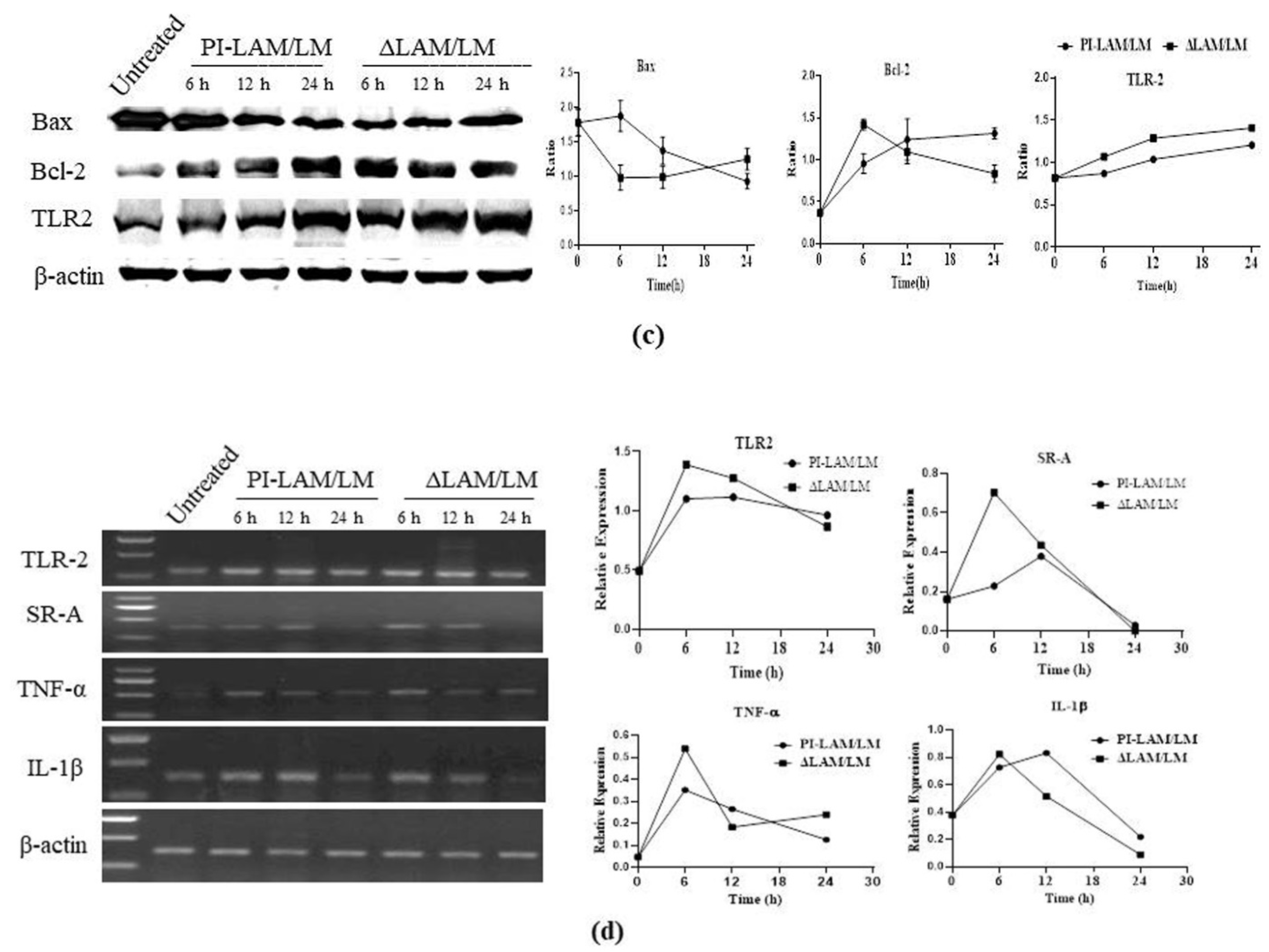
3.8. LAM/LM Molecules Regulated Gene Expression Related to Pathogenic Recognition and Immune Responses
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Global Tuberculosis Report 2021; World Health Organization: Geneva, Switzerland, 2021. [Google Scholar]
- Chakaya, J.; Khan, M.; Ntoumi, F.; Aklillu, E.; Fatima, R.; Mwaba, P.; Kapata, N.; Mfinanga, S.; Hasnain, S.E.; Katoto, P.D.M.C.; et al. Global Tuberculosis Report 2020—Reflections on the Global TB burden, treatment and prevention efforts. Int. J. Infect. Dis. 2021, 113, S7–S12. [Google Scholar] [CrossRef]
- Grayfer, L.; Hodgkinson, J.W.; Belosevic, M. Antimicrobial responses of teleost phagocytes and innate immune evasion strategies of intracellular bacteria. Dev. Comp. Immunol. 2014, 43, 223–242. [Google Scholar] [CrossRef] [PubMed]
- Ryndak, M.B.; Laal, S. Mycobacterium tuberculosis Primary Infection and Dissemination: A Critical Role for Alveolar Epithelial Cells. Front. Cell Infect. Microbiol. 2019, 9, 299. [Google Scholar] [CrossRef]
- Scordo, J.M.; Knoell, D.L.; Torrelles, J.B. Alveolar Epithelial Cells in Mycobacterium tuberculosis Infection: Active Players or Innocent Bystanders? J. Innate. Immun. 2016, 8, 3–14. [Google Scholar] [CrossRef]
- Paik, S.; Kim, J.K.; Chung, C.; Jo, E.K. Autophagy: A new strategy for host-directed therapy of tuberculosis. Virulence 2019, 10, 448–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kramarska, E.; Squeglia, F.; de Maio, F.; Delogu, G.; Berisio, R. PE_PGRS33, an Important Virulence Factor of Mycobacterium tuberculosis and Potential Target of Host Humoral Immune Response. Cells 2021, 10, 161. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.; Raqib, R.; Guðmundsson, G.H.; Bergman, P.; Agerberth, B.; Rekha, R.S. Host-Directed Therapy as a Novel Treatment Strategy to Overcome Tuberculosis: Targeting Immune Modulation. Antibiotics 2020, 9, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhatt, K.; Bhagavathula, M.; Verma, S.; Timmins, G.S.; Deretic, V.P.; Ellner, J.J.; Salgame, P. Rapamycin modulates pulmonary pathology in a murine model of Mycobacterium tuberculosis infection. Dis. Model. Mech. 2021, 14, dmm049018. [Google Scholar] [CrossRef] [PubMed]
- Young, C.; Walzl, G.; Plessis, N.D. Therapeutic host-directed strategies to improve outcome in tuberculosis. Mucosal. Immunol. 2020, 13, 190–204. [Google Scholar] [CrossRef] [Green Version]
- Gutierrez, M.G.; Master, S.S.; Singh, S.B.; Taylor, G.A.; Colombo, M.I.; Deretic, V. Autophagy is a defense mechanism inhibiting BCG and Mycobacterium tuberculosis survival in infected macrophages. Cell 2004, 119, 753–766. [Google Scholar] [CrossRef] [Green Version]
- Scarlatti, F.; Maffei, R.; Beau, I.; Ghidoni, R.; Codogno, P. Non-canonical autophagy: An exception or an underestimated form of autophagy? Autophagy 2008, 4, 1083–1085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Z.; Klionsky, D.J. Autophagosome formation: Core machinery and adaptations. Nat. Cell Biol. 2007, 9, 1102–1109. [Google Scholar] [CrossRef]
- Inomata, M.; Xu, S. Macrophage LC3-associated phagocytosis is an immune defense against Streptococcus pneumoniae that diminishes with host aging. Proc. Natl. Acad. Sci. USA 2020, 117, 33561–33569. [Google Scholar] [CrossRef] [PubMed]
- Köster, S.; Upadhyay, S.; Chandra, P.; Papavinasasundaram, K.; Yang, G.; Hassan, A.; Grigsby, S.J.; Mittal, E.; Park, H.S.; Jones, V.; et al. Mycobacterium tuberculosis is protected from NADPH oxidase and LC3-associated phagocytosis by the LCP protein CpsA. Proc. Natl. Acad. Sci. USA 2017, 114, E8711–E8720. [Google Scholar] [CrossRef] [Green Version]
- Upadhyay, S.; Philips, J.A. LC3-associated phagocytosis: Host defense and microbial response. Curr. Opin. Immunol. 2019, 60, 81–90. [Google Scholar] [CrossRef]
- Castillo, E.F.; Dekonenko, A.; Arko-Mensah, J.; Mandell, M.A.; Dupont, N.; Jiang, S.; Delgado-Vargas, M.; Timmins, G.S.; Bhattacharya, D.; Yang, H.; et al. Autophagy protects against active tuberculosis by suppressing bacterial burden and inflammation. Proc. Natl. Acad. Sci. USA 2012, 109, E3168–E3176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.-S.; Lee, J.-S.; Rodgers, M.; Min, C.-K.; Lee, J.-Y.; Kim, H.J.; Lee, K.-H.; Kim, C.-J.; Oh, B.; Zandi, E.; et al. Autophagy protein Rubicon mediates phagocytic NADPH oxidase activation in response to microbial infection or TLR stimulation. Cell Host. Microbe. 2012, 11, 264–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vergne, I.; Gilleron, M.; Nigou, J. Manipulation of the endocytic pathway and phagocyte functions by Mycobacterium tuberculosis lipoarabinomannan. Front. Cell Infect. Microbiol. 2014, 4, 187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsunaga, K.; Saitoh, T.; Tabata, K.; Omori, H.; Satoh, T.; Kurotori, N.; Maejima, I.; Shirahama-Noda, K.; Ichimura, T.; Isobe, T.; et al. Two Beclin 1-binding proteins, Atg14L and Rubicon, reciprocally regulate autophagy at different stages. Nat. Cell Biol. 2009, 11, 385–396. [Google Scholar] [CrossRef]
- Ogawa, M.; Takada, N.; Shizukuishi, S.; Tomokiyo, M.; Chang, B.; Yoshida, M.; Kakuta, S.; Tanida, I.; Ryo, A.; Guan, J.; et al. Streptococcus pneumoniae triggers hierarchical autophagy through reprogramming of LAPosome-like vesicles via NDP52-delocalization. Commun. Biol. 2020, 3, 25. [Google Scholar] [CrossRef] [Green Version]
- Köster, S.; Upadhyay, S.; Philips, J.A. Why macrophages cannot LAP up TB. Autophagy 2018, 14, 552–554. [Google Scholar] [CrossRef]
- Sanjuan, M.A.; Dillon, C.P.; Tait, S.W.G.; Moshiach, S.; Dorsey, F.; Connell, S.; Komatsu, M.; Tanaka, K.; Cleveland, J.L.; Withoff, S.; et al. Toll-like receptor signalling in macrophages links the autophagy pathway to phagocytosis. Nature 2007, 450, 1253–1257. [Google Scholar] [CrossRef]
- Cadwell, K.; Philips, J.A. Autophagy meets phagocytosis. Immunity 2013, 39, 425–427. [Google Scholar] [CrossRef] [Green Version]
- Bonilla, D.L.; Bhattacharya, A.; Sha, Y.; Xu, Y.; Xiang, Q.; Kan, A.; Jagannath, C.; Komatsu, M.; Eissa, N.T. Autophagy regulates phagocytosis by modulating the expression of scavenger receptors. Immunity 2013, 39, 537–547. [Google Scholar] [CrossRef] [Green Version]
- Barry, C.E.; Crick, D.C.; McNeil, M.R. Targeting the formation of the cell wall core of M. tuberculosis. Infect. Disord. Drug Targets 2007, 7, 182–202. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, D. The mycobacterial cell wall: Structure, biosynthesis and sites of drug action. Curr. Opin. Chem. Biol. 1997, 1, 579–588. [Google Scholar] [CrossRef]
- Turner, J.; Torrelles, J.B. Mannose-capped lipoarabinomannan in Mycobacterium tuberculosis pathogenesis. Pathog. Dis. 2018, 76, fty026. [Google Scholar] [CrossRef] [Green Version]
- Feng, X.; Lu, J.; He, Z.; Wang, Y.; Qi, F.; Pi, R.; Zhang, G. Mycobacterium smegmatis Induces Neurite Outgrowth and Differentiation in an Autophagy-Independent Manner in PC12 and C17.2 Cells. Front. Cell Infect. Microbiol. 2018, 8, 201. [Google Scholar] [CrossRef]
- Berg, S.; Kaur, D.; Jackson, M.; Brennan, P.J. The glycosyltransferases of Mycobacterium tuberculosis-roles in the synthesis of arabinogalactan, lipoarabinomannan, and other glycoconjugates. Glycobiology 2007, 17, 35R–56R. [Google Scholar] [CrossRef] [PubMed]
- Cui, Z.; Li, Y.; Cheng, S.; Yang, H.; Lu, J.; Hu, Z.; Ge, B. Mutations in the embC-embA intergenic region contribute to Mycobacterium tuberculosis resistance to ethambutol. Antimicrob. Agents Chemother. 2014, 58, 6837–6843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korkegian, A.; Roberts, D.M.; Blair, R.; Parish, T. Mutations in the essential arabinosyltransferase EmbC lead to alterations in Mycobacterium tuberculosis lipoarabinomannan. J. Biol. Chem. 2014, 289, 35172–35181. [Google Scholar] [CrossRef] [Green Version]
- Torrelles, J.B.; Khoo, K.H.; Sieling, P.A.; Modlin, R.L.; Zhang, N.; Marques, A.M.; Treumann, A.; Rithner, C.D.; Brennan, P.J.; Chatterjee, D. Truncated structural variants of lipoarabinomannan in Mycobacterium leprae and an ethambutol-resistant strain of Mycobacterium tuberculosis. J. Biol. Chem. 2004, 279, 41227–41239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aliprantis, A.O.; Yang, R.B.; Mark, M.R.; Suggett, S.; Devaux, B.; Radolf, J.D.; Klimpel, G.R.; Godowski, P.; Zychlinsky, A. Cell activation and apoptosis by bacterial lipoproteins through toll-like receptor-2. Science 1999, 285, 736–739. [Google Scholar] [CrossRef] [PubMed]
- Shui, W.; Petzold, C.J.; Redding, A.; Liu, J.; Pitcher, A.; Sheu, L.; Hsieh, T.-y.; Keasling, J.D.; Bertozzi, C.R. Organelle membrane proteomics reveals differential influence of mycobacterial lipoglycans on macrophage phagosome maturation and autophagosome accumulation. J. Proteome. Res. 2011, 10, 339–348. [Google Scholar] [CrossRef]
- Dao, D.N.; Kremer, L.; Guérardel, Y.; Molano, A.; Jacobs, W.R., Jr.; Porcelli, S.A.; Briken, V. Mycobacterium tuberculosis lipomannan induces apoptosis and interleukin-12 production in macrophages. Infect. Immun. 2004, 72, 2067–2074. [Google Scholar] [CrossRef] [Green Version]
- Roca, F.J.; Whitworth, L.J.; Redmond, S.; Jones, A.A.; Ramakrishnan, L. TNF Induces Pathogenic Programmed Macrophage Necrosis in Tuberculosis through a Mitochondrial-Lysosomal-Endoplasmic Reticulum Circuit. Cell 2019, 178, 1344–1361.e11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vignal, C.; Guérardel, Y.; Kremer, L.; Masson, M.; Legrand, D.; Mazurier, J.; Elass, E. Lipomannans, but not lipoarabinomannans, purified from Mycobacterium chelonae and Mycobacterium kansasii induce TNF-alpha and IL-8 secretion by a CD14-toll-like receptor 2-dependent mechanism. J. Immunol. 2003, 171, 2014–2023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, C.; Qu, Z.L.; Tang, X.L.; Liu, Q.; Luo, W.; Huang, C.; Pan, Q.; Zhang, X.L. Mycobacterium tuberculosis Mannose-Capped Lipoarabinomannan Induces IL-10-Producing B Cells and Hinders CD4(+)Th1 Immunity. iScience 2019, 11, 13–30. [Google Scholar] [CrossRef] [Green Version]
- O’Connor, R.A.; Wittmer, S.; Dalton, D.K. Infection-induced apoptosis deletes bystander CD4+ T cells: A mechanism for suppression of autoimmunity during BCG infection. J. Autoimmun. 2005, 24, 93–100. [Google Scholar] [CrossRef]
- Paik, S.; Jo, E.K. An Interplay Between Autophagy and Immunometabolism for Host Defense Against Mycobacterial Infection. Front. Immunol. 2020, 11, 603951. [Google Scholar] [CrossRef]
- Shi, L.; Berg, S.; Lee, A.; Spencer, J.S.; Zhang, J.; Vissa, V.; McNeil, M.R.; Khoo, K.-H.; Chatterjee, D. The carboxy terminus of EmbC from Mycobacterium smegmatis mediates chain length extension of the arabinan in lipoarabinomannan. J. Biol. Chem. 2006, 281, 19512–19526. [Google Scholar] [CrossRef] [Green Version]
- Augenstreich, J.; Briken, V. Host Cell Targets of Released Lipid and Secreted Protein Effectors of Mycobacterium tuberculosis. Front. Cell Infect. Microbiol. 2020, 10, 595029. [Google Scholar] [CrossRef]
- Li, W.; Xin, Y.; McNeil, M.R.; Ma, Y. rmlB and rmlC genes are essential for growth of mycobacteria. Biochem. Biophys. Res. Commun. 2006, 342, 170–178. [Google Scholar] [CrossRef]
- Jiang, T.; He, L.; Zhan, Y.; Zang, S.; Ma, Y.; Zhao, X.; Zhang, C.; Xin, Y. The effect of MSMEG_6402 gene disruption on the cell wall structure of Mycobacterium smegmatis. Microb. Pathog. 2011, 51, 156–160. [Google Scholar] [CrossRef]
- Banaei, N.; Kincaid, E.Z.; Lin, S.-G.; Desmond, E.; William, R.J., Jr.; Ernst, J.D. Lipoprotein processing is essential for resistance of Mycobacterium tuberculosis to malachite green. Antimicrob. Agents Chemother. 2009, 53, 3799–3802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukuda, T.; Matsumura, T.; Ato, M.; Hamasaki, M.; Nishiuchi, Y.; Murakami, Y.; Maeda, Y.; Yoshimori, T.; Matsumoto, S.; Kobayashi, K.; et al. Critical roles for lipomannan and lipoarabinomannan in cell wall integrity of mycobacteria and pathogenesis of tuberculosis. mBio 2013, 4, e00472-12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Küssau, T.; van Wyk, N.; Johansen, M.D.; Alsarraf, H.M.A.B.; Neyret, A.; Hamela, C.; Sørensen, K.K.; Thygesen, M.B.; Beauvineau, C.; Kremer, L.; et al. Functional Characterization of the N-Acetylmuramyl-l-Alanine Amidase, Ami1, from Mycobacterium abscessus. Cell 2020, 9, 2410. [Google Scholar] [CrossRef] [PubMed]
- Eitson, J.L.; Medeiros, J.J.; Hoover, A.R.; Srivastava, S.; Roybal, K.T.; Aínsa, J.A.; Hansen, E.J.; Gumbo, T.; van Oers, N.S.C. Mycobacterial shuttle vectors designed for high-level protein expression in infected macrophages. Appl. Environ. Microbiol. 2012, 78, 6829–6837. [Google Scholar] [CrossRef] [Green Version]
- Şentürk, M.; Lin, G.; Zuo, Z.; Mao, D.; Watson, E.; Mikos, A.G.; Bellen, H.J. Ubiquilins regulate autophagic flux through mTOR signalling and lysosomal acidification. Nat. Cell Biol. 2019, 21, 384–396. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wang, X.; Vikash, V.; Ye, Q.; Wu, D.; Liu, Y.; Dong, W. ROS and ROS-Mediated Cellular Signaling. Oxid. Med. Cell. Longev. 2016, 2016, 4350965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Correia-Neves, M.; Sundling, C.; Cooper, A.; Källenius, G. Lipoarabinomannan in Active and Passive Protection against Tuberculosis. Front. Immunol. 2019, 10, 1968. [Google Scholar] [CrossRef]
- Yan, Z.H.; Zhao, B.; Pang, Y.; Wang, X.J.; Yi, L.; Wang, H.L.; Yang, B.; Wei, P.J.; Jia, H.Y.; Li, S.P.; et al. Generation of mycobacterial lipoarabinomannan-specific monoclonal antibodies and their ability to identify mycobacterium isolates. J. Microbiol. Immunol. Infect. 2021, 54, 437–446. [Google Scholar] [CrossRef]
- Rodrigues, T.S.; Conti, B.J.; Fraga-Silva, T.F.d.; Almeida, F.; Bonato, V.L.D. Interplay between alveolar epithelial and dendritic cells and Mycobacterium tuberculosis. J. Leukoc. Biol. 2020, 108, 1139–1156. [Google Scholar] [CrossRef] [PubMed]
- Nigou, J.; Gilleron, M.; Rojas, M.; García, L.F.; Thurnher, M.; Puzo, G. Mycobacterial lipoarabinomannans: Modulators of dendritic cell function and the apoptotic response. Microbe. Infect. 2002, 4, 945–953. [Google Scholar] [CrossRef]
- Wu, M.Y.; Lu, J.H. Autophagy and Macrophage Functions: Inflammatory Response and Phagocytosis. Cells 2019, 9, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Wu, Q.; Lei, H.; Zheng, H.; Zhou, F.; Sun, Z.; Zhao, J.; Yu, X.; Zhang, S. Mannosylated structures of mycobacterial lipoarabinomannans facilitate the maturation and activation of dendritic cells. Cell Immunol. 2019, 335, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Brumell, J.H. Bacteria-autophagy interplay: A battle for survival. Nat. Rev. Microbiol. 2014, 12, 101–114. [Google Scholar] [CrossRef]
- Sharma, M.; Sharma, S.; Roy, S.; Varma, S.; Bose, M. Pulmonary epithelial cells are a source of interferon-gamma in response to Mycobacterium tuberculosis infection. Immunol. Cell Biol. 2007, 85, 229–237. [Google Scholar] [CrossRef]
- Miralda, I.; Klaes, C.K.; Graham, J.E.; Uriarte, S.M. Human Neutrophil Granule Exocytosis in Response to Mycobacterium smegmatis. Pathogens 2020, 9, 123. [Google Scholar] [CrossRef] [Green Version]
- García-Pérez, B.E.; Hernández-González, J.C.; García-Nieto, S.; Luna-Herrera, J. Internalization of a non-pathogenic mycobacteria by macropinocytosis in human alveolar epithelial A549 cells. Microb. Pathog. 2008, 45, 1–6. [Google Scholar] [CrossRef]
- Bowdish, D.M.E.; Sakamoto, K.; Kim, M.-J.; Kroos, M.; Mukhopadhyay, S.; Leifer, C.A.; Tryggvason, K.; Gordon, S.; Russell, D.G. MARCO, TLR2, and CD14 are required for macrophage cytokine responses to mycobacterial trehalose dimycolate and Mycobacterium tuberculosis. PLoS Pathog. 2009, 5, e1000474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inoue, M.; Niki, M.; Ozeki, Y.; Nagi, S.; Chadeka, E.A.; Yamaguchi, T.; Osada-Oka, M.; Ono, K.; Oda, T.; Mwende, F.; et al. High-density lipoprotein suppresses tumor necrosis factor alpha production by mycobacteria-infected human macrophages. Sci. Rep. 2018, 8, 6736. [Google Scholar] [CrossRef] [PubMed] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequences (5′–3′) | Annealing Temperature | Product Length (bp) |
|---|---|---|---|
| MSMEG_6387 | F: CCGACCCTGCTGAAACTGCT | 58 | 494 |
| R: AGCCAGAACGCCAGGAACAG | |||
| MSMEG_6388 | F: GTCGGTGCGCATCAAGTACG | 58 | 680 |
| R: CCTTGACCATCGAGCCGAGT | |||
| MSMEG_6389 | F: CGATCGTGTCGACCGTCATC | 58 | 292 |
| R: ATGAGCGCCAGCACGTTGTA | |||
| MSMEG_6390 | F: CTGGCTGGTGCTCGAACTCA | 58 | 773 |
| R: CCAGCACACATCCGTTGAGG | |||
| gap | F: GGAAAGCTGTGGCGTGATGG | 54 | 399 |
| R: GTAGGCCATGAGGTCCACCA |
| Gene | Primer Sequences (5′–3′) | Product Length (bp) |
|---|---|---|
| TLR-2 | F: GGAATCGGTGAGGTCCTGTCCTG | 296 |
| R: GGCGTCACATGCAGAAAGCCC | ||
| TLR-4 | F: AAGTGTCTGAACTCCCTCCAGG | 278 |
| R: ATGGTCTTATTCATCTGACAGGTGATA | ||
| SR-A | F: ATGTCCGTTCAGCGTCTT | 380 |
| R: TAGGTCCTGATGCTTCTTTA | ||
| Rubicon | R: AGTGGGTTACTTGGGAGTG | 310 |
| F: CTTTGGCTAATAGTTCTGC | ||
| IFN-γ | F: TCGGTAACTGACTTGAATGTCCA | 93 |
| R: TCGCTTCCCTGTTTTAGCTGC | ||
| IL-1β | F: GGACAAGCTGAGGAAGATGC | 360 |
| R: TCTTTCAACACGCAGGACAG | ||
| TNF-α | F: TGCTTGTTCCTCAGCCTCTT | 514 |
| R: GGAAGACCCCTCCCAGATAG | ||
| β-actin | F: CATGGATGATGATATCGCCGCG | 371 |
| R: ACATGATCTGGGTCATCTTCTCG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, H.; Gui, X.; Chen, S.; Fu, W.; Li, X.; Xiao, T.; Hou, J.; Jiang, T. Structural Variability of Lipoarabinomannan Modulates Innate Immune Responses within Infected Alveolar Epithelial Cells. Cells 2022, 11, 361. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11030361
Liu H, Gui X, Chen S, Fu W, Li X, Xiao T, Hou J, Jiang T. Structural Variability of Lipoarabinomannan Modulates Innate Immune Responses within Infected Alveolar Epithelial Cells. Cells. 2022; 11(3):361. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11030361
Chicago/Turabian StyleLiu, Hanrui, Xuwen Gui, Shixing Chen, Weizhe Fu, Xiang Li, Tingyuan Xiao, Jie Hou, and Tao Jiang. 2022. "Structural Variability of Lipoarabinomannan Modulates Innate Immune Responses within Infected Alveolar Epithelial Cells" Cells 11, no. 3: 361. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11030361