
BMP9 Promotes an Epithelial Phenotype and a Hepatocyte-like Gene Expression Profile in Adult Hepatic Progenitor Cells

 and
and 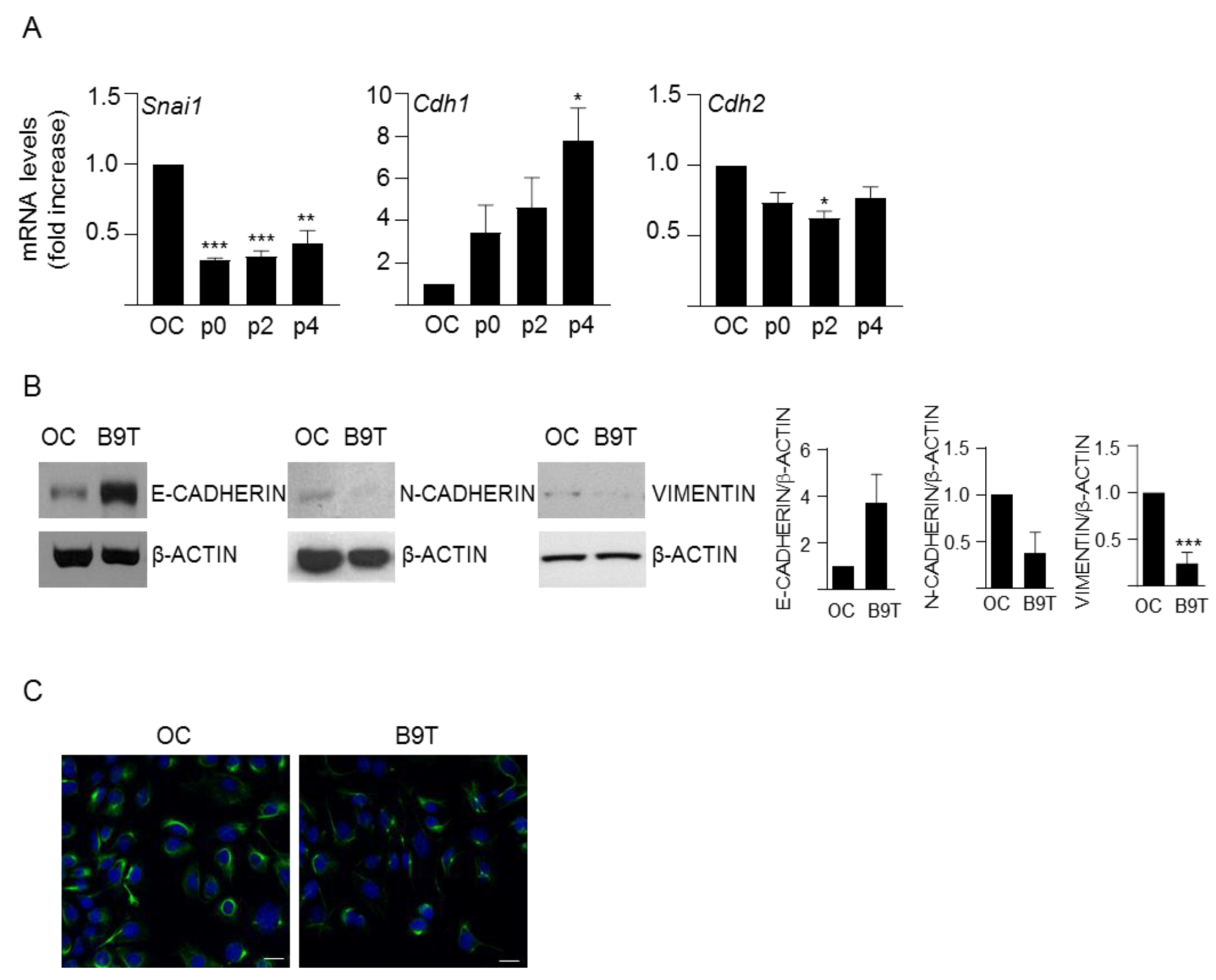{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines and Culture Conditions
2.2. Confocal Microscopy Analysis
2.3. Analysis of Cell Number
2.4. Measurement of Apoptotic Index
2.5. Measurement of Caspase-3-like Enzymatic Activity
2.6. RNA Isolation and Quantitative Reverse Transcriptase-Polymerase Chain Reaction
2.7. Clonogenic Assay
2.8. Soft Agar Assays
2.9. Protein Isolation and Western Blot Analysis
2.10. Analysis of Urea Production
2.11. Statistical Analysis
3. Results
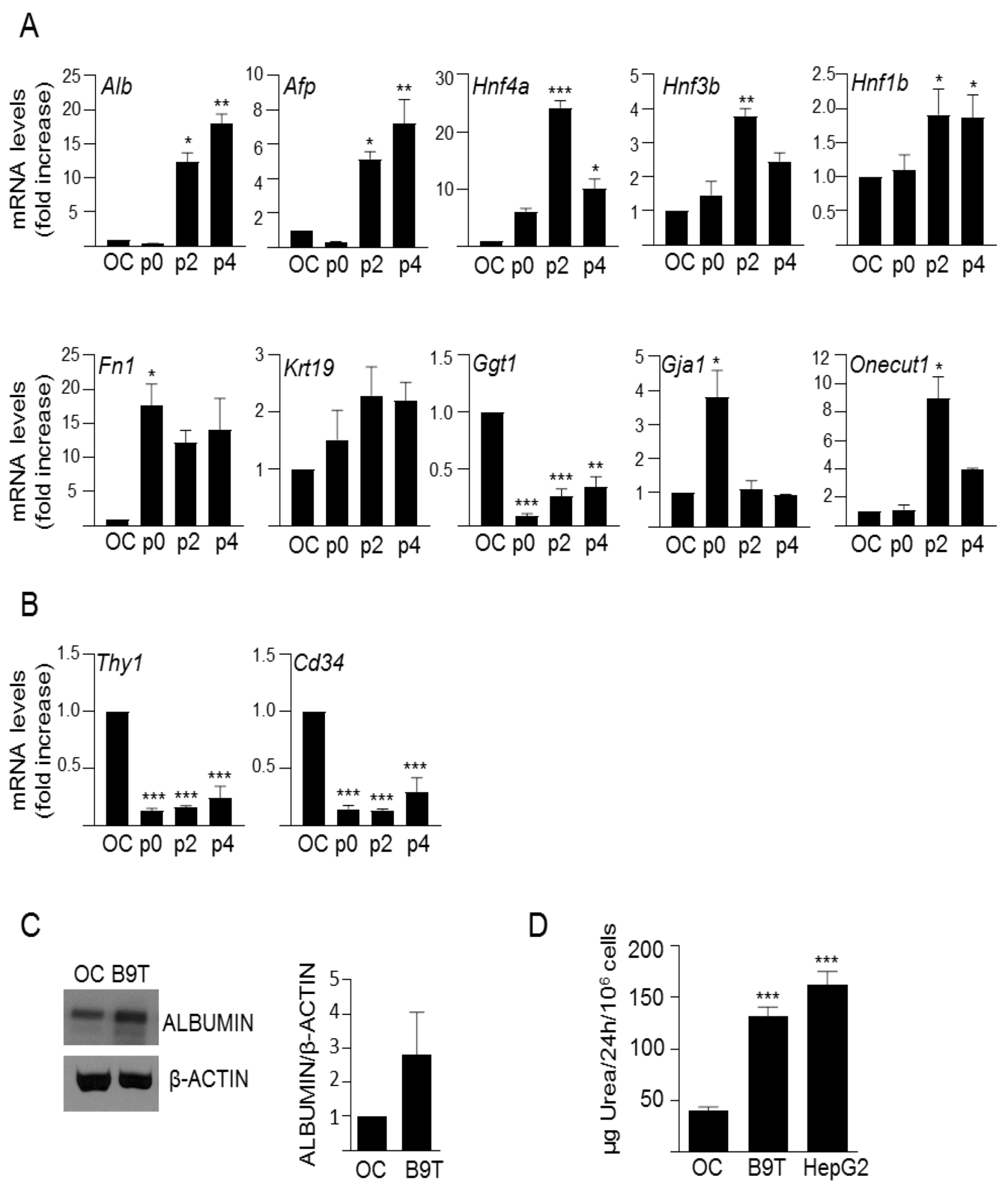
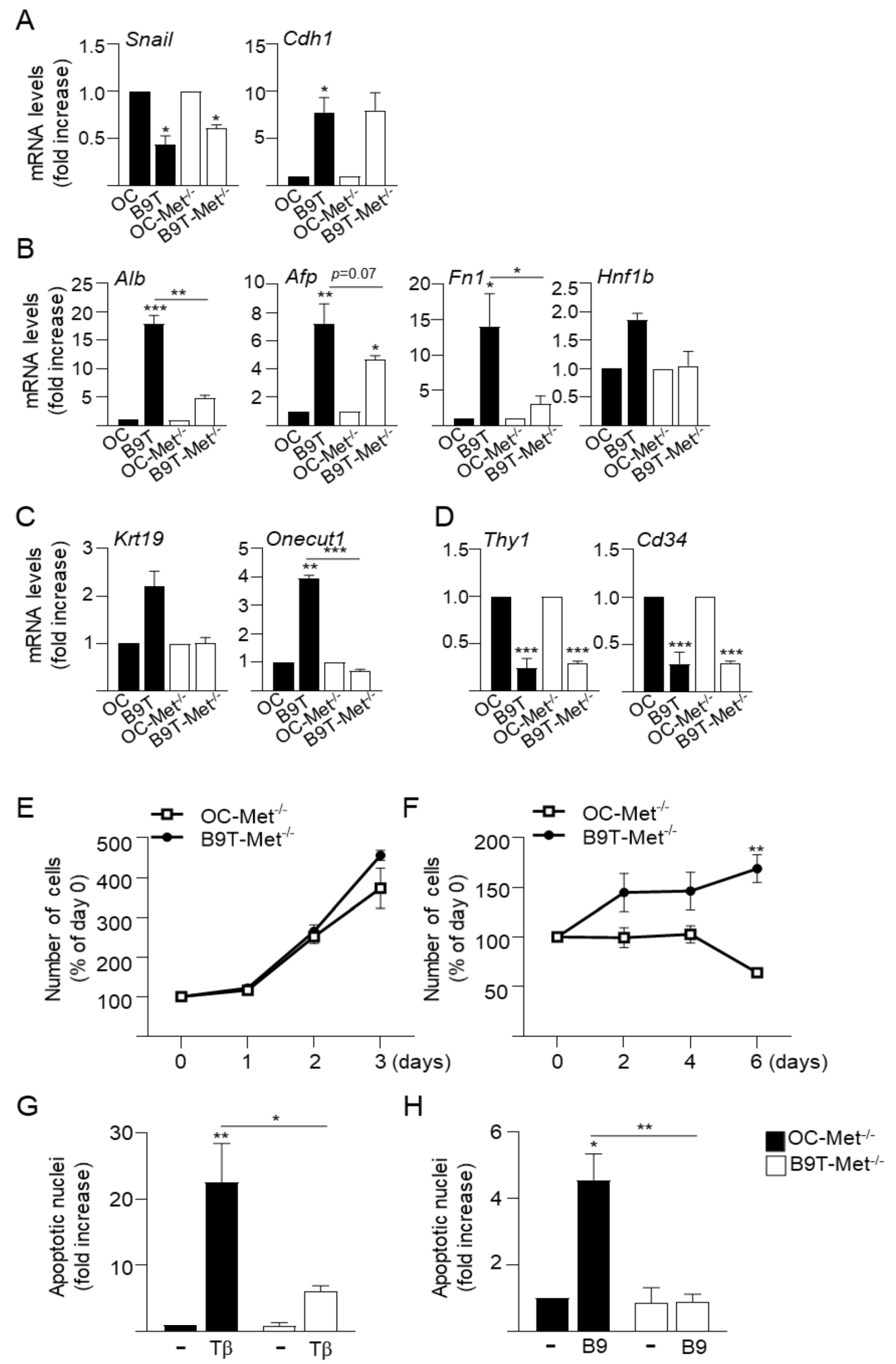
3.1. Chronic Exposure to BMP9 Leads to Acquisition of Hepatocyte-like Properties in HPC/Oval Cells
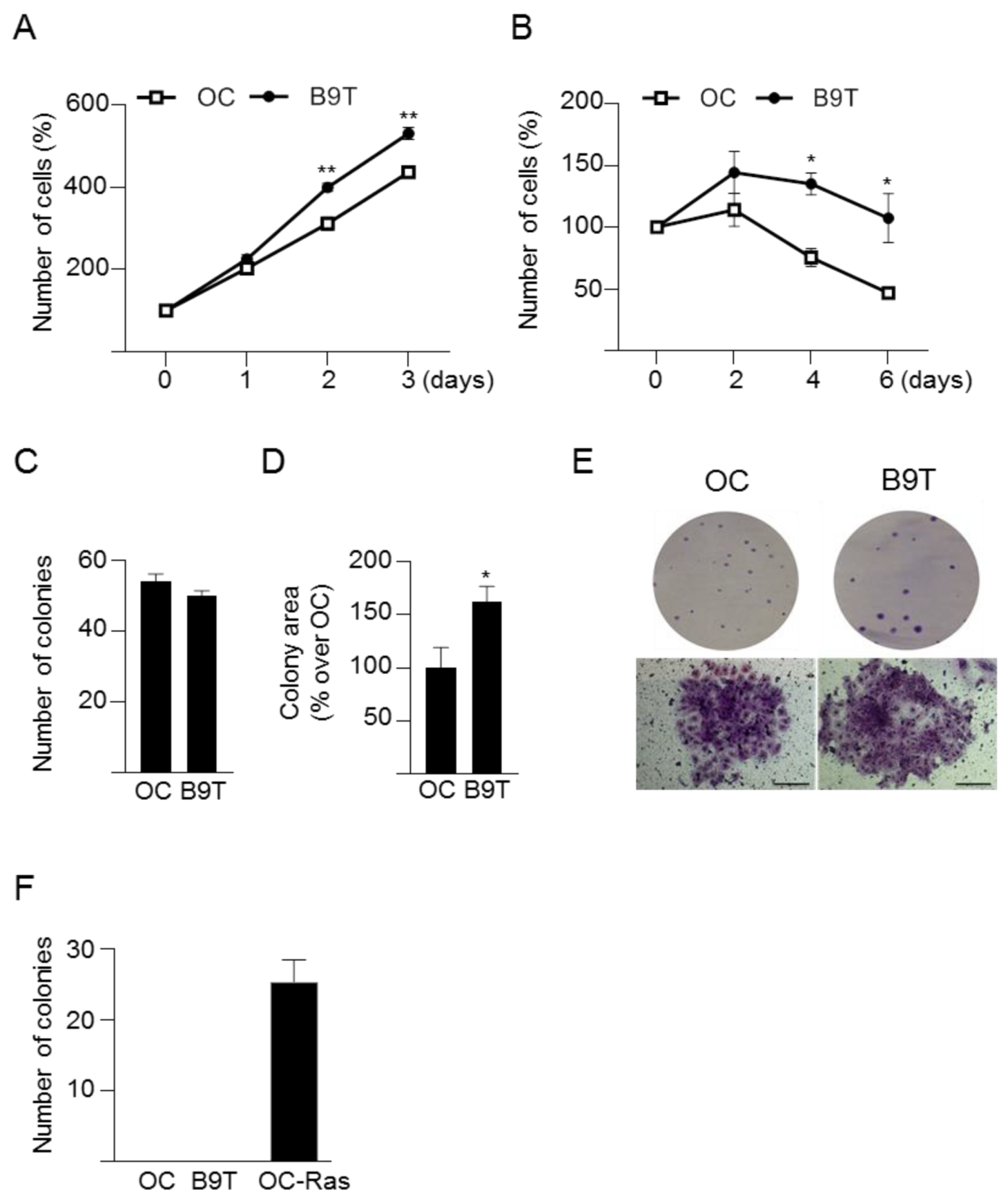
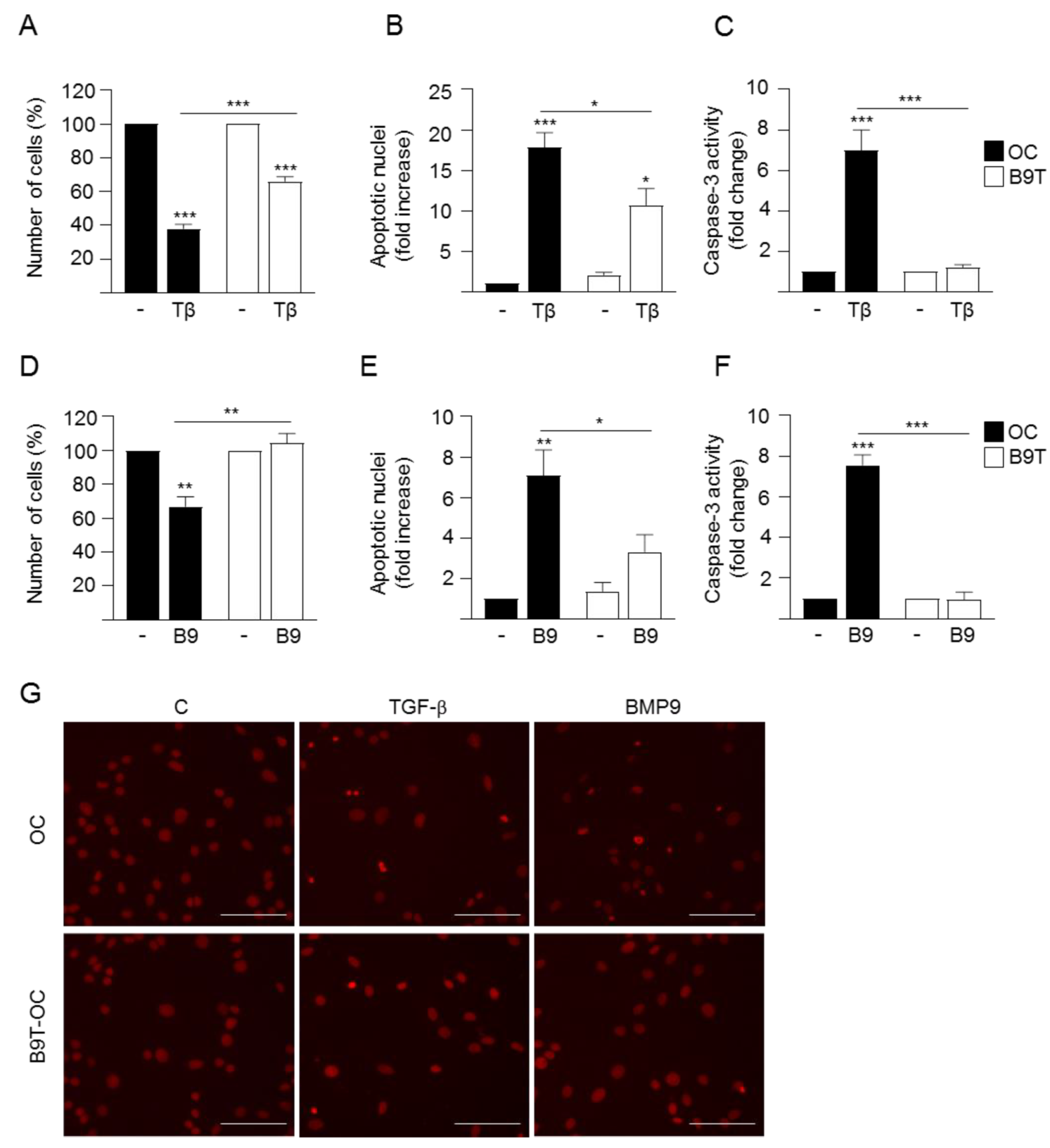
3.2. Chronic Exposure to BMP9 Confers Growth and Survival Advantages to HPC/Oval Cells
3.3. c-Met Signaling Contributes to the Acquisition of B9T-OC Properties
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Desroches-Castan, A.; Tillet, E.; Ricard, N.; Ouarné, M.; Mallet, C.; Belmudes, L.; Couté, Y.; Boillot, O.; Scoazec, J.-Y.; Bailly, S.; et al. Bone Morphogenetic Protein 9 Is a Paracrine Factor Controlling Liver Sinusoidal Endothelial Cell Fenestration and Protecting Against Hepatic Fibrosis. Hepatology 2019, 70, 1392–1408. [Google Scholar] [CrossRef]
- Desroches-Castan, A.; Tillet, E.; Ricard, N.; Ouarné, M.; Mallet, C.; Feige, J.-J.; Bailly, S. Differential Consequences of Bmp9 Deletion on Sinusoidal Endothelial Cell Differentiation and Liver Fibrosis in 129/Ola and C57BL/6 Mice. Cells 2019, 8, 1079. [Google Scholar] [CrossRef] [Green Version]
- Herrera, B.; Addante, A.; Sánchez, A. BMP Signalling at the Crossroad of Liver Fibrosis and Regeneration. Int. J. Mol. Sci. 2017, 19, 39. [Google Scholar] [CrossRef] [Green Version]
- Herrera, B.; Dooley, S.; Breitkopf-Heinlein, K. Potential Roles of Bone Morphogenetic Protein (BMP)-9 in Human Liver Diseases. Int. J. Mol. Sci. 2014, 15, 5199–5220. [Google Scholar] [CrossRef]
- Xiao, X.; Alfaro-Magallanes, V.M.; Babitt, J.L. Bone morphogenic proteins in iron homeostasis. Bone 2020, 138, 115495. [Google Scholar] [CrossRef]
- Chen, H.; Nio, K.; Yamashita, T.; Okada, H.; Li, R.; Suda, T.; Li, Y.; Doan, P.T.B.; Seki, A.; Nakagawa, H.; et al. BMP9-ID1 signaling promotes EpCAM-positive cancer stem cell properties in hepatocellular carcinoma. Mol. Oncol. 2021, 15, 2203–2218. [Google Scholar] [CrossRef]
- Herrera, B.; García-Álvaro, M.; Cruz, S.; Walsh, P.; Fernandez, M.; Roncero, C.; Fabregat, I.; Sanchez, A.; Inman, G.J. BMP9 Is a Proliferative and Survival Factor for Human Hepatocellular Carcinoma Cells. PLoS ONE 2013, 8, e69535. [Google Scholar] [CrossRef]
- Li, Q.; Gu, X.; Weng, H.; Ghafoory, S.; Liu, Y.; Feng, T.; Dzieran, J.; Li, L.; Ilkavets, I.; Kruithof-de Julio, M.; et al. Bone morphogenetic protein-9 (BMP-9) induces epithelial to mesenchymal transition (EMT) in hepatocellular carcinoma cells. Cancer Sci. 2013, 104, 398–408. [Google Scholar] [CrossRef]
- Jiang, Q.; Li, Q.; Liu, B.; Li, G.; Riedemann, G.; Gaitantzi, H.; Breitkopf-Heinlein, K.; Zeng, A.; Ding, H.; Xu, K. BMP9 promotes methionine- and choline-deficient diet-induced nonalcoholic steatohepatitis in non-obese mice by enhancing NF-κB dependent macrophage polarization. Int. Immunopharmacol. 2021, 96, 107591. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.-Q.; Liu, B.-B.; Xu, K.-S. New insights into BMP9 signaling in liver diseases. Mol. Cell. Biochem. 2021, 476, 3591–3600. [Google Scholar] [CrossRef]
- Sun, Q.-J.; Cai, L.-Y.; Jian, J.; Cui, Y.-L.; Huang, C.-K.; Liu, S.-Q.; Lu, J.-L.; Wang, W.; Zeng, X.; Zhong, L. The Role of Bone Morphogenetic Protein 9 in Nonalcoholic Fatty Liver Disease in Mice. Front. Pharmacol. 2021, 11, 605967. [Google Scholar] [CrossRef]
- Yang, Z.; Li, P.; Shang, Q.; Wang, Y.; He, J.; Ge, S.; Jia, R.; Fan, X. CRISPR-mediated BMP9 ablation promotes liver steatosis via the down-regulation of PPARα expression. Sci. Adv. 2020, 6, eabc5022. [Google Scholar] [CrossRef]
- Breitkopf-Heinlein, K.; Meyer, C.; König, C.; Gaitantzi, H.; Addante, A.; Thomas, M.; Wiercinska, E.; Cai, C.; Li, Q.; Wan, F.; et al. BMP-9 interferes with liver regeneration and promotes liver fibrosis. Gut 2017, 66, 939–954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, P.; Li, Y.; Zhu, L.; Yang, Z.; He, J.; Wang, L.; Shang, Q.; Pan, H.; Wang, H.; Ma, X.; et al. Targeting secreted cytokine BMP9 gates the attenuation of hepatic fibrosis. Biochim. Biophys. Acta Mol. Basis Dis. 2018, 1864, 709–720. [Google Scholar] [CrossRef] [PubMed]
- Addante, A.; Roncero, C.; Almalé, L.; Lazcanoiturburu, N.; García-Álvaro, M.; Fernández, M.; Sanz, J.; Hammad, S.; Nwosu, Z.C.; Lee, S.; et al. Bone morphogenetic protein 9 as a key regulator of liver progenitor cells in DDC-induced cholestatic liver injury. Liver Int. 2018, 38, 1664–1675. [Google Scholar] [CrossRef]
- Bria, A.; Marda, J.; Zhou, J.; Sun, X.; Cao, Q.; Petersen, B.E.; Pi, L. Hepatic progenitor cell activation in liver repair. Liver Res. 2017, 1, 81–87. [Google Scholar] [CrossRef]
- Ko, S.; Russell, J.O.; Molina, L.M.; Monga, S.P. Liver Progenitors and Adult Cell Plasticity in Hepatic Injury and Repair: Knowns and Unknowns. Annu. Rev. Pathol. Mech. Dis. 2020, 15, 23–50. [Google Scholar] [CrossRef] [Green Version]
- Williams, M.J.; Clouston, A.; Forbes, S.J. Links Between Hepatic Fibrosis, Ductular Reaction, and Progenitor Cell Expansion. Gastroenterology 2014, 146, 349–356. [Google Scholar] [CrossRef]
- Lee, K.W.; Castilho, A.; Ma, S.K.Y.; Ng, I.O.L. Liver cancer stem cells: Implications for a new therapeutic target. Liver Int. 2009, 29, 955–965. [Google Scholar] [CrossRef] [PubMed]
- Carpino, G.; Renzi, A.; Franchitto, A.; Cardinale, V.; Onori, P.; Reid, L.; Alvaro, D.; Gaudio, E. Stem/Progenitor Cell Niches Involved in Hepatic and Biliary Regeneration. Stem Cells Int. 2016, 2016, 3658013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kordes, C.; Häussinger, D. Hepatic stem cell niches. J. Clin. Investig. 2013, 123, 1874–1880. [Google Scholar] [CrossRef] [Green Version]
- Addante, A.; Roncero, C.; Lazcanoiturburu, N.; Méndez, R.; Almalé, L.; García-Álvaro, M.; Dijke, P.T.; Fabregat, I.; Herrera, B.; Sánchez, A. A Signaling Crosstalk between BMP9 and HGF/c-Met Regulates Mouse Adult Liver Progenitor Cell Survival. Cells 2020, 9, 752. [Google Scholar] [CrossRef] [Green Version]
- del Castillo, G.; Factor, V.M.; Fernández, M.; Álvarez-Barrientos, A.; Fabregat, I.; Thorgeirsson, S.S.; Sánchez, A. Deletion of the Met Tyrosine Kinase in Liver Progenitor Oval Cells Increases Sensitivity to Apoptosis in Vitro. Am. J. Pathol. 2008, 172, 1238–1247. [Google Scholar] [CrossRef] [Green Version]
- Almalé, L.; García-Álvaro, M.; Martínez-Palacián, A.; García-Bravo, M.; Lazcanoiturburu, N.; Addante, A.; Roncero, C.; Sanz, J.; de la O López, M.; Bragado, P.; et al. c-Met Signaling Is Essential for Mouse Adult Liver Progenitor Cells Expansion After Transforming Growth Factor-β-Induced Epithelial–Mesenchymal Transition and Regulates Cell Phenotypic Switch. Stem Cells 2019, 37, 1108–1118. [Google Scholar] [CrossRef] [Green Version]
- Suárez-Causado, A.; Caballero-Díaz, D.; Bertrán, E.; Roncero, C.; Addante, A.; García-Álvaro, M.; Fernández, M.; Herrera, B.; Porras, A.; Fabregat, I.; et al. HGF/c-Met signaling promotes liver progenitor cell migration and invasion by an epithelial–mesenchymal transition-independent, phosphatidyl inositol-3 kinase-dependent pathway in an in vitro model. Biochim. Biophys. Acta 2015, 1853, 2453–2463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-Palacián, A.; Del Castillo, G.; Suárez-Causado, A.; García-Álvaro, M.; De La Morena-Frutos, D.; Fernandez, M.; Roncero, C.; Fabregat, I.; Herrera, B.; Sanchez, A. Mouse Hepatic Oval Cells Require Met-Dependent PI3K to Impair TGF-β-Induced Oxidative Stress and Apoptosis. PLoS ONE 2013, 8, e53108. [Google Scholar] [CrossRef] [Green Version]
- Valdés, F.; Alvarez, A.M.; Locascio, A.; Vega, S.; Herrera, B.; Fernández, M.; Benito, M.; Nieto, M.A.; Fabregat, I. The epithelial mesenchymal transition confers resistance to the apoptotic effects of transforming growth factor Beta in fetal rat hepatocytes. Mol. Cancer Res. 2002, 1, 68–78. [Google Scholar]
- Lorenzini, S.; Bird, T.; Boulter, L.; Bellamy, C.; Samuel, K.; Aucott, R.; Clayton, E.; Andreone, P.; Bernardi, M.; Golding, M.; et al. Characterisation of a stereotypical cellular and extracellular adult liver progenitor cell niche in rodents and diseased human liver. Gut 2010, 59, 645–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hengstler, J.G.; Brulport, M.; Schormann, W.; Bauer, A.; Hermes, M.; Nussler, A.K.; Fändrich, F.; Ruhnke, M.; Ungefroren, H.; Griffin, L.; et al. Generation of human hepatocytes by stem cell technology: Definition of the hepatocyte. Expert Opin. Drug Metab. Toxicol. 2005, 1, 61–74. [Google Scholar] [CrossRef]
- Kalluri, R.; Weinberg, R.A. The basics of epithelial-mesenchymal transition. J. Clin. Investig. 2009, 119, 1420–1428. [Google Scholar] [CrossRef] [Green Version]
- Zaret, K.S. Regulatory phases of early liver development: Paradigms of organogenesis. Nat. Rev. Genet. 2002, 3, 499–512. [Google Scholar] [CrossRef]
- Yahoo, N.; Pournasr, B.; Rostamzadeh, J.; Hakhamaneshi, M.S.; Ebadifar, A.; Fathi, F.; Baharvand, H. Forced expression of Hnf1b/Foxa3 promotes hepatic fate of embryonic stem cells. Biochem. Biophys. Res. Commun. 2016, 474, 199–205. [Google Scholar] [CrossRef] [Green Version]
- Masson, N.M.; Currie, I.S.; Terrace, J.D.; Garden, O.J.; Parks, R.W.; Ross, J.A. Hepatic progenitor cells in human fetal liver express the oval cell marker Thy-1. Am. J. Physiol. Liver Physiol. 2006, 291, G45–G54. [Google Scholar] [CrossRef] [PubMed]
- Petersen, B.E.; Grossbard, B.; Hatch, H.; Pi, L.; Deng, J.; Scott, E.W. Mouse A6–positive hepatic oval cells also express several hematopoietic stem cell markers. Hepatology 2003, 37, 632–640. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.-W.; Taniguchi, H. Diversity of Hepatic Stem Cells in the Fetal and Adult Liver. Semin. Liver Dis. 2003, 23, 337–348. [Google Scholar] [CrossRef]
- Bladt, F.; Riethmacher, D.; Isenmann, S.; Aguzzi, A.; Birchmeier, C. Essential role for the c-met receptor in the migration of myogenic precursor cells into the limb bud. Nature 1995, 376, 768–771. [Google Scholar] [CrossRef] [PubMed]
- Uehara, Y.; Minowa, O.; Mori, C.; Shiota, K.; Kuno, J.; Noda, T.; Kitamura, N. Placental defect and embryonic lethality in mice lacking hepatocyte growth factor/scatter factor. Nature 1995, 373, 702–705. [Google Scholar] [CrossRef]
- Prior, N.; Inacio, P.; Huch, M. Liver organoids: From basic research to therapeutic applications. Gut 2019, 68, 2228–2237. [Google Scholar] [CrossRef] [Green Version]
- Kitade, M.; Factor, V.M.; Andersen, J.B.; Tomokuni, A.; Kaji, K.; Akita, H.; Holczbauer, A.; Seo, D.; Marquardt, J.U.; Conner, E.A.; et al. Specific fate decisions in adult hepatic progenitor cells driven by MET and EGFR signaling. Genes Dev. 2013, 27, 1706–1717. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, A.; Zheng, Y.-W.; Fukao, K.; Nakauchi, H.; Taniguchi, H. Liver repopulation by c-Met-positive stem/progenitor cells isolated from the developing rat liver. Hepatogastroenterology 2004, 51, 423–426. [Google Scholar]
- Ishikawa, T.; Factor, V.M.; Marquardt, J.U.; Raggi, C.; Seo, D.; Kitade, M.; Conner, E.A.; Thorgeirsson, S.S. Hepatocyte growth factor/c-metsignaling is required for stem-cell-mediated liver regeneration in mice. Hepatology 2012, 55, 1215–1226. [Google Scholar] [CrossRef] [Green Version]
- Dooley, S.; ten Dijke, P. TGF-β in progression of liver disease. Cell Tissue Res. 2012, 347, 245–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giannelli, G.; Mikulits, W.; Dooley, S.; Fabregat, I.; Moustakas, A.; Dijke, P.T.; Portincasa, P.; Winter, P.; Janssen, R.; Leporatti, S.; et al. The rationale for targeting TGF-β in chronic liver diseases. Eur. J. Clin. Investig. 2016, 46, 349–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sukowati, C.H.C.; Anfuso, B.; Torre, G.; Francalanci, P.; Crocè, L.S.; Tiribelli, C. The Expression of CD90/Thy-1 in Hepatocellular Carcinoma: An In Vivo and In Vitro Study. PLoS ONE 2013, 8, e76830. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.F.; Ho, D.W.; Ng, M.N.; Lau, C.K.; Yu, W.C.; Ngai, P.; Chu, P.W.; Lam, C.T.; Poon, R.T.; Fan, S.T. Significance of CD90+ Cancer Stem Cells in Human Liver Cancer. Cancer Cell 2008, 13, 153–166. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Addante, A.; González-Corralejo, C.; Roncero, C.; Lazcanoiturburu, N.; García-Sáez, J.; Herrera, B.; Sánchez, A. BMP9 Promotes an Epithelial Phenotype and a Hepatocyte-like Gene Expression Profile in Adult Hepatic Progenitor Cells. Cells 2022, 11, 365. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11030365
Addante A, González-Corralejo C, Roncero C, Lazcanoiturburu N, García-Sáez J, Herrera B, Sánchez A. BMP9 Promotes an Epithelial Phenotype and a Hepatocyte-like Gene Expression Profile in Adult Hepatic Progenitor Cells. Cells. 2022; 11(3):365. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11030365
Chicago/Turabian StyleAddante, Annalisa, Carlos González-Corralejo, Cesáreo Roncero, Nerea Lazcanoiturburu, Juan García-Sáez, Blanca Herrera, and Aránzazu Sánchez. 2022. "BMP9 Promotes an Epithelial Phenotype and a Hepatocyte-like Gene Expression Profile in Adult Hepatic Progenitor Cells" Cells 11, no. 3: 365. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11030365