Intra- and Intercellular Quality Control Mechanisms of Mitochondria


Kagawa School of Pharmaceutical Sciences, Tokushima Bunri University, Shido 1314-1, Sanuki, Kagawa 769-2193, Japan
*
Author to whom correspondence should be addressed.
Cells 2018, 7(1), 1; https://0-doi-org.brum.beds.ac.uk/10.3390/cells7010001
Submission received: 30 November 2017
/
Revised: 22 December 2017
/
Accepted: 23 December 2017
/
Published: 26 December 2017
{kind=link}
{kind=link}
Abstract
:Mitochondria function to generate ATP and also play important roles in cellular homeostasis, signaling, apoptosis, autophagy, and metabolism. The loss of mitochondrial function results in cell death and various types of diseases. Therefore, quality control of mitochondria via intra- and intercellular pathways is crucial. Intracellular quality control consists of biogenesis, fusion and fission, and degradation of mitochondria in the cell, whereas intercellular quality control involves tunneling nanotubes and extracellular vesicles. In this review, we outline the current knowledge on the intra- and intercellular quality control mechanisms of mitochondria.
1. Introduction
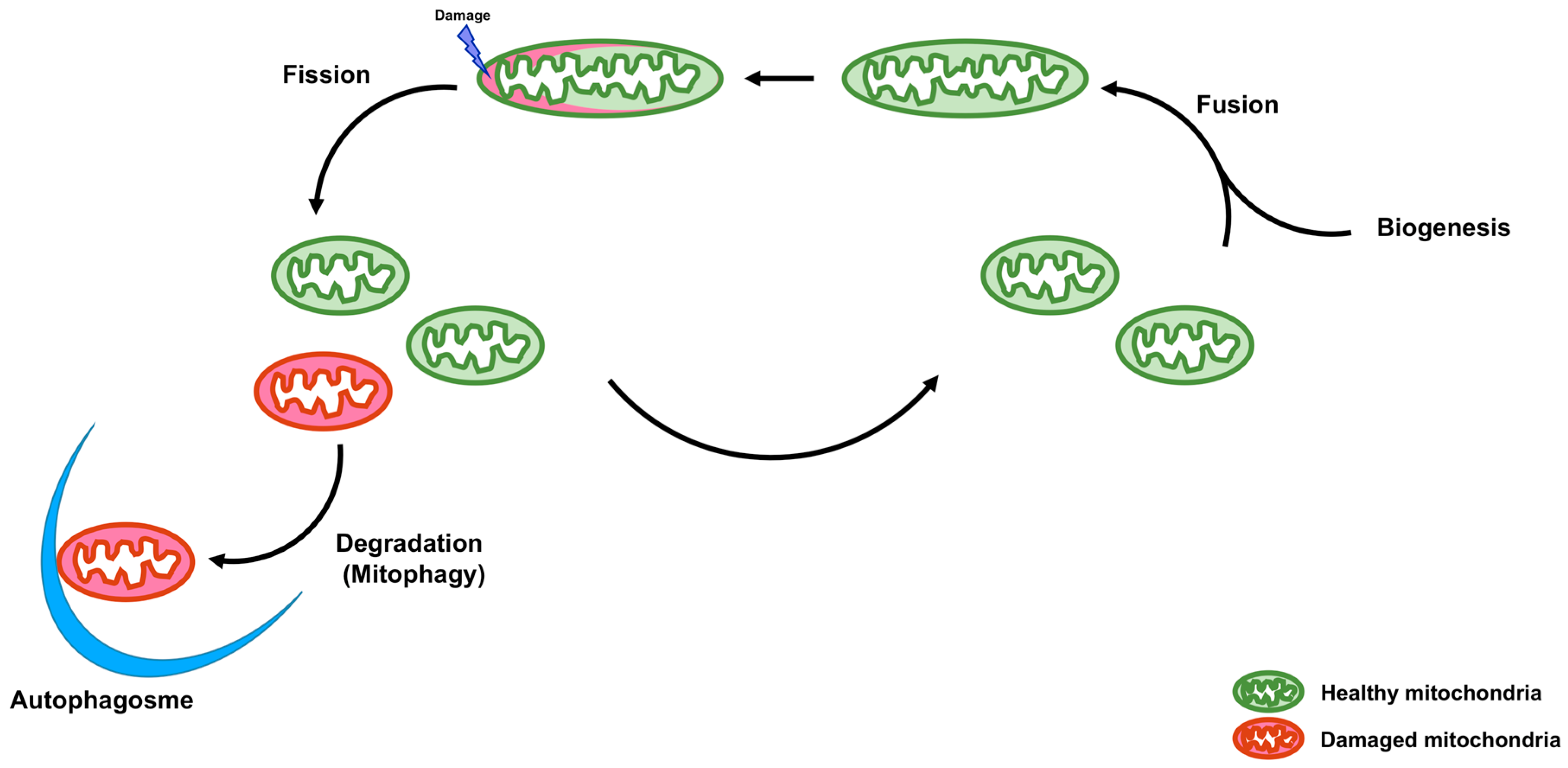
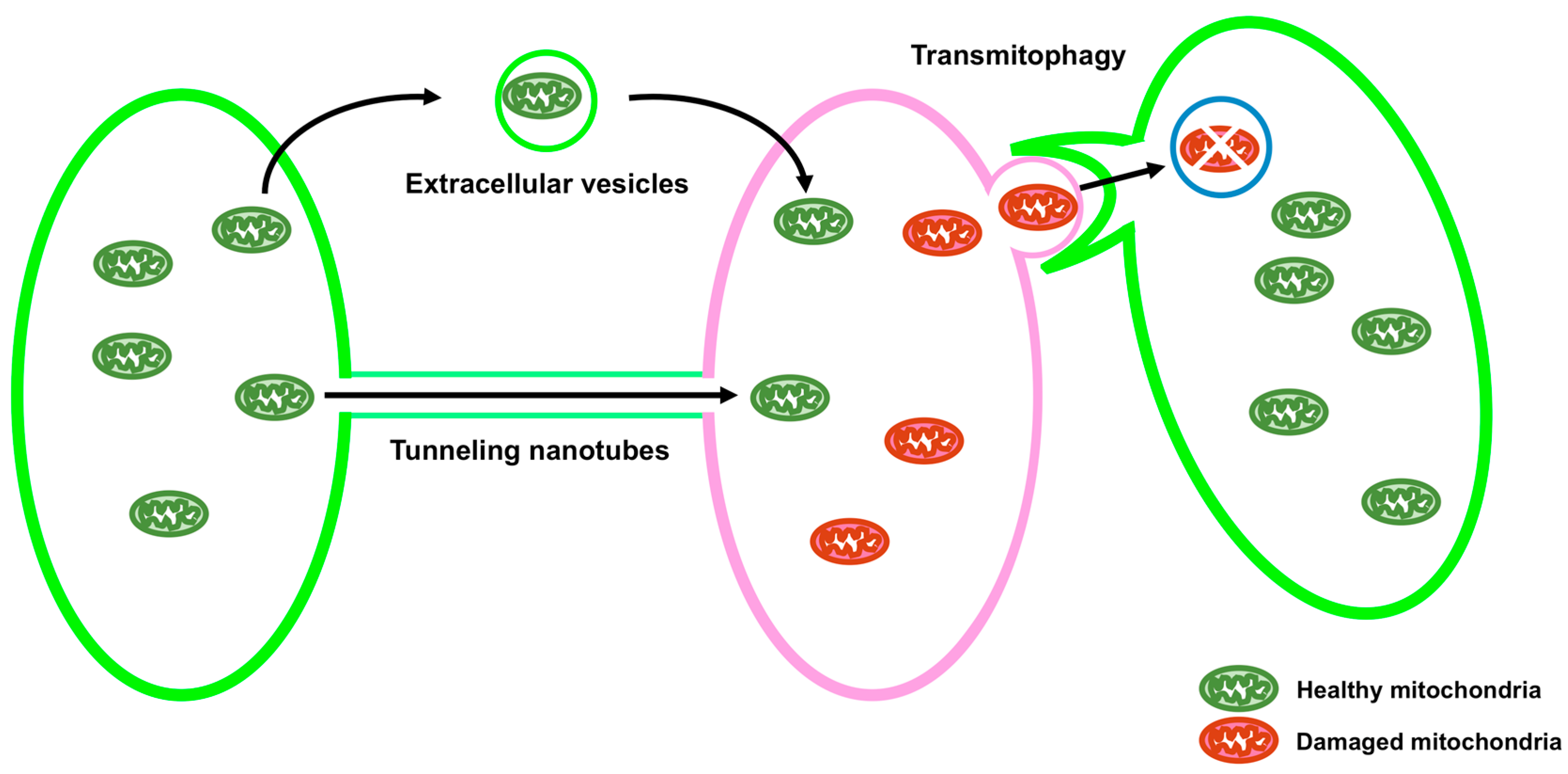
Mitochondria are double membrane organelles and are referred to as powerhouses of the cell because their major function is the generation of cellular ATP. Mitochondria also play important roles in other processes, including calcium homeostasis, reactive oxygen species production, apoptosis, autophagy, and the metabolism of amino acids, lipids, and glucose [1]. Loss of mitochondrial function is associated with various types of diseases [2]. Quality control of mitochondria is, therefore, crucial. A functional population of mitochondria is strictly controlled by intra- and intercellular quality control mechanisms. Intracellular quality control consists of biogenesis, fusion and fission, and degradation of mitochondria (Figure 1). On the other hand, intercellular quality control consists of tunneling nanotubes (TNTs) and extracellular vesicles (EVs) (Figure 2).
2. Intracellular Quality Control
2.1. Biogenesis
Mitochondria do not originate de novo; rather, proteins involved in the maintenance of mitochondrial population and mass regulate the biogenesis of mitochondria. These proteins are encoded by both mitochondrial DNA (mtDNA) and nuclear DNA (nDNA). Mitochondrial transcription factor A (TFAM), transcription factor B2, mitochondrial (TFB2M), nuclear respiratory factor 1 (NRF1) and NRF2, estrogen-related receptors (ERRs), and peroxisome proliferator-activated receptor gamma coactivator 1α (PGC-1α) play important roles in activating the transcription of genes required for mitochondrial biogenesis.
TFAM is encoded by nDNA and transported from the cytosol to mitochondria. In mitochondria, TFAM plays dual roles: as a transcription factor (TF) for mitochondrial genes and as a regulator of mtDNA replication [3,4,5]. TFB2M is also encoded by nDNA and transported from the cytosol to mitochondria to function as TF for mitochondrial genes. By contrast, transcription factor B1, mitochondrial (TFB1M), a paralog of TFB2M, is a dimethyltransferase that methylates adenine residues of mt12S rRNA [6]. TFAM binds to mtDNA and changes its structure. TFB2M and mitochondrial RNA polymerase (POLRMT) interact with TFAM to induce target gene expression. Mitochondrial transcription machinery, composed of TFAM, TFB2M, and POLRMT, initiates the expression of mtDNA [7,8,9]. A recent study has shown that POLRMT also functions as a switch between the transcription and replication of mtDNA [10].
NRF1 and NRF2 are nuclear TFs. NRF1 binds to GC-rich palindromes [11,12], whereas NRF2 binds to direct tandem repeats with GGAA core motifs [13,14]. Human NRF2 consists of two subunits: α and β (β1 or β2) or γ (γ1 or γ2). Subunit α contains the DNA-binding domain, whereas subunit β or γ contains the transcription activation domain [15]. Both NRF1 and NRF2 positively regulate the expression of genes encoding proteins related to the oxidative phosphorylation system (OXPHOS) complexes, heme biosynthesis, mitochondrial protein import and assembly, and mitochondrial translation [16]. NRF1 and NRF2 also regulate the expression of TFAM [17] and TFB2M [18].
ERRs α, β, and γ are nuclear receptors (NRs) whose endogenous ligands are unknown [19]. ERRs bind to the ERR response element with a consensus DNA sequence of TCAAGGTCA. ERRα and ERRγ bind to promoters of genes encoding mitochondrial proteins. However, the role of ERRβ in the expression of mitochondrial genes is unknown. The transcriptional function of ERRs is positively regulated by PGC-1α [20,21].
PGC-1α is a coactivator that lacks DNA-binding activity, but activates transcription of TFs or NRs by binding to these proteins. PGC-1α interacts with NRF1, NRF2, and ERRs to positively regulate these TFs and NRs for mitochondrial biogenesis [22]. Although PGC-1α is not a coactivator of TFAM and TFB2M, it indirectly activates the expression of TFAM and TFB2M via the activation of NRFs [17,18]. Thus, PGC-1α is considered a master regulator of mitochondrial biogenesis. PGC-1α is a member of the PGC1 family, which also includes PGC-1β and PGC-related coactivator (PRC). PGC-1β associates with NRF1 and ERRs, and positively regulates the expression of mitochondrial biogenesis proteins [22]. PRC also binds to NRF1 and ERRα. In addition, PRC forms a complex with NRF2 by binding to the host cell factor 1 (HCF-1) [23]. Post-translational modifications of PGC-1α include phosphorylation, methylation, acetylation, and deacetylation, whereas those of PGC-1β and PRC remain unclear [24].
2.2. Fusion and Fission
2.2.1. Fusion of Mitochondria
Mitochondrial fusion results in the exchange of mtDNA, proteins, and metabolites from healthy and damaged mitochondria, in order to repress the accumulation of damaged contents in a single mitochondrion. The fusion of the outer mitochondrial membrane (OMM) is controlled by mitofusin (MFN), whereas that of the inner mitochondrial membrane (IMM) is regulated by optic atrophy 1 (OPA1).
Two isoforms of MFN, MFN1 and MFN2, exist in mammals. Both MFNs are transmembrane GTPases, located in the OMM. MFNs play a crucial role in the tethering and fusion of the OMMs [25]. However, the GTPase and tethering activities of MFN1 are greater than that those of MFN2 [26]. Thus, MFN1 is considered the main GTP-dependent membrane tethering protein for mitochondrial fusion. MFN2 is expressed on the mitochondria-associated endoplasmic reticulum (ER) membrane. It tethers mitochondria to the ER by binding to MFNs on the OMM and functions in the uptake of mitochondrial Ca2+ from the ER [27].
OPA1 is a GTPase and transmembrane protein localized to the IMM. The long form of OPA1 (L-OPA1) is processed to its short form (S-OPA1) by OMA1 and YME1L1 [28,29]. The transmembrane domain of L-OPA1 is anchored to the IMM. By contrast, S-OPA1 is cleaved and does not possess a transmembrane domain. Knockdown of OPA1 results in mitochondrial fragmentation, which is recovered by the induction of L-OPA1 [29]. In addition, a decrease in the mitochondrial membrane potential induces complete conversion of L-OPA1 to S-OPA1 by OMA1, leading to the inhibition of mitochondrial fusion and induction of mitochondrial fragmentation [30]. Thus, L-OPA1 functions to regulate the fusion of the IMM.
2.2.2. Fission of Mitochondria
It is known that mitochondrial fission is necessary for the transmission of mitochondria to daughter cells during mitosis and the dissociation of damaged DNA, proteins, and metabolites of mitochondria. Mitochondrial fission is mainly regulated by dynamin-related protein 1 (DRP1). DRP1 is a cytosolic dynamin-like GTPase. It is recruited to the OMM to form multimeric ring-like structures at mitochondrial fission sites, which leads to the constriction and scission of mitochondria in a GTPase dependent manner. The activity of DRP1 is regulated by post-translational modifications, such as phosphorylation, ubiquitination, sumorylation, S-nitrosylation, and O-GlcNAcylation [31].
The recruitment of DRP1 onto the OMM is mediated by OMM-localized DRP1 receptors, namely mitochondrial fission factor (MFF), fission mitochondrial 1 (FIS1), mitochondrial elongation factor 1 [MIEF1; also known as the mitochondrial dynamic 51 kDa protein (MiD51)], and MIEF2 (also known as MiD49). MFF is localized to the OMM. Overexpression of MFF leads to the recruitment of DRP1 to the OMM, whereas knockdown of MFF results in the elongation of mitochondria [32,33]. FIS1 is also an anchored membrane protein and an ortholog of yeast Fis1. Yeast Fis1 acts as a receptor for Dnm1, the yeast ortholog of DRP1, and functions in the fission of yeast mitochondria [34]. Overexpression of FIS1 enhances the activity of mitochondrial fission, whereas blocking of FIS1 results in the elongation of mitochondria [35,36,37,38]. MIEF1 and MIEF2 are anchored to the OMM. Overexpression of the MIEF proteins results in the elongation of mitochondria, due to blocking of mitochondrial fission [35,39,40]. However, depletion of MIEFs also results in the elongation of mitochondria [35,39,41]. Recently, MIEFs and MFF have been shown to coordinately function with DRP1 on the OMM and have been used as models to demonstrate the contradictory results obtained from the overexpression and knockout of MIEFs. Although both MIEFs and MFF bind to DRP1, MIEFs have higher binding affinities to DRP1 than MFF. Thus, MIEF overexpression inhibits the binding of endogenous MFF to DRP1, leading to mitochondrial fusion. MIEFs also bind to MFF, thereby linking DRP1 to MFF and forming a DRP1-MIEF-MFF complex, resulting in mitochondrial fission. MIEF knockout inhibits the formation of this complex, thus leading to mitochondrial fusion [42].
2.3. Degradation (Mitophagy)
Damaged and dysfunctional mitochondria are deleterious to the cell. Degradation of such mitochondria is, therefore, crucial. Mitochondrial degradation is executed via the process of autophagy, which removes unwanted cytosolic components [43,44,45]. Selective degradation of mitochondria via autophagy is called mitophagy. Several proteins mediate the process of mitophagy, including phosphatase and tensin homolog (PTEN), induced putative kinase 1 (PINK1), Parkin, BCL2 interacting protein 3 (BNIP3), NIX [also known as BNIP3 like (BNIP3L)], Bcl2-like protein 13 (Bcl2-L-13), and FUN14 domain containing 1 (FUNDC1).
PINK1 is a serine/threonine kinase localized to mitochondria [46]. In healthy mitochondria, presenilin-associated rhomboid-like (PARL) processes PINK1, leading to the degradation of PINK1. In depolarized mitochondria, the processing of PINK1 by PARL is blocked. This results in the accumulation of PINK1 on the OMM [47,48,49]. PINK1 undergoes autophosphorylation and phosphorylates ubiquitin moieties of originally ubiquitinated OMM proteins. In addition, PINK1 phosphorylates and activates Parkin, an E3 ubiquitin ligase that adds ubiquitin molecules to originally ubiquitinated OMM proteins [50,51,52]. PINK1 also phosphorylates the ubiquitin molecules added by Parkin. The ubiquitin-binding autophagic adaptor proteins, nuclear dot protein 52 kDa (NDP52) and optineurin (OPTN), recruit microtubule-associated protein 1 light chain 3 (LC3) to the OMM proteins ubiquitinated and phosphorylated by PINK1 and Parkin, leading to mitophagy [53,54,55].
BNIP3, NIX, Bcl2-L-13, and FUNDC1 are localized to the OMM and mediate mitophagy by associating with the LC3 subfamily proteins, including LC3 alpha (LC3A), LC3 beta (LC3B), and LC3C, and the γ-aminobutyric-acid-type-A receptor-associated protein (GABARAP) subfamily proteins, including GABARAP, GABARAP-like 1 (GABARAPL1), and GABARAP-like 2 (GABARAPL2) [56]. BNIP3, NIX, and Bcl2-L-13 belong to the Bcl-2 family. Although Bcl-2 harbors four Bcl-2 homology motifs (BH1–4), BNIP3 harbors only the BH3 motif [57]. Overexpression of BNIP3 leads to the induction of mitophagy [58,59], whereas BNIP3 knockdown suppresses mitophagy [60]. Phosphorylation of Ser17 of BNIP3 is necessary for the binding of BNIP3 to LC3B, whereas phosphorylation of both Ser17 and Ser24 mediates the binding of BNIP3 to GABARAPL2 (also known as GATE-16) [61]. Like BNIP3, NIX also harbors only the BH3 motif. Deletion of NIX results in defective mitophagy [62,63]. Although NIX associates with all members of the LC3 and GABARAP subfamilies [56], phosphorylation of Ser34 and Ser35 residues of NIX enhances its ability to bind to LC3A and LC3B [64]. Bcl2-L-13 is a Bcl-2 homolog protein that also plays key roles in mitophagy and mitochondrial fragmentation. Phosphorylation at Ser272 of Bcl2-L-13 is necessary for its interaction with LC3B [65]. FUNDC1, which is also located in the OMM, functions to link mitochondria with LC3. Casein kinase 2 (CK2) and Src kinase phosphorylate Ser13 and Tyr18 residues of FUNDC1, respectively, blocking the induction of mitophagy. By contrast, dephosphorylation of Ser13 and Tyr18 residues of FUNDC1 by PGAM5, a mitochondrial phosphatase, or inhibition of CK2 and Src kinase results in the induction of mitophagy [66,67].
Mitochondrial-derived vesicles (MDVs) and mitochondrial spheroids are also important pathways for mitochondrial degradation and quality control, in addition to canonical autophagy [68]. MDVs deliver oxidized components of the mitochondria to the lysosome in response to oxidative stress. MDV formation is dependent on PINK1 and Parkin. However, key proteins controlling canonical autophagy, such as ATG5 and LC3, are not necessary for MDV formation. In addition, MDV formation and turnover precede mitophagy [69,70]. A recent study has shown that Syntaxin-17 is required for fusing the MDV and the lysosome [71]. Mitochondrial spheroids are the transformed structures of the mitochondria. They have a ring or cup-like shape and are generated in response to oxidative stress and mitochondrial damage [72,73,74]. As mitochondrial spheroids are formed independently of ATG5 or ATG7, their formation is distinct from canonical autophagy. Although mitochondrial spheroids contain lysosomal proteins, the degradation of contents via mitochondrial spheroids remains to be elucidated. MFN1 and MFN2 are required for mitochondrial spheroid formation, and Parkin inhibits mitochondrial spheroid formation by degrading MFN1 and MFN2. Thus, Parkin induces mitophagy and prevents mitochondrial spheroid formation [68,72,74].
3. Intercellular Transport
Mitochondria are transported between cells via TNTs and EVs (Figure 2). It has been reported that intercellular transport of mitochondria via TNTs rescues cells containing damaged mitochondria by sending healthy mitochondria from adjacent cells [75,76,77,78,79,80]. The transport of mitochondria via TNTs can be bidirectional [81]. It has been shown that mitochondrial rho GTPase 1 (MIRO1), also known as ras homolog family member T1 (RHOT1), plays an important role in the intercellular transport of mitochondria through TNTs [82]. MIRO1 is anchored to the OMM and is also related to the intracellular transport of mitochondria in the cell [83]. Knockdown of MIRO1 inhibits mitochondrial transport from mesenchymal stem cells (MSCs) to epithelial cells injured by rotenone, without reducing TNT formation, whereas overexpression of MIRO1 leads to an increase in the transport of mitochondria from MSCs to rotenone-injured epithelial cells [82].
EVs are divided into two types: exosomes (30–150 nm) and microvesicles (MVs) (30–1000 nm) [84]. The intercellular transport of mitochondria via EVs has been demonstrated [80,85,86]. It has been recently shown that the release of mitochondria-containing EVs from the astrocyte, mediated by CD38-cyclic ADP ribose (cADPR)-Ca2+ signaling, saves neurons damaged by oxygen-glucose deprivation or ischemic stroke [87]. CD38 is an ADP ribosyl cyclase that generates cADPR, leading to the release of Ca2+ ions [88]. Activation of CD38 and addition of cADPR enhances the release of mitochondria from astrocytes, whereas knockdown of CD38 or BAPTA-AM, an intracellular Ca2 chelator, reduced the release of mitochondria from astrocytes [87]. Damaged mitochondria are also transported into adjacent cells for their degradation. The retinal ganglion cells produce mitochondria-containing protrusions surrounded by vesicles. The adjacent astrocyte tears off, internalizes these protrusions, and degrades the mitochondria in these protrusions. This process is termed transmitophagy [89].
4. Concluding Remarks
Although the major function of mitochondria is the generation of energy, mitochondria are also associated with cellular homeostasis, cell signaling, metabolism, and cell death. Thus, intra- and intercellular quality controls of mitochondria are crucial and rigorously regulated. The impairment of quality control results in the accumulation of damaged mitochondria, leading to cell death and various types of diseases. Modifications of molecules and/or signaling associated with intra- and intercellular quality controls of mitochondria helps rescue cells containing damaged mitochondria. Moreover, intercellular quality control of mitochondria via EVs can lead to the possibility of the delivering of healthy mitochondria into cells containing damaged mitochondria. The elucidation of detailed mechanisms of intra- and intercellular quality controls of mitochondria will help in the development of therapeutic strategies for the management of diseases caused by defective or dysfunctional mitochondria.
Author Contributions
Y.K. and H.N. contributed to the writing of the review.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Vakifahmetoglu-Norberg, H.; Ouchida, A.T.; Norberg, E. The role of mitochondria in metabolism and cell death. Biochem. Biophys. Res. Commun. 2017, 482, 426–431. [Google Scholar] [CrossRef] [PubMed]
- Chan, D.C. Mitochondria: Dynamic organelles in disease, aging, and development. Cell 2006, 125, 1241–1252. [Google Scholar] [CrossRef] [PubMed]
- Ekstrand, M.I.; Falkenberg, M.; Rantanen, A.; Park, C.B.; Gaspari, M.; Hultenby, K.; Rustin, P.; Gustafsson, C.M.; Larsson, N.G. Mitochondrial transcription factor A regulates mtDNA copy number in mammals. Hum. Mol. Genet. 2004, 13, 935–944. [Google Scholar] [CrossRef] [PubMed]
- Ngo, H.B.; Lovely, G.A.; Phillips, R.; Chan, D.C. Distinct structural features of TFAM drive mitochondrial DNA packaging versus transcriptional activation. Nat. Commun. 2014, 5, 3077. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Dierckx, A.; Wanrooij, P.H.; Wanrooij, S.; Larsson, N.G.; Wilhelmsson, L.M.; Falkenberg, M.; Gustafsson, C.M. Mammalian transcription factor A is a core component of the mitochondrial transcription machinery. Proc. Natl. Acad. Sci. USA 2012, 109, 16510–16515. [Google Scholar] [CrossRef] [PubMed]
- Metodiev, M.D.; Lesko, N.; Park, C.B.; Camara, Y.; Shi, Y.; Wibom, R.; Hultenby, K.; Gustafsson, C.M.; Larsson, N.G. Methylation of 12S rRNA is necessary for in vivo stability of the small subunit of the mammalian mitochondrial ribosome. Cell Metab. 2009, 9, 386–397. [Google Scholar] [CrossRef] [PubMed]
- Morozov, Y.I.; Agaronyan, K.; Cheung, A.C.; Anikin, M.; Cramer, P.; Temiakov, D. A novel intermediate in transcription initiation by human mitochondrial RNA polymerase. Nucleic Acids Res. 2014, 42, 3884–3893. [Google Scholar] [CrossRef] [PubMed]
- Posse, V.; Gustafsson, C.M. Human Mitochondrial Transcription Factor B2 Is Required for Promoter Melting during Initiation of Transcription. J. Biol. Chem. 2017, 292, 2637–2645. [Google Scholar] [CrossRef] [PubMed]
- Ramachandran, A.; Basu, U.; Sultana, S.; Nandakumar, D.; Patel, S.S. Human mitochondrial transcription factors TFAM and TFB2M work synergistically in promoter melting during transcription initiation. Nucleic Acids Res. 2017, 45, 861–874. [Google Scholar] [CrossRef] [PubMed]
- Kuhl, I.; Miranda, M.; Posse, V.; Milenkovic, D.; Mourier, A.; Siira, S.J.; Bonekamp, N.A.; Neumann, U.; Filipovska, A.; Polosa, P.L.; et al. POLRMT regulates the switch between replication primer formation and gene expression of mammalian mtDNA. Sci. Adv. 2016, 2, e1600963. [Google Scholar] [CrossRef] [PubMed]
- Evans, M.J.; Scarpulla, R.C. NRF-1: A trans-activator of nuclear-encoded respiratory genes in animal cells. Genes Dev. 1990, 4, 1023–1034. [Google Scholar] [CrossRef] [PubMed]
- Virbasius, C.A.; Virbasius, J.V.; Scarpulla, R.C. NRF-1, an activator involved in nuclear-mitochondrial interactions, utilizes a new DNA-binding domain conserved in a family of developmental regulators. Genes Dev. 1993, 7, 2431–2445. [Google Scholar] [CrossRef] [PubMed]
- Virbasius, J.V.; Scarpulla, R.C. Transcriptional activation through ETS domain binding sites in the cytochrome c oxidase subunit IV gene. Mol. Cell. Biol. 1991, 11, 5631–5638. [Google Scholar] [CrossRef] [PubMed]
- Virbasius, J.V.; Virbasius, C.A.; Scarpulla, R.C. Identity of GABP with NRF-2, a multisubunit activator of cytochrome oxidase expression, reveals a cellular role for an ETS domain activator of viral promoters. Genes Dev. 1993, 7, 380–392. [Google Scholar] [CrossRef] [PubMed]
- Gugneja, S.; Virbasius, J.V.; Scarpulla, R.C. Four structurally distinct, non-DNA-binding subunits of human nuclear respiratory factor 2 share a conserved transcriptional activation domain. Mol. Cell. Biol. 1995, 15, 102–111. [Google Scholar] [CrossRef] [PubMed]
- Kelly, D.P.; Scarpulla, R.C. Transcriptional regulatory circuits controlling mitochondrial biogenesis and function. Genes Dev. 2004, 18, 357–368. [Google Scholar] [CrossRef] [PubMed]
- Virbasius, J.V.; Scarpulla, R.C. Activation of the human mitochondrial transcription factor A gene by nuclear respiratory factors: A potential regulatory link between nuclear and mitochondrial gene expression in organelle biogenesis. Proc. Natl. Acad. Sci. USA 1994, 91, 1309–1313. [Google Scholar] [CrossRef] [PubMed]
- Gleyzer, N.; Vercauteren, K.; Scarpulla, R.C. Control of mitochondrial transcription specificity factors (TFB1M and TFB2M) by nuclear respiratory factors (NRF-1 and NRF-2) and PGC-1 family coactivators. Mol. Cell. Biol. 2005, 25, 1354–1366. [Google Scholar] [CrossRef] [PubMed]
- Gronemeyer, H.; Gustafsson, J.A.; Laudet, V. Principles for modulation of the nuclear receptor superfamily. Nat. Rev. Drug Discov. 2004, 3, 950–964. [Google Scholar] [CrossRef] [PubMed]
- Fan, W.; Evans, R. PPARs and ERRs: Molecular mediators of mitochondrial metabolism. Curr. Opin. Cell Biol. 2015, 33, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Huss, J.M.; Garbacz, W.G.; Xie, W. Constitutive activities of estrogen-related receptors: Transcriptional regulation of metabolism by the ERR pathways in health and disease. Biochim. Biophys. Acta 2015, 1852, 1912–1927. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Handschin, C.; Spiegelman, B.M. Metabolic control through the PGC-1 family of transcription coactivators. Cell Metab. 2005, 1, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Vercauteren, K.; Gleyzer, N.; Scarpulla, R.C. PGC-1-related coactivator complexes with HCF-1 and NRF-2beta in mediating NRF-2(GABP)-dependent respiratory gene expression. J. Biol. Chem. 2008, 283, 12102–12111. [Google Scholar] [CrossRef] [PubMed]
- Villena, J.A. New insights into PGC-1 coactivators: Redefining their role in the regulation of mitochondrial function and beyond. FEBS J. 2015, 282, 647–672. [Google Scholar] [CrossRef] [PubMed]
- Koshiba, T.; Detmer, S.A.; Kaiser, J.T.; Chen, H.; McCaffery, J.M.; Chan, D.C. Structural basis of mitochondrial tethering by mitofusin complexes. Science 2004, 305, 858–862. [Google Scholar] [CrossRef] [PubMed]
- Ishihara, N.; Eura, Y.; Mihara, K. Mitofusin 1 and 2 play distinct roles in mitochondrial fusion reactions via GTPase activity. J. Cell Sci. 2004, 117, 6535–6546. [Google Scholar] [CrossRef] [PubMed]
- De Brito, O.M.; Scorrano, L. Mitofusin 2 tethers endoplasmic reticulum to mitochondria. Nature 2008, 456, 605–610. [Google Scholar] [CrossRef] [PubMed]
- Ehses, S.; Raschke, I.; Mancuso, G.; Bernacchia, A.; Geimer, S.; Tondera, D.; Martinou, J.C.; Westermann, B.; Rugarli, E.I.; Langer, T. Regulation of OPA1 processing and mitochondrial fusion by m-AAA protease isoenzymes and OMA1. J. Cell Biol. 2009, 187, 1023–1036. [Google Scholar] [CrossRef] [PubMed]
- Ishihara, N.; Fujita, Y.; Oka, T.; Mihara, K. Regulation of mitochondrial morphology through proteolytic cleavage of OPA1. EMBO J. 2006, 25, 2966–2977. [Google Scholar] [CrossRef] [PubMed]
- Anand, R.; Wai, T.; Baker, M.J.; Kladt, N.; Schauss, A.C.; Rugarli, E.; Langer, T. The i-AAA protease YME1L and OMA1 cleave OPA1 to balance mitochondrial fusion and fission. J. Cell Biol. 2014, 204, 919–929. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Huang, Y.; Li, L. Drp1-Dependent Mitochondrial Fission Plays Critical Roles in Physiological and Pathological Progresses in Mammals. Int. J. Mol. Sci. 2017, 18, 144. [Google Scholar] [CrossRef] [PubMed]
- Gandre-Babbe, S.; van der Bliek, A.M. The novel tail-anchored membrane protein Mff controls mitochondrial and peroxisomal fission in mammalian cells. Mol. Biol. Cell 2008, 19, 2402–2412. [Google Scholar] [CrossRef] [PubMed]
- Otera, H.; Wang, C.; Cleland, M.M.; Setoguchi, K.; Yokota, S.; Youle, R.J.; Mihara, K. Mff is an essential factor for mitochondrial recruitment of Drp1 during mitochondrial fission in mammalian cells. J. Cell Biol. 2010, 191, 1141–1158. [Google Scholar] [CrossRef] [PubMed]
- Hettema, E.H.; Motley, A.M. How peroxisomes multiply. J. Cell Sci. 2009, 122, 2331–2336. [Google Scholar] [CrossRef] [PubMed]
- Loson, O.C.; Song, Z.; Chen, H.; Chan, D.C. Fis1, Mff, MiD49, and MiD51 mediate Drp1 recruitment in mitochondrial fission. Mol. Biol. Cell 2013, 24, 659–667. [Google Scholar] [CrossRef] [PubMed]
- Stojanovski, D.; Koutsopoulos, O.S.; Okamoto, K.; Ryan, M.T. Levels of human Fis1 at the mitochondrial outer membrane regulate mitochondrial morphology. J. Cell Sci. 2004, 117, 1201–1210. [Google Scholar] [CrossRef] [PubMed]
- Yoon, Y.; Krueger, E.W.; Oswald, B.J.; McNiven, M.A. The mitochondrial protein hFis1 regulates mitochondrial fission in mammalian cells through an interaction with the dynamin-like protein DLP1. Mol. Cell. Biol. 2003, 23, 5409–5420. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.; Fox, R.J.; Burwell, L.S.; Yoon, Y. Regulation of mitochondrial fission and apoptosis by the mitochondrial outer membrane protein hFis1. J. Cell Sci. 2005, 118, 4141–4151. [Google Scholar] [CrossRef] [PubMed]
- Palmer, C.S.; Osellame, L.D.; Laine, D.; Koutsopoulos, O.S.; Frazier, A.E.; Ryan, M.T. MiD49 and MiD51, new components of the mitochondrial fission machinery. EMBO Rep. 2011, 12, 565–573. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Liu, T.; Jin, S.; Wang, X.; Qu, M.; Uhlen, P.; Tomilin, N.; Shupliakov, O.; Lendahl, U.; Nister, M. Human MIEF1 recruits Drp1 to mitochondrial outer membranes and promotes mitochondrial fusion rather than fission. EMBO J. 2011, 30, 2762–2778. [Google Scholar] [CrossRef] [PubMed]
- Otera, H.; Miyata, N.; Kuge, O.; Mihara, K. Drp1-dependent mitochondrial fission via MiD49/51 is essential for apoptotic cristae remodeling. J. Cell Biol. 2016, 212, 531–544. [Google Scholar] [CrossRef] [PubMed]
- Yu, R.; Liu, T.; Jin, S.B.; Ning, C.; Lendahl, U.; Nister, M.; Zhao, J. MIEF1/2 function as adaptors to recruit Drp1 to mitochondria and regulate the association of Drp1 with Mff. Sci. Rep. 2017, 7, 880. [Google Scholar] [CrossRef] [PubMed]
- Ariosa, A.R.; Klionsky, D.J. Autophagy core machinery: Overcoming spatial barriers in neurons. J. Mol. Med. 2016, 94, 1217–1227. [Google Scholar] [CrossRef] [PubMed]
- Kiriyama, Y.; Nochi, H. The Function of Autophagy in Neurodegenerative Diseases. Int. J. Mol. Sci. 2015, 16, 26797–26812. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Klionsky, D.J. Eaten alive: A history of macroautophagy. Nat. Cell Biol. 2010, 12, 814–822. [Google Scholar] [CrossRef] [PubMed]
- Valente, E.M.; Abou-Sleiman, P.M.; Caputo, V.; Muqit, M.M.; Harvey, K.; Gispert, S.; Ali, Z.; Del Turco, D.; Bentivoglio, A.R.; Healy, D.G.; et al. Hereditary early-onset Parkinson’s disease caused by mutations in PINK1. Science 2004, 304, 1158–1160. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.M.; Lazarou, M.; Wang, C.; Kane, L.A.; Narendra, D.P.; Youle, R.J. Mitochondrial membrane potential regulates PINK1 import and proteolytic destabilization by PARL. J. Cell Biol. 2010, 191, 933–942. [Google Scholar] [CrossRef] [PubMed]
- Yamano, K.; Youle, R.J. PINK1 is degraded through the N-end rule pathway. Autophagy 2013, 9, 1758–1769. [Google Scholar] [CrossRef] [PubMed]
- Meissner, C.; Lorenz, H.; Weihofen, A.; Selkoe, D.J.; Lemberg, M.K. The mitochondrial intramembrane protease PARL cleaves human Pink1 to regulate Pink1 trafficking. J. Neurochem. 2011, 117, 856–867. [Google Scholar] [CrossRef] [PubMed]
- Kazlauskaite, A.; Kondapalli, C.; Gourlay, R.; Campbell, D.G.; Ritorto, M.S.; Hofmann, K.; Alessi, D.R.; Knebel, A.; Trost, M.; Muqit, M.M. Parkin is activated by PINK1-dependent phosphorylation of ubiquitin at Ser65. Biochem. J. 2014, 460, 127–139. [Google Scholar] [CrossRef] [PubMed]
- Koyano, F.; Okatsu, K.; Kosako, H.; Tamura, Y.; Go, E.; Kimura, M.; Kimura, Y.; Tsuchiya, H.; Yoshihara, H.; Hirokawa, T.; et al. Ubiquitin is phosphorylated by PINK1 to activate parkin. Nature 2014, 510, 162–166. [Google Scholar] [CrossRef] [PubMed]
- Wauer, T.; Simicek, M.; Schubert, A.; Komander, D. Mechanism of phospho-ubiquitin-induced PARKIN activation. Nature 2015, 524, 370–374. [Google Scholar] [CrossRef] [PubMed]
- Fiesel, F.C.; Ando, M.; Hudec, R.; Hill, A.R.; Castanedes-Casey, M.; Caulfield, T.R.; Moussaud-Lamodière, E.L.; Stankowski, J.N.; Bauer, P.O.; Lorenzo-Betancor, O.; et al. (Patho-)physiological relevance of PINK1-dependent ubiquitin phosphorylation. EMBO Rep. 2015, 16, 1114–1130. [Google Scholar] [CrossRef] [PubMed]
- Heo, J.M.; Ordureau, A.; Paulo, J.A.; Rinehart, J.; Harper, J.W. The PINK1-PARKIN Mitochondrial Ubiquitylation Pathway Drives a Program of OPTN/NDP52 Recruitment and TBK1 Activation to Promote Mitophagy. Mol. Cell 2015, 60, 7–20. [Google Scholar] [CrossRef] [PubMed]
- Lazarou, M.; Sliter, D.A.; Kane, L.A.; Sarraf, S.A.; Wang, C.; Burman, J.L.; Sideris, D.P.; Fogel, A.I.; Youle, R.J. The ubiquitin kinase PINK1 recruits autophagy receptors to induce mitophagy. Nature 2015, 524, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Wild, P.; McEwan, D.G.; Dikic, I. The LC3 interactome at a glance. J. Cell Sci. 2014, 127, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Aouacheria, A.; Rech de Laval, V.; Combet, C.; Hardwick, J.M. Evolution of Bcl-2 homology motifs: Homology versus homoplasy. Trends Cell Biol. 2013, 23, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Hamacher-Brady, A.; Brady, N.R.; Logue, S.E.; Sayen, M.R.; Jinno, M.; Kirshenbaum, L.A.; Gottlieb, R.A.; Gustafsson, A.B. Response to myocardial ischemia/reperfusion injury involves Bnip3 and autophagy. Cell Death Differ. 2007, 14, 146–157. [Google Scholar] [CrossRef] [PubMed]
- Quinsay, M.N.; Thomas, R.L.; Lee, Y.; Gustafsson, A.B. Bnip3-mediated mitochondrial autophagy is independent of the mitochondrial permeability transition pore. Autophagy 2010, 6, 855–862. [Google Scholar] [CrossRef] [PubMed]
- Ishihara, M.; Urushido, M.; Hamada, K.; Matsumoto, T.; Shimamura, Y.; Ogata, K.; Inoue, K.; Taniguchi, Y.; Horino, T.; Fujieda, M.; et al. Sestrin-2 and BNIP3 regulate autophagy and mitophagy in renal tubular cells in acute kidney injury. Am. J. Physiol. Renal Physiol. 2013, 305, F495–F509. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Massen, S.; Terenzio, M.; Lang, V.; Chen-Lindner, S.; Eils, R.; Novak, I.; Dikic, I.; Hamacher-Brady, A.; Brady, N.R. Modulation of serines 17 and 24 in the LC3-interacting region of Bnip3 determines pro-survival mitophagy versus apoptosis. J. Biol. Chem. 2013, 288, 1099–1113. [Google Scholar] [CrossRef] [PubMed]
- Sandoval, H.; Thiagarajan, P.; Dasgupta, S.K.; Schumacher, A.; Prchal, J.T.; Chen, M.; Wang, J. Essential role for Nix in autophagic maturation of erythroid cells. Nature 2008, 454, 232–235. [Google Scholar] [CrossRef] [PubMed]
- Schweers, R.L.; Zhang, J.; Randall, M.S.; Loyd, M.R.; Li, W.; Dorsey, F.C.; Kundu, M.; Opferman, J.T.; Cleveland, J.L.; Miller, J.L.; et al. NIX is required for programmed mitochondrial clearance during reticulocyte maturation. Proc. Natl. Acad. Sci. USA 2007, 104, 19500–19505. [Google Scholar] [CrossRef] [PubMed]
- Rogov, V.V.; Suzuki, H.; Marinkovic, M.; Lang, V.; Kato, R.; Kawasaki, M.; Buljubasic, M.; Sprung, M.; Rogova, N.; Wakatsuki, S.; et al. Phosphorylation of the mitochondrial autophagy receptor Nix enhances its interaction with LC3 proteins. Sci. Rep. 2017, 7, 1131. [Google Scholar] [CrossRef] [PubMed]
- Murakawa, T.; Yamaguchi, O.; Hashimoto, A.; Hikoso, S.; Takeda, T.; Oka, T.; Yasui, H.; Ueda, H.; Akazawa, Y.; Nakayama, H.; et al. Bcl-2-like protein 13 is a mammalian Atg32 homologue that mediates mitophagy and mitochondrial fragmentation. Nat. Commun. 2015, 6, 7527. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Han, Z.; Feng, D.; Chen, Y.; Chen, L.; Wu, H.; Huang, L.; Zhou, C.; Cai, X.; Fu, C.; et al. A regulatory signaling loop comprising the PGAM5 phosphatase and CK2 controls receptor-mediated mitophagy. Mol. Cell 2014, 54, 362–377. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Feng, D.; Chen, G.; Chen, M.; Zheng, Q.; Song, P.; Ma, Q.; Zhu, C.; Wang, R.; Qi, W.; et al. Mitochondrial outer-membrane protein FUNDC1 mediates hypoxia-induced mitophagy in mammalian cells. Nat. Cell Biol. 2012, 14, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Ni, H.M.; Williams, J.A.; Ding, W.X. Mitochondrial dynamics and mitochondrial quality control. Redox Biol. 2015, 4, 6–13. [Google Scholar] [CrossRef] [PubMed]
- McLelland, G.L.; Soubannier, V.; Chen, C.X.; McBride, H.M.; Fon, E.A. Parkin and PINK1 function in a vesicular trafficking pathway regulating mitochondrial quality control. EMBO J. 2014, 33, 282–295. [Google Scholar] [CrossRef] [PubMed]
- Soubannier, V.; McLelland, G.L.; Zunino, R.; Braschi, E.; Rippstein, P.; Fon, E.A.; McBride, H.M. A vesicular transport pathway shuttles cargo from mitochondria to lysosomes. Curr. Biol. 2012, 22, 135–141. [Google Scholar] [CrossRef] [PubMed]
- McLelland, G.L.; Lee, S.A.; McBride, H.M.; Fon, E.A. Syntaxin-17 delivers PINK1/parkin-dependent mitochondrial vesicles to the endolysosomal system. J. Cell Biol. 2016, 214, 275–291. [Google Scholar] [CrossRef] [PubMed]
- Ding, W.X.; Guo, F.; Ni, H.M.; Bockus, A.; Manley, S.; Stolz, D.B.; Eskelinen, E.L.; Jaeschke, H.; Yin, X.M. Parkin and mitofusins reciprocally regulate mitophagy and mitochondrial spheroid formation. J. Biol. Chem. 2012, 287, 42379–42388. [Google Scholar] [CrossRef] [PubMed]
- Ding, W.X.; Li, M.; Biazik, J.M.; Morgan, D.G.; Guo, F.; Ni, H.M.; Goheen, M.; Eskelinen, E.L.; Yin, X.M. Electron microscopic analysis of a spherical mitochondrial structure. J. Biol. Chem. 2012, 287, 42373–42378. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.M.; Ding, W.X. The reciprocal roles of PARK2 and mitofusins in mitophagy and mitochondrial spheroid formation. Autophagy 2013, 9, 1687–1692. [Google Scholar] [CrossRef] [PubMed]
- Koyanagi, M.; Brandes, R.P.; Haendeler, J.; Zeiher, A.M.; Dimmeler, S. Cell-to-cell connection of endothelial progenitor cells with cardiac myocytes by nanotubes: A novel mechanism for cell fate changes? Circ. Res. 2005, 96, 1039–1041. [Google Scholar] [CrossRef] [PubMed]
- Onfelt, B.; Nedvetzki, S.; Benninger, R.K.; Purbhoo, M.A.; Sowinski, S.; Hume, A.N.; Seabra, M.C.; Neil, M.A.; French, P.M.; Davis, D.M. Structurally distinct membrane nanotubes between human macrophages support long-distance vesicular traffic or surfing of bacteria. J. Immunol. 2006, 177, 8476–8483. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Gerdes, H.H. Transfer of mitochondria via tunneling nanotubes rescues apoptotic PC12 cells. Cell Death Differ. 2015, 22, 1181–1191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saenz-de-Santa-Maria, I.; Bernardo-Castineira, C.; Enciso, E.; Garcia-Moreno, I.; Chiara, J.L.; Suarez, C.; Chiara, M.D. Control of long-distance cell-to-cell communication and autophagosome transfer in squamous cell carcinoma via tunneling nanotubes. Oncotarget 2017, 8, 20939–20960. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, K.; Park, H.C.; Ratliff, B.; Addabbo, F.; Hatzopoulos, A.K.; Chander, P.; Goligorsky, M.S. Adriamycin nephropathy: A failure of endothelial progenitor cell-induced repair. Am. J. Pathol. 2010, 176, 1685–1695. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.N.; Das, S.R.; Emin, M.T.; Wei, M.; Sun, L.; Westphalen, K.; Rowlands, D.J.; Quadri, S.K.; Bhattacharya, S.; Bhattacharya, J. Mitochondrial transfer from bone-marrow-derived stromal cells to pulmonary alveoli protects against acute lung injury. Nat. Med. 2012, 18, 759–765. [Google Scholar] [CrossRef] [PubMed]
- He, K.; Shi, X.; Zhang, X.; Dang, S.; Ma, X.; Liu, F.; Xu, M.; Lv, Z.; Han, D.; Fang, X.; et al. Long-distance intercellular connectivity between cardiomyocytes and cardiofibroblasts mediated by membrane nanotubes. Cardiovasc. Res. 2011, 92, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, T.; Mukherjee, S.; Pattnaik, B.; Kumar, M.; Singh, S.; Kumar, M.; Rehman, R.; Tiwari, B.K.; Jha, K.A.; Barhanpurkar, A.P.; et al. Miro1 regulates intercellular mitochondrial transport & enhances mesenchymal stem cell rescue efficacy. EMBO J. 2014, 33, 994–1010. [Google Scholar] [CrossRef] [PubMed]
- Reis, K.; Fransson, A.; Aspenstrom, P. The Miro GTPases: At the heart of the mitochondrial transport machinery. FEBS Lett. 2009, 583, 1391–1398. [Google Scholar] [CrossRef] [PubMed]
- Welsh, J.A.; Holloway, J.A.; Wilkinson, J.S.; Englyst, N.A. Extracellular Vesicle Flow Cytometry Analysis and Standardization. Front. Cell Dev. Biol. 2017, 5, 78. [Google Scholar] [CrossRef] [PubMed]
- Spees, J.L.; Olson, S.D.; Whitney, M.J.; Prockop, D.J. Mitochondrial transfer between cells can rescue aerobic respiration. Proc. Natl. Acad. Sci. USA 2006, 103, 1283–1288. [Google Scholar] [CrossRef] [PubMed]
- Phinney, D.G.; Di Giuseppe, M.; Njah, J.; Sala, E.; Shiva, S.; St Croix, C.M.; Stolz, D.B.; Watkins, S.C.; Di, Y.P.; Leikauf, G.D.; et al. Mesenchymal stem cells use extracellular vesicles to outsource mitophagy and shuttle microRNAs. Nat. Commun. 2015, 6, 8472. [Google Scholar] [CrossRef] [PubMed]
- Hayakawa, K.; Esposito, E.; Wang, X.; Terasaki, Y.; Liu, Y.; Xing, C.; Ji, X.; Lo, E.H. Transfer of mitochondria from astrocytes to neurons after stroke. Nature 2016, 535, 551–555. [Google Scholar] [CrossRef] [PubMed]
- Malavasi, F.; Deaglio, S.; Funaro, A.; Ferrero, E.; Horenstein, A.L.; Ortolan, E.; Vaisitti, T.; Aydin, S. Evolution and function of the ADP ribosyl cyclase/CD38 gene family in physiology and pathology. Physiol. Rev. 2008, 88, 841–886. [Google Scholar] [CrossRef] [PubMed]
- Davis, C.H.; Kim, K.Y.; Bushong, E.A.; Mills, E.A.; Boassa, D.; Shih, T.; Kinebuchi, M.; Phan, S.; Zhou, Y.; Bihlmeyer, N.A.; et al. Transcellular degradation of axonal mitochondria. Proc. Natl. Acad. Sci. USA 2014, 111, 9633–9638. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Schematic representation of intracellular quality control of mitochondria. Intracellular quality control of mitochondria consists of biogenesis, fusion, fission, and degradation (mitophagy) to maintain functions of mitochondria. Mitochondrial fission is necessary for the dissociation of damaged and dysfunctional mitochondria. Damaged and dysfunctional mitochondria are degraded by mitophagy. Mitochondrial biogenesis supplements decreased mitochondrial mass. Mitochondrial fusion leads to the exchange of mitochondrial DNA (mtDNA), proteins, and metabolites between healthy and damaged mitochondria to prevent the accumulation of damaged contents in a single mitochondrion.
Figure 1.
Schematic representation of intracellular quality control of mitochondria. Intracellular quality control of mitochondria consists of biogenesis, fusion, fission, and degradation (mitophagy) to maintain functions of mitochondria. Mitochondrial fission is necessary for the dissociation of damaged and dysfunctional mitochondria. Damaged and dysfunctional mitochondria are degraded by mitophagy. Mitochondrial biogenesis supplements decreased mitochondrial mass. Mitochondrial fusion leads to the exchange of mitochondrial DNA (mtDNA), proteins, and metabolites between healthy and damaged mitochondria to prevent the accumulation of damaged contents in a single mitochondrion.

Figure 2.
Schematic showing intercellular quality control of mitochondria. Mitochondria are transported between cells via tunneling nanotubes (TNTs) and extracellular vesicles (EVs), which help rescue cells containing damaged mitochondria by transporting healthy mitochondria from the adjacent cells. In neurons, damaged mitochondria packed in a neuron are degraded by an adjacent astrocyte (transmitophagy).
Figure 2.
Schematic showing intercellular quality control of mitochondria. Mitochondria are transported between cells via tunneling nanotubes (TNTs) and extracellular vesicles (EVs), which help rescue cells containing damaged mitochondria by transporting healthy mitochondria from the adjacent cells. In neurons, damaged mitochondria packed in a neuron are degraded by an adjacent astrocyte (transmitophagy).

© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kiriyama, Y.; Nochi, H. Intra- and Intercellular Quality Control Mechanisms of Mitochondria. Cells 2018, 7, 1. https://0-doi-org.brum.beds.ac.uk/10.3390/cells7010001
AMA Style
Kiriyama Y, Nochi H. Intra- and Intercellular Quality Control Mechanisms of Mitochondria. Cells. 2018; 7(1):1. https://0-doi-org.brum.beds.ac.uk/10.3390/cells7010001
Chicago/Turabian StyleKiriyama, Yoshimitsu, and Hiromi Nochi. 2018. "Intra- and Intercellular Quality Control Mechanisms of Mitochondria" Cells 7, no. 1: 1. https://0-doi-org.brum.beds.ac.uk/10.3390/cells7010001
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.