Natural Resistance Associated Macrophage Protein Is Involved in Immune Response of Blunt Snout Bream, Megalobrama amblycephala

Abstract
:1. Introduction
2. Materials and Methods
2.1. Fish, LPS Stimulation, and RNA Isolation
2.2. Cloning and Characterization of maNramp
2.3. Quantitative Analysis of maNramp
2.4. Statistical Analysis
3. Results
3.1. Identification and Characterization of maNramp
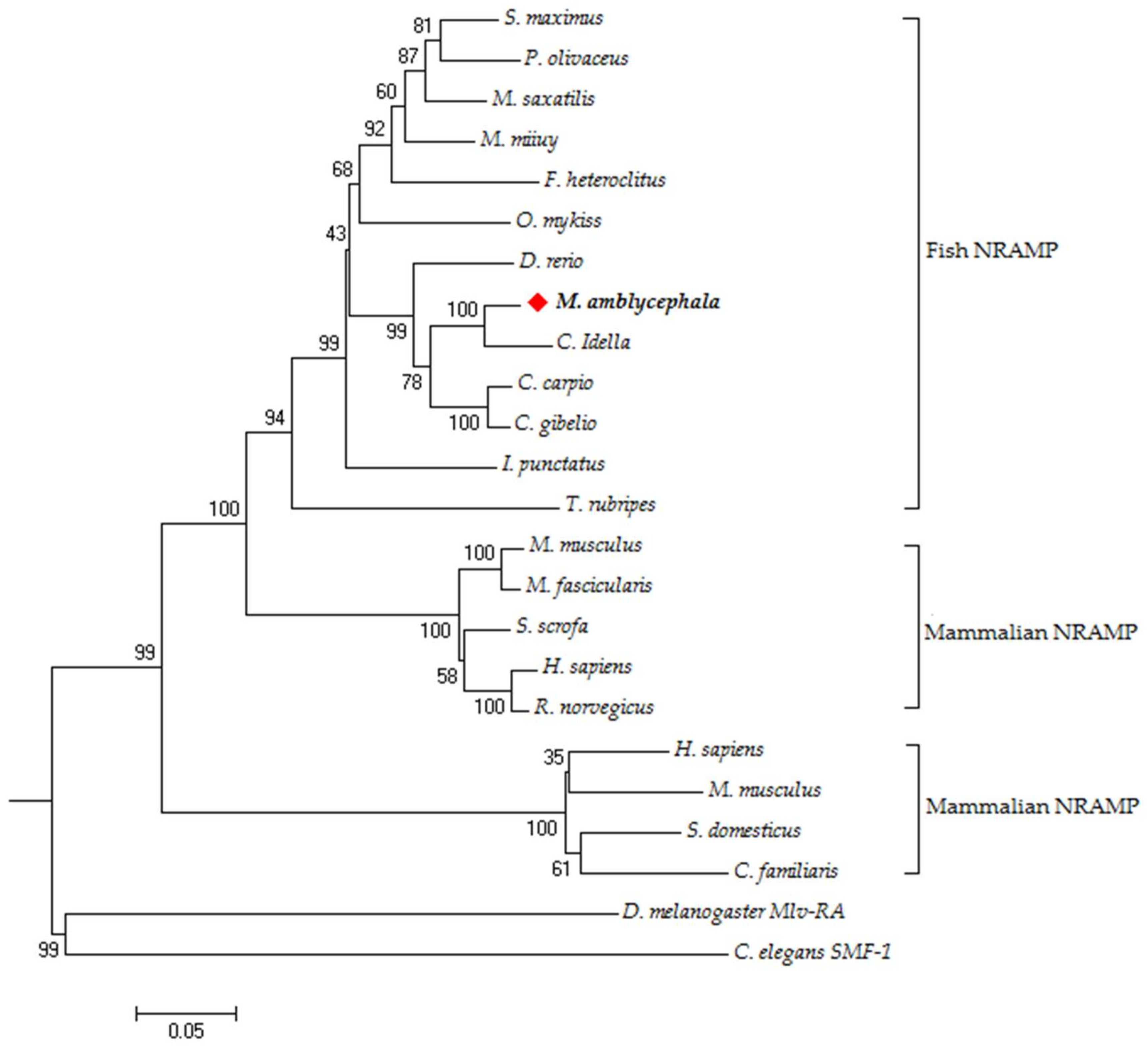
3.2. Phylogenetic Analysis
3.3. Tissue Distribution of maNramp
3.4. Temporal Expression Analysis of maNramp Post LPS Stimulation
4. Discussion and Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Tang, L.L.; Liang, Y.H.; Jiang, Y.H.; Liu, S.J.; Zhang, F.Y.; He, X.; Wang, T.Y.; Zhou, Y.; Zhong, H.; Yan, J.P. Identification and expression analysis on bactericidal permeability-increasing protein/lipopolysaccharide-binding protein of blunt snout bream, Megalobrama amblycephala. Fish Shellfish Immunol. 2015, 45, 630–640. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wei, X.L.; Chen, L.P.; Chen, N.; Li, Y.H.; Wang, W.M.; Wang, H.L. Sequence analysis and expression differentiation of chemokine receptor CXCR4b among three populations of Megalobrama amblycephala. Dev. Comp. Immunol. 2013, 40, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Luo, W.; Liu, H.; Zeng, C.; Liu, X.; Yi, S.; Wang, W. Transcriptome analysis and SSR/SNP markers information of the blunt snout bream (Megalobrama amblycephala). PLoS ONE 2012, 7, e42637. [Google Scholar] [CrossRef] [PubMed]
- Lv, Y.; Xiang, X.; Jiang, Y.; Tang, L.; Zhou, Y.; Zhong, H.; Xiao, J.; Yan, J. Identification and Characterization of Lipopolysaccharide Induced TNFalpha Factor from Blunt Snout Bream, Megalobrama amblycephala. Int. J. Mol. Sci. 2017, 18, 233. [Google Scholar] [CrossRef] [PubMed]
- Food and Agriculture Organization. China Fishery Statistical Year Book; Chinese Agriculture Press: Beijing, China, 2016. [Google Scholar]
- Nielsen, M.E.; Hoi, L.; Schmidt, A.S.; Qian, D.; Shimada, T.; Shen, J.Y.; Larsen, J.L. Is Aeromonas hydrophila the dominant motile Aeromonas species that causes disease outbreaks in aquaculture production in the Zhejiang Province of China? Dis. Aquat. Organ. 2001, 46, 23–29. [Google Scholar] [CrossRef] [PubMed]
- He, L.J.; Liao, L.K.; Yuan, J.F.; Tang, H.Y.; Wu, Q.; Zhang, G.W. Pathological observation of bacterial septicemia in Megalobrama amblycephala. J. Southwest Agric. Univ. 2006, 28, 483–485. [Google Scholar]
- Rauta, P.R.; Samanta, M.; Dash, H.R.; Nayak, B.; Das, S. Toll-like receptors (TLRs) in aquatic animals: Signaling pathways, expressions and immune responses. Immunol. Lett. 2014, 158, 14–24. [Google Scholar] [CrossRef] [PubMed]
- Star, B.; Nederbragt, A.J.; Jentoft, S.; Grimholt, U.; Malmstrom, M.; Gregers, T.F.; Rounge, T.B.; Paulsen, J.; Solbakken, M.H.; Sharma, A.; et al. The genome sequence of Atlantic cod reveals a unique immune system. Nature 2011, 477, 207–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, W.; Wu, H.; Xu, H.; Xu, T.; Zhang, X.; Chang, K.; Zhang, Y. Cloning and molecular characterization of two complement Bf/C2 genes in large yellow croaker (Pseudosciaena crocea). Fish Shellfish Immunol. 2009, 27, 285–295. [Google Scholar] [CrossRef] [PubMed]
- Kugapreethan, R.; Wan, Q.; Nilojan, J.; Lee, J. Identification and characterization of a calcium-dependent lily-type lectin from black rockfish (Sebastes schlegelii): Molecular antennas are involved in host defense via pathogen recognition. Dev. Comp. Immunol. 2017, 81, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Tan, W.; Ma, M.; Yu, X.; Lai, Q.; Wu, Z.; Lin, G.; Hu, C. Molecular characterization and transcription regulation analysis of type I IFN gene in grass carp (Ctenopharyngodon idella). Gene 2012, 504, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Perera, N.C.N.; Godahewa, G.I.; Hwang, J.Y.; Kwon, M.G.; Hwang, S.D.; Lee, J. Molecular, structural, and functional comparison of N lobe and C lobe of the transferrin from rock bream, Oplegnathus fasciatus, with respect to its immune response. Fish Shellfish Immunol. 2017, 68, 299–309. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Wang, Y.; Xu, P.; Liu, Z. NK-lysin of channel catfish: Gene triplication, sequence variation, and expression analysis. Mol. Immunol. 2006, 43, 1676–1686. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.; Xiang, X.; Jiang, Y.; Lv, Y.; Zhou, Y.; Zhong, H.; Xiao, J.; Zhang, F.; Jiang, H.; Yan, J. Identification and characterization of a novel Toll-like receptor 4 homologue in blunt snout bream, Megalobrama amblycephala. Fish Shellfish Immunol. 2016, 57, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.L.; Xu, M.Y.; Ji, X.S.; Yu, G.C. Cloning and characterisation of natural resistance associated macrophage protein (Nramp) cDNA from red sea bream (Pagrus major). Fish Shellfish Immunol. 2004, 17, 305–313. [Google Scholar] [CrossRef] [PubMed]
- Ehrnstorfer, I.A.; Geertsma, E.R.; Pardon, E.; Steyaert, J.; Dutzler, R. Crystal structure of a SLC11 (NRAMP) transporter reveals the basis for transition-metal ion transport. Nat. Struct. Mol. Biol. 2014, 21, 990–996. [Google Scholar] [CrossRef] [PubMed]
- Bairoch, A. The PROSITE dictionary of sites and patterns in proteins, its current status. Nucleic Acids Res. 1993, 21, 3097–3103. [Google Scholar] [CrossRef] [PubMed]
- Vidal, S.M.; Malo, D.; Vogan, K.; Skamene, E.; Gros, P. Natural resistance to infection with intracellular parasites: Isolation of a candidate for Bcg. Cell 1993, 73, 469–485. [Google Scholar] [CrossRef]
- Illing, A.C.; Shawki, A.; Cunningham, C.L.; Mackenzie, B. Substrate profile and metal-ion selectivity of human divalent metal-ion transporter-1. J. Biol. Chem. 2012, 287, 30485–30496. [Google Scholar] [CrossRef] [PubMed]
- Vidal, S.; Belouchi, A.M.; Cellier, M.; Beatty, B.; Gros, P. Cloning and characterization of a second human NRAMP gene on chromosome 12q13. Mamm. Genome 1995, 6, 224–230. [Google Scholar] [CrossRef] [PubMed]
- Gruenheid, S.; Cellier, M.; Vidal, S.; Gros, P. Identification and characterization of a second mouse Nramp gene. Genomics 1995, 25, 514–525. [Google Scholar] [CrossRef]
- Gruenheid, S.; Pinner, E.; Desjardins, M.; Gros, P. Natural resistance to infection with intracellular pathogens: The Nramp1 protein is recruited to the membrane of the phagosome. J. Exp. Med. 1997, 185, 717–730. [Google Scholar] [CrossRef] [PubMed]
- Searle, S.; Bright, N.A.; Roach, T.I.; Atkinson, P.G.; Barton, C.H.; Meloen, R.H.; Blackwell, J.M. Localisation of Nramp1 in macrophages: Modulation with activation and infection. J. Cell Sci. 1998, 111 Pt 19, 2855–2866. [Google Scholar] [PubMed]
- Evans, C.A.; Harbuz, M.S.; Ostenfeld, T.; Norrish, A.; Blackwell, J.M. Nramp1 is expressed in neurons and is associated with behavioural and immune responses to stress. Neurogenetics 2001, 3, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Li, Y.; Hashad, M.; Schurr, E.; Gros, P.; Adams, L.G.; Templeton, J.W. Bovine natural resistance associated macrophage protein 1 (Nramp1) gene. Genome Res. 1996, 6, 956–964. [Google Scholar] [CrossRef] [PubMed]
- Cellier, M.; Govoni, G.; Vidal, S.; Kwan, T.; Groulx, N.; Liu, J.; Sanchez, F.; Skamene, E.; Schurr, E.; Gros, P. Human natural resistance-associated macrophage protein: CDNA cloning, chromosomal mapping, genomic organization, and tissue-specific expression. J. Exp. Med. 1994, 180, 1741–1752. [Google Scholar] [CrossRef] [PubMed]
- Peracino, B.; Buracco, S.; Bozzaro, S. The Nramp (Slc11) proteins regulate development, resistance to pathogenic bacteria and iron homeostasis in Dictyostelium discoideum. J. Cell Sci. 2013, 126 Pt 1, 301–311. [Google Scholar] [CrossRef] [PubMed]
- Saeij, J.P.; Wiegertjes, G.F.; Stet, R.J. Identification and characterization of a fish natural resistance-associated macrophage protein (NRAMP) cDNA. Immunogenetics 1999, 50, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Donovan, A.; Brownlie, A.; Dorschner, M.O.; Zhou, Y.; Pratt, S.J.; Paw, B.H.; Phillips, R.B.; Thisse, C.; Thisse, B.; Zon, L.I. The zebrafish mutant gene chardonnay (cdy) encodes divalent metal transporter 1 (DMT1). Blood 2002, 100, 4655–4659. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Waldbieser, G.C.; Rice, C.D.; Elibol, B.; Wolters, W.R.; Hanson, L.A. Isolation and characterization of channel catfish natural resistance associated macrophage protein gene. Dev. Comp. Immunol. 2002, 26, 517–531. [Google Scholar] [CrossRef]
- Sibthorpe, D.; Baker, A.M.; Gilmartin, B.J.; Blackwell, J.M.; White, J.K. Comparative analysis of two slc11 (Nramp) loci in Takifugu rubripes. DNA Cell Biol. 2004, 23, 45–58. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.D.; Xu, J.; Luo, X.S.; Zhou, Y.; Xiao, Y.; Zeng, L.B. Molecular cloning, characterization and expression of natural resistance associated macrophage protein (Nramp) gene cDNA from grass carp (Ctenopharyngodon idella). J. Fishery Sci. China 2011, 18, 38–47. [Google Scholar] [CrossRef]
- Burge, E.J.; Gauthier, D.T.; Van Veld, P.A. In vitro response of the striped bass natural resistance-associated macrophage protein, Nramp, to LPS and Mycobacterium marinum exposure. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2004, 138, 391–400. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods (San Diego, Calif.) 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Casey, J.L.; Hentze, M.W.; Koeller, D.M.; Caughman, S.W.; Rouault, T.A.; Klausner, R.D.; Harford, J.B. Iron-responsive elements: Regulatory RNA sequences that control mRNA levels and translation. Science 1988, 240, 924–928. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.L.; Wang, Z.J.; Xu, M.Y.; Gui, J.F. Molecular identification and expression analysis of natural resistance associated macrophage protein (Nramp) cDNA from Japanese flounder (Paralichthys olivaceus). Fish Shellfish Immunol. 2006, 20, 365–373. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.L.; Zhang, Y.X.; Xu, J.Y.; Meng, L.; Sha, Z.X.; Ren, G.C. Molecular cloning, characterization and expression analysis of natural resistance associated macrophage protein (Nramp) cDNA from turbot (Scophthalmus maximus). Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2007, 147, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Gunshin, H.; Mackenzie, B.; Berger, U.V.; Gunshin, Y.; Romero, M.F.; Boron, W.F.; Nussberger, S.; Gollan, J.L.; Hediger, M.A. Cloning and characterization of a mammalian proton-coupled metal-ion transporter. Nature 1997, 388, 482–488. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.L.; Gelbart, T.; West, C.; Halloran, C.; Beutler, E. The human Nramp2 gene: Characterization of the gene structure, alternative splicing, promoter region and polymorphisms. Blood Cells Mol. Dis. 1998, 24, 199–215. [Google Scholar] [CrossRef] [PubMed]
- Neves, J.V.; Wilson, J.M.; Kuhl, H.; Reinhardt, R.; Castro, L.F.; Rodrigues, P.N. Natural history of SLC11 genes in vertebrates: Tales from the fish world. BMC Evol. Biol. 2011, 11, 106. [Google Scholar] [CrossRef] [PubMed]
- Xing, H.F.; Gao, F.T.; Zhang, Y.Z.; Dong, Z.D.; Chen, S.L. Expression and SNP Screening of Natural Resistance-Associated Macrophage Protein (Nramp) Gene cDNA from Half Smooth Tongue Sole (Cynoglossus semilaevis). Prog. Fish Sci. 2016, 37, 116–127. [Google Scholar]
- Jia, A.; Zhang, X.H. Molecular cloning, characterization and expression analysis of cathepsin D gene from turbot Scophthalmus maximus. Fish Shellfish Immunol. 2009, 26, 606–613. [Google Scholar] [CrossRef] [PubMed]
- Press, C.M.; Evensen, Ø. The morphology of the immune system in teleost fishes. Fish Shellfish Immunol. 1999, 9, 309–318. [Google Scholar] [CrossRef]
- Fänge, R.; Nilsson, S. The fish spleen: Structure and function. Experientia 1985, 41, 152–158. [Google Scholar] [CrossRef] [PubMed]
- Van Muiswinkel, W.B.; Lamers, C.H.; Rombout, J.H. Structural and functional aspects of the spleen in bony fish. Res. Immunol. 1991, 142, 362–366. [Google Scholar] [CrossRef]
- Wardrop, S.L.; Richardson, D.R. Interferon-gamma and lipopolysaccharide regulate the expression of Nramp2 and increase the uptake of iron from low relative molecular mass complexes by macrophages. Eur. J. Biochem. 2000, 267, 6586–6593. [Google Scholar] [CrossRef] [PubMed]
- Bonecchi, R.; Garlanda, C.; Mantovani, A.; Riva, F. Cytokine decoy and scavenger receptors as key regulators of immunity and inflammation. Cytokine 2016, 87, 37–45. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence(5′ → 3′) | Comment |
|---|---|---|
| Nramp-F | ACYATYATYGGGCTSCTG | Gene cloning |
| Nramp-R | ACCACHCCCCCTTTGTAGAT | - |
| 3′-Adaptor primer | GCTGTCAACGATACGCTACGTAACGGCATGACAGTG(T)18 | 3′RACE |
| 3′-Primer | GCTGTCAACGATACGCTACGTAACG | - |
| 3′-Nested primer | CGCTACGTAACGGCATGACAGTG | - |
| Nramp-3′-GSP | ATCGCCCTCAATCTGCTGTCGGTCGG | - |
| Nramp-3′-NGSP | GTTTTTGTGGTGTCCGTCTTCGCCG | - |
| AAP | GGCCACGCGTCGACTAGTACGGGIIGGGIIGGGIIG | 5′RACE |
| AUAP | GGCCACGCGTCGACTAGTAC | - |
| Nramp-5′-GSP | TCCTGCATGTCAGAGCCGA | - |
| Nramp-5′-NGSP | CGGCACCGTGGGGTAATGG | - |
| Nramp-qF | GGACATCTACAAAGGGGGTG | Real-time PCR |
| Nramp-qR | AAATCCAGACGAGAGACGCC | - |
| β-actin-qF | TCTACAACGAGCTGCGTGTTG | - |
| β-actin-qR | TCAATCCCAAAGCCAACAGG | - |
| 18S rRNA-qF | CAAGACGGACGAGAGCGAAA | - |
| 18S rRNA-qR | GCGGGTTGGCATAGTTTACG | - |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, Y.-H.; Mao, Y.; Lv, Y.-N.; Tang, L.-L.; Zhou, Y.; Zhong, H.; Xiao, J.; Yan, J.-P. Natural Resistance Associated Macrophage Protein Is Involved in Immune Response of Blunt Snout Bream, Megalobrama amblycephala. Cells 2018, 7, 27. https://0-doi-org.brum.beds.ac.uk/10.3390/cells7040027
Jiang Y-H, Mao Y, Lv Y-N, Tang L-L, Zhou Y, Zhong H, Xiao J, Yan J-P. Natural Resistance Associated Macrophage Protein Is Involved in Immune Response of Blunt Snout Bream, Megalobrama amblycephala. Cells. 2018; 7(4):27. https://0-doi-org.brum.beds.ac.uk/10.3390/cells7040027
Chicago/Turabian StyleJiang, Yu-Hong, Ying Mao, Yi-Na Lv, Lei-Lei Tang, Yi Zhou, Huan Zhong, Jun Xiao, and Jin-Peng Yan. 2018. "Natural Resistance Associated Macrophage Protein Is Involved in Immune Response of Blunt Snout Bream, Megalobrama amblycephala" Cells 7, no. 4: 27. https://0-doi-org.brum.beds.ac.uk/10.3390/cells7040027