Shape-Shifted Red Blood Cells: A Novel Red Blood Cell Stage?

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Cell Cultures
2.3. Transmission Electronic Microscopy (TEM) and Wright-Giemsa Staining
2.4. Flow Cytometry and Immunofluorescence
2.5. Antibodies
2.6. Cell Viability Assay
2.7. RNA-seq of RBCs and shRBCs
2.8. Proteomic Sequencing
2.9. Pathway Enrichment Analysis
2.10. Preparation of CM from shRBC Cell Cultures
2.11. TPS-2 and shRBCs Infection with VHSV
2.12. Rainbow Trout In Vivo Heat Stress
2.13. RBCs Exposed to Different Pathogen-Associated Molecular Patterns (PAMPs)
2.14. Software and Statistics
3. Results
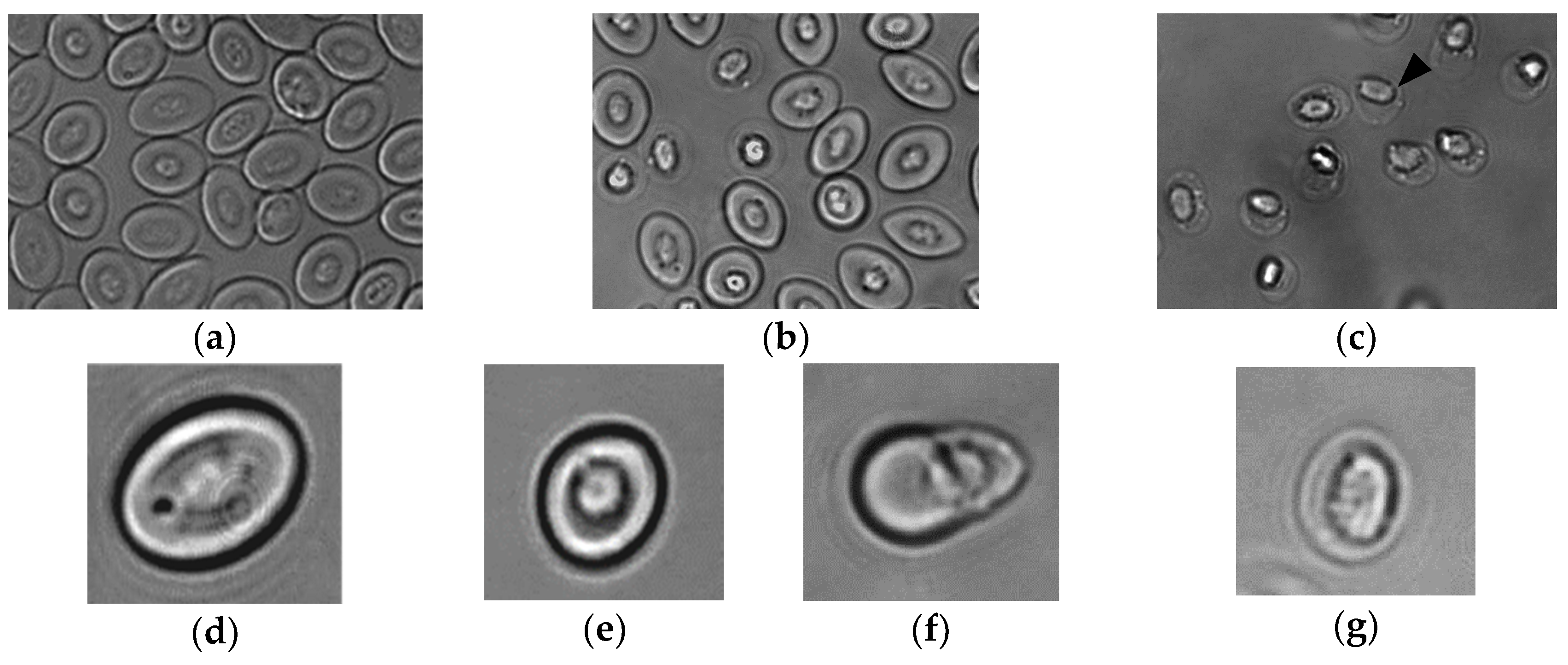
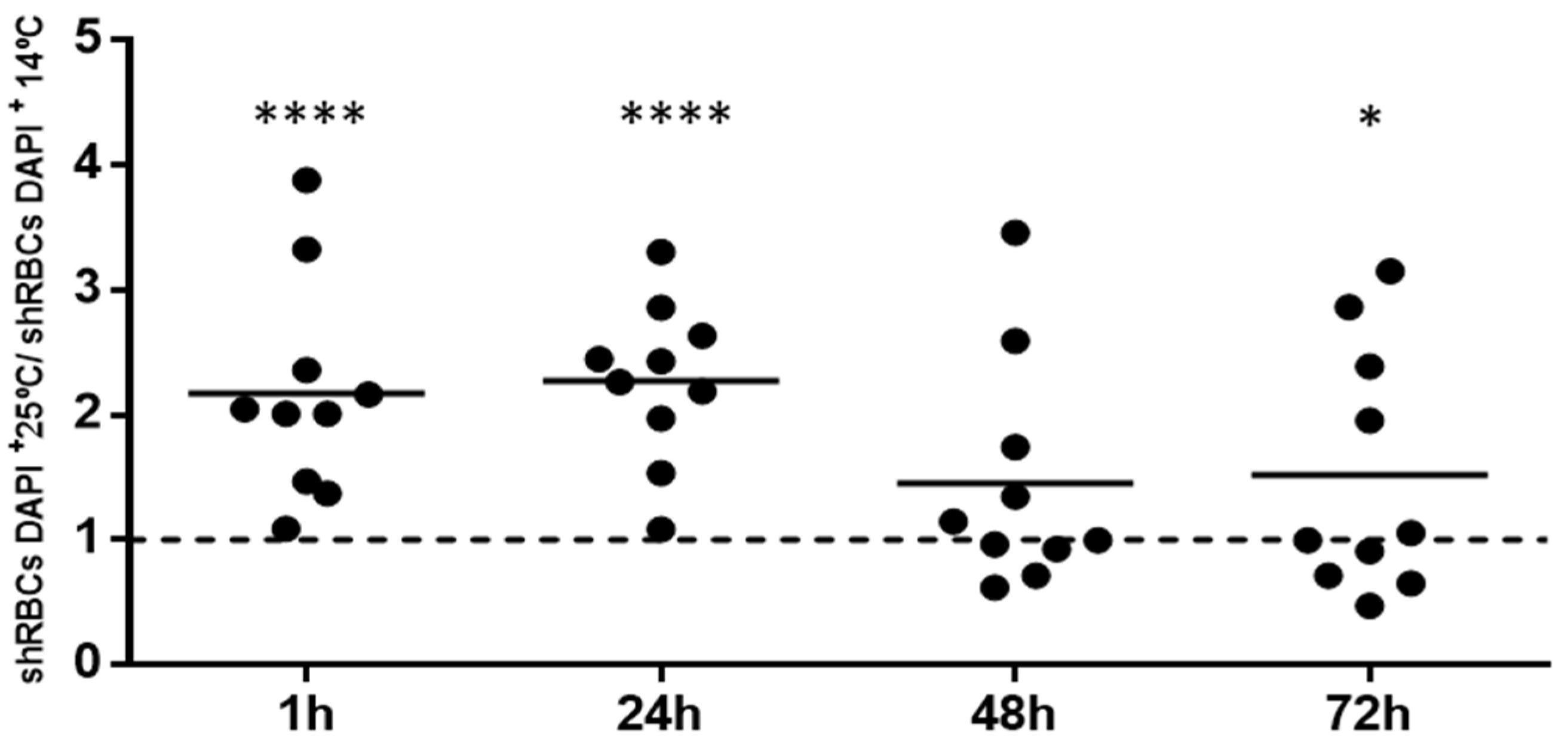
3.1. Rainbow Trout RBC Transformation Sequence to shRBC In Vitro
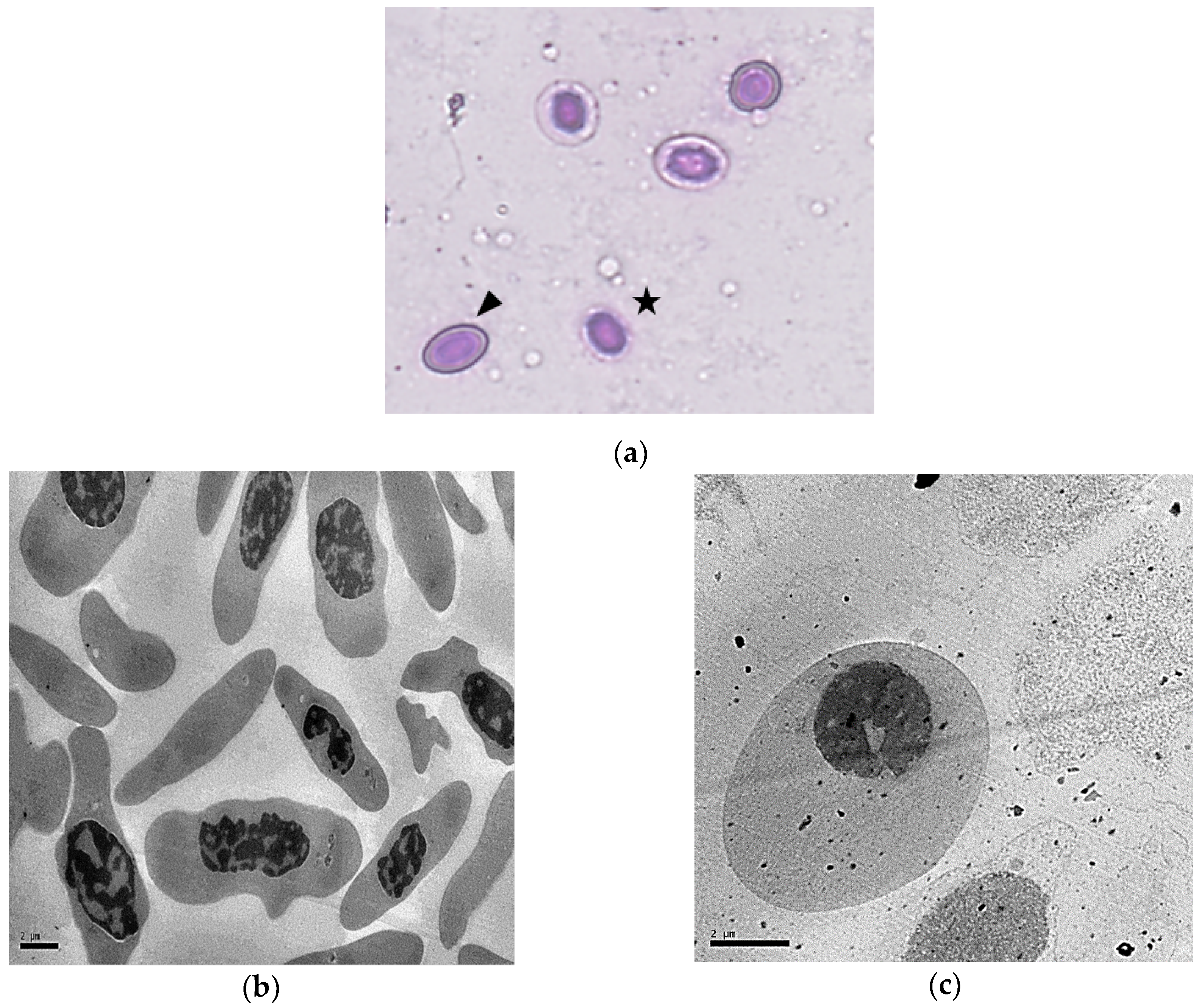
3.2. Morphological Characterization of Rainbow Trout shRBCs
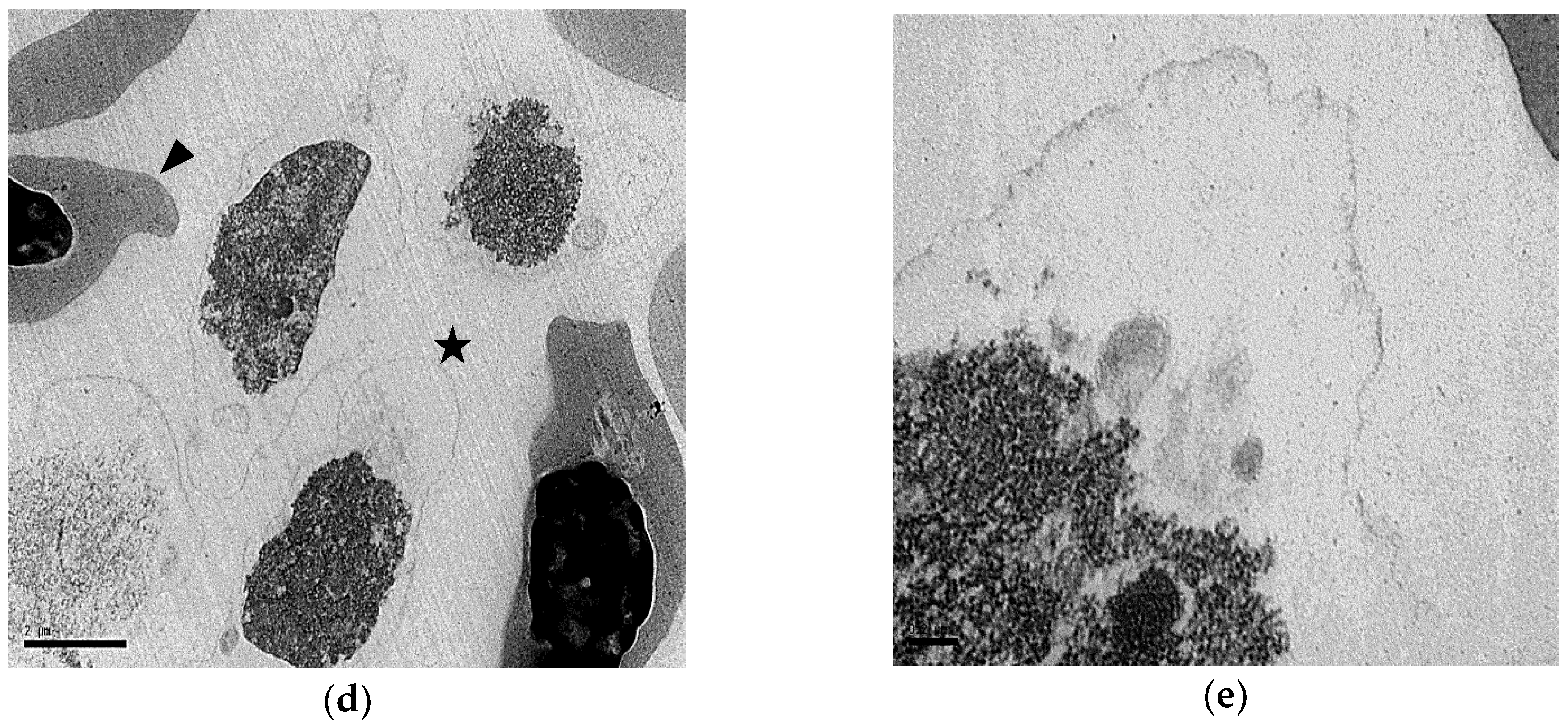
3.3. shRBCs Share Properties with Mammalian Primitive Pyrenocytes
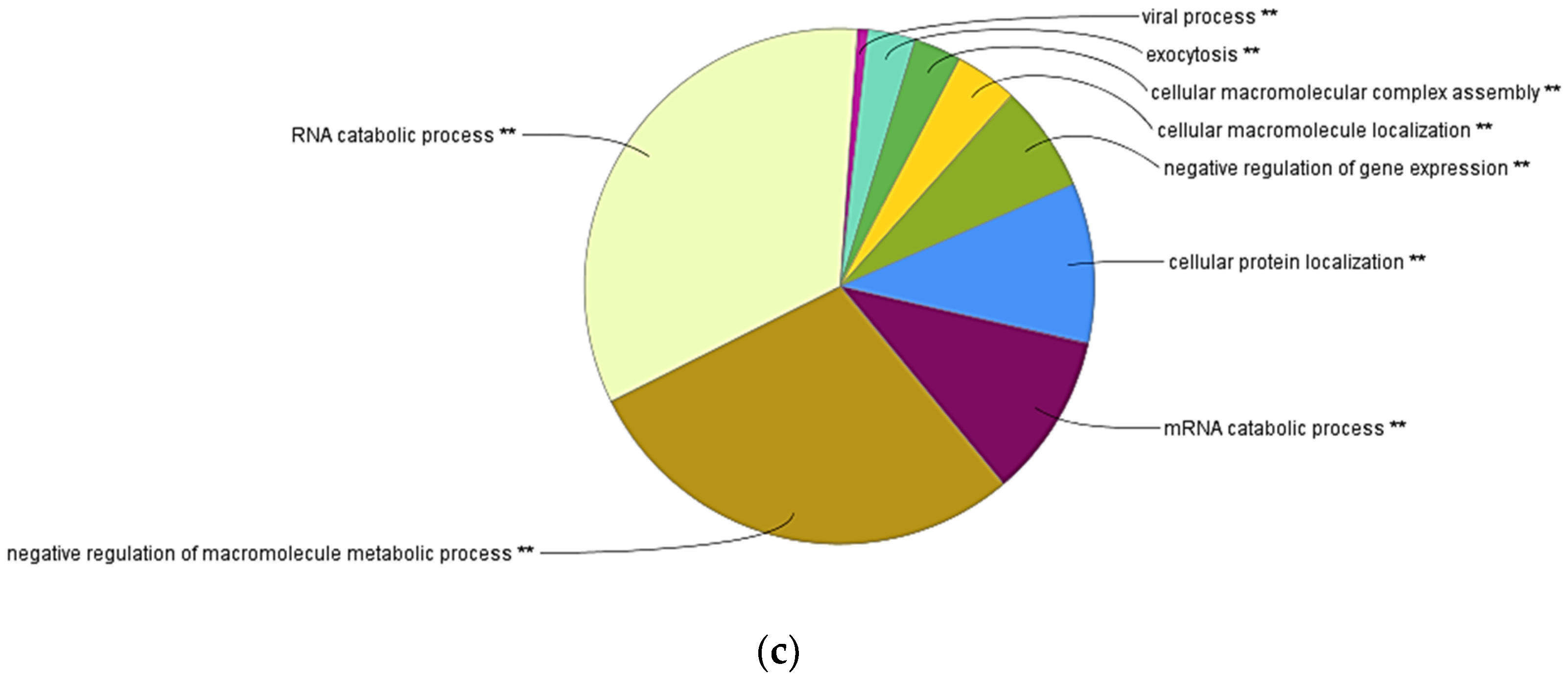
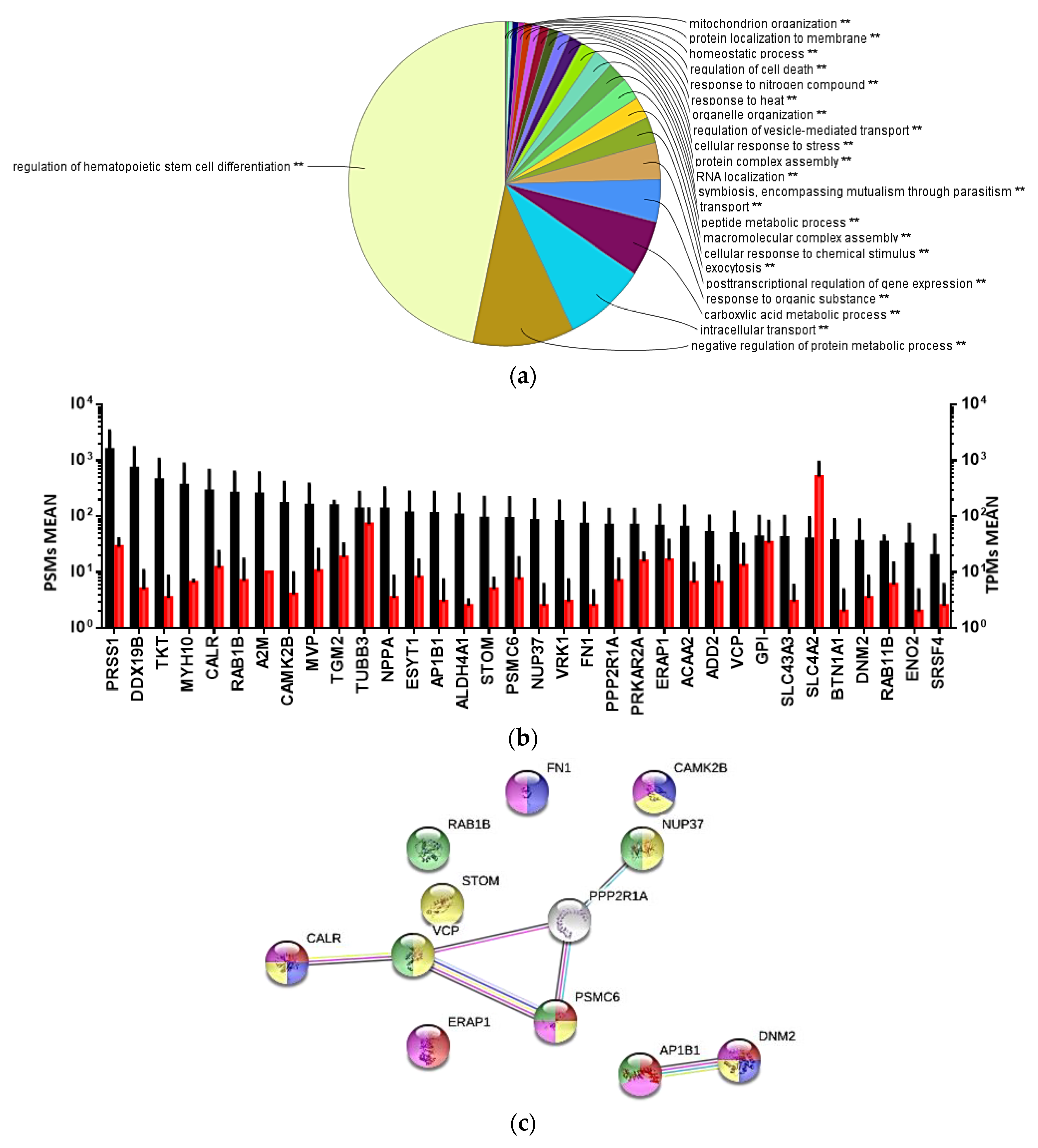
3.4. Transcriptome Evaluation of shRBCs
3.5. Proteomic Sequencing of shRBCs
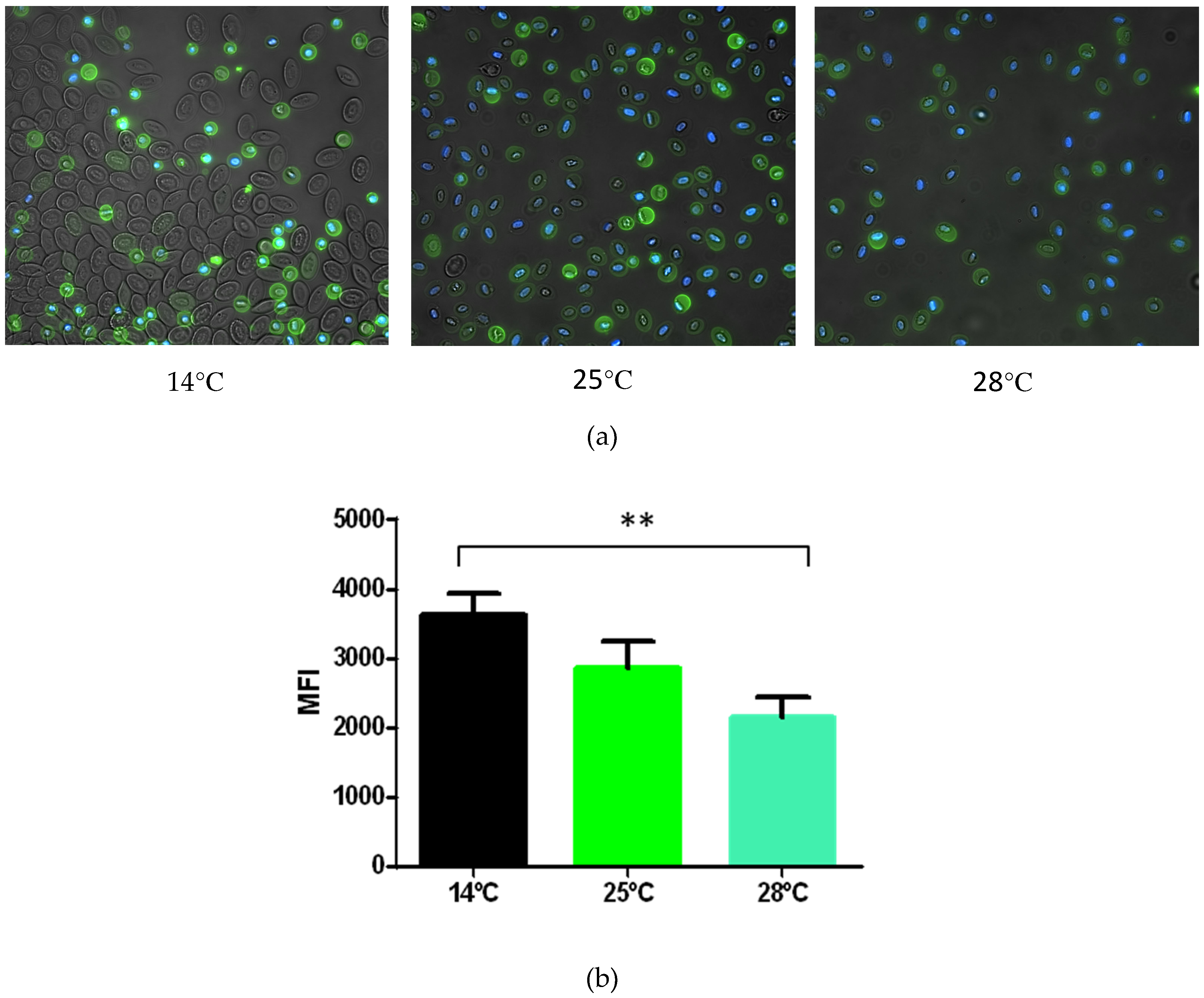
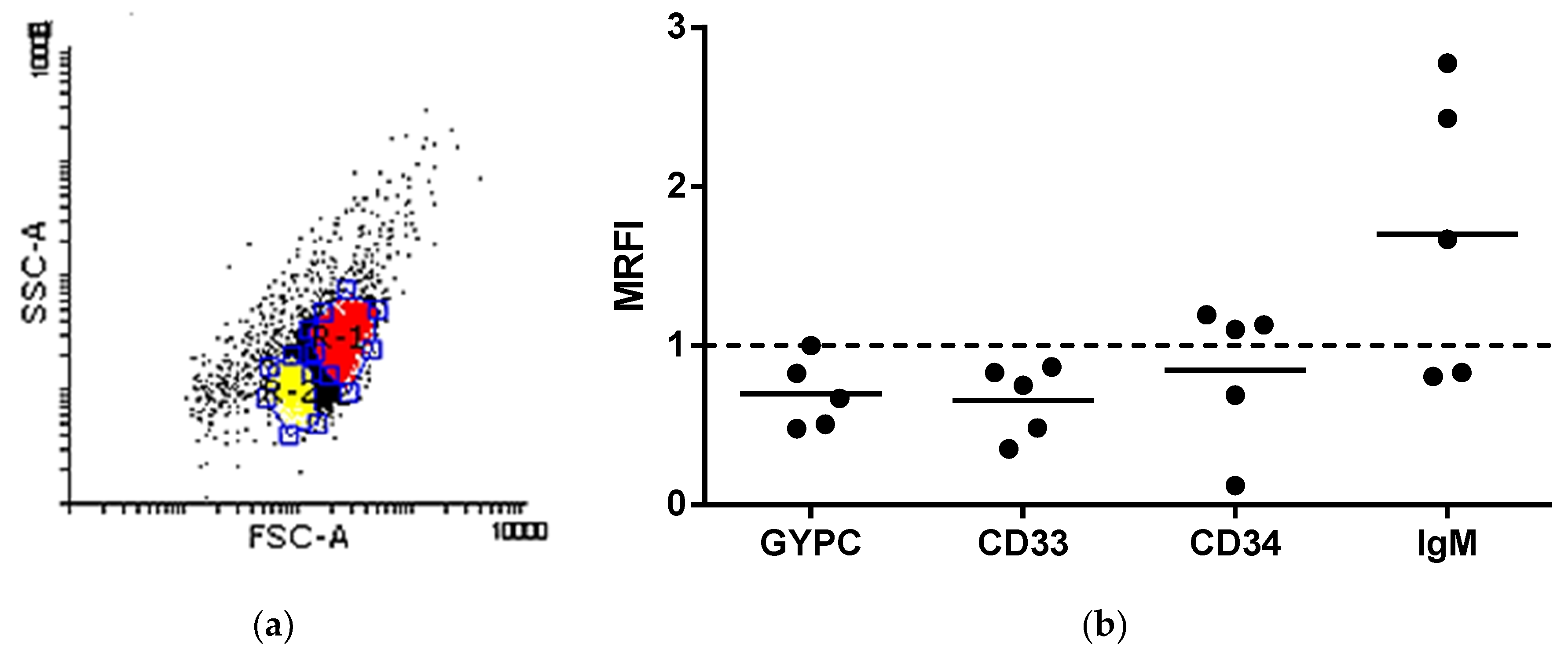
3.6. Protein Cell Marker Screening in shRBCs
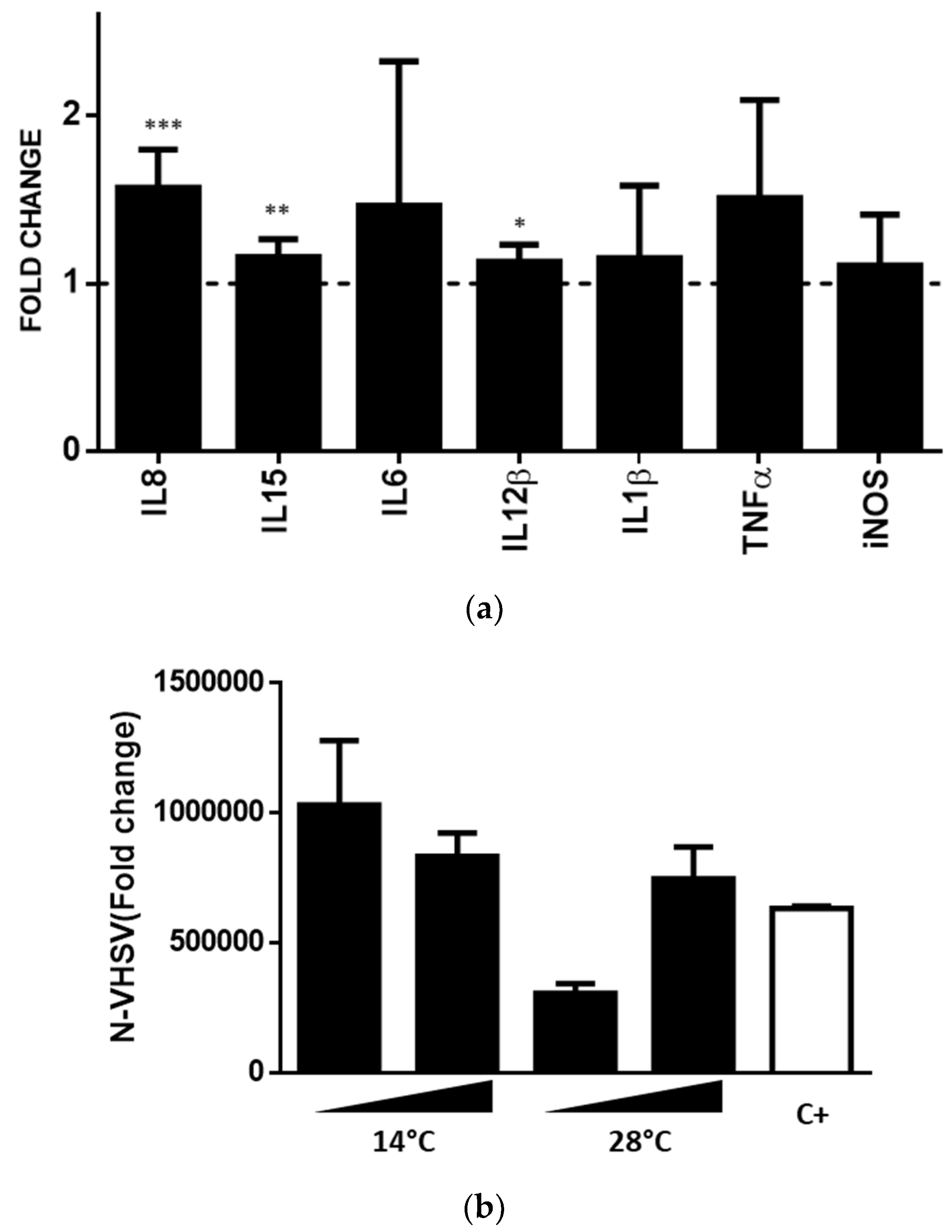
3.7. shRBCs Immune Response to VHSV, a Viral Pathogen
3.8. shRBCs CM Triggered TPS-2 Cytokine Signalling
3.9. shRBCs in Circulating Blood of Rainbow Trout
4. Discussion
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Passantino, L.; Altamura, M.; Cianciotta, A.; Patruno, R.; Tafaro, A.; Jirillo, E.; Passantino, G.F. Fish immunology. I. Binding and engulfment of candida albicans by erythrocytes of rainbow trout (salmo gairdneri richardson). Immunopharmacol. Immunotoxicol. 2002, 24, 665–678. [Google Scholar] [CrossRef] [PubMed]
- Morera, D.; Mackenzie, S.A. Is there a direct role for erythrocytes in the immune response? Vet. Res. 2011, 42, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morera, D.; Roher, N.; Ribas, L.; Balasch, J.C.; Donate, C.; Callol, A.; Boltana, S.; Roberts, S.; Goetz, G.; Goetz, F.W.; et al. Rna-seq reveals an integrated immune response in nucleated erythrocytes. PLoS ONE 2011, 6, e26998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Workenhe, S.T.; Kibenge, M.J.; Wright, G.M.; Wadowska, D.W.; Groman, D.B.; Kibenge, F.S. Infectious salmon anaemia virus replication and induction of alpha interferon in atlantic salmon erythrocytes. Virol. J. 2008, 5, 36. [Google Scholar] [CrossRef] [PubMed]
- Dahle, M.K.; Wessel, O.; Timmerhaus, G.; Nyman, I.B.; Jorgensen, S.M.; Rimstad, E.; Krasnov, A. Transcriptome analyses of atlantic salmon (salmo salar L.) erythrocytes infected with piscine orthoreovirus (prv). Fish Shellfish Immunol. 2015, 45, 780–790. [Google Scholar] [CrossRef] [PubMed]
- Nombela, I.; Puente-Marin, S.; Chico, V.; Villena, A.; Carracedo, B.; Ciordia, S.; Mena, M.; Mercado, L.; Perez, L.; Coll, J.; et al. Identification of Diverse Defense Mechanisms in Trout Red Blood Cells in Response to Vhsv Halted Viral Replication. F1000Research 2017, 6. [Google Scholar] [CrossRef]
- Nombela, I.; Carrion, A.; Puente-Marin, S.; Chico, V.; Mercado, L.; Perez, L.; Coll, J.; Ortega-Villaizan, M.D.M. Infectious pancreatic necrosis virus triggers antiviral immune response in rainbow trout red blood cells, despite not being infective. F1000Research 2017, 6. [Google Scholar] [CrossRef] [PubMed]
- Nombela, I.; Ortega-Villaizan, M. Nucleated red blood cells: Immune cell mediators of the antiviral response. PLoS Pathog. 2018, in press. [Google Scholar]
- Pereiro, P.; Romero, A.; Diaz-Rosales, P.; Estepa, A.; Figueras, A.; Novoa, B. Nucleated teleost erythrocytes play an nk-lysin- and autophagy-dependent role in antiviral immunity. Front. Immunol. 2017, 8, 1458. [Google Scholar] [CrossRef] [PubMed]
- Cui, L.; Takada, H.; Takimoto, T.; Fujiyoshi, J.; Ishimura, M.; Hara, T. Immunoregulatory function of neonatal nucleated red blood cells in humans. Immunobiology 2016, 221, 853–861. [Google Scholar] [CrossRef] [PubMed]
- Akbari, A. The Role of Red Blood Cells in Wound Healing. Master’s Thesis, University of British Columbia, Vancouver, BC, Canada, 2011. [Google Scholar]
- Lewis, J.M.; Hori, T.S.; Rise, M.L.; Walsh, P.J.; Currie, S. Transcriptome responses to heat stress in the nucleated red blood cells of the rainbow trout (Oncorhynchus mykiss). Physiol. Genom. 2010, 42, 361–373. [Google Scholar] [CrossRef] [PubMed]
- McGrath, K.E.; Kingsley, P.D.; Koniski, A.D.; Porter, R.L.; Bushnell, T.P.; Palis, J. Enucleation of primitive erythroid cells generates a transient population of “pyrenocytes” in the mammalian fetus. Blood 2008, 111, 2409–2417. [Google Scholar] [CrossRef] [PubMed]
- Gautier, E.F.; Ducamp, S.; Leduc, M.; Salnot, V.; Guillonneau, F.; Dussiot, M.; Hale, J.; Giarratana, M.C.; Raimbault, A.; Douay, L.; et al. Comprehensive proteomic analysis of human erythropoiesis. Cell Rep. 2016, 16, 1470–1484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kingsley, P.D.; Malik, J.; Fantauzzo, K.A.; Palis, J. Yolk sac-derived primitive erythroblasts enucleate during mammalian embryogenesis. Blood 2004, 104, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, H.; Kawane, K.; Koike, M.; Mori, Y.; Uchiyama, Y.; Nagata, S. Phosphatidylserine-dependent engulfment by macrophages of nuclei from erythroid precursor cells. Nature 2005, 437, 754–758. [Google Scholar] [CrossRef] [PubMed]
- Carracedo, B. Caracterización de Cultivos Celulares Angiogénicos Obtenidos de Órganos Linfoides de Trucha Arco Iris, Oncorhynchus Mykiss. Ph.D. Thesis, University of León, León, Spain, 2003. [Google Scholar]
- Winton, J.; Batts, W.; deKinkelin, P.; LeBerre, M.; Bremont, M.; Fijan, N. Current lineages of the epithelioma papulosum cyprini (epc) cell line are contaminated with fathead minnow, pimephales promelas, cells. J. Fish Dis. 2010, 33, 701–704. [Google Scholar] [CrossRef] [PubMed]
- Chico, V.; Martinez-Lopez, A.; Ortega-Villaizan, M.; Falco, A.; Perez, L.; Coll, J.M.; Estepa, A. Pepscan mapping of viral hemorrhagic septicemia virus glycoprotein g major lineal determinants implicated in triggering host cell antiviral responses mediated by type i interferon. J. Virol. 2010, 84, 7140–7150. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Lopez, A.; Garcia-Valtanen, P.; Ortega-Villaizan, M.; Chico, V.; Gomez-Casado, E.; Coll, J.M.; Estepa, A. Vhsv g glycoprotein major determinants implicated in triggering the host type i ifn antiviral response as DNA vaccine molecular adjuvants. Vaccine 2014, 32, 6012–6019. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, P.; Wacyk, J.; Morales-Lange, B.; Rojas, V.; Guzman, F.; Dixon, B.; Mercado, L. Immunomodulatory effect of cathelicidins in response to a beta-glucan in intestinal epithelial cells from rainbow trout. Dev. Comp. Immunol. 2015, 51, 160–169. [Google Scholar] [CrossRef] [PubMed]
- Santana, P.; Palacios, C.; Narváez, E.; Guzmán, F.; Gallardo, J.A.; Mercado, L. Anti-peptide antibodies: A tool for detecting il-8 in salmonids. Electron. J. Biotechnol. 2012, 15. [Google Scholar] [CrossRef]
- Ordas, M.C.; Cuesta, A.; Mercado, L.; Bols, N.C.; Tafalla, C. Viral hemorrhagic septicaemia virus (vhsv) up-regulates the cytotoxic activity and the perforin/granzyme pathway in the rainbow trout rts11 cell line. Fish Shellfish Immunol. 2011, 31, 252–259. [Google Scholar] [CrossRef] [PubMed]
- Sanz, F.; Basurco, B.; Babin, M.; Dominguez, J.; Coll, J.M. Monoclonal antibodies against the structural proteins of viral haemorrhagic septicaemia virus isolates. J. Fish Dis. 1993, 16, 53–63. [Google Scholar] [CrossRef]
- Sanchez, C.; Lopez-Fierro, P.; Zapata, A.; Dominguez, J. Characterization of monoclonal antibodies against heavy and light chains of trout immunoglobulin. Fish Shellfish Immunol. 1993, 3, 237–251. [Google Scholar] [CrossRef]
- Sanchez, C.; Dominguez, J. Trout immuglobulin populations differing in light chains revealed by monoclonal antibodies. Mol. Immunol. 1991, 28, 1271–1277. [Google Scholar] [CrossRef]
- Puente-Marin, S.; Nombela, I.; Ciordia, S.; Mena, M.; Chico, V.; Coll, J.; Ortega-Villaizan, M. In silico functional networks identified in fish nucleated red blood cells by means of transcriptomic and proteomic profiling. Genes 2018, 9, 202. [Google Scholar] [CrossRef] [PubMed]
- Bindea, G.; Mlecnik, B.; Hackl, H.; Charoentong, P.; Tosolini, M.; Kirilovsky, A.; Fridman, W.H.; Pages, F.; Trajanoski, Z.; Galon, J. Cluego: A cytoscape plug-in to decipher functionally grouped gene ontology and pathway annotation networks. Bioinformatics 2009, 25, 1091–1093. [Google Scholar] [CrossRef] [PubMed]
- Bindea, G.; Galon, J.; Mlecnik, B. Cluepedia cytoscape plugin: Pathway insights using integrated experimental and in silico data. Bioinformatics 2013, 29, 661–663. [Google Scholar] [CrossRef] [PubMed]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Franceschini, A.; Wyder, S.; Forslund, K.; Heller, D.; Huerta-Cepas, J.; Simonovic, M.; Roth, A.; Santos, A.; Tsafou, K.P.; et al. String v10: Protein-protein interaction networks, integrated over the tree of life. Nucleic Acids Res. 2015, 43, D447–D452. [Google Scholar] [CrossRef] [PubMed]
- Gotz, S.; Garcia-Gomez, J.M.; Terol, J.; Williams, T.D.; Nagaraj, S.H.; Nueda, M.J.; Robles, M.; Talon, M.; Dopazo, J.; Conesa, A. High-throughput functional annotation and data mining with the blast2go suite. Nucleic Acids Res. 2008, 36, 3420–3435. [Google Scholar] [CrossRef] [PubMed]
- LeBerre, M.; De Kinkelin, P.; Metzger, A. Quantification sérologique des rhabddovirus de salmonidés. Bull. Off. Int. Epizoot. 1977, 87, 391–393. [Google Scholar]
- Basurco, B.; Coll, J.M. Spanish isolates and reference strains of viral haemorrhagic septicaemia virus shown similar protein size patterns. Bull. Eur. Assoc. Fish Pathol. 1989, 9, 92–95. [Google Scholar]
- Raida, M.K.; Buchmann, K. Temperature-dependent expression of immune-relevant genes in rainbow trout following yersinia ruckeri vaccination. Dis. Aquat. Organ. 2007, 77, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Chico, V.; Gomez, N.; Estepa, A.; Perez, L. Rapid detection and quantitation of viral hemorrhagic septicemia virus in experimentally challenged rainbow trout by real-time rt-pcr. J. Virol. Methods 2006, 132, 154–159. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Bird, S.; Koussounadis, A.; Holland, J.W.; Carrington, A.; Zou, J.; Secombes, C.J. Identification of a novel il-1 cytokine family member in teleost fish. J. Immunol. 2009, 183, 962–974. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Diaz-Rosales, P.; Costa, M.M.; Campbell, S.; Snow, M.; Collet, B.; Martin, S.A.; Secombes, C.J. Functional characterization of a nonmammalian il-21: Rainbow trout Oncorhynchus mykiss il-21 upregulates the expression of the th cell signature cytokines ifn-gamma, il-10, and il-22. J. Immunol. 2011, 186, 708–721. [Google Scholar] [CrossRef] [PubMed]
- Purcell, M.K.; Nichols, K.M.; Winton, J.R.; Kurath, G.; Thorgaard, G.H.; Wheeler, P.; Hansen, J.D.; Herwig, R.P.; Park, L.K. Comprehensive gene expression profiling following DNA vaccination of rainbow trout against infectious hematopoietic necrosis virus. Mol. Immunol. 2006, 43, 2089–2106. [Google Scholar] [CrossRef] [PubMed]
- Rendell, J.L.; Fowler, S.; Cockshutt, A.; Currie, S. Development-dependent differences in intracellular localization of stress proteins (hsps) in rainbow trout, Oncorhynchus mykiss, following heat shock. Comp. Biochem. Physiol. Part D Genom. Proteom. 2006, 1, 238–252. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Lu, Z.H.; Leno, G.H. The binding of orc2 to chromatin from terminally differentiated cells. Exp. Cell Res. 2002, 274, 334–341. [Google Scholar] [CrossRef] [PubMed]
- Li, D.Q.; Nair, S.S.; Kumar, R. The morc family: New epigenetic regulators of transcription and DNA damage response. Epigenetics 2013, 8, 685–693. [Google Scholar] [CrossRef] [PubMed]
- Jadhav, G.; Teguh, D.; Kenny, J.; Tickner, J.; Xu, J. Morc3 mutant mice exhibit reduced cortical area and thickness, accompanied by altered haematopoietic stem cells niche and bone cell differentiation. Sci. Rep. 2016, 6, 25964. [Google Scholar] [CrossRef] [PubMed]
- Lo, Y.H.; Chung, E.; Li, Z.; Wan, Y.W.; Mahe, M.M.; Chen, M.S.; Noah, T.K.; Bell, K.N.; Yalamanchili, H.K.; Klisch, T.J.; et al. Transcriptional regulation by atoh1 and its target spdef in the intestine. Cell. Mol. Gastroenterol. Hepatol. 2017, 3, 51–71. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Huang, X.; An, Y.; Ren, F.; Yang, Z.Z.; Zhu, H.; Zhou, L.; He, X.; Schachner, M.; Xiao, Z.; et al. Cell recognition molecule l1 promotes embryonic stem cell differentiation through the regulation of cell surface glycosylation. Biochem. Biophys. Res. Commun. 2013, 440, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Lang, F.; Henke, G.; Embark, H.M.; Waldegger, S.; Palmada, M.; Bohmer, C.; Vallon, V. Regulation of channels by the serum and glucocorticoid-inducible kinase—Implications for transport, excitability and cell proliferation. Cell. Physiol. Biochem. 2003, 13, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Nemoto, E.; Kots, A.Y.; Kaslow, H.R.; Dennert, G. Regulation of cytotoxic t cells by ecto-nicotinamide adenine dinucleotide (nad) correlates with cell surface gpi-anchored/arginine adp-ribosyltransferase. J. Immunol. 1994, 153, 4048–4058. [Google Scholar] [PubMed]
- Peng, X.; Qin, Z.; Zhang, G.; Guo, Y.; Huang, J. Integration of the proteome and transcriptome reveals multiple levels of gene regulation in the rice dl2 mutant. Front. Plant Sci. 2015, 6, 351. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Secombes, C.J. The cytokine networks of adaptive immunity in fish. Fish Shellfish Immunol. 2013, 35, 1703–1718. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Holland, J.W.; Carrington, A.; Zou, J.; Secombes, C.J. Molecular and functional characterization of il-15 in rainbow trout Oncorhynchus mykiss: A potent inducer of ifn-gamma expression in spleen leukocytes. J. Immunol. 2007, 179, 1475–1488. [Google Scholar] [CrossRef] [PubMed]
- Meenakshi, S. Ultrastructure of spleen in the freshwater fish, tilapia mossambica (peters). Eur. Acad. Res. 2014, 2, 109–122. [Google Scholar]
- Lu, Z.H.; Xu, H.; Leno, G.H. DNA replication in quiescent cell nuclei: Regulation by the nuclear envelope and chromatin structure. Mol. Biol. Cell 1999, 10, 4091–4106. [Google Scholar] [CrossRef] [PubMed]
- Seledtsova, G.V.; Seledtsov, V.I.; Taraban, V.Y.; Samarin, D.M.; Kozlov, V.A. A role for interferon-gamma and transforming growth factor-beta in erythroid cell-mediated regulation of nitric oxide production in macrophages. Immunology 1997, 91, 109–113. [Google Scholar] [PubMed]
- Passantino, L.; Massaro, M.A.; Jirillo, F.; Di Modugno, D.; Ribaud, M.R.; Modugno, G.D.; Passantino, G.F.; Jirillo, E. Antigenically activated avian erythrocytes release cytokine-like factors: A conserved phylogenetic function discovered in fish. Immunopharmacol. Immunotoxicol. 2007, 29, 141–152. [Google Scholar] [CrossRef] [PubMed]
- St Paul, M.; Paolucci, S.; Barjesteh, N.; Wood, R.D.; Sharif, S. Chicken erythrocytes respond to toll-like receptor ligands by up-regulating cytokine transcripts. Res. Vet. Sci. 2013, 95, 87–91. [Google Scholar] [CrossRef] [PubMed]
- Shau, H.; Gupta, R.K.; Golub, S.H. Identification of a natural killer enhancing factor (nkef) from human erythroid cells. Cell. Immunol. 1993, 147, 1–11. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward primer (5′–3′) | Reverse primer (5′–3′) | Probe (5′–3′) | Reference or Accession Number |
|---|---|---|---|---|
| ef1α | ACCCTCCTCTTGGTCGTTTC | TGATGACACCAACAGCAACA | GCTGTGCGTGACATGAGGCA | [35] |
| N-VHSV | GACTCAACGGGACAGGAATGA | GGGCAATGCCCAAGTTGTT | TGGGTTGTTCACCCAGGCCGC | [36] |
| il8 | AGAGACACTGAGATCATTGCCAC | CCCTCTTCATTTGTTGTTGGC | TCCTGGCCCTCCTGACCATTACTGAG | [37] |
| ifnγ | TCCAAGGACCAGCTGTTCAAC | TCATCAACACCCTCTGCTCACT | CCTGTTTTCCCCAAGGACACGTTTGAG | [38] |
| tnfα | AGCATGGAAGACCGTCAACGAT | ACCCTCTAAATGGATGGCTGCTT | AAAAGATACCCACCATACATTGAAGCAGATTGCC | [39] |
| il1β | GCCCCCAACCGCCTTA | CAGTGTTTGCGGCCATCTTA | ACCTTCACCATCCAGCGCCACAA | AJ278242.2 |
| il15 | TACTATCCACACCAGCGTCTGAAC | TTTCAGCAGCACCAGCAATG | TTCATAATATTGAGCTGCCTGAGTGCCACC | [6] |
| il6 | ACTCCCCTCTGTCACACACC | GGCAGACAGGTCCTCCACTA | CCACTGTGCTGATAGGGCTGG | HF913655.1 |
| il12β | TGACAGCCAGGAATCTTGCA | GAAAGCGAATGTGTCAGTTCAAA | ACCCAACGACCAGCCTCCAAGATG | AJ548829.1 |
| inos | TCAGAACCTCCTCCACAA | GTGTACTCCTGAGAGTCCTTT | GCACCGACAGCGTCTA | AJ300555.1 |
| Gene Symbol | Gene Description | Log2 Fold Change |
|---|---|---|
| MT-ND2 | NADH-ubiquinone oxidoreductase chain 2 | 17.462 |
| MICU1 | Calcium uptake protein 1, mitochondrial | 16.460 |
| TFPI2 | Tissue factor pathway inhibitor 2 | 16.342 |
| ORC2 | Origin recognition complex subunit 2 | 15.899 |
| Art2a | T-cell ecto-ADP-ribosyltransferase 1 | 15.871 |
| SLIT3 | Slit homolog 3 protein | 15.443 |
| DLG2 | Disks large homolog 2 | 15.364 |
| PDZD2 | PDZ domain-containing protein 2 | 15.272 |
| SYT14 | Synaptotagmin-14 | 15.111 |
| PGM2 | Phosphoglucomutase-2 | 15.080 |
| GNAS | Guanine nucleotide-binding protein G(s) subunit alpha isoforms short | 14.969 |
| TRADD | Tumor necrosis factor receptor type 1-associated DEATH domain protein | 14.919 |
| TBX5 | T-box 5 | 14.700 |
| MUC2 | Mucin 2 | 14.648 |
| FUT9 | Fucosyltransferase 9 | 14.464 |
| SGK2 | Serum/glucocorticoid regulated kinase 2 | 14.451 |
| SHANK3 | SH3 and multiple ankyrin repeat domains 3 | 14.314 |
| SPDEF | SAM pointed domain containing ets transcription factor | 14.266 |
| MORC3 | MORC family CW-type zinc finger 3 | 14.213 |
| SNX14 | Sorting nexin 14 | 14.139 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chico, V.; Puente-Marin, S.; Nombela, I.; Ciordia, S.; Mena, M.C.; Carracedo, B.; Villena, A.; Mercado, L.; Coll, J.; Ortega-Villaizan, M.D.M. Shape-Shifted Red Blood Cells: A Novel Red Blood Cell Stage? Cells 2018, 7, 31. https://0-doi-org.brum.beds.ac.uk/10.3390/cells7040031
Chico V, Puente-Marin S, Nombela I, Ciordia S, Mena MC, Carracedo B, Villena A, Mercado L, Coll J, Ortega-Villaizan MDM. Shape-Shifted Red Blood Cells: A Novel Red Blood Cell Stage? Cells. 2018; 7(4):31. https://0-doi-org.brum.beds.ac.uk/10.3390/cells7040031
Chicago/Turabian StyleChico, Verónica, Sara Puente-Marin, Iván Nombela, Sergio Ciordia, María Carmen Mena, Begoña Carracedo, Alberto Villena, Luis Mercado, Julio Coll, and María Del Mar Ortega-Villaizan. 2018. "Shape-Shifted Red Blood Cells: A Novel Red Blood Cell Stage?" Cells 7, no. 4: 31. https://0-doi-org.brum.beds.ac.uk/10.3390/cells7040031