Genome-Wide Identification and Transcriptional Expression of the PAL Gene Family in Common Walnut (Juglans Regia L.)


1
Key Laboratory of Resource Biology and Biotechnology in Western China, Ministry of Education, College of Life Sciences, Northwest University, Xi’an, Shaanxi 710069, China
2
School of Chemistry and Chemical Engineering, Xianyang Normal University, Xianyang 712000, China
*
Author to whom correspondence should be addressed.
Genes 2019, 10(1), 46; https://0-doi-org.brum.beds.ac.uk/10.3390/genes10010046
Submission received: 2 November 2018
/
Revised: 10 January 2019
/
Accepted: 10 January 2019
/
Published: 15 January 2019
(This article belongs to the Section Plant Genetics and Genomics)
Abstract
:Juglans regia L. is an economically important crop cultivated worldwide for its high quality and quantity of wood and nuts. Phenylalanine ammonia-lyase (PAL) is the first enzyme in the phenylpropanoid pathway that plays a critical role in plant growth, development, and adaptation, but there have been few reports of the PAL gene family in common walnut. Here, we report a genome-wide study of J. regia PAL genes and analyze their phylogeny, duplication, microRNA, and transcriptional expression. A total of 12 PAL genes were identified in the common walnut and clustered into two subfamilies based on phylogenetic analysis. These common walnut PALs are distributed on eight different pseudo-chromosomes. Seven of the 12 PALs (JrPAL2-3, JrPAL4-2, JrPAL2-1, JrPAL4-1, JrPAL8, JrPAL9, and JrPAL6) were specific found in J. regia, and JrPAL3, JrPAL5, JrPAL1-2, JrPAL7, and JrPAL2-2 were found to be closely associated with the woody plant Populus trichocarpa. Additionally, the expression patterns of JrPAL3, JrPAL7, JrPAL9, and JrPAL2-1 showed that they had high expression in female and male flowers. The miRNA ath-miR830-5p regulates two genes, JrPAL5 and JrPAL1, such that they have low expression in the male and female flowers of the common walnut. Our research provides useful information for further research into the function of PAL genes in common walnut and Juglans.
1. Introduction
Phenylalanine ammonia-lyase (PAL, EC4.3.1.5), the first enzyme of the phenylpropanoid pathway, produces precursors to a variety of important secondary metabolites, such as phytoalexin, lignin, and phenolic compounds [1,2,3,4,5]. PAL, first reported in 1961 [6], potentially comprises protective compounds, such as flavonoids, furanocoumarin phytoalexins, and cell wall components [7]. The PAL gene is widely present in higher plants, and is also found in some fungi, yeasts, and very few bacteria, but not in animals [8,9]. PAL encoding genes are generally well studied and are commonly found as small gene families comprising one to five members [10,11,12,13,14]. For example, four PAL genes have been identified and functionally characterized in Arabidopsis thaliana [15,16], five in Populus trichocarpa [17], three in Scutellaria baicalensis [18], and three in Coffeac anephora [19]. However, some studies have reported that the PAL gene family has more than five members; for example, 13 PAL genes have been found in Cucumis sativus [20], 12 in Citrullus lanatus [20], 13 in Cucumis melo [21], and 16 in Vitis vinifera [22]. In Pinus taeda, five distinct PAL genes have been identified [12]. In Salix babylonica, PAL1, PAL2, PAL3, and PAL4 genes are orthologous to the poplar genes [13]. In A. thaliana, AtPAL1 and AtPAL2 are highly expressed in roots and mature flowers and are barely detectable in leaf tissues [15,16], and in most woody plants cluster into two clusters [13,14,17,20]. Moreover, low-temperature stress enhances PAL activity [23]. CaPAL1 acts as a positive regulator in the phenylpropanoid pathway [24]. PAL functions as a positive regulator of rice allelopathic potential [25].
Common walnut (Juglans regia L.), also known as English walnut, Persian walnut, or hú táo in Chinese, is an economically important hardwood tree species cultivated worldwide for its high quality and quantity of wood, nutritious nuts, and traditional medicinal value [26,27,28,29,30]. The PAL gene (JX069977.1) has been cloned in the common walnut as PAL [31] and its expression has been studied [32]. Transcription analysis revealed that JrPAL was expressed in all tested tissues including roots, stems, and leaves, with the highest transcription level found in roots [31]. From June to September, the related JrPAL expression was most pronounced in August, indicating a high synthesis rate of phenolic compounds at this development stage in different varieties of J. regia [33]. In black walnut (J. nigra), PAL genes were also strongly expressed in autumn, suggesting that their transcription in these tissues contributes to phenolic biosynthesis [34]. In a previous study, increases in total phenolics in harvested fresh walnut kernels due to cold stress were accompanied by increases in PAL specific and total activity [32]. These studies show that the PAL gene plays an important role in plant growth and explore its evolutionary mechanism and expression pattern, which lays a foundation for further research on gene function, which is a meaningful perspective for studying the evolutionary mechanism of the PAL gene. However, comprehensive information and functional characterization of the PAL gene family of the common walnut (J. regia) still remain unclear, especially the expression in male and female flowers.
In this study, we systematically characterized 12 PAL genes in the common walnut. We used an iterative process of manual and computational analysis to identify members of the common walnut PAL-encoding gene family within the latest released common walnut whole-genome sequence [29]. We constructed a phylogenetic tree of PAL genes. Conserved motifs, gene structure, protein domain, chromosome localization, and miRNA prediction were further analyzed in common walnut PAL genes. To understand the function of the PAL genes in the common walnut, we also studied the transcriptional-level expression profiles of PALs in male and female flowers at different development stages. Our results provide useful theoretical support for the functional characterization of these PAL genes that are involved in the flower development process in common walnut.
2. Materials and Methods
2.1. Identification of PAL Genes in the Common Walnut (Juglans Regia)
To identify putative PAL gene family members in J. regia, we performed BLASTP (Protein-protein Basic Local Alignment Search Tool) searches in the common walnut genome database, version 1.0 (https://treegenesdb.org/FTP/Genomes/Jure/), with a top E value less than 1 × 10−20 [35]. The available PAL protein sequences were obtained from the following sources: the dicotyledons A. thaliana (At) and P. trichocarpa (Pt), and the monocotyledons Zea mays (Zm) and Oryza sativa (Os) from the Ensembl Plants database (http://archive.plants.ensembl.org/info/website/ftp/index.html) [35]. The walnut whole protein database was downloaded from the National Center for Biotechnology (NCBI) genome plant database (https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk /genome/17683). All walnut PAL potential candidate protein sequences were examined using the Pfam database (http://pfam.xfam.org) [36], the Conserved Domain Database (CDD) (cdd/wrpsb.cgi), and the Simple Modular Architecture Research Tool (SMART) database (http://smart.embl-heidelberg.de/) with an E value cutoff of 1.0 by the domain analysis programs [37]. Sequences lacking PAL–TAL motifs were removed using ClustalX version 2.1 software with default parameters to verify potential common walnut PAL protein candidates by comparing all of the protein sequences with known PAL proteins [38].
2.2. Analysis of Protein Sequence Properties
The PAL sequence name and position information was acquired through BLAST with the parameters E-value < 10−15 and ID % > 50%. [39]. The PAL sequences were predicted on the Plant-mPLoc website to predict subcellular localization of plant proteins, including those with multiple sites [40]. The theoretical isoelectric point and molecular weight were predicted in the ProtParam tool (https://web.expasy.org/protparam/) [41].
2.3. Phylogenetic Analysis
Multiple sequence alignments were generated using ClustalX version 2.1 with default parameters [38]. The evolutionary relationship of J. regia PAL proteins with A. thaliana, Z. mays, O. sativa, and P. trichocarpa PAL proteins was studied using MEGA7 [42]. A phylogenetic tree was constructed using the neighbor-joining (NJ) and maximum likelihood (ML) methods using MEGA version 7.1, the tree topology support was assessed by means of a bootstrap analysis with 1000 replicates, and the phylogenetic tree displayed bootstrap values greater than 50 [42]. The syntenic relationship of PAL genes in the common walnut was conducted using Multiple Collinearity Scan Toolkit (MCScanX) [43]. Initially, potential gene pairs (E-value < 10−5, top 5 matches) obtained by BLASTP across J. regia genomes, were used as input file for MCScanX to analyze segmental and tandem duplications [35,43].
2.4. Conserved Domain and Motifs Displayed in J. Regia PAL Proteins
The motifs were identified using the Multiple EM for Motif Elicitation (MEME) program with default parameters (http://meme-suite.org/) [44]. The parameters were as follows: the maximum number of motifs was set to 20 and the optimum motif width was set to 30–50. We searched each motif sequence of the PAL genes using the SMART database (http://smart.embl-heidelberg.de/) with default parameters. The maximum number of motifs was set to 20, with conserved domains through the NCBI-Batch-CDD software [37].
2.5. Analysis of Gene Exon–Intron Structures
The exon–intron structure of each J. regia PAL gene was confirmed from alignment of the coding sequence (CDS) with the corresponding common walnut genomic sequences through the est2genome program (http://emboss.bioinformatics.nl/cgi-bin/emboss/est2genome). The exon–intron structure was illustrated using the online Gene Structure Display Server program (http://gsds.cbi.pku.edu.cn) by comparing their CDS with genomic sequences of A. thaliana (At) and P. trichocarpa (Pt) by aligning the FASTA-formatted CDS and genomic DNA sequences [1,45,46]. Related walnut gene sequences were searched for on the Genome Browser (https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/genome/).
2.7. Chromosome Location of Common Walnut PAL Genes
The chromosomal locations of J. regia PAL genes were generated using MapInspect software [47] (http://www.softsea.com/download/MapInspect.html) based on the initial position information. The required chromosomal location information was downloaded from the Ensembl Plants database (http://archive.plants.ensembl.org/Triticum_aestivum/Info/Index). To further explore the ways of gene replication, the CDS sequences of the J. regia PAL genes were compared homologously using ClustalX version 2.1 with default settings [38].
2.8. miRNA Predicted in Common Walnut PAL Family Genes
All of the genome sequences of the common walnut PAL family genes were submitted as candidate genes to predict potential miRNAs by searching against the available walnut reference of miRNA sequences using the psRNATarget Server with default parameters [48]. We visualized the interactions between the predicted miRNAs and the corresponding target walnut PAL genes using Cytoscape software with default parameters [49].
2.9. Plant Materials, Treatments, Collections, and RNA Isolation and Analysis of Gene Expression Profiles
To assess the expression of common walnut PAL genes, we collected 12 samples of fresh female and male flowers of individual common walnut trees grown in the Qinling Mountains, which were collected at different development stages (1 to 3 biological replications, on 10, 15, and 22 April, and 1 May), frozen in liquid nitrogen prior to storage at −80 °C until use (Table S1). Total RNA was isolated by an RNA-prep Pure Plant Kit (Tiangen, Beijing, China) [50]. Libraries for RNA sequencing (RNA-seq) were produced using a NEBNext Ultra RNA Library Prep Kit (NEB, Beverly, MA, USA). Paired-end sequencing was performed on the Illumina HiSeq2500 platform to generate 100 bp reads with default parameters by Novogene Bioinformatics Technology Co. Ltd., Beijing, China. The de novo transcriptome was assembled using default settings in Trinity based on the well genome reference of J. regia [29,51]. To further characterize the different temporal and spatial gene expression patterns of the JrPAL gene family, we analyzed RNA-seq data. The transcriptome sequencing datasets were deposited in BioProject under ID PRJNA358784, which was used to perform RNA-seq of different J. regia wild female and male flowers. We analyzed the total RNA-seq data of the female and male flowers at the initial germination flowering stages. We quantified these gene expression levels on the basis of their fragments per kilobase of transcript per million mapped reads (FPKM) values using Cufflinks with default parameters [52], and represented these results using HemI 1.0 software with default parameters [53]. The differential gene expression (DESeq) analysis was performed using the DESeq R package (1.10.1). Genes with an adjusted p-value < 0.05 found by DESeq were assigned as differentially expressed [54]. We normalized the number of reads for the differential gene expression from the RNA-seq data based on a method described by Anders and Huber using the DESeq Bioconductor package with default parameters [55,56,57]. Unigenes were annotated using data from the NCBI Gene Ontology (GO) and Pfam databases. GO annotations were performed in Blast2GO v2.5 with a cutoff E value of 1 × 10−6 [58].
3. Results
3.1. Identification and Characterization of Common Walnut PAL Genes
A total of 12 full-length genes coding putative phenylalanine ammonia-lyase (PAL) were identified in the J. regia genome (Table 1). The 12 sequences containing PAL-HAL domains belong to the PAL gene family. It is evident that these 12 sequences form a family, named the PAL gene family. These walnut PAL proteins ranged in length from 281 to 760 amino acids, with molecular weights from 31.19 kDa to 83.97 kDa and isoelectric points ranging from 5.58 to 8.75. Subcellular localization analysis indicated that all 12 walnut PAL genes are localized in the cytoplasm (Table 1).
3.2. Phylogenetic Relationship of Common Walnut and Other Four Plants in the PAL Gene Family
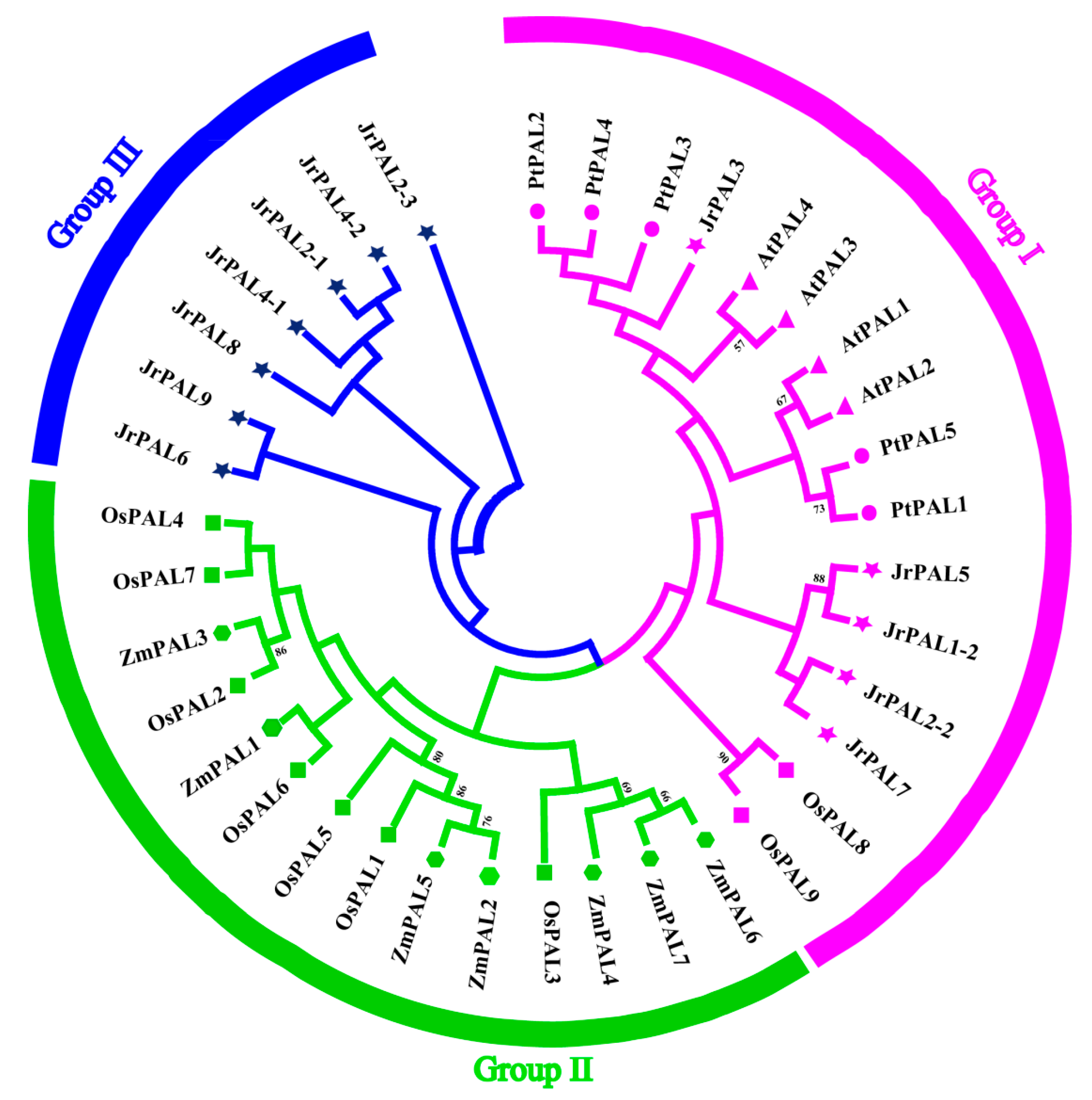
To investigate the lineage-specific expansion of PAL genes in the J. regia genome, we performed a phylogenetic analysis of all PALs from J. regia, A. thaliana proteins, O. sativa proteins, Z. mays proteins, and P. trichocarpa proteins. These plants had 12, 4, 7, 6, and 5 PAL genes, respectively. Based on the completed alignment of the sequences, PALs were clustered into three groups, designated group I, group II, and group III (Figure 1). Overall, group I contained 7 PALs, group II contained 14 PALs, and group III contained 14 PALs (Table S2). Seven PAL genes (JrPAL2-3, JrPAL4-2, JrPAL2-1, JrPAL4-1, JrPAL8, JrPAL9, and JrPAL6) were specifically found in J. regia, and five common walnut PALs (JrPAL3, JrPAL5, JrPAL1-2, JrPAL7, and JrPAL2-2) were found to be closely associated with P. trichocarpa (Figure 1 and Figure S1).
3.3. Position, Conserved Motifs, and Exon–Intron of Common Walnut PALs
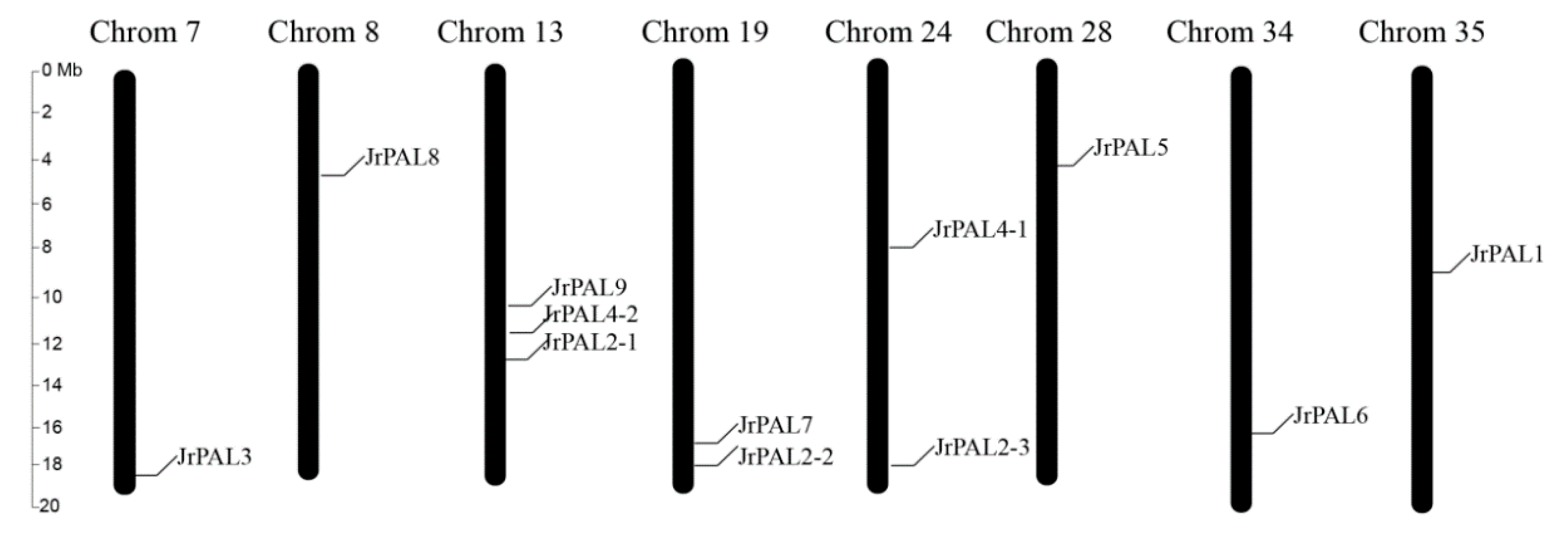
The 12 PAL genes were assigned to eight pseudo-chromosomes of J. regia based on the physical positions (Figure 2; Table 1). The distribution of the PAL genes was different on each pseudo-chromosome. Pseudo-chromosome 13 contained the largest number of PAL genes (3), followed by pseudo chromosome 19 (2 genes) and pseudo-chromosome 24 (2 genes). All other pseudo-chromosomes contained one gene (Figure 2; Table 1). JrPAL3 and JrPAL2-2 is pairs of segmentally duplicated PALs in the J. regia genome. (Figure 2; Table 1).
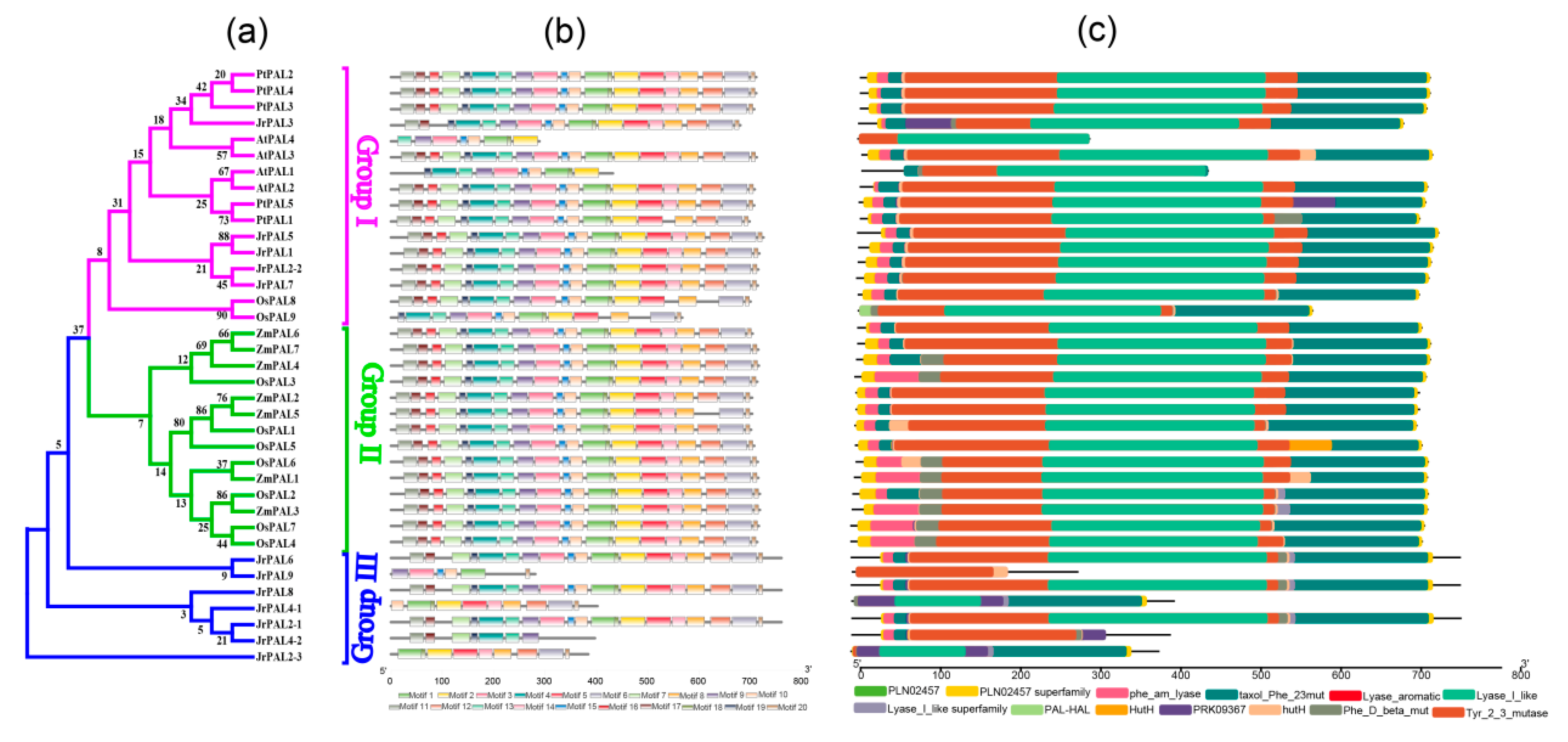
The differences within the PAL family were further analyzed by examining the conserved motifs and domains using the MEME program and the NCBI-CDD database, respectively. After going through the MEME program, the result of the PAL genes was 20 motifs. The predicted walnut PAL gene motifs ranged from 8 to 50 amino acids in length. Motif 1 was mostly present in all species except JrPAL4-2 (Figure 3a,b). Common walnut PALs containing the 13 domains belong to the PAL family, and PRK09367 domains exist in group I and group III; in group II it is a specific domain, and the Lyase_I_like domain is a conserved domain in the three groups. These results suggest that all PAL genes in the walnut contain at least one typical domain (Figure 3b,c).
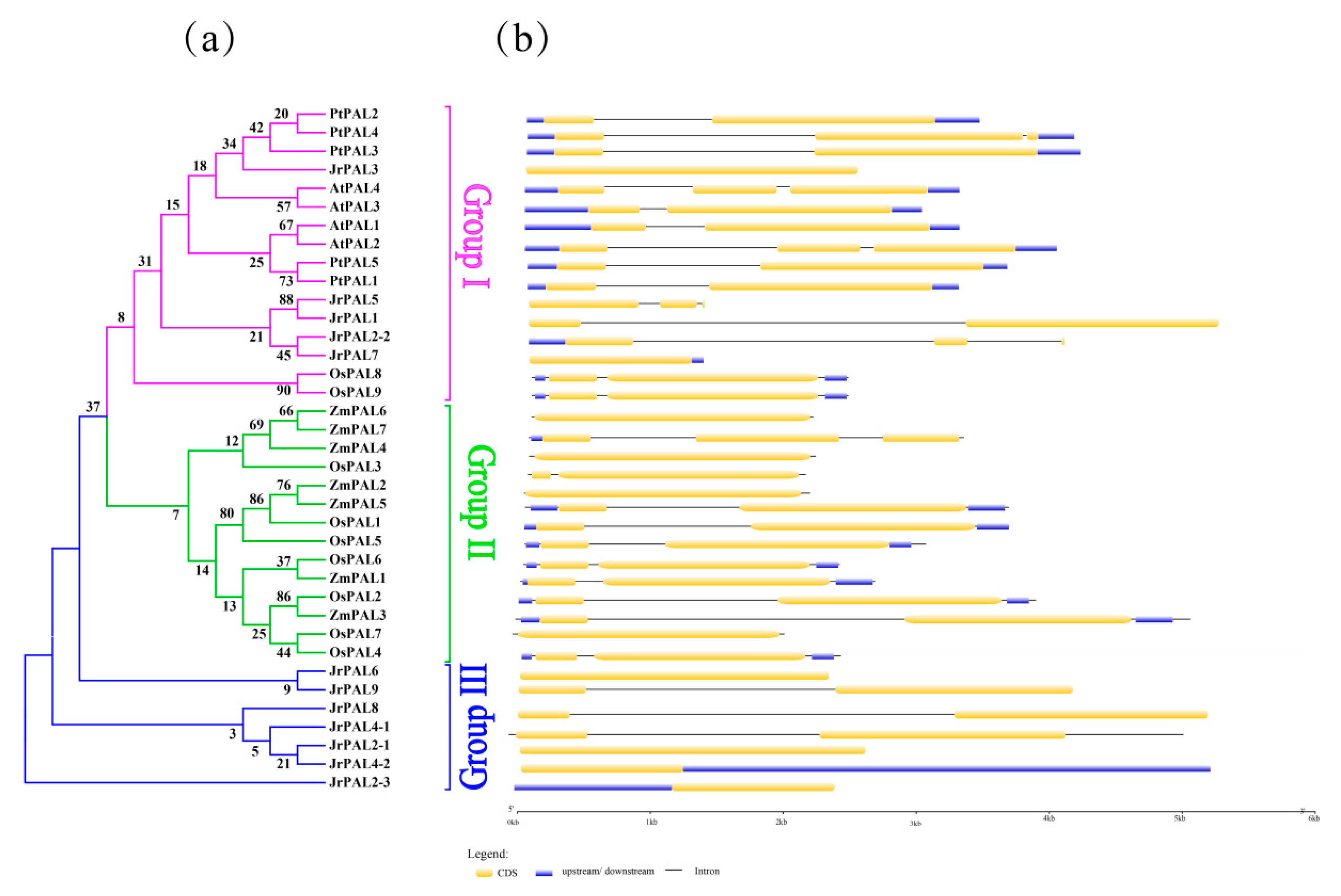
The number of introns per gene varied from one to two based on the exon–intron organization structure analysis of PAL genes in Arabidopsis, common walnut, rice, maize and poplar (Figure 4). The intron positions of orthologous PAL genes in Arabidopsis and poplar and their insertion with symmetric exons are well conserved, indicating that all these PAL genes might have a common ancestor. The structure of PAL genes of common walnut have a different exon–intron organization compared with Arabidopsis and poplar (Figure 4). The exon–intron organization structure of A. thaliana and P. trichocarpa shows an intron insertion in the front, and none exists in J. regia (Figure 4). The gene structure of the PAL gene family in J. regia has an intron in the middle (Figure 4). Overall, the whole gene length of woody plants is longer than that of herbs (Figure 4b).
3.4. MicroRNA Targeting and Expression Profile Analysis of Common Walnut PAL Genes
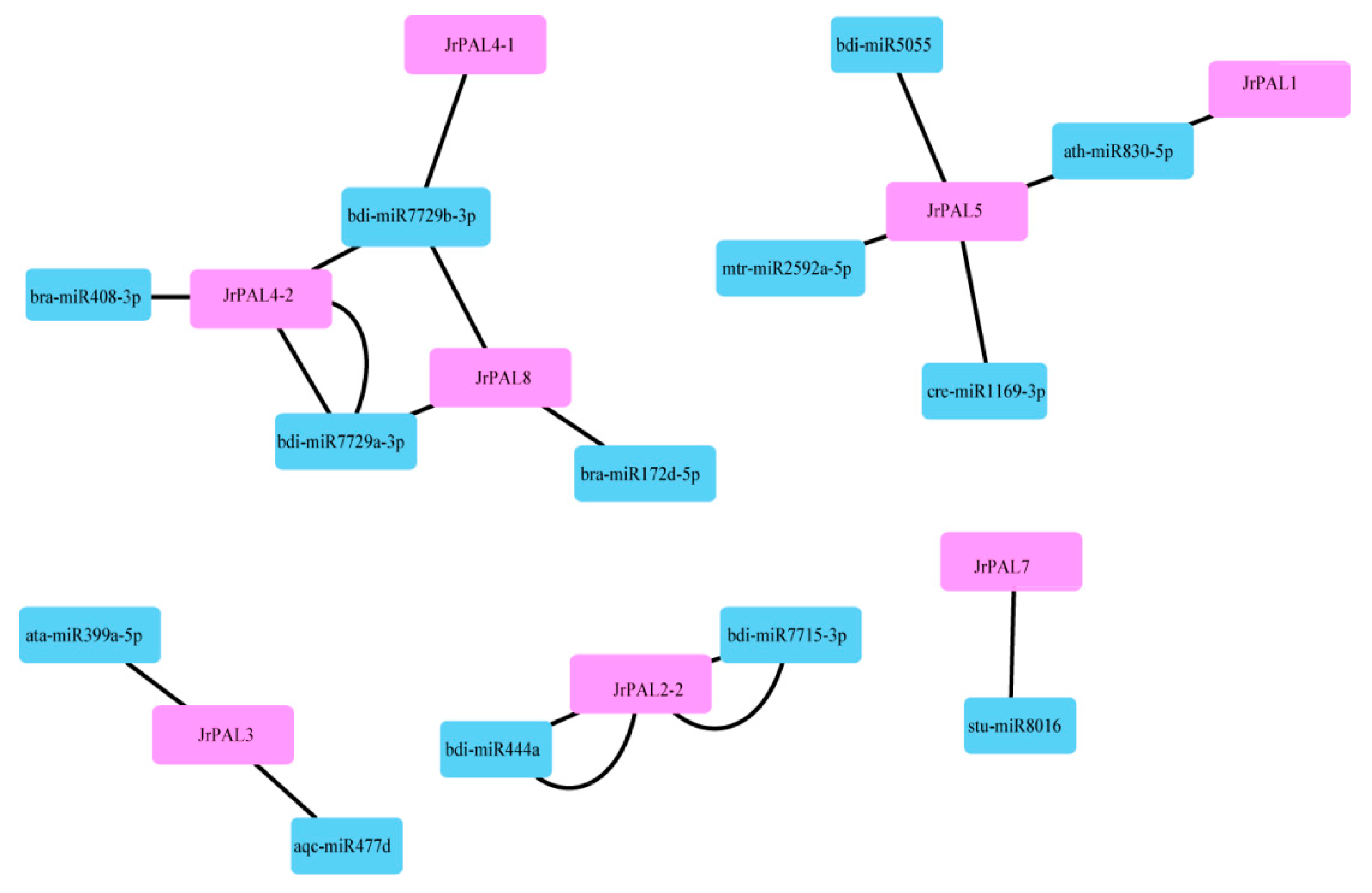
To understand the underlying regulatory mechanism of miRNAs involved in the regulation of PALs, we identified 13 putative miRNAs targeting 12 common walnut PAL genes to construct a relationship network using Cytoscape software (Figure 5; Table S3). We analyzed the connection distribution of the regulation network and found that JrPAL4-2 is one of the most targeted PAL genes of common walnut. The bdi-miRNA 7729b-3p targets walnut genes JrPAL4-1, JrPAL4-2, and JrPAL8, JrPAL5, JrPAL7, JrPAL2-2, JrPAL3 (Figure 5). Our results show that the miRNA ath-miR830-5p targets JrPAL5 and JrPAL1. The two genes JrPAL5 and JrPAL1 have low expression in common walnut flowers (Figure 6). Furthermore, our results also show that the miRNA bdi-miR7729b-3p targeting JrPAL4-1 has high expression in flowers; while three miRNAs (bdi-miR7729b-3p, bdi-miR408-3p, and bdi-miR7729a-3p) targeting JrPAL4-2 have high expression in common walnut flowers (Figure 5 and Figure 6).
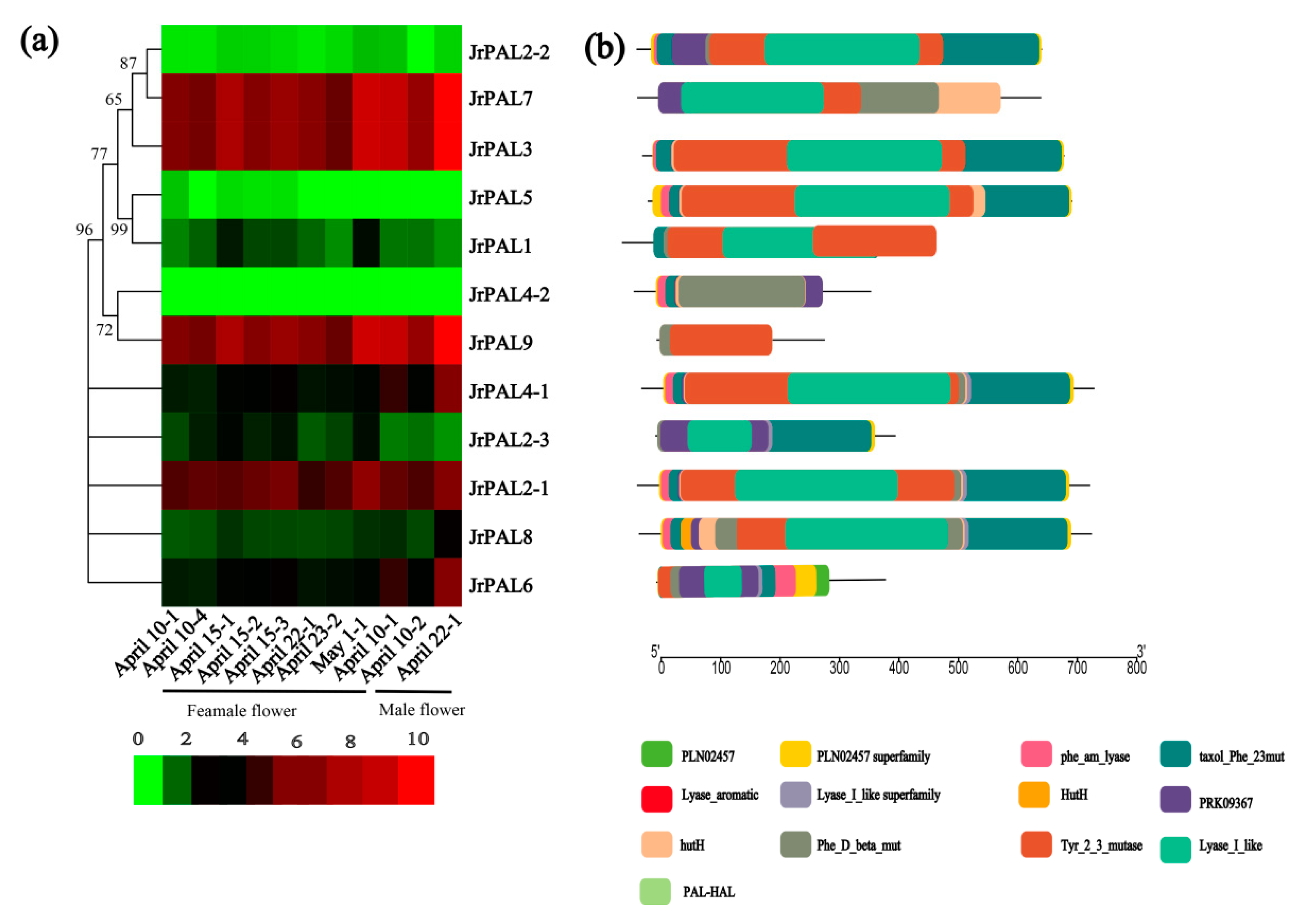
To gain insight into the putative functions of walnut PAL genes, the temporal and spatial expression profiles of the identified PAL genes were examined using the RNA-seq data (Figure 5; Table S4). Four PALs (JrPAL7, JrPAL3, JrPAL9, and JrPAL2-1) had high expression in female and male flowers (Figure 5). We found that the PAL genes with high expression had almost a Tyr_2_3 mutase motif. The expression results showed that the maturity of male flowers had increased expression in all PAL genes (Figure 5). In addition, the phylogenetic relationships are close, and the expression patterns of the two genes are quite different, resulting in JrPAL7, JrPAL3, JrPAL9, and JrPAL2-1 missing motif PRK09367.
4. Discussion
4.1. Characteristics of Phenylalanine Ammonia-Lyase Gene in J. Regia
Phenylalanine ammonia-lyase (PAL) is the first enzyme in the phenylpropanoid pathway [1,27]. PAL plays a critical role in plant growth, development, and adaptation. Recently, there have been reports on the functional analysis of PAL genes in the common walnut [1,27,31,32]. In this study, we found that JrPAL2-2 has a high similarity with JrPAL (JX069977.1) [31] (Table S5). At the molecular level, the identification and isolation of the PAL gene are very important, as the gene was shown to be closely related to stress resistance in previous studies [1,23,24]. Increases in specific and total activity of PAL during cold storage is consistent with the previous study, suggesting that the PAL gene family in the walnut may also have a basic function of resisting stress from cold, salt, and disease [32]. Recently, some studies have shown that the PAL gene is localized to the cytoplasm [13,59,60]. Our results also show that all 12 PAL genes were predicted to be in the cytoplasm of the common walnut (Table 1).
4.2. Comparisons of PAL Gene Family in Plants; Expansion and Evolution of Common Walnut PALs
The PAL gene family members have significant differences among various plant species (Figure 1). We identified 12 PAL genes (JrPAL1-12) in J. regia based on the complete reference common walnut genome sequence (Table 1; Figure 1). Our results show that the number of PAL genes in J. regia far exceeds the four AtPALs in modern A. thaliana, suggesting that genome duplication may have occurred in the evolution of J. regia [15,16]. From JrPAL1 to JrPAL12, despite differences in the genome size and the total number of protein-encoding genes among the sequenced plant species, the number of PAL genes seemed not to increase or decrease proportionally (Table 1) [20]. We searched the PAL genes in 40 common walnut scaffolds. (pseudo-chromosomes) by anchoring the scaffolds to the walnut linkage groups (Table S6) [61]. These 12 PAL genes were localized to eight scaffolds in the common walnut (Table 1; Figure 2) [61]. Duplication events are important in the expansion and evolution of gene families, including whole-genome duplications, small-scale segmental duplications, local tandem duplications, or combinations of these possibilities [62,63,64]. The one gene pair (JrPAL3 and JrPAL2-2) choose the segmental duplicated events in common walnut, which might be caused by gene expansion based on their chromosomal distribution and phylogenetic and syntenic relationships (Figure 2) [20,43].
Based on phylogenetic analysis, our 12 JrPALs were separated into two distinct groups, as PAL genes from cucurbit species A. thaliana [15,16,20], suggesting similar evolutionary trajectories between J. regia and cucurbit species. The phylogenetic tree constructed for PAL genes was divided into four clusters in C. lanatus [65], four clusters in P. taeda [12], and two clusters in most woody plants (S. babylonica, Ornithogalum saundersiae, and P. trichocarpa) [13,14,17] (Figure 1). Motif 1 was only detected in walnut PAL protein sequences compared to other plants (Figure 1 and Figure 3b), indicating that the walnut PAL gene family has a highly conserved protein structure. The PAL genes of these five plants contain all the conserved domains identified in NCBI-Batch-CDD, suggesting that the PAL gene family is extremely conservative in evolution, presuming that PAL genes have important antiretroviral effects. The key domain is phe_am_lyase, which exists in all genes including all species. In group I, JrPAL3, JrPAL5, JrPAL1, JrPAL2-2, and JrPAL7 are distributed in different subclades of the phylogenetic tree in Figure 3c, mainly because of specific PRK09367 domains in the front of the protein structure. JrPAL2-1 and JrPAL4-2, and JrPAL6 and JrPAL9 show quite a difference in protein structure length, but they exist on the same branch of the phylogenetic tree, and we can notice that Tyr_2_3_mutase domains are distributed in the protein structure. These genes (JrPAL2-1, JrPAL4-2, JrPAL6, and JrPAL9) compared to the other genes (JrPAL4-1 and JrPAL2-3) have a clear difference in protein structure, which is the lack of the obvious major domain PRK09367. A phylogenetic analysis of walnut PAL genes shows that they share similar motifs in each subfamily (Figure 3a,b), while motif 1 may be consistent with Tyr_2_3_mutase, so it can be deduced that motif 1 will perform the function of Tyr_2_3_mutase (Figure 3c).
Studying exon–intron gene structure can provide important clues for gene evolution [65]. Genome-wide characterization, molecular cloning, and expression analysis of the structure of PAL genes in walnut and the overall gene structure of Arabidopsis show great differences in exon and intron regions, and one of them showed the same performance (Figure 4a,c) [1]. Compared to the structure of the CiPAL gene in watermelon, the structure of PAL in common walnut has undergone evolutionary changes [66]. There is genetic similarity in A. thaliana and C. lanatus and PAL gene families [65,66]. The information of PAL gene structure in walnut and poplar indicates that the PAL family has undergone major variations in evolution [66] (Figure 4c).
4.3. Comprehensive Analysis of microRNAs Targeting Common Walnut PAL Genes and Expression Levels of PALs in Common Walnut Flowers
In recent years, many studies have shown that miRNAs in plants mainly respond to stress by regulating the expression of genes associated with stress [67]. JrPAL is expressed in roots, shoots, and leaves, but little is known about its expression in flowers [31]. There is no doubt that ath-miR830-5p plays an important role in A. thaliana by targeting two genes with lower expression in flowers than in other tissues; for example, roots and leaves [67,68]. Our experiments of RNA-seq from female and male flowers and miRNA prediction of PAL genes in the common walnut indicated that ath-miR830-5p targeting JrPAL5 and JrPAL1 has low expression in common walnut flowers (Figure 5).
As an entry point to the phenylpropanoid pathway, PAL is tightly regulated at the pre- and post-transcriptional levels. In previous studies, the PAL genes showed different expression patterns in different organs (xylem, roots, leaves, stems, and flowers) in willow, C. sativus, and Salvia miltiorrhiza [11,13,15,16,17,18]. However, there have been few reports on the PAL gene family in common walnut flowers. AtPAL1 and AtPAL2 were shown to be highly expressed in Arabidopsis flowers, while JrPAL3 also showed high expression in female and male flowers of the common walnut (Figure 6) [15]. Furthermore, we found that AtPAL1 and AtPAL2 were clustered into the same group with JrPAL3 based on the phylogenetic tree (Figure 1 and Figure 6). Therefore, it is speculated that the PAL gene is more important at the time of male flower maturation, which can be explained from the side, as the plant becomes more resistant as a mature individual than a young individual (Figure 6b). Different members of gene families generally exhibit disparities in abundance in different tissues or with different stressors [69,70]. This correlation indicates that PALs with similar evolutionary status might play a similar role in plant growth, which allowed us to investigate the functions of PALs from other cucurbits using a comparative genomic approach. The results showed that six genes (JrPAL7, JrPAL3, JrPAL9, JrPAL4-1, JrPAL2-1, and JrPAL6) have high expression in both female and male flowers; another six genes (JrPAL2-2, JrPAL5, JrPAL1, JrPAL4-2, JrPAL2-3, and JrPAL8) have a low expression pattern in both female and male flower tissues. This result of PAL gene family members is due to the different protein structure, gene structure, and microRNA network between the former six genes (high expression) and the latter six genes (low expression) ( Figure 3, Figure 4 and Figure 6). We found that the expression patterns of the PAL gene family in various growing stages of female and male flowers are similar, which shows that they are resistant genes in plant flowering. JrPAL2-2, JrPAL7, JrPAL3, JrPAL5, JrPAL1, JrPAL4-2, JrPAL2-1, JrPAL8, and JrPAL9 have the same expression level at different stages of female and male flowers, but JrPAL6 and JrPAL4-1 have high expression in male flowers compared to female flowers (Figure 6a). Changes in expression levels during the female and male flowering process indicated the important role of PAL genes in sex determination and resistance adaptation. In addition, the expression of JrPAL1 and JrPAL3 was upregulated at the May 1 time point for female flowers, and the expression of JrPAL8 was upregulated at the 22 April time point for male flowers, indicating their different roles during the flowering process. The diverse expression patterns of the PAL family genes indicate a complex regulation of the PAL-mediated phenylpropanoid pathway during the flowering process of the common walnut. The structure of domain Lyase_I_like of JrPAL2-3 was different between female and male flowers. Meanwhile, JrPAL2-3 has high expression in female flowers compared to male flowers, which indicates that the gene domain might affect the pattern of expression of the plant-flowering process (Figure 6). In addition, the microRNAs bra-miR408-3p, bdi-miR7729b-3p, and bdi-miR7729a-3p target the gene JrPAL4-2, and bdi-miR7729b-3p, bra-miR172d-5p, and bdi-miR7729a-3p target the gene JrPAL8 (Figure 5). Moreover, the microRNA ath-miR830-5p targets two genes, JrPAL5 and JrPAL1; however, they have low expression levels in both female and male flowers at the whole growth stage (Figure 5 and Figure 6). With the same pattern, the microRNAs bdi-miR7715-3p and bdi-miR444a both target gene JrPAL2-2, which has low expression in female and male flowers. The two genes JrPAL7 and JrPAL3 have high expression levels among the PAL gene family in the common walnut; JrPAL 7 is targeted by the microRNA stu-miR8016, and JrPAL3 is targeted by two microRNAs, ata-miR399a-5p and aqc-miR477d. The diverse patterns of microRNA-targeted PAL genes indicate that the networks of microRNA may be key regulator network for the PAL gene family in the common walnut.
5. Conclusions
We identified 12 PAL genes in the common walnut. These members are distributed on eight pseudo-chromosomes. The PAL genes were divided into two subfamilies. Seven PALs (JrPAL2-3, JrPAL4-2, JrPAL2-1, JrPAL4-1, JrPAL8, JrPAL9, and JrPAL6) were specifically found in J. regia, and JrPAL3, JrPAL5, JrPAL1-2, JrPAL7, and JrPAL2-2 were found to be closely associated with P. trichocarpa. The exon–intron gene structure of PALs in the common walnut indicated that the PAL family has undergone major variations in evolution compared with Arabidopsis. The relative expression levels of PALs varied in different developmental stages of female and male flowers of common walnut; JrPAL3, JrPAL7, JrPAL9, and JrPAL2-1 were expressed at high levels in most samples. The transcriptional level of JrPAL6 increased in the flowers with the expression of all PALs at different developmental stages. The miRNA ath-miR830-5p targeting JrPAL5 and JrPAL1 has a low expression in common walnut flowers. These findings could lay a theoretical foundation for the functional study of PALs and the further construction of common walnut light regulation networks.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/2073-4425/10/1/46/s1, Figure S1. Phylogenetic analysis of PAL proteins among common walnut (12), Arabidopsis (4), rice (7), maize (6), and poplar (5). These 34 sequences were used to construct a maximum likelihood (ML) tree. Table S1. A total of 12 samples of female and male flowers of common walnut were used for expression profiling in this study. Table S2. Distribution of common walnut PAL genes in Class I, II, and III. Table S3. The putative miRNAs and their targeted JrPAL genes. Table S4. Expression data of PAL genes in flowers at different developmental stages in common walnut (J. regia). Table S5. Kinship between JX069977.1 and JrPAL1-12 in common walnut. Table S6. Information on 40 pseudo-chromosomes for common walnut (J. regia).
Author Contributions
Conceptualization, P.Z. and F.Y.; Conceived and designed the experiments: P.Z. and F.Y.; Data curation, P.Z., H.L., and F.Y.; Analyzed the data: P.Z., H.L., and F.Y.; Funding acquisition, P.Z., Project administration, P.Z.; Contributed materials/analysis tools: P.Z., H.L., and F.Y.; Wrote original draft, P.Z., H.L., and F.Y.; Writing—review and editing, P.Z., H.L, and F.Y.
Funding
This research was funded by the National Natural Science Foundation of China (no. 41471038), the Program for Excellent Young Academic Backbones funding by Northwest University (grant no. 338050070), and the Northwest University Training Programs of Innovation and Entrepreneurship for Graduates (grant nos. 2017037 and 201807075).
Acknowledgments
We thank Huijuan Zhou, Meng Dang, and Yiheng Hu for sample collection.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Huang, J.; Gu, M.; Lai, Z.; Fan, B.; Shi, K.; Zhou, Y.H.; Yu, J.Q.; Chen, Z. Functional analysis of the Arabidopsis PAL gene family in plant growth, development, and response to environmental stress. Plant Physiol. 2010, 153, 1526–1538. [Google Scholar] [CrossRef]
- Wu, P.; Guo, Q.Q.; Qin, Z.W. The fungicide propamocarb increases lignin by activating the phenylpropanoid pathway in Cucumis sativus L. Hortic. Environ. Biotechnol. 2016, 57, 511–518. [Google Scholar] [CrossRef]
- Dixon, R.A.; Paiva, N.L. Stress-induced phenylpropanoid metabolism. Plant Cell 1995, 7, 1085. [Google Scholar] [CrossRef]
- Camera, S.L.; Gouzerh, G.; Dhondt, S.; Hoffmann, L.; Fritig, B.; Legrand, M.; Heitz, T. Metabolic reprogramming in plant innate immunity: The contributions of phenylpropanoid and oxylipin pathways. Immunol. Rev. 2010, 198, 267–284. [Google Scholar] [CrossRef]
- Vogt, T. Phenylpropanoid biosynthesis. Mol. Plant 2010, 3, 2–20. [Google Scholar] [CrossRef] [PubMed]
- Koukol, J.; Conn, E.E. The metabolism of aromatic compounds in higher plants. IV. Purification and properties of the phenylalanine deaminase of Hordeum vulgare. J. Boil. Chem. 1966, 236, 2692–2698. [Google Scholar]
- Lois, R.; Dietrich, A.; Hahlbrock, K.; Schulz, W. A phenylalanine ammonia-lyase gene from parsley: Structure, regulation and identification of elicitor and light responsive cis-acting elements. EMBO J. 1989, 8, 1641–1648. [Google Scholar] [CrossRef] [PubMed]
- Macdonald, M.J.; D’Cunha, G.B. A modern view of phenylalanine ammonia lyase. Biochem. Cell Biol. 2007, 85, 273. [Google Scholar] [CrossRef]
- Cochrane, F.C.; Davin, L.B.; Lewis, N.G. The Arabidopsis phenylalanine ammonia lyase gene family: Kinetic characterization of the four PAL isoforms. Phytochemistry 2004, 65, 1557–1564. [Google Scholar] [CrossRef] [PubMed]
- Rawal, H.C.; Singh, N.K.; Sharma, T.R. Conservation, divergence, and genome-wide distribution of PAL and POX A gene families in plants. Int. J. of Genom. 2013, 2013, 678–969. [Google Scholar]
- Hou, X.; Shao, F.; Ma, Y.; Lu, S. The phenylalanine ammonia-lyase gene family in Salvia miltiorrhiza: Genome-wide characterization, molecular cloning and expression analysis. Mol. Biol. Rep. 2013, 40, 4301–4310. [Google Scholar] [CrossRef] [PubMed]
- Bagal, U.R. The phenylalanine ammonia lyase (PAL) gene family shows a gymnosperm-specific lineage. BMC Genom. 2012, 13, S1. [Google Scholar]
- De, J.F.; Hanley, S.J.; Beale, M.H.; Karp, A. Characterisation of the willow phenylalanine ammonia-lyase (PAL) gene family reveals expression differences compared with poplar. Phytochemistry 2015, 117, 90–97. [Google Scholar]
- Wang, Z.B.; Chen, X.; Wang, W.; Cheng, K.D.; Kong, J.Q. Transcriptome-wide identification and characterization of Ornithogalum saundersiae phenylalanine ammonia lyase gene family. RSC Adv. 2014, 4, 27159–27175. [Google Scholar] [CrossRef]
- Wanner, L.A.; Li, G.; Ware, D.; Somssich, I.E.; Davis, K.R. The phenylalanine ammonia-lyase gene family in Arabidopsis thaliana. Plant Mol. Biol. 1995, 27, 327–338. [Google Scholar] [CrossRef] [PubMed]
- Raes, J.; Rohde, A.; Christensen, J.H.; Van, Y.D.P.; Boerjan, W. Genome-wide characterization of the lignification toolbox in Arabidopsis. Plant Physiol. 2003, 133, 1051–1071. [Google Scholar] [CrossRef] [PubMed]
- Shi, R.; Sun, Y.H.; Li, Q.; Heber, S.; Sederoff, R.; Chiang, V.L. Towards a systems approach for lignin biosynthesis in Populus trichocarpa: Transcript abundance and specificity of the monolignol biosynthetic genes. Plant Cell Physiol. 2010, 51, 144–163. [Google Scholar] [CrossRef]
- Hui, X.; Park, N.I.; Li, X.; Yong, K.K.; Lee, S.Y.; Sang, U.P. Molecular cloning and characterization of phenylalanine ammonia-lyase, cinnamate 4-hydroxylase and genes involved in flavone biosynthesis in Scutellaria baicalensis. Bioresour. Technol. 2010, 101, 9715–9722. [Google Scholar]
- Lepelley, M.; Mahesh, V.; Mccarthy, J.; Rigoreau, M.; Crouzillat, D.; Chabrillange, N.; De, K.A.; Campa, C. Characterization, high-resolution mapping and differential expression of three homologous PAL genes in Coffea canephora pierre (Rubiaceae). Planta 2012, 236, 313–326. [Google Scholar] [CrossRef] [PubMed]
- Dong, C.J.; Ning, C.; Zhang, Z.G.; Shang, Q.M. Phenylalanine ammonia-lyase gene families in cucurbit species: Structure, evolution, and expression. J. Integr. Agric. 2016, 15, 1239–1255. [Google Scholar] [CrossRef]
- Shang, Q.M.; Li, L.; Dong, C.J. Multiple tandem duplication of the phenylalanine ammonia-lyase genes in Cucumis sativus L. Planta 2012, 236, 1093–1105. [Google Scholar] [CrossRef] [PubMed]
- Jaillon, O.; Aury, J.M.; Noel, B.; Policriti, A.; Clepet, C.; Casagrande, A.; Choisne, N.; Aubourg, S.; Vitulo, N.; Jubin, C. The grapevine genome sequence suggests ancestral hexaploidization in major angiosperm phyla. Nature 2007, 449, 463. [Google Scholar] [PubMed]
- Wada, K.C.; Mizuuchi, K.; Koshio, A.; Kaneko, K.; Mitsui, T.; Takeno, K. Stress enhances the gene expression and enzyme activity of phenylalanine ammonia-lyase and the endogenous content of salicylic acid to induce flowering in pharbitis. J. Plant Physiol. 2014, 171, 895–902. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.S.; Hwang, B.K. An important role of the pepper phenylalanine ammonia-lyase gene (PAL1) in salicylic acid-dependent signalling of the defence response to microbial pathogens. J. Exp. Bot. 2014, 65, 2295. [Google Scholar] [CrossRef] [PubMed]
- Fang, C.; Zhuang, Y.; Xu, T.; Li, Y.; Li, Y.; Lin, W. Changes in rice allelopathy and rhizosphere microflora by inhibiting rice phenylalanine ammonia-lyase gene expression. J. Chem. Ecol. 2013, 39, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Pollegioni, P.; Woeste, K.E.; Chiocchini, F.; Lungo, S.D.; Olimpieri, I.; Tortolano, V.; Clark, J.; Hemery, G.E.; Mapelli, S.; Malvolti, M.E. Ancient humans influenced the current spatial genetic structure of common walnut populations in Asia. PLoS ONE 2015, 10, e0135980. [Google Scholar] [CrossRef] [PubMed]
- Han, H.; Woeste, K.E.; Hu, Y.; Dang, M.; Zhang, T.; Gao, X.X.; Zhou, H.; Feng, X.; Zhao, G.; Zhao, P. Genetic diversity and population structure of common walnut (Juglans regia) in China based on EST-SSRs and the nuclear gene phenylalanine ammonia-lyase (PAL). Tree Genet. Genomes 2016, 12, 111. [Google Scholar] [CrossRef]
- Panth, N.; Paudel, K.R.; Karki, R. Phytochemical profile and biological activity of Juglans regia. Chin J. Integr. Med. 2016, 14, 359–373. [Google Scholar] [CrossRef]
- Martínez-García, P.J.; Crepeau, M.W.; Puiu, D.; Gonzalez-Ibeas, D.; Whalen, J.; Stevens, K.A.; Paul, R.; Butterfield, T.S.; Britton, M.T.; Reagan, R.L.; et al. The walnut (Juglans regia) genome sequence reveals diversity in genes coding for the biosynthesis of non-structural polyphenols. Plant J. 2016, 87, 507–532. [Google Scholar] [CrossRef]
- Tsoukas, M.A.; Ko, B.J.; Witte, T.R.; Dincer, F.; Hardman, W.E.; Mantzoros, C.S. Dietary walnut suppression of colorectal cancer in mice: Mediation by miRNA patterns and fatty acid incorporation. J. Nutr. Biochem. 2015, 26, 776–783. [Google Scholar] [CrossRef]
- Xu, F.; Deng, G.; Cheng, S.; Zhang, W.; Huang, X.; Li, L.; Cheng, H.; Rong, X.; Li, J. Molecular cloning, characterization and expression of the phenylalanine ammonia-lyase gene from Juglans regia. Molecules 2012, 17, 7810–7823. [Google Scholar] [CrossRef] [PubMed]
- Christopoulos, M.V.; Tsantili, E. Participation of phenylalanine ammonia-lyase (PAL) in increased phenolic compounds in fresh cold stressed walnut (Juglans regia L.) kernels. Postharvest Biol. Technol. 2015, 104, 17–25. [Google Scholar] [CrossRef]
- Persic, M.; Mikulicpetkovsek, M.; Halbwirth, H.; Solar, A.; Veberic, R.; Slatnar, A. Red walnut: Characterization of the phenolic profiles, activities and gene expression of selected enzymes related to the phenylpropanoid pathway in pellicle during walnut development. J. Agric. Food Chem. 2018, 66, 2742–2748. [Google Scholar] [CrossRef]
- Beritognolo, I.; Magel, E.; Abdellatif, A.; Charpentier, J.P.; Jayallemand, C.; Breton, C. Expression of genes encoding chalcone synthase, flavanone 3-hydroxylase and dihydroflavonol 4-reductase correlates with flavanol accumulation during heartwood formation in Juglans nigra. Tree Physiol. 2002, 22, 291. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Meng, D.; Abdullah, M.; Jin, Q.; Lin, Y.; Cai, Y. Genome wide identification, evolutionary, and expression analysis of VQ genes from two Pyrus species. Genes 2018, 9, 224. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Penelope, C.; Eberhardt, R.Y.; Eddy, S.R.; Jaina, M.; Mitchell, A.L.; Potter, S.C.; Marco, P.; Matloob, Q.; Amaia, S.V. The Pfam protein families database: Towards a more sustainable future. Nucleic Acids Res. 2016, 44, D279–D285. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Doerks, T.; Bork, P. Smart 7: Recent updates to the protein domain annotation resource. Nucleic Acids Res. 2012, 40, 302–305. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. Muscle: A multiple sequence alignment method with reduced time and space complexity. BMC Bioinform. 2004, 5, 113. [Google Scholar] [CrossRef] [PubMed]
- Lobo, I. Basic local alignment search tool (BLAST). J. Mol. Biol. 2012, 215, 403–410. [Google Scholar]
- Chou, K.C.; Shen, H.B. Plant-mPLoc: A top-down strategy to augment the power for predicting plant protein subcellular localization. PLoS ONE 2010, 5, e11335. [Google Scholar] [CrossRef]
- Ma, M.; Ming, L.; Jian, C.; Song, Y.; Wang, S.; Li, P. Molecular and biological characterization of Chinese Sacbrood virus LN isolate. Comp. Funct. Genom. 2011, 2011, 386–409. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tang, H.; DeBarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.-H.; Jin, H.; Marler, B.; Guo, H. Mcscanx: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef]
- Bailey, T.L.; Johnson, J.; Grant, CE.; Noble, W.S. The MEME suite. Nucleic Acids Res. 2015, 43, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.J.; Harding, S.A.; Tschaplinski, T.J.; Lindroth, R.L.; Yuan, Y. Genome-wide analysis of the structural genes regulating defense phenylpropanoid metabolism in Populus. New Phytol. 2010, 172, 47–62. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Jin, J.; Guo, A.Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2014, 31, 1296–1297. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Li, W.; He, Q.; Daud, M.K.; Chen, J.; Zhu, S. Characterization of 19 genes encoding membrane-bound fatty acid desaturases and their expression profiles in Gossypium raimondii under low temperature. PLoS ONE 2015, 10, e0123281. [Google Scholar] [CrossRef]
- Dai, X.; Zhao, P.X. psRNATarget: A plant small RNA target analysis server. Nucleic Acids Res. 2011, 39, W155–W159. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Zhu, H.; Han, X.; Lv, J.; Zhao, L.; Xu, X.; Zhang, T.; Guo, W. Structure, expression differentiation and evolution of duplicated fiber developmental genes in Gossypium barbadense and G. hirsutum. BMC Plant Biol. 2011, 11, 11–40. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q. Full-length transcriptome assembly from RNA-seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Roberts, A.; Goff, L.A.; Pertea, G.; Kim, D.; Kelley, D.R.; Pimentel, H.; Salzberg, S.L.; Rinn, J.L.; Pachter, L. Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks. Nat. Protoc. 2012, 7, 562–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, W.; Wang, Y.; Liu, Z.; Cheng, H.; Xue, Y. HEMI: A toolkit for illustrating heatmaps. PLoS ONE 2014, 9, e111988. [Google Scholar] [CrossRef]
- Likun, W.; Zhixing, F.; Xi, W.; Xiaowo, W.; Xuegong, Z. Degseq: An R package for identifying differentially expressed genes from RNA-seq data. Bioinformatics 2010, 26, 136–138. [Google Scholar]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010, 11, R106. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Piao, Y.; Shon, H.S.; Ryu, K.H. Comparing the normalization methods for the differential analysis of Illumina high-throughput RNA-seq data. BMC Bioinform. 2015, 16, 347. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Love, M.I.; Anders, S.; Huber, W. DESeq2: Differential gene expression analysis based on the negative binomial distribution. 2014. Available online: https://rdrr.io/bioc/DESeq2/ (accessed on 24 September 2018).
- Conesa, A.; Götz, S.; Garcíagómez, J.M.; Terol, J.; Talón, M.; Robles, M. Blast2go: A universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 2005, 21, 3674–3676. [Google Scholar] [CrossRef]
- Achnine, L.; Blancaflor, E.B.; Rasmussen, S.; Dixon, R.A. Colocalization of l-phenylalanine ammonia-lyase and cinnamate 4-hydroxylase for metabolic channeling in phenylpropanoid biosynthesis. Plant Cell 2004, 16, 3098–3109. [Google Scholar] [CrossRef]
- Ma, R.F.; Liu, Q.Z.; Xiao, Y.; Zhang, L.; Li, Q.; Yin, J.; Chen, W.S. The phenylalanine ammonia-lyase gene family in Isatis indigotica fort.: Molecular cloning, characterization, and expression analysis. Chin. J. Nat. Med. 2016, 14, 801–812. [Google Scholar] [CrossRef]
- Lei, L.; Zhou, S.L.; Ma, H.; Zhang, L.S. Expansion and diversification of the set domain gene family following whole-genome duplications in Populus trichocarpa. BMC Evol. Biol. 2012, 12, 51. [Google Scholar] [CrossRef] [PubMed]
- Xue, Z.; Duan, L.; Liu, D.; Guo, J.; Ge, S.; Dicks, J.; ÓMáille, P.; Osbourn, A.; Qi, X. Divergent evolution of oxidosqualene cyclases in plants. New Phytol. 2012, 193, 1022–1038. [Google Scholar] [CrossRef] [PubMed]
- Ober, D. Seeing double: Gene duplication and diversification in plant secondary metabolism. Trends Plant Sci. 2005, 10, 444–449. [Google Scholar] [CrossRef] [PubMed]
- Freeling, M. Bias in plant gene content following different sorts of duplication: Tandem, whole-genome, segmental, or by transposition. Annu. Rev. Plant Biol. 2009, 60, 433–453. [Google Scholar] [CrossRef] [PubMed]
- Dong, C.J.; Shang, Q.M. Genome-wide characterization of phenylalanine ammonia-lyase gene family in watermelon (Citrullus lanatus). Planta 2013, 238, 35–49. [Google Scholar] [CrossRef] [PubMed]
- Li, W.X.; Oono, Y.; Zhu, J.; He, X.J.; Wu, J.M.; Iida, K.; Lu, X.Y.; Cui, X.; Jin, H.; Zhu, J.K. The Arabidopsis NFYA5 transcription factor is regulated transcriptionally and posttranscriptionally to promote drought resistance. Plant Cell 2008, 20, 2238–2251. [Google Scholar] [CrossRef] [PubMed]
- Borges, F.; Pereira, P.A.; Slotkin, R.K.; Martienssen, R.A.; Becker, J.D. MicroRNA activity in the Arabidopsis male germline. J. Exp. Bot. 2011, 62, 3699. [Google Scholar] [CrossRef] [PubMed]
- Jeong, D.H.; Green, P.J. Comprehensive investigation of microRNAs enhanced by analysis of sequence variants, expression patterns, argonaute loading, and target cleavage. Plant Physiol. 2013, 162, 1225–1245. [Google Scholar] [CrossRef] [PubMed]
- Baranwal, V.K.; Negi, N.; Khurana, P. Auxin response factor genes repertoire in Mulberry: Identification, and structural, functional and evolutionary analyses. Genes 2017, 8, 202. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Phylogenetic analysis of phenylalanine ammonia-lyase (PAL) proteins among common walnut (12), Arabidopsis (4), rice (7), maize (6), and poplar (5). These 34 sequences were used to construct a neighbor-joining (NJ) tree. The tree was divided into three groups, represented by different colors. Triangles represent Arabidopsis, rectangles represent rice, pentagons represent maize, circles represent poplar, and pentagrams represent walnut. The number on each node of branch indicates the bootstrap support value more than 50.
Figure 1.
Phylogenetic analysis of phenylalanine ammonia-lyase (PAL) proteins among common walnut (12), Arabidopsis (4), rice (7), maize (6), and poplar (5). These 34 sequences were used to construct a neighbor-joining (NJ) tree. The tree was divided into three groups, represented by different colors. Triangles represent Arabidopsis, rectangles represent rice, pentagons represent maize, circles represent poplar, and pentagrams represent walnut. The number on each node of branch indicates the bootstrap support value more than 50.

Figure 2.
Chromosome locations of PAL genes of common walnut on 40 scaffolds (pseudo-chromosomes). The pseudo-chromosome name is at the top of each bar. The scale of the pseudo-chromosomes is millions of base pairs (Mb).
Figure 2.
Chromosome locations of PAL genes of common walnut on 40 scaffolds (pseudo-chromosomes). The pseudo-chromosome name is at the top of each bar. The scale of the pseudo-chromosomes is millions of base pairs (Mb).

Figure 3.
(a) Phylogenetic relationships, (b) motif compositions, and (c) conserved domains of the 12 PAL genes identified in the common walnut. Phylogenetic relationships used the neighbor-joining (NJ) method, and different colors represent different groups. Colored boxes indicate conserved motifs and gray lines represent nonconserved sequences. The lengths of motifs in each protein are shown proportionally.
Figure 3.
(a) Phylogenetic relationships, (b) motif compositions, and (c) conserved domains of the 12 PAL genes identified in the common walnut. Phylogenetic relationships used the neighbor-joining (NJ) method, and different colors represent different groups. Colored boxes indicate conserved motifs and gray lines represent nonconserved sequences. The lengths of motifs in each protein are shown proportionally.

Figure 4.
(a) Phylogenetic relationships and (b) gene structures of PAL genes in Arabidopsis, poplar, rice, maize, and common walnut. CDS, upstream/downstream, introns, and intron insertion are shown. Orange boxes indicate coding sequence, blue boxes indicate upstreams or downstreams genes, and gray lines represent introns.
Figure 4.
(a) Phylogenetic relationships and (b) gene structures of PAL genes in Arabidopsis, poplar, rice, maize, and common walnut. CDS, upstream/downstream, introns, and intron insertion are shown. Orange boxes indicate coding sequence, blue boxes indicate upstreams or downstreams genes, and gray lines represent introns.

Figure 5.
A schematic representation of the regulatory network relationships between the putative miRNAs and their targeted walnut PAL genes.
Figure 5.
A schematic representation of the regulatory network relationships between the putative miRNAs and their targeted walnut PAL genes.

Figure 6.
(a) Phylogenetic relationships and expression patterns and (b) conserved motif compositions of the 12 PAL genes in walnut. The phylogenetic tree was constructed based on full-length protein sequences using MEGA6.0 (http://web.megasoftware.net/) with hierarchical clustering of the relative expression levels of PAL genes. RNA-seq data of female and male flowers in common walnut were used to analyze expression patterns. The heat map was drawn in log10-transformed expression values. Red and green represent decreased and increased expression levels in each sample, respectively; 1 represents one biological replication; 2 represents two biological replications; 3 represents three biological replications; 4 represents four replications. The Multiple EM for Motif Elicitation (MEME) program was used to predict conserved motifs. Each motif is represented by a different colored box. Heat map shows expression patterns of walnut PAL family genes in six stages. Red and green represent relatively high and low expression compared to control, respectively.
Figure 6.
(a) Phylogenetic relationships and expression patterns and (b) conserved motif compositions of the 12 PAL genes in walnut. The phylogenetic tree was constructed based on full-length protein sequences using MEGA6.0 (http://web.megasoftware.net/) with hierarchical clustering of the relative expression levels of PAL genes. RNA-seq data of female and male flowers in common walnut were used to analyze expression patterns. The heat map was drawn in log10-transformed expression values. Red and green represent decreased and increased expression levels in each sample, respectively; 1 represents one biological replication; 2 represents two biological replications; 3 represents three biological replications; 4 represents four replications. The Multiple EM for Motif Elicitation (MEME) program was used to predict conserved motifs. Each motif is represented by a different colored box. Heat map shows expression patterns of walnut PAL family genes in six stages. Red and green represent relatively high and low expression compared to control, respectively.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Phenylalanine ammnia-lyase (PAL) gene family information in common walnut (J. regia).
| Gene Name | Protein ID | Gene ID | CDS ID | Subcellular Location | Amino Acids (aa) | Molecular Weight (kDa) | Theoretical pI | Chromosome | Chromosome Length | Gene Position | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Start | End | ||||||||||
| JrPAL3 | XP_018828772.1 | NW_017443600.1:c859630-855244 | XM_018973227.1 | Cytoplasm | 708 | 77.04 | 5.96 | Chr7 | 19,001,705 | 18,431,863 | 1,8427,477 |
| JrPAL5 | XP_018859391.1 | NW_017437924.1:10925-15347 | XM_019003846.1 | Cytoplasm | 712 | 77.71 | 6.06 | Chr28 | 18,296,634 | 2,940,954 | 2,945,376 |
| JrPAL2-2 | XP_018844813.1 | NW_017389549.1:c62915-57410 | XM_018989268.1 | Cytoplasm | 680 | 74.11 | 5.91 | Chr19 | 18,508,379 | 1,8341,787 | 18,337,338 |
| JrPAL2-1 | XP_018827035.1 | NW_017443587.1:471936-474695 | XM_018971490.1 | Cytoplasm | 760 | 83.67 | 6.16 | Chr13 | 18,490,500 | 1,2030,270 | 12,033,029 |
| JrPAL4-1 | XP_018845411.1 | NW_017389589.1:c87043-84575 | XM_018989866.1 | Cytoplasm | 760 | 83.92 | 6.38 | Chr24 | 18,356,466 | 3,643,363 | 3,640,895 |
| JrPAL8 | XP_018817312.1 | NW_017443031.1:261176-263621 | XM_018961767.1 | Cytoplasm | 760 | 83.97 | 6.35 | Chr8 | 19,363,960 | 5,898,473 | 5,900,918 |
| JrPAL1 | XP_018853318.1 | NW_017394290.1:c1301-6 | XM_018997773.1 | Cytoplasm | 432 | 47.05 | 6.57 | Chr35 | 18,286,198 | 9,845,144 | 9,843,849 |
| JrPAL2-3 | XP_018845408.1 | NW_017389589.1:c6632-4075 | XM_018989863.1 | Cytoplasm | 402 | 44.94 | 5.74 | Chr24 | 18,356,466 | 3,562,298 | 3,560,395 |
| JrPAL4-2 | XP_018827000.1 | NW_017443587.1:336894-338297 | XM_018971455.1 | Cytoplasm | 397 | 44.11 | 8.75 | Chr13 | 18,490,500 | 11,895,228 | 11,896,204 |
| JrPAL6 | XP_018855337.1 | NW_017419648.1:c5073-3779 | XM_018989863.1 | Cytoplasm | 384 | 42.92 | 5.85 | Chr34 | 18,288,579 | 16,505,050 | 16,503,756 |
| JrPAL7 | AAX18624.1 | NW_017389549.1:c62915-57410 | XM_018989268.1 | Cytoplasm | 289 | 31.19 | 5.58 | Chr19 | 18,508,379 | 18,341,787 | 18,337,338 |
| JrPAL9 | XP_018827002.1 | NW_017443587.1:394996-399270 | XM_018971457.1 | Cytoplasm | 281 | 31.47 | 6.40 | Chr13 | 18,490,500 | 11,954,327 | 11,956,842 |
Note: Protein ID, Gene ID, and CDS (coding sequence) ID indicate that the accession number of the PAL gene family member sequences were downloaded from the National Center for Biotechnology (NCBI). kDa indicates that kilodaltons (unified atomic mass unit), pI indicates that isoelectric point.
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Yan, F.; Li, H.; Zhao, P. Genome-Wide Identification and Transcriptional Expression of the PAL Gene Family in Common Walnut (Juglans Regia L.). Genes 2019, 10, 46. https://0-doi-org.brum.beds.ac.uk/10.3390/genes10010046
AMA Style
Yan F, Li H, Zhao P. Genome-Wide Identification and Transcriptional Expression of the PAL Gene Family in Common Walnut (Juglans Regia L.). Genes. 2019; 10(1):46. https://0-doi-org.brum.beds.ac.uk/10.3390/genes10010046
Chicago/Turabian StyleYan, Feng, Huaizhu Li, and Peng Zhao. 2019. "Genome-Wide Identification and Transcriptional Expression of the PAL Gene Family in Common Walnut (Juglans Regia L.)" Genes 10, no. 1: 46. https://0-doi-org.brum.beds.ac.uk/10.3390/genes10010046
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.