A Comparison Between Two Assays for Measuring Seminal Oxidative Stress and their Relationship with Sperm DNA Fragmentation and Semen Parameters

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Semen Samples
2.3. MeasuringReactive Oxygen Species Using Chemiluminescence
2.4. Measuring Oxidation-Reduction Potential Using MiOXSYS
2.5. Measuring Sperm DNA Fragmentation
2.6. Statistical Analysis
3. Results
3.1. Correlation between OS and Sperm DNA Damage; Comparison between Two Methods of OS Measurement
3.2. Sperm DNA Damage and HDS Levels in Oxidative Balanced versus Oxidative Stressed Semen Samples
3.3. Correlation between Oxidative Stress, Sperm DNA Damage and Semen Parameters
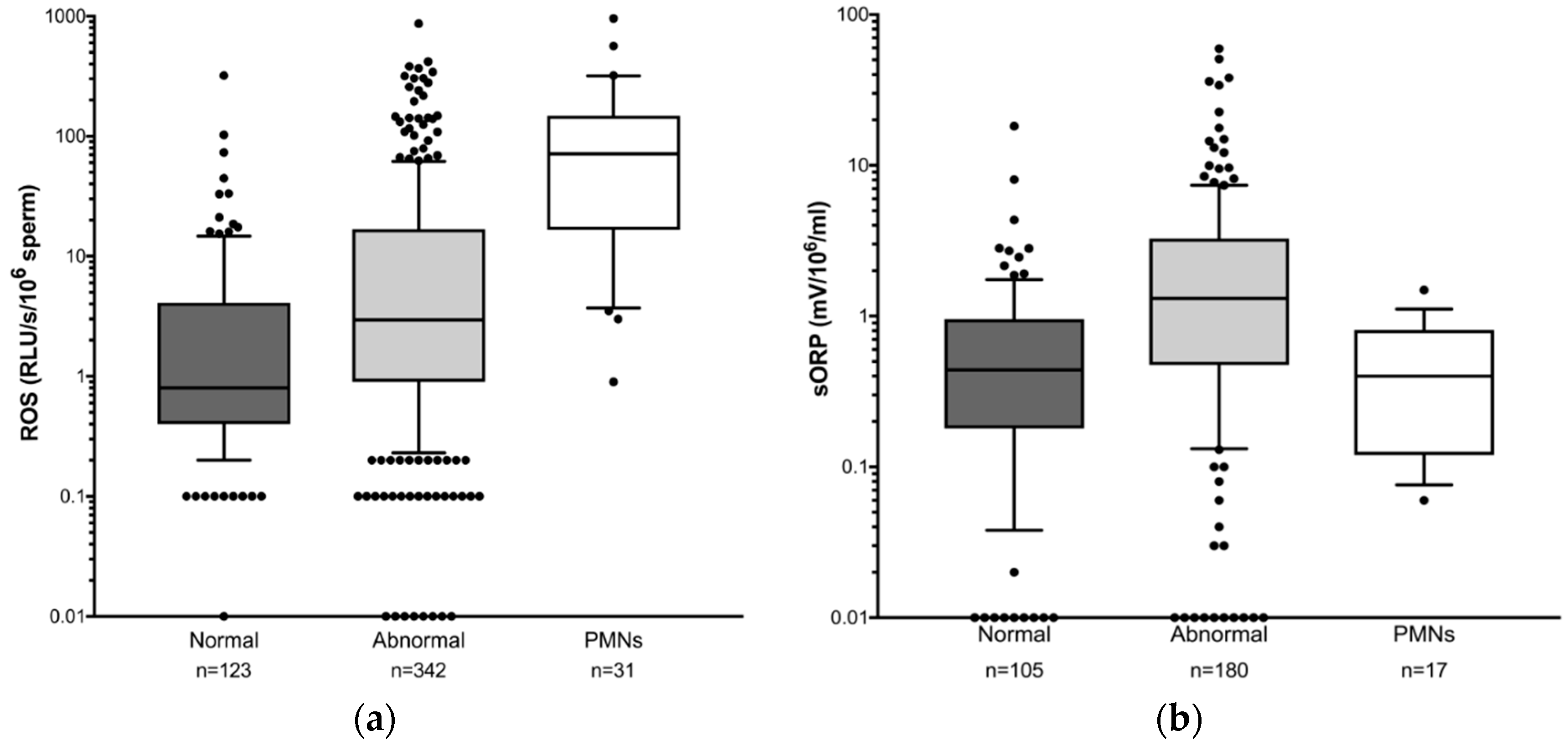
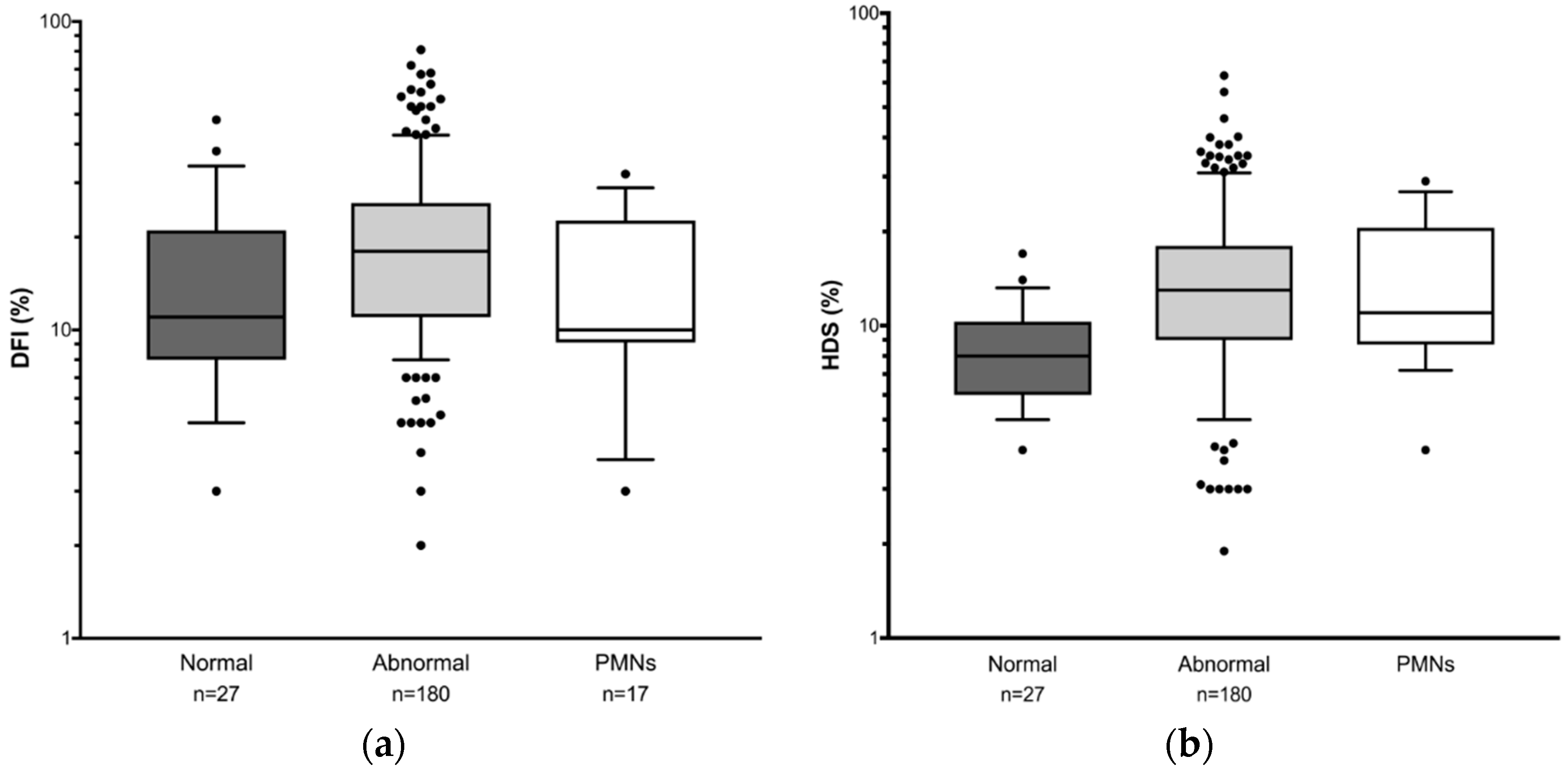
3.4. Comparison of Sperm DNA Damage and OS among Different Patient Groups Selected According to Semen Parameters
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Darbandi, M.; Darbandi, S.; Agarwal, A.; Sengupta, P.; Durairajanayagam, D.; Henkel, R.; Sadeghi, M. Reactive Oxygen Species and Male Reproductive Hormones. Reprod. Biol. Endocrinol. 2018, 16, 87. [Google Scholar] [CrossRef] [PubMed]
- Wagner, H.; Cheng, J.; Ko, E. Role of Reactive Oxygen Species in Male Infertility: An Updated Review of Literature. Arab J. Urol. 2017, 16, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Wright, C.; Milne, S.; Leeson, H. Sperm DNA Damage Caused by Oxidative Stress: Modifiable Clinical, Lifestyle and Nutritional Factors in Male Infertility. Reproduct. BioMed. Online 2014, 28, 684–703. [Google Scholar] [CrossRef] [PubMed]
- Tremellen, K. Oxidative Stress and Male Infertility—A Clinical Perspective. Hum. Reproduct. Update 2008, 14, 243–258. [Google Scholar] [CrossRef] [PubMed]
- Dada, R.; Bisht, S. Oxidative Stress Major Executioner in Disease Pathology Role in Sperm DNA Damage and Preventive Strategies. Front. Biosci. 2017, 9, 420–447. [Google Scholar] [CrossRef]
- Aitken, R. Reactive Oxygen Species as Mediators of Sperm Capacitation and Pathological Damage. Mol. Reproduct. Dev. 2017, 84, 1039–1052. [Google Scholar] [CrossRef]
- Du Plessis, S.; Agarwal, A.; Halabi, J.; Tvrda, E. Contemporary Evidence on The Physiological Role of Reactive Oxygen Species In Human Sperm Function. J. Assist. Reproduct. Genet. 2015, 32, 509–520. [Google Scholar] [CrossRef] [PubMed]
- Aitken, R.; Curry, B. Redox Regulation of Human Sperm Function: From the Physiological Control of Sperm Capacitation to the Etiology of Infertility and DNA Damage in The Germ Line. Antioxid. Redox Signal. 2011, 14, 367–381. [Google Scholar] [CrossRef] [PubMed]
- de Lamirande, E.; Jiang, H.; Zini, A.; Kodama, H.; Gagnon, C. Reactive Oxygen Species and Sperm Physiology. Rev. Reproduct. 1997, 2, 48–54. [Google Scholar] [CrossRef]
- O’Flaherty, C.; Matsushita-Fournier, D. Reactive Oxygen Species and Protein Modifications in Spermatozoa. Biol. Reproduct. 2017, 97, 577–585. [Google Scholar] [CrossRef]
- Sakkas, D.; Alvarez, J. Sperm DNA Fragmentation: Mechanisms of Origin, Impact on Reproductive Outcome, and Analysis. Fertil. Steril. 2010, 93, 1027–1036. [Google Scholar] [CrossRef] [PubMed]
- Lobascio, A.; De Felici, M.; Anibaldi, M.; Greco, P.; Minasi, M.; Greco, E. Involvement of Seminal Leukocytes, Reactive Oxygen Species, and Sperm Mitochondrial Membrane Potential in the DNA Damage of the Human Spermatozoa. Andrology 2015, 3, 265–270. [Google Scholar] [CrossRef]
- Aitken, R.; Baker, M. Oxidative Stress, Spermatozoa and Leukocytic Infiltration: Relationships Forged by the Opposing Forces of Microbial Invasion and the Search for Perfection. J. Reproduct. Immunol. 2013, 100, 11–19. [Google Scholar] [CrossRef]
- Mupfiga, C.; Fisher, D.; Kruger, T.; Henkel, R. The Relationship Between Seminal Leukocytes, Oxidative Status in the Ejaculate, and Apoptotic Markers in Human Spermatozoa. Syst. Biol. Reproduct. Med. 2013, 59, 304–311. [Google Scholar] [CrossRef] [PubMed]
- Henkel, R. Leukocytes and Oxidative Stress: Dilemma for Sperm Function and Male Fertility. Asian J. Androl. 2011, 13, 43–52. [Google Scholar] [CrossRef]
- Koppers, A.; De Iuliis, G.; Finnie, J.; McLaughlin, E.; Aitken, R. Significance of Mitochondrial Reactive Oxygen Species in the Generation of Oxidative Stress in Spermatozoa. J. Clin. Endocrinol. Metab. 2008, 93, 3199–3207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ford, W. Regulation of Sperm Function by Reactive Oxygen Species. Hum. Reproduct. Update 2004, 10, 387–399. [Google Scholar] [CrossRef] [PubMed]
- Gomez, E.; Buckingham, D.; Brindle, J.; Lanzafame, F.; Irvine, D.; Aitken, R. Development of an Image Analysis System to Monitor the Retention of Residual Cytoplasm by Human Spermatozoa: Correlation With Biochemical Markers of the Cytoplasmic Space, Oxidative Stress, and Sperm Function. J. Androl. 1996, 17, 276–287. [Google Scholar]
- Aitken, R.; Gibb, Z.; Baker, M.; Drevet, J.; Gharagozloo, P. Causes and Consequences of Oxidative Stress in Spermatozoa. Reproduct. Fertil. Dev. 2016, 28, 1–10. [Google Scholar] [CrossRef]
- Moazamian, R.; Polhemus, A.; Connaughton, H.; Fraser, B.; Whiting, S.; Gharagozloo, P.; Aitken, R. Oxidative Stress and Human Spermatozoa: Diagnostic and Functional Significance of Aldehydes Generated as a Result of Lipid Peroxidation. MHR Basic Sci. Reproduct. Med. 2015, 21, 502–515. [Google Scholar] [CrossRef]
- Muratori, M.; Tamburrino, L.; Marchiani, S.; Cambi, M.; Olivito, B.; Azzari, C.; Forti, G.; Baldi, E. Investigation on the Origin of Sperm DNA Fragmentation: Role of Apoptosis, Immaturity and Oxidative Stress. Mol. Med. 2015, 21, 109–122. [Google Scholar] [CrossRef] [PubMed]
- Aitken, R.; Koppers, A. Apoptosis and DNA Damage in Human Spermatozoa. Asian J. Androl. 2010, 13, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Darbandi, M.; Darbandi, S.; Agarwal, A.; Baskaran, S.; Dutta, S.; Sengupta, P.; Khorram Khorshid, H.; Esteves, S.; Gilany, K.; Hedayati, M.; et al. Reactive Oxygen Species-Induced Alterations in H19-Igf2 Methylation Patterns, Seminal Plasma Metabolites, and Semen Quality. J. Assist. Reproduct. Genet. 2018. [Google Scholar] [CrossRef] [PubMed]
- Menezo, Y.; Silvestris, E.; Dale, B.; Elder, K. Oxidative Stress and Alterations in DNA Methylation: Two Sides of the Same Coin in Reproduction. Reproduct. BioMed. Online 2016, 33, 668–683. [Google Scholar] [CrossRef]
- Tunc, O.; Tremellen, K. Oxidative DNA Damage Impairs Global Sperm DNA Methylation in Infertile Men. J. Assist. Reproduct. Genet. 2009, 26, 537–544. [Google Scholar] [CrossRef] [PubMed]
- Santi, D.; Spaggiari, G.; Simoni, M. Sperm DNA Fragmentation Index as a Promising Predictive Tool for Male Infertility Diagnosis and Treatment Management—Meta-Analyses. Reproduct. BioMed. Online 2018, 37, 315–326. [Google Scholar] [CrossRef]
- Khosravi, F.; Valojerdi, M.; Amanlou, M.; Karimian, L.; Abolhassani, F. Relationship of Seminal Reactive Nitrogen and Oxygen Species and Total Antioxidant Capacity with Sperm DNA Fragmentation in Infertile Couples with Normal and Abnormal Sperm Parameters. Andrologia 2014, 46, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Sharma, R.; Sharma, R.; Assidi, M.; Abuzenadah, A.; Alshahrani, S.; Durairajanayagam, D.; Sabanegh, E. Characterizing Semen Parameters and Their Association with Reactive Oxygen Species in Infertile Men. Reproduct. Biol. Endocrinol. 2014, 12, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Zini, A.; Lamirande, E.; Gagnon, C. Reactive Oxygen Species in Semen of Infertile Patients: Levels of Superoxide Dismutase- And Catalase-Like Activities in Seminal Plasma and Spermatozoa. Int. J. Androl. 1993, 16, 183–188. [Google Scholar] [CrossRef]
- Kamkar, N.; Ramezanali, F.; Sabbaghian, M. The Relationship Between Sperm DNA Fragmentation, Free Radicals and Antioxidant Capacity with Idiopathic Repeated Pregnancy Loss. Reproduct. Biol. 2018, 18, 330–335. [Google Scholar] [CrossRef]
- Leach, M.; Aitken, R.; Sacks, G. Sperm DNA Fragmentation Abnormalities in Men from Couples with a History of Recurrent Miscarriage. Aust. N. Z. J. Obstet. Gynaecol. 2015, 55, 379–383. [Google Scholar] [CrossRef]
- Evenson, D. Sperm Chromatin Structure Assay (SCSA®). In Spermatogenesis. Methods in Molecular Biology (Methods and Protocols); Carrell, D., Aston, K., Eds.; Humana Press: Totowa, NJ, USA, 2013; pp. 147–164. [Google Scholar]
- Homa, S.; Vessey, W.; Perez-Miranda, A.; Riyait, T.; Agarwal, A. Reactive Oxygen Species (ROS) In Human Semen: Determination of a Reference Range. J. Assist. Reproduct. Genet. 2015, 32, 757–764. [Google Scholar] [CrossRef]
- Vessey, W.; Perez-Miranda, A.; Macfarquhar, R.; Agarwal, A.; Homa, S. Reactive Oxygen Species in Human Semen: Validation and Qualification of a Chemiluminescence Assay. Fertil. Steril. 2014, 102, 1576–1583. [Google Scholar] [CrossRef]
- Saleh, R.; Agarwal, A. Oxidative Stress and Male Infertility: From Research Bench to Clinical Practice. J. Androl. 2002, 23, 737–752. [Google Scholar]
- Agarwal, A.; Bui, A. Oxidation-Reduction Potential as a New Marker for Oxidative Stress: Correlation to Male Infertility. Investig. Clin. Urol. 2017, 58, 385–399. [Google Scholar] [CrossRef]
- Bar-Or, D.; Bar-Or, R.; Rael, L.; Brody, E. Oxidative Stress in Severe Acute Illness. Redox Biol. 2015, 4, 340–345. [Google Scholar] [CrossRef]
- Takeshima, T.; Yumura, Y.; Yasuda, K.; Sanjo, H.; Kuroda, S.; Yamanaka, H.; Iwasaki, A. Inverse Correlation Between Reactive Oxygen Species in Unwashed Semen and Sperm Motion Parameters as Measured by a Computer-Assisted Semen Analyzer. Asian J. Androl. 2017, 19, 350–354. [Google Scholar] [CrossRef]
- Pasqualotto, F.; Sharma, R.; Nelson, D.; Thomas, A.; Agarwal, A. Relationship Between Oxidative Stress, Semen Characteristics, and Clinical Diagnosis in Men Undergoing Infertility Investigation. Fertil. Steril. 2000, 73, 459–464. [Google Scholar] [CrossRef]
- Barroso, G.; Morshedi, M.; Oehninger, S. Analysis of DNA Fragmentation, Plasma Membrane Translocation of Phosphatidylserine and Oxidative Stress in Human Spermatozoa. Hum. Reproduct. 2000, 15, 1338–1344. [Google Scholar] [CrossRef]
- Aitken, R.; Buckingham, D.; Brindle, J.; Gomez, E.; Baker, H.; Irvine, D. Andrology: Analysis of Sperm Movement in Relation to the Oxidative Stress Created by Leukocytes in Washed Sperm Preparations and Seminal Plasma. Hum. Reproduct. 1995, 10, 2061–2071. [Google Scholar] [CrossRef]
- Khodair, H.; Omran, T. Evaluation of Reactive Oxygen Species (ROS) and DNA Integrity Assessment in Cases of Idiopathic Male Infertility. Egypt. J. Dermatol. Venerol. 2013, 33, 51–55. [Google Scholar] [CrossRef]
- Saleh, R.; Agarwal, A.; Nada, E.; El-Tonsy, M.; Sharma, R.; Meyer, A.; Nelson, D.; Thomas, A. Negative Effects of Increased Sperm DNA Damage in Relation to Seminal Oxidative Stress in Men with Idiopathic and Male Factor Infertility. Fertil. Steril. 2003, 79, 1597–1605. [Google Scholar] [CrossRef]
- Lopes, S.; Jurisicova, A.; Sun, J.; Casper, R. Reactive Oxygen Species: Potential Cause for DNA Fragmentation in Human Spermatozoa. Hum. Reproduct. 1998, 13, 896–900. [Google Scholar] [CrossRef]
- Majzoub, A.; Arafa, M.; Mahdi, M.; Agarwal, A.; Al Said, S.; Al-Emadi, I.; El Ansari, W.; Alattar, A.; Al Rumaihi, K.; Elbardisi, H. Oxidation–Reduction Potential and Sperm DNA Fragmentation, and Their Associations with Sperm Morphological Anomalies Amongst Fertile and Infertile Men. Arab J. Urol. 2018, 16, 87–95. [Google Scholar] [CrossRef]
- Arafa, M.; Agarwal, A.; Al Said, S.; Majzoub, A.; Sharma, R.; Bjugstad, K.; AlRumaihi, K.; Elbardisi, H. Semen Quality and Infertility Status Can Be Identified Through Measures of Oxidation-Reduction Potential. Andrologia 2017, 50, e12881. [Google Scholar] [CrossRef]
- World Health Organization. WHO Laboratory Manual for the Examination and Processing of Human Semen, 5th ed.; WHO Press: Geneva, Switzerland, 2010. [Google Scholar]
- Rael, L.; Bar-Or, R.; Kelly, M.; Carrick, M.; Bar-Or, D. Assessment of Oxidative Stress in Patients with an Isolated Traumatic Brain Injury Using Disposable Electrochemical Test Strips. Electroanalysis 2015, 27, 2567–2573. [Google Scholar] [CrossRef]
- Stahl, P.; Cogan, C.; Mehta, A.; Bolyakov, A.; Paduch, D.; Goldstein, M. Concordance Among Sperm Deoxyribonucleic Acid Integrity Assays and Semen Parameters. Fertil. Steril. 2015, 104, 56–61. [Google Scholar] [CrossRef]
- Agarwal, A.; Qiu, E.; Sharma, R. Laboratory Assessment of Oxidative Stress in Semen. Arab J. Urol. 2018, 16, 77–86. [Google Scholar] [CrossRef]
- Wardman, P. Fluorescent and Luminescent Probes for Measurement of Oxidative and Nitrosative Species in Cells and Tissues: Progress, Pitfalls, and Prospects. Free Radic. Biol. Med. 2007, 43, 995–1022. [Google Scholar] [CrossRef]
- Aitken, R.; Baker, M.; O’Bryan, M. Andrology Lab Corner*: Shedding Light on Chemiluminescence: The Application of Chemiluminescence in Diagnostic Andrology. J. Androl. 2004, 25, 455–465. [Google Scholar] [CrossRef]
- Spanidis, Y.; Mpesios, A.; Stagos, D.; Goutzourelas, N.; Bar-Or, D.; Karapetsa, M.; Zakynthinos, E.; Spandidos, D.; Tsatsakis, A.; Leon, G.; et al. Assessment of the Redox Status in Patients with Metabolic Syndrome and Type 2 Diabetes Reveals Great Variations. Exp. Ther. Med. 2016, 11, 895–903. [Google Scholar] [CrossRef]
- Aitken, R.; De Iuliis, G.; Finnie, J.; Hedges, A.; McLachlan, R. Analysis of the Relationships Between Oxidative Stress, DNA Damage and Sperm Vitality in a Patient Population: Development of Diagnostic Criteria. Hum. Reproduct. 2010, 25, 2415–2426. [Google Scholar] [CrossRef]
- Smit, M.; Romijn, J.; Wildhagen, M.; Weber, R.; Dohle, G. Sperm Chromatin Structure is Associated with the Quality of Spermatogenesis in Infertile Patients. Fertil. Steril. 2010, 94, 1748–1752. [Google Scholar] [CrossRef]
- Moskovtsev, S.; Willis, J.; White, J.; Mullen, J. Sperm DNA Damage: Correlation to Severity of Semen Abnormalities. Urology 2009, 74, 789–793. [Google Scholar] [CrossRef]
- Agarwal, A.; Wang, S. Clinical Relevance of Oxidation-Reduction Potential in the Evaluation of Male Infertility. Urology 2017, 104, 84–89. [Google Scholar] [CrossRef]
- Henkel, R.; Kierspel, E.; Stalf, T.; Mehnert, C.; Menkveld, R.; Tinneberg, H.; Schill, W.; Kruger, T. Effect of Reactive Oxygen Species Produced by Spermatozoa and Leukocytes on Sperm Functions in Non-Leukocytospermic Patients. Fertil. Steril. 2005, 83, 635–642. [Google Scholar] [CrossRef]
- Micillo, A.; Vassallo, M.; Cordeschi, G.; D’Andrea, S.; Necozione, S.; Francavilla, F.; Francavilla, S.; Barbonetti, A. Semen Leukocytes and Oxidative-Dependent DNA Damage of Spermatozoa in Male Partners of Subfertile Couples With No Symptoms of Genital Tract Infection. Andrology 2016, 4, 808–815. [Google Scholar] [CrossRef]
- Alvarez, J.G.; Sharma, R.K.; Ollero, M.; Saleh, R.A.; Lopez, M.C.; Thomas, A.J., Jr.; Evenson, D.P.; Agarwal, A. Increased DNA Damage in Sperm from Leukocytospermic Semen Samples as Determined by the Sperm Chromatin Structure Assay. Fertil. Steril. 2002, 7, 319–329. [Google Scholar] [CrossRef]
- Simbini, T.; Umapathy, E.; Jacobus, E.; Tendaupenyu, G.; Mbizvo, M. Study on the Origin of Seminal Leucocytes Using Split Ejaculate Technique and the Effect of Leucocytospermia on Sperm Characteristics. Urol. Int. 1998, 61, 95–100. [Google Scholar] [CrossRef]
- Halliwell, B.; Gutteridge, J. The Antioxidants of Human Extracellular Fluids. Arch. Biochem. Biophys. 1990, 280, 1–8. [Google Scholar] [CrossRef]
- Ma, Y.; Xie, N.; Li, Y.; Zhang, B.; Xie, D.; Zhang, W.; Li, Q.; Yu, H.; Zhang, Q.; Ni, Y.; et al. Teratozoospermia with Amorphous Sperm Head Associate with Abnormal Chromatin Condensation in a Chinese Family. Syst. Biol. Reproduct. Med. 2018, 19, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Zini, A.; Phillips, S.; Courchesne, A.; Boman, J.; Baazeem, A.; Bissonnette, F.; Kadoch, I.; San Gabriel, M. Sperm Head Morphology is Related to High Deoxyribonucleic Acid Stainability Assessed by Sperm Chromatin Structure Assay. Fertil. Steril. 2009, 91, 2495–2500. [Google Scholar] [CrossRef] [PubMed]
- Aitken, R.J.; De Iuliis, G.N. On the Possible Origins of DNA Damage in Human Spermatozoa. Mol. Hum. Reproduct. 2010, 16, 3–13. [Google Scholar] [CrossRef]
- De Iuliis, G.N.; Thomson, L.K.; Mitchell, L.A.; Finnie, J.M.; Koppers, A.J.; Hedges, A.; Nixon, B.; Aitken, R.J. DNA Damage in Human Spermatozoa is Highly Correlated with the Efficiency of Chromatin Remodeling and the Formation of 8-hydroxy-20-deoxyguanosine, a Marker of Oxidative Stress. Biol. Reproduct. 2009, 81, 517–524. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| ROS | sORP | |||
|---|---|---|---|---|
| Number of Patients | % | Number of Patients | % | |
| Normozoopsermia | 172 | 34.6 | 139 | 46.2 |
| Oligozoospermia | 18 | 3.6 | 9 | 3.0 |
| Asthenozoospermia | 8 | 1.6 | 6 | 2.0 |
| Teratozoospermia | 119 | 24.0 | 72 | 23.9 |
| Oligoasthenozoospermia | 1 | <1 | 1 | <1 |
| Oligoteratozoospermia | 69 | 13.9 | 32 | 10.6 |
| Asthenoteratozoospermia | 29 | 5.8 | 11 | 3.7 |
| Oligoasthenoteratozoospermia | 49 | 9.9 | 15 | 5.0 |
| Leukocytospermia | 31 | 6.3 | 16 | 5.3 |
| TOTAL | 496 | - | 301 | - |
| All Samples | All Samples Excluding those with Leukocytospermia | |
|---|---|---|
| Oxidative stress measured by ROS | ||
| Oxidative balanced | 18.78 ± 1.10 (161) | 18.75 ± 1.12 (159) |
| Oxidative stressed | 22.86 ± 1.59 (77) | 24.67 ± 1.78 (63) |
| p value | 0.0359 | 0.0052 |
| Oxidative stress measured by sORP | ||
| Oxidative balanced | 11.97 ± 1.41 (30) | 12.14 ± 1.49 (28) |
| Oxidative stressed | 19.39 ± 1.83 (18) | 19.39 ± 1.86 (18) |
| p value | 0.0024 | 0.004 |
| All Samples | All Samples Excluding those with Leukocytospermia | |
|---|---|---|
| Oxidative stress measured by ROS | ||
| Oxidative balanced | 13.45 ± 0.74 (161) | 13.49 ± 0.75 (159) |
| Oxidative stressed | 15.78 ± 1.02 (77) | 16.19 ± 1.16 (63) |
| p value | 0.0097 | 0.0077 |
| Oxidative stress measured by sORP | ||
| Oxidative balanced | 11.07 ± 1.11 (30) | 10.61 ± 1.09 (28) |
| Oxidative stressed | 17.89 ± 3.40 (18) | 17.89 ± 3.40 (18) |
| I value | 0.0881 | 0.0672 |
| Value | Count/ml | Total Motility | Progressive Motility | Total Motile Sperm Count | Vitality | Morphology | |
|---|---|---|---|---|---|---|---|
| ROS | R2 | −0.15729 | −0.14482 | −0.14444 | −0.17395 | −0.20832 | −0.12536 |
| p value | 0.0004 | 0.0012 | 0.0013 | 0.0001 | <0.0001 | 0.0053 | |
| n | 496 | 495 | 495 | 495 | 495 | 493 | |
| sORP | R2 | −0.24628 | −0.21101 | −0.23561 | −0.25055 | −0.13519 | −0.22642 |
| p value | <0.0001 | 0.0002 | <0.0001 | <0.0001 | 0.019 | <0.0001 | |
| n | 301 | 301 | 301 | 301 | 301 | 300 | |
| DFI | R2 | -0.19182 | −0.53951 | −0.48693 | −0.27539 | −0.5727 | −0.19016 |
| p value | 0.0041 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | 0.0047 | |
| n | 222 | 221 | 221 | 221 | 221 | 220 | |
| HDS | R2 | −0.36663 | −0.23638 | −0.24938 | −0.27703 | −0.11497 | −0.48848 |
| p value | <0.0001 | 0.0004 | 0.0002 | <0.0001 | 0.0882 | <0.0001 | |
| n | 222 | 221 | 221 | 221 | 221 | 220 | |
| PMN | R2 | 0.2098 | 0.03497 | 0.04169 | 0.15498 | 0.03037 | 0.04361 |
| p value | <0.0001 | 0.4389 | 0.3561 | 0.0006 | 0.5015 | 0.3354 | |
| n | 493 | 492 | 492 | 492 | 492 | 490 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Homa, S.T.; Vassiliou, A.M.; Stone, J.; Killeen, A.P.; Dawkins, A.; Xie, J.; Gould, F.; Ramsay, J.W.A. A Comparison Between Two Assays for Measuring Seminal Oxidative Stress and their Relationship with Sperm DNA Fragmentation and Semen Parameters. Genes 2019, 10, 236. https://0-doi-org.brum.beds.ac.uk/10.3390/genes10030236
Homa ST, Vassiliou AM, Stone J, Killeen AP, Dawkins A, Xie J, Gould F, Ramsay JWA. A Comparison Between Two Assays for Measuring Seminal Oxidative Stress and their Relationship with Sperm DNA Fragmentation and Semen Parameters. Genes. 2019; 10(3):236. https://0-doi-org.brum.beds.ac.uk/10.3390/genes10030236
Chicago/Turabian StyleHoma, Sheryl T., Anna M. Vassiliou, Jesse Stone, Aideen P. Killeen, Andrew Dawkins, Jingyi Xie, Farley Gould, and Jonathan W. A. Ramsay. 2019. "A Comparison Between Two Assays for Measuring Seminal Oxidative Stress and their Relationship with Sperm DNA Fragmentation and Semen Parameters" Genes 10, no. 3: 236. https://0-doi-org.brum.beds.ac.uk/10.3390/genes10030236