Pigment Intensity in Dogs is Associated with a Copy Number Variant Upstream of KITLG

 , and
, and 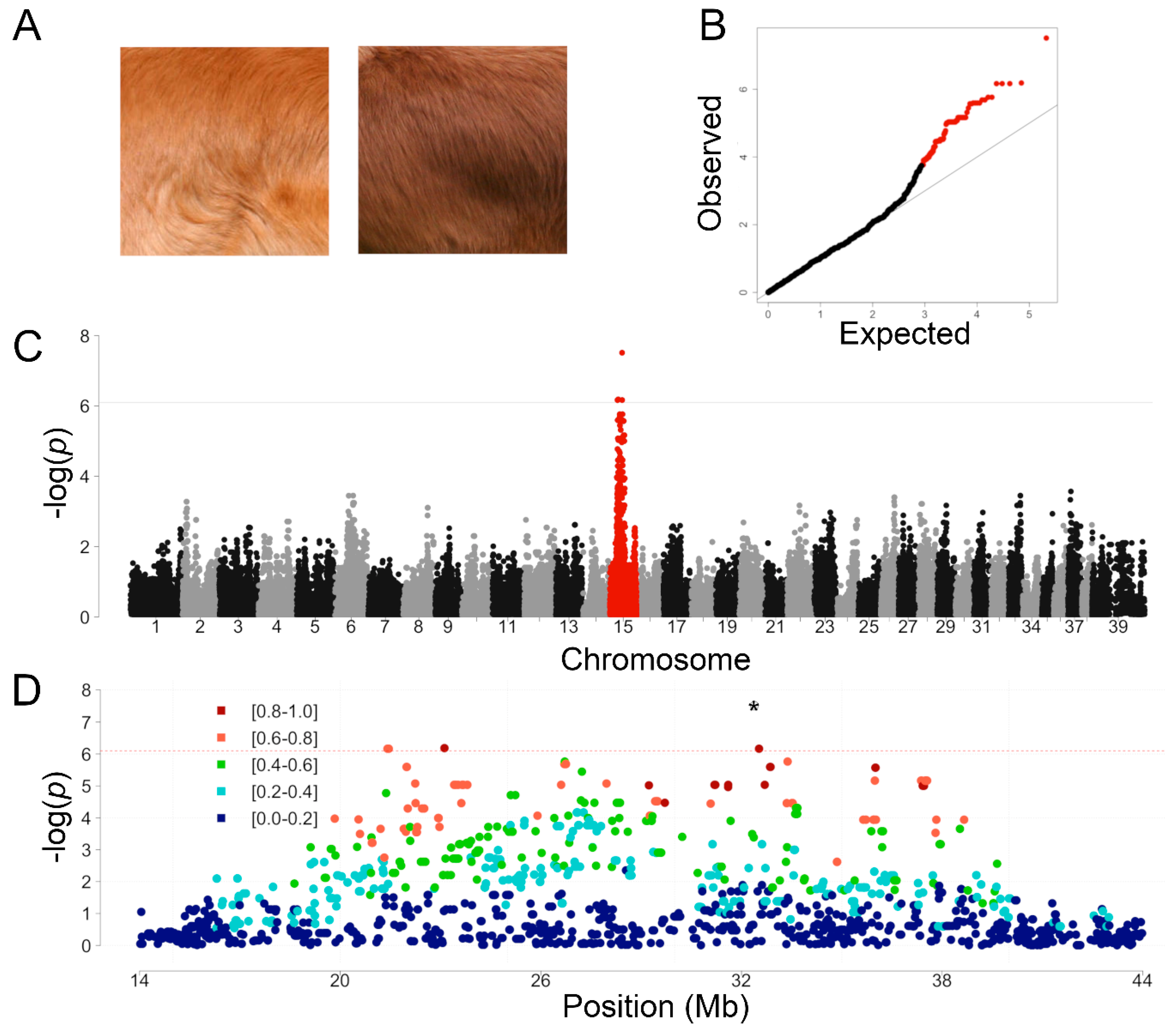{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and DNA Extraction
2.2. Genome-Wide Association Study
2.3. Whole-Genome Sequencing
2.4. Digital Droplet PCR Genotyping
2.5. Quantitative Hair Pigment Analysis
3. Results
3.1. Genome-Wide Association Study in Light and Dark Red NSDTR
3.2. Whole Genome Sequence Variant and Coverage Analysis
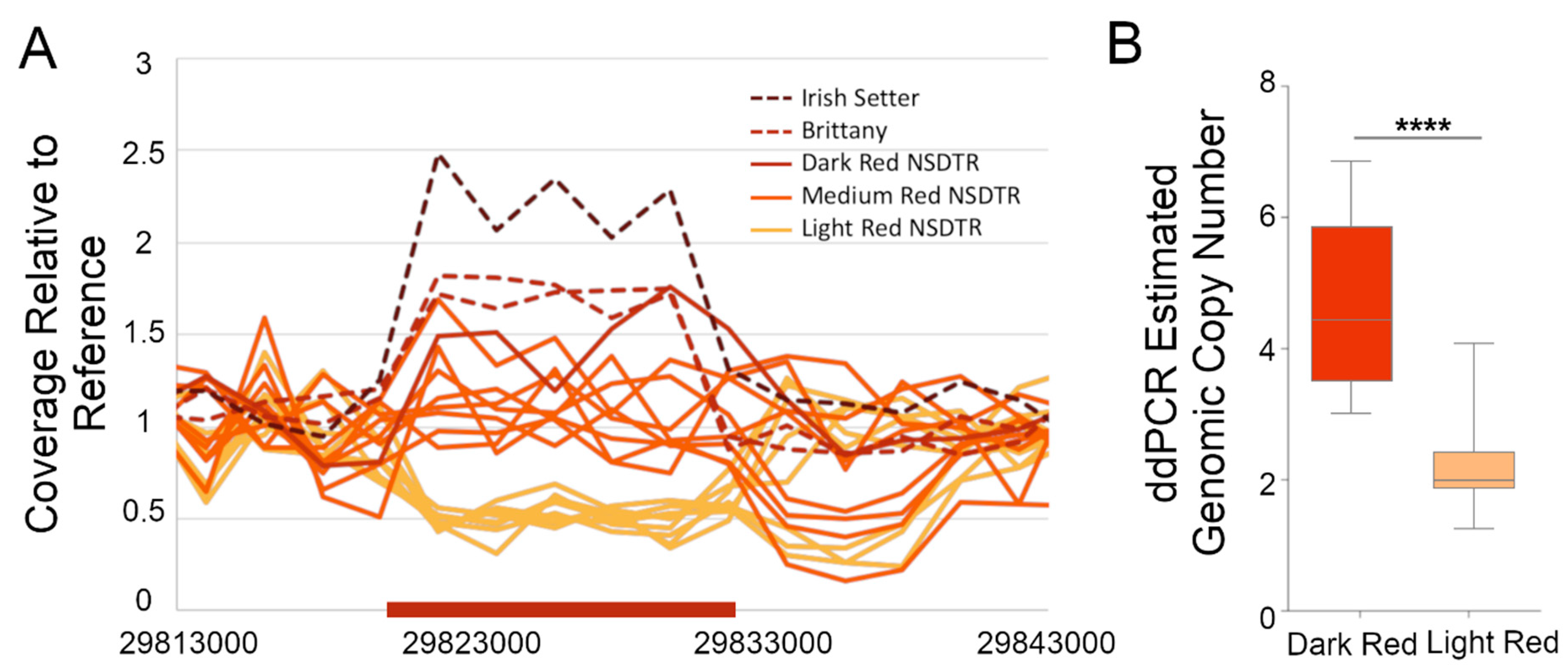
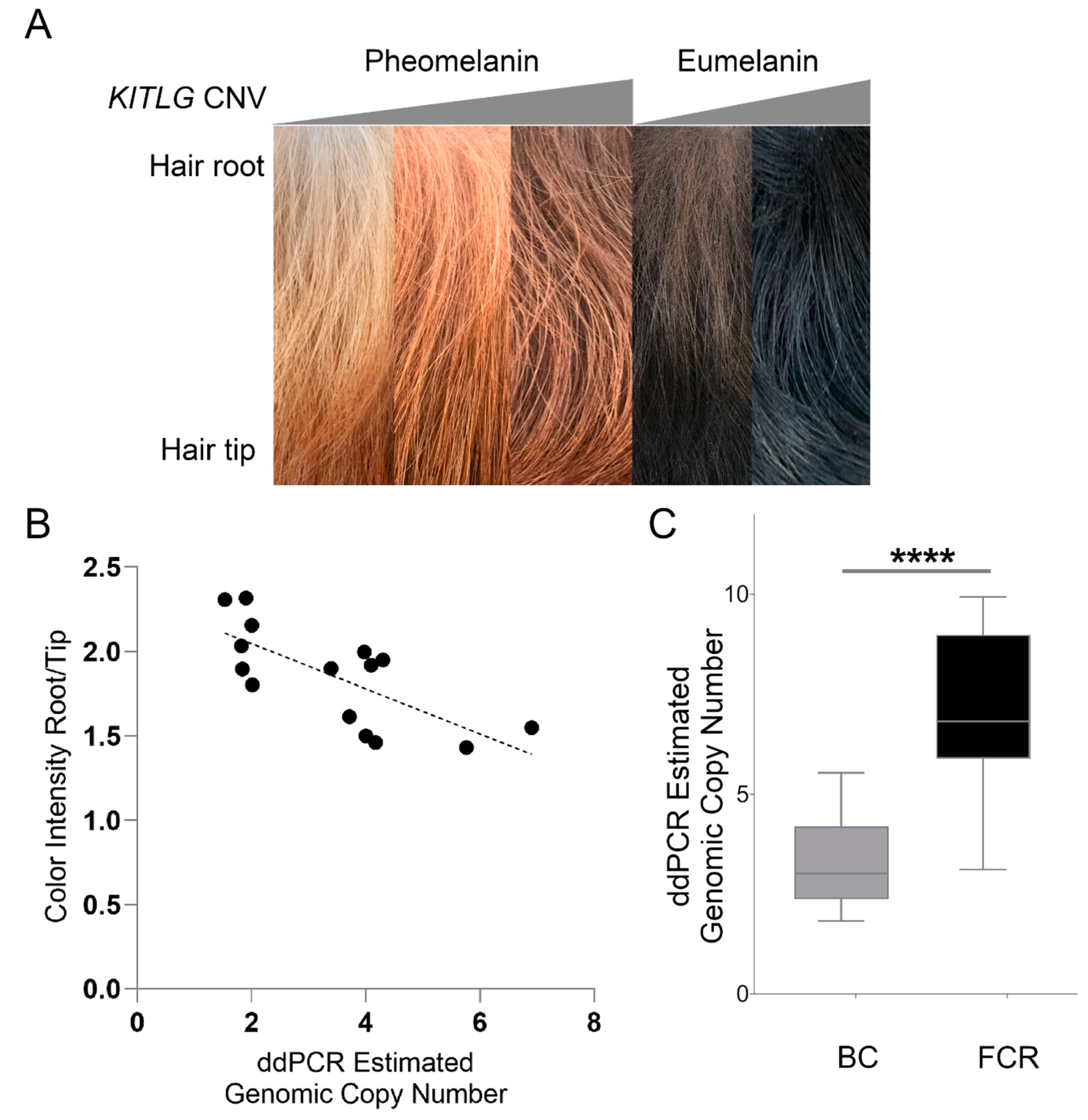
3.3. Validation of the KITLG CNV in Pheomelanin-Based Coat Colors
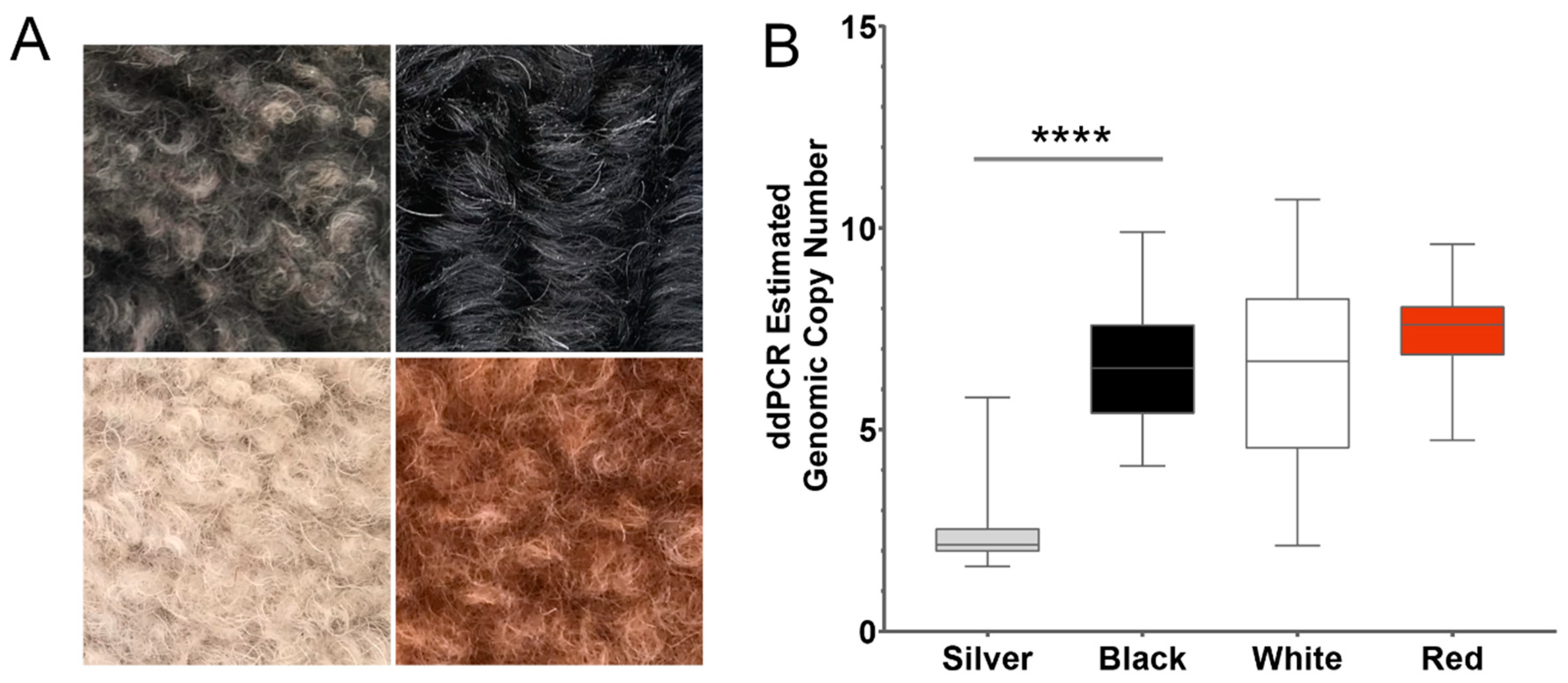
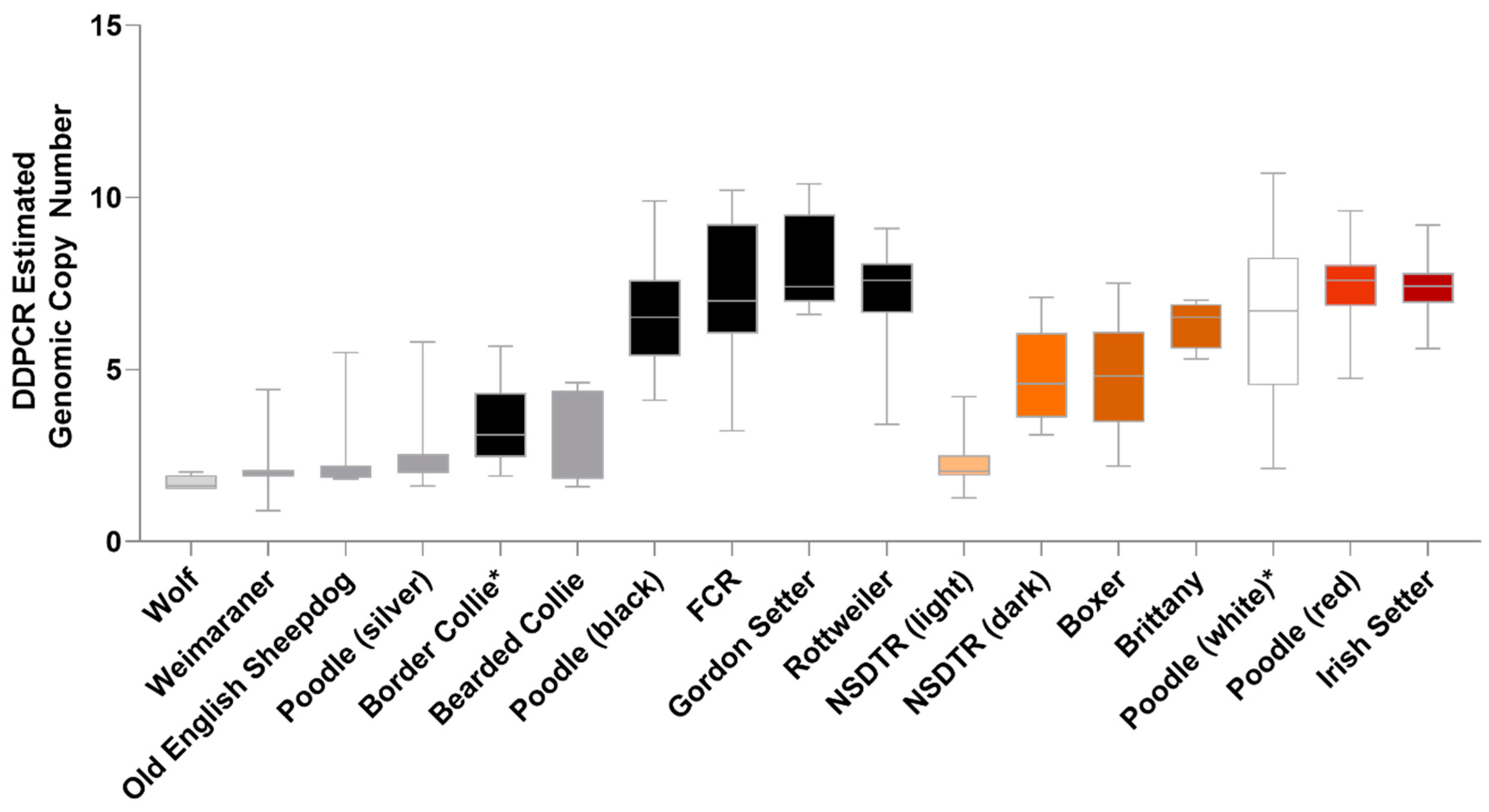
3.4. Validation of the KITLG CNV in Eumelanin-Based Coat Colors
3.5. Comparison of the Hair Shaft in High and Low Copy Number Dogs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Larson, G.; Karlsson, E.K.; Perri, A.; Webster, M.T.; Ho, S.Y.W.; Peters, J.; Stahl, P.W.; Piper, P.J.; Lingaas, F.; Fredholm, M.; et al. Rethinking dog domestication by integrating genetics, archeology, and biogeography. Proc. Natl. Acad. Sci. USA 2012, 109, 8878–8883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shannon, L.M.; Boyko, R.H.; Castelhano, M.; Corey, E.; Hayward, J.J.; McLean, C.; White, M.E.; Abi Said, M.; Anita, B.A.; Bondjengo, N.I.; et al. Genetic structure in village dogs reveals a Central Asian domestication origin. Proc. Natl. Acad. Sci. USA 2015, 112, 13639–13644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, G.-D.; Peng, M.-S.; Yang, H.-C.; Savolainen, P.; Zhang, Y.-P. Questioning the evidence for a Central Asian domestication origin of dogs. Proc. Natl. Acad. Sci. USA 2016, 113, E2554–E2555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Botigue, L.R.; Song, S.; Scheu, A.; Gopalan, S.; Pendleton, A.L.; Oetjens, M.; Taravella, A.M.; Seregely, T.; Zeeb-Lanz, A.; Arbogast, R.M.; et al. Ancient European dog genomes reveal continuity since the Early Neolithic. Nat. Commun. 2017, 8, 16082. [Google Scholar] [CrossRef]
- Anderson, T.M.; vonHoldt, B.M.; Candille, S.I.; Musiani, M.; Greco, C.; Stahler, D.R.; Smith, D.W.; Padhukasahasram, B.; Randi, E.; Leonard, J.A.; et al. Molecular and evolutionary history of melanism in North American gray wolves. Science (New York N.Y.) 2009, 323, 1339–1343. [Google Scholar] [CrossRef] [Green Version]
- Ollivier, M.; Tresset, A.; Hitte, C.; Petit, C.; Hughes, S.; Gillet, B.; Duffraisse, M.; Pionnier-Capitan, M.; Lagoutte, L.; Arbogast, R.-M.; et al. Evidence of Coat Color Variation Sheds New Light on Ancient Canids. PLoS ONE 2013, 8, e75110. [Google Scholar] [CrossRef]
- Kerns, J.A.; Newton, J.; Berryere, T.G.; Rubin, E.M.; Cheng, J.F.; Schmutz, S.M.; Barsh, G.S. Characterization of the dog Agouti gene and a nonagoutimutation in German Shepherd Dogs. Mamm. Genome 2004, 15, 798–808. [Google Scholar] [CrossRef]
- Newton, J.M.; Wilkie, A.L.; He, L.; Jordan, S.A.; Metallinos, D.L.; Holmes, N.G.; Jackson, I.J.; Barsh, G.S. Melanocortin 1 receptor variation in the domestic dog. Mamm. Genome 2000, 11, 24–30. [Google Scholar] [CrossRef]
- Candille, S.I.; Kaelin, C.B.; Cattanach, B.M.; Yu, B.; Thompson, D.A.; Nix, M.A.; Kerns, J.A.; Schmutz, S.M.; Millhauser, G.L.; Barsh, G.S. A -defensin mutation causes black coat color in domestic dogs. Science (New York N.Y.) 2007, 318, 1418–1423. [Google Scholar] [CrossRef] [Green Version]
- Berryere, T.G.; Kerns, J.A.; Barsh, G.S.; Schmutz, S.M. Association of an Agouti allele with fawn or sable coat color in domestic dogs. Mamm. Genome 2005, 16, 262–272. [Google Scholar] [CrossRef]
- Robbins, L.S.; Nadeau, J.H.; Johnson, K.R.; Kelly, M.A.; Rosellirehfuss, L.; Baack, E.; Mountjoy, K.G.; Cone, R.D. Pigmentation phenotypes of variant extension locus alleles result from point mutations that alter msh receptor function. Cell 1993, 72, 827–834. [Google Scholar] [CrossRef]
- Hedan, B.; Cadieu, E.; Botherel, N.; Dufaure de Citres, C.; Letko, A.; Rimbault, M.; Drogemuller, C.; Jagannathan, V.; Derrien, T.; Schmutz, S.; et al. Identification of a Missense Variant in MFSD12 Involved in Dilution of Phaeomelanin Leading to White or Cream Coat Color in Dogs. Genes 2019, 10, 386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Philipp, U.; Hamann, H.; Mecklenburg, L.; Nishino, S.; Mignot, E.; Gunzel-Apel, A.R.; Schmutz, S.M.; Leeb, T. Polymorphisms within the canine MLPH gene are associated with dilute coat color in dogs. BMC Genet. 2005, 6, 34. [Google Scholar] [CrossRef] [Green Version]
- Schmutz, S.M.; Berryere, T.G.; Goldfinch, A.D. TYRP1 and MC1R genotypes and their effects on coat color in dogs. Mamm. Genome 2002, 13, 380–387. [Google Scholar] [CrossRef] [PubMed]
- Cargill, E.J.; Famula, T.R.; Schnabel, R.D.; Strain, G.M.; Murphy, K.E. The color of a Dalmatian’s spots: Linkage evidence to support the TYRP1gene. BMC Vet. Res. 2005, 1, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hrckova Turnova, E.; Majchrakova, Z.; Bielikova, M.; Soltys, K.; Turna, J.; Dudas, A. A novel mutation in the TYRP1 gene associated with brown coat colour in the Australian Shepherd Dog Breed. Anim. Genet. 2017, 48, 626. [Google Scholar] [CrossRef]
- Nicholas, T.J.; Cheng, Z.; Ventura, M.; Mealey, K.; Eichler, E.E.; Akey, J.M. The genomic architecture of segmental duplications and associated copy number variants in dogs. Genome Res. 2009, 19, 491–499. [Google Scholar] [CrossRef] [Green Version]
- Lindblad-Toh, K.; Wade, C.M.; Mikkelsen, T.S.; Karlsson, E.K.; Jaffe, D.B.; Kamal, M.; Clamp, M.; Chang, J.L.; Kulbokas, E.J., III; Zody, M.C.; et al. Genome sequence, comparative analysis and haplotype structure of the domestic dog. Nature 2005, 438, 803–819. [Google Scholar] [CrossRef]
- Kim, K.S.; Lee, S.E.; Jeong, H.W.; Ha, J.H. The complete nucleotide sequence of the domestic dog (Canis familiaris) mitochondrial genome. Mol. Phylogenet. Evol. 1998, 10, 210–220. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.C.; Chow, C.C.; Tellier, L.C.A.M.; Vattikuti, S.; Purcell, S.M.; Lee, J.J. Second-generation PLINK: Rising to the challenge of larger and richer datasets. GigaScience 2015, 4. [Google Scholar] [CrossRef]
- Steiß, V.; Letschert, T.; Schäfer, H.; Pahl, R. PERMORY-MPI: A program for high-speed parallel permutation testing in genome-wide association studies. Bioinformatics (Oxf. Engl.) 2012, 28, 1168–1169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, S.D. qqman: An R package for visualizing GWAS results using Q-Q and manhattan plots. bioRxiv 2014. [Google Scholar] [CrossRef]
- Karssen, L.C.; van Duijn, C.M.; Aulchenko, Y.S. The GenABEL Project for statistical genomics. F1000Res 2016, 5, 914. [Google Scholar] [CrossRef] [PubMed]
- Kierczak, M.; Jablonska, J.; Forsberg, S.K.; Bianchi, M.; Tengvall, K.; Pettersson, M.; Scholz, V.; Meadows, J.R.; Jern, P.; Carlborg, O.; et al. cgmisc: Enhanced genome-wide association analyses and visualization. Bioinformatics (Oxf. Engl. ) 2015, 31, 3830–3831. [Google Scholar] [CrossRef] [PubMed]
- Brown, E.A.; Dickinson, P.J.; Mansour, T.; Sturges, B.K.; Aguilar, M.; Young, A.E.; Korff, C.; Lind, J.; Ettinger, C.L.; Varon, S.; et al. FGF4 retrogene on CFA12 is responsible for chondrodystrophy and intervertebral disc disease in dogs. Proc. Natl. Acad. Sci. USA 2017, 114, 11476–11481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoeppner, M.P.; Lundquist, A.; Pirun, M.; Meadows, J.R.S.; Zamani, N.; Johnson, J.; Sundström, G.; Cook, A.; FitzGerald, M.G.; Swofford, R.; et al. An Improved Canine Genome and a Comprehensive Catalogue of Coding Genes and Non-Coding Transcripts. PLoS ONE 2014, 9, e91172. [Google Scholar] [CrossRef] [PubMed]
- Kent, W.J.; Sugnet, C.W.; Furey, T.S.; Roskin, K.M.; Pringle, T.H.; Zahler, A.M.; Haussler, D. The human genome browser at UCSC. Genome Res. 2002, 12, 996–1006. [Google Scholar] [CrossRef] [Green Version]
- Kent, W.J.; Baertsch, R.; Hinrichs, A.; Miller, W.; Haussler, D. Evolution’s cauldron: Duplication, deletion, and rearrangement in the mouse and human genomes. Proc. Natl. Acad. Sci. USA 2003, 100, 11484–11489. [Google Scholar] [CrossRef] [Green Version]
- Chiaromonte, F.; Yap, V.B.; Miller, W. Scoring pairwise genomic sequence alignments. Pac. Symp. Biocomput. 2002, 115–126. [Google Scholar] [CrossRef]
- Blanchette, M.; Kent, W.J.; Riemer, C.; Elnitski, L.; Smit, A.F.; Roskin, K.M.; Baertsch, R.; Rosenbloom, K.; Clawson, H.; Green, E.D.; et al. Aligning multiple genomic sequences with the threaded blockset aligner. Genome Res. 2004, 14, 708–715. [Google Scholar] [CrossRef] [Green Version]
- McLaren, W.; Gil, L.; Hunt, S.E.; Riat, H.S.; Ritchie, G.R.S.; Thormann, A.; Flicek, P.; Cunningham, F. The Ensembl Variant Effect Predictor. Genome Biol. 2016, 17, 122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McLaren, W.; Pritchard, B.; Rios, D.; Chen, Y.; Flicek, P.; Cunningham, F. Deriving the consequences of genomic variants with the Ensembl API and SNP Effect Predictor. Bioinformatics (Oxf. Engl.) 2010, 26, 2069–2070. [Google Scholar] [CrossRef] [PubMed]
- Robinson, J.T.; Thorvaldsdóttir, H.; Winckler, W.; Guttman, M.; Lander, E.S.; Getz, G.; Mesirov, J.P. Integrative genomics viewer. Nat. Biotechnol. 2011, 29, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Besmer, P. The kit ligand encoded at the murine Steel locus: A pleiotropic growth and differentiation factor. Curr. Opin. Cell Biol. 1991, 3, 939–946. [Google Scholar] [CrossRef]
- Reissmann, M.; Ludwig, A. Pleiotropic effects of coat colour-associated mutations in humans, mice and other mammals. Semin. Cell Dev. Biol. 2013, 24, 576–586. [Google Scholar] [CrossRef]
- Geissler, E.N.; Ryan, M.A.; Housman, D.E. The dominant-white spotting (W) locus of the mouse encodes the c-kit proto-oncogene. Cell 1988, 55, 185–192. [Google Scholar] [CrossRef]
- Brannan, C.I.; Lyman, S.D.; Williams, D.E.; Eisenman, J.; Anderson, D.M.; Cosman, D.; Bedell, M.A.; Jenkins, N.A.; Copeland, N.G. Steel-Dickie mutation encodes a c-kit ligand lacking transmembrane and cytoplasmic domains. Proc. Natl. Acad. Sci. USA 1991, 88, 4671–4674. [Google Scholar] [CrossRef] [Green Version]
- Grichnik, J.M.; Burch, J.A.; Burchette, J.; Shea, C.R. The SCF/KIT pathway plays a critical role in the control of normal human melanocyte homeostasis. J. Investig. Dermatol. 1998, 111, 233–238. [Google Scholar] [CrossRef] [Green Version]
- Liao, C.P.; Booker, R.C.; Morrison, S.J.; Le, L.Q. Identification of hair shaft progenitors that create a niche for hair pigmentation. Genes Dev. 2017, 31, 744–756. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.Q.; Si, L.; Tang, Q.; Lin, D.; Fu, Z.; Zhang, J.; Cui, B.; Zhu, Y.; Kong, X.; Deng, M.; et al. Gain-of-function mutation of KIT ligand on melanin synthesis causes familial progressive hyperpigmentation. Am. J. Hum. Genet. 2009, 84, 672–677. [Google Scholar] [CrossRef] [Green Version]
- Zazo Seco, C.; Serrao de Castro, L.; van Nierop, J.W.; Morin, M.; Jhangiani, S.; Verver, E.J.; Schraders, M.; Maiwald, N.; Wesdorp, M.; Venselaar, H.; et al. Allelic Mutations of KITLG, Encoding KIT Ligand, Cause Asymmetric and Unilateral Hearing Loss and Waardenburg Syndrome Type 2. Am. J. Hum. Genet. 2015, 97, 647–660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giebel, L.B.; Spritz, R.A. Mutation of the KIT (mast/stem cell growth factor receptor) protooncogene in human piebaldism. Proc. Natl. Acad. Sci. USA 1991, 88, 8696–8699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.J.; Zhao, R.; He, X.Y.; Li, L.P.; Wang, K.W.; Zhao, L.; Tu, M.; Tang, J.S.; Xie, Z.G.; Zhu, Y.M. A novel splicing mutation of KIT results in piebaldism and auburn hair color in a Chinese family. BioMed Res. Int. 2013, 2013, 689756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richards, K.A.; Fukai, K.; Oiso, N.; Paller, A.S. A novel KIT mutation results in piebaldism with progressive depigmentation. J. Am. Acad. Dermatol. 2001, 44, 288–292. [Google Scholar] [CrossRef]
- Murakami, T.; Fukai, K.; Oiso, N.; Hosomi, N.; Kato, A.; Garganta, C.; Barnicoat, A.; Poppelaars, F.; Aquaron, R.; Paller, A.S.; et al. New KIT mutations in patients with piebaldism. J. Dermatol. Sci. 2004, 35, 29–33. [Google Scholar] [CrossRef]
- Sulem, P.; Gudbjartsson, D.F.; Stacey, S.N.; Helgason, A.; Rafnar, T.; Magnusson, K.P.; Manolescu, A.; Karason, A.; Palsson, A.; Thorleifsson, G.; et al. Genetic determinants of hair, eye and skin pigmentation in Europeans. Nat. Genet. 2007, 39, 1443–1452. [Google Scholar] [CrossRef]
- Miller, C.T.; Beleza, S.; Pollen, A.A.; Schluter, D.; Kittles, R.A.; Shriver, M.D.; Kingsley, D.M. cis-Regulatory changes in Kit ligand expression and parallel evolution of pigmentation in sticklebacks and humans. Cell 2007, 131, 1179–1189. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Shi, H.; Ma, P.; Zhao, S.; Kong, Q.; Bian, T.; Gong, C.; Zhao, Q.; Liu, Y.; Qi, X.; et al. Darwinian positive selection on the pleiotropic effects of KITLG explain skin pigmentation and winter temperature adaptation in Eurasians. Mol. Biol. Evol. 2018. [Google Scholar] [CrossRef]
- Guenther, C.A.; Tasic, B.; Luo, L.; Bedell, M.A.; Kingsley, D.M. A molecular basis for classic blond hair color in Europeans. Nat. Genet. 2014, 46, 748–752. [Google Scholar] [CrossRef] [Green Version]
- Sarvella, P.A.; Russell, L.B. Steel, a new dominant gene in the house mouse. J. Hered. 1956, 47, 123–128. [Google Scholar] [CrossRef] [Green Version]
- Bedell, M.A.; Brannan, C.I.; Evans, E.P.; Copeland, N.G.; Jenkins, N.A.; Donovan, P.J. DNA rearrangements located over 100 kb 5’ of the Steel (Sl)-coding region in Steel-panda and Steel-contrasted mice deregulate Sl expression and cause female sterility by disrupting ovarian follicle development. Genes Dev. 1995, 9, 455–470. [Google Scholar] [CrossRef] [Green Version]
- Song, X.; Xu, C.; Liu, Z.; Yue, Z.; Liu, L.; Yang, T.; Cong, B.; Yang, F. Comparative Transcriptome Analysis of Mink (Neovison vison) Skin Reveals the Key Genes Involved in the Melanogenesis of Black and White Coat Colour. Sci. Rep. 2017, 7, 12461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, S.; Li, J.; Ma, T.; Li, J.; Li, Y.; Jiang, H.; Zhang, Q. MiR-27a regulates WNT3A and KITLG expression in Cashmere goats with different coat colors. Anim. Biotechnol. 2019. [Google Scholar] [CrossRef]
- Karyadi, D.M.; Karlins, E.; Decker, B.; vonHoldt, B.M.; Carpintero-Ramirez, G.; Parker, H.G.; Wayne, R.K.; Ostrander, E.A. A copy number variant at the KITLG locus likely confers risk for canine squamous cell carcinoma of the digit. PLoS Genet. 2013, 9, e1003409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duffy, D.L.; Zhu, G.; Li, X.; Sanna, M.; Iles, M.M.; Jacobs, L.C.; Evans, D.M.; Yazar, S.; Beesley, J.; Law, M.H.; et al. Novel pleiotropic risk loci for melanoma and nevus density implicate multiple biological pathways. Nat. Commun. 2018, 9, 4774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landero-Huerta, D.A.; Vigueras-Villasenor, R.M.; Yokoyama-Rebollar, E.; Arechaga-Ocampo, E.; Rojas-Castaneda, J.C.; Jimenez-Trejo, F.; Chavez-Saldana, M. Epigenetic and risk factors of testicular germ cell tumors: A brief review. Front. Biosci. (Landmark. Ed.) 2017, 22, 1073–1098. [Google Scholar] [PubMed] [Green Version]
- Yang, S.; Li, W.S.; Dong, F.; Sun, H.M.; Wu, B.; Tan, J.; Zou, W.J.; Zhou, D.S. KITLG is a novel target of miR-34c that is associated with the inhibition of growth and invasion in colorectal cancer cells. J. Cell. Mol. Med. 2014, 18, 2092–2102. [Google Scholar] [CrossRef]
- Zeron-Medina, J.; Wang, X.; Repapi, E.; Campbell, M.R.; Su, D.; Castro-Giner, F.; Davies, B.; Peterse, E.F.; Sacilotto, N.; Walker, G.J.; et al. A polymorphic p53 response element in KIT ligand influences cancer risk and has undergone natural selection. Cell 2013, 155, 410–422. [Google Scholar] [CrossRef] [Green Version]
- Belluco, S.; Brisebard, E.; Watrelot, D.; Pillet, E.; Marchal, T.; Ponce, F. Digital squamous cell carcinoma in dogs: Epidemiological, histological, and immunohistochemical study. Vet. Pathol. 2013, 50, 1078–1082. [Google Scholar] [CrossRef] [Green Version]
- Williamson, S.H.; Hubisz, M.J.; Clark, A.G.; Payseur, B.A.; Bustamante, C.D.; Nielsen, R. Localizing recent adaptive evolution in the human genome. PLoS Genet. 2007, 3, e90. [Google Scholar] [CrossRef]
- Boyko, A.R.; Quignon, P.; Li, L.; Schoenebeck, J.J.; Degenhardt, J.D.; Lohmueller, K.E.; Zhao, K.; Brisbin, A.; Parker, H.G.; vonHoldt, B.M.; et al. A simple genetic architecture underlies morphological variation in dogs. PLoS Biol. 2010, 8, e1000451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaysse, A.; Ratnakumar, A.; Derrien, T.; Axelsson, E.; Rosengren Pielberg, G.; Sigurdsson, S.; Fall, T.; Seppala, E.H.; Hansen, M.S.; Lawley, C.T.; et al. Identification of genomic regions associated with phenotypic variation between dog breeds using selection mapping. PLoS Genet. 2011, 7, e1002316. [Google Scholar] [CrossRef] [PubMed] [Green Version]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Weich, K.; Affolter, V.; York, D.; Rebhun, R.; Grahn, R.; Kallenberg, A.; Bannasch, D. Pigment Intensity in Dogs is Associated with a Copy Number Variant Upstream of KITLG. Genes 2020, 11, 75. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11010075
Weich K, Affolter V, York D, Rebhun R, Grahn R, Kallenberg A, Bannasch D. Pigment Intensity in Dogs is Associated with a Copy Number Variant Upstream of KITLG. Genes. 2020; 11(1):75. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11010075
Chicago/Turabian StyleWeich, Kalie, Verena Affolter, Daniel York, Robert Rebhun, Robert Grahn, Angelica Kallenberg, and Danika Bannasch. 2020. "Pigment Intensity in Dogs is Associated with a Copy Number Variant Upstream of KITLG" Genes 11, no. 1: 75. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11010075