Centric Fusions behind the Karyotype Evolution of Neotropical Nannostomus Pencilfishes (Characiforme, Lebiasinidae): First Insights from a Molecular Cytogenetic Perspective

 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Chromosome Preparation and C-Banding
2.3. Repetitive DNA Mapping with Fluorescence In Situ Hybridization (FISH)
2.4. Comparative Genomic Hybridization (CGH)
2.5. Microscopy and Image Processing
3. Results
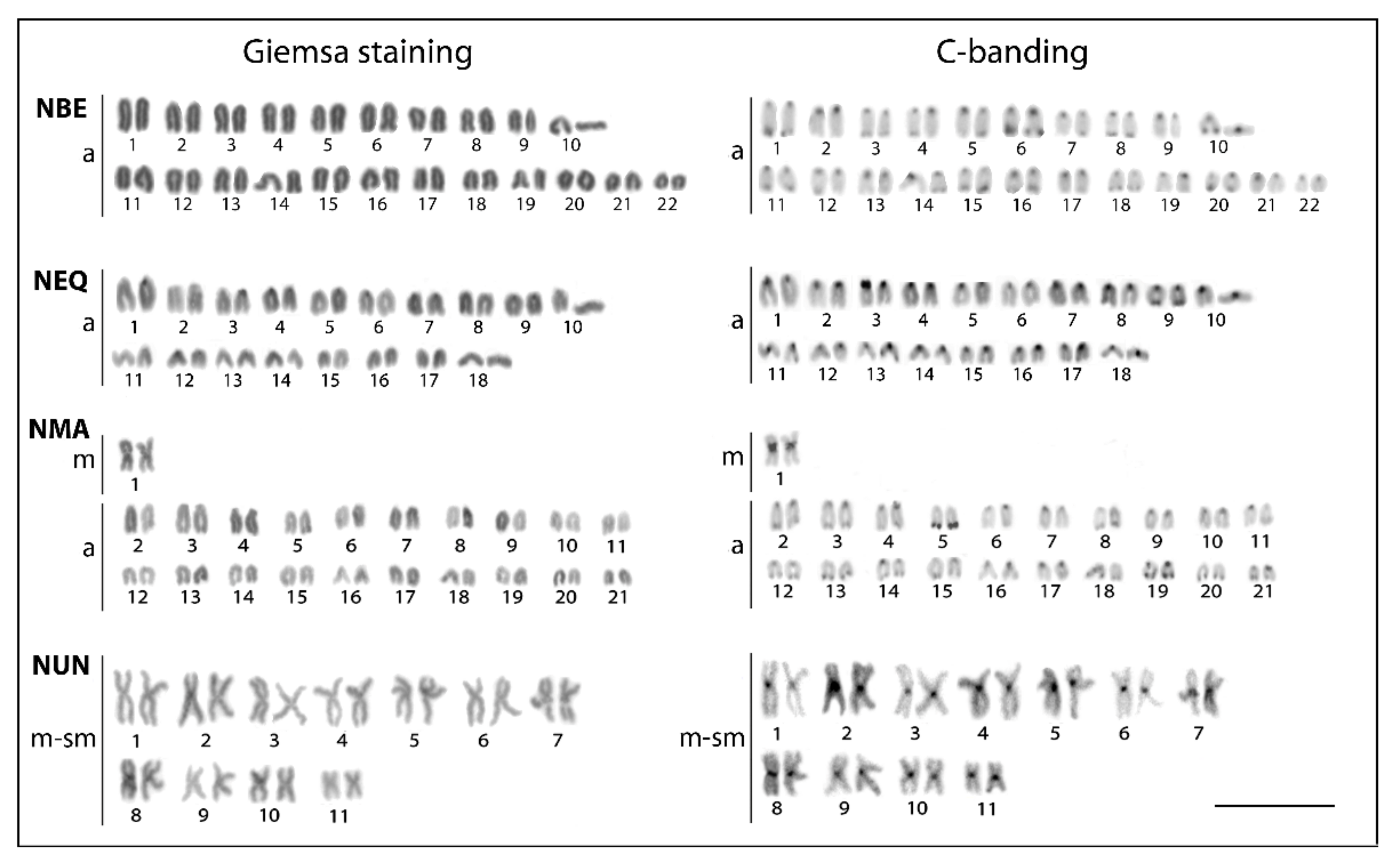
3.1. Conventional Cytogenetic Characteristics
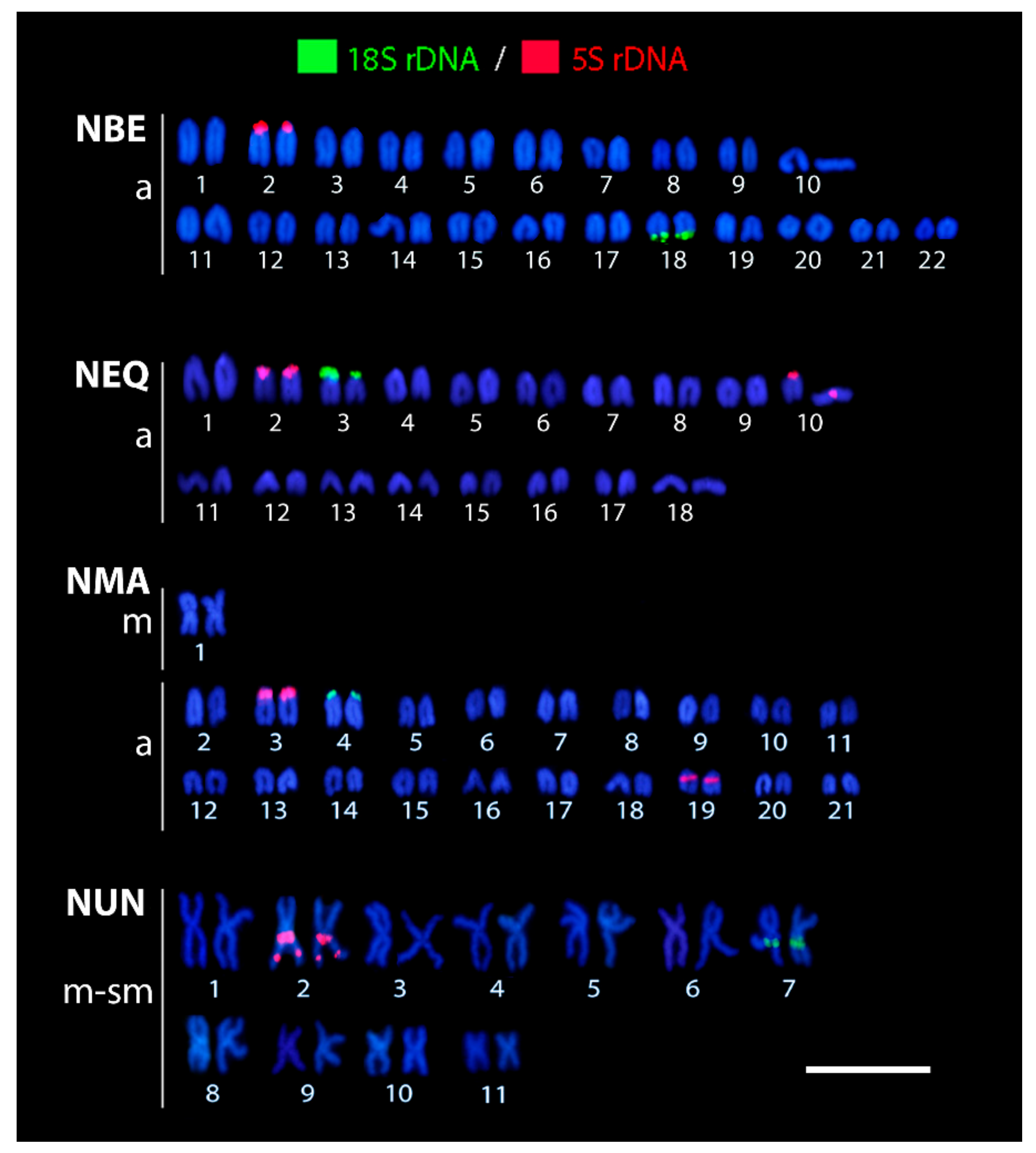
3.2. Patterns of 5S and 18S rDNA Distribution as Revealed by FISH
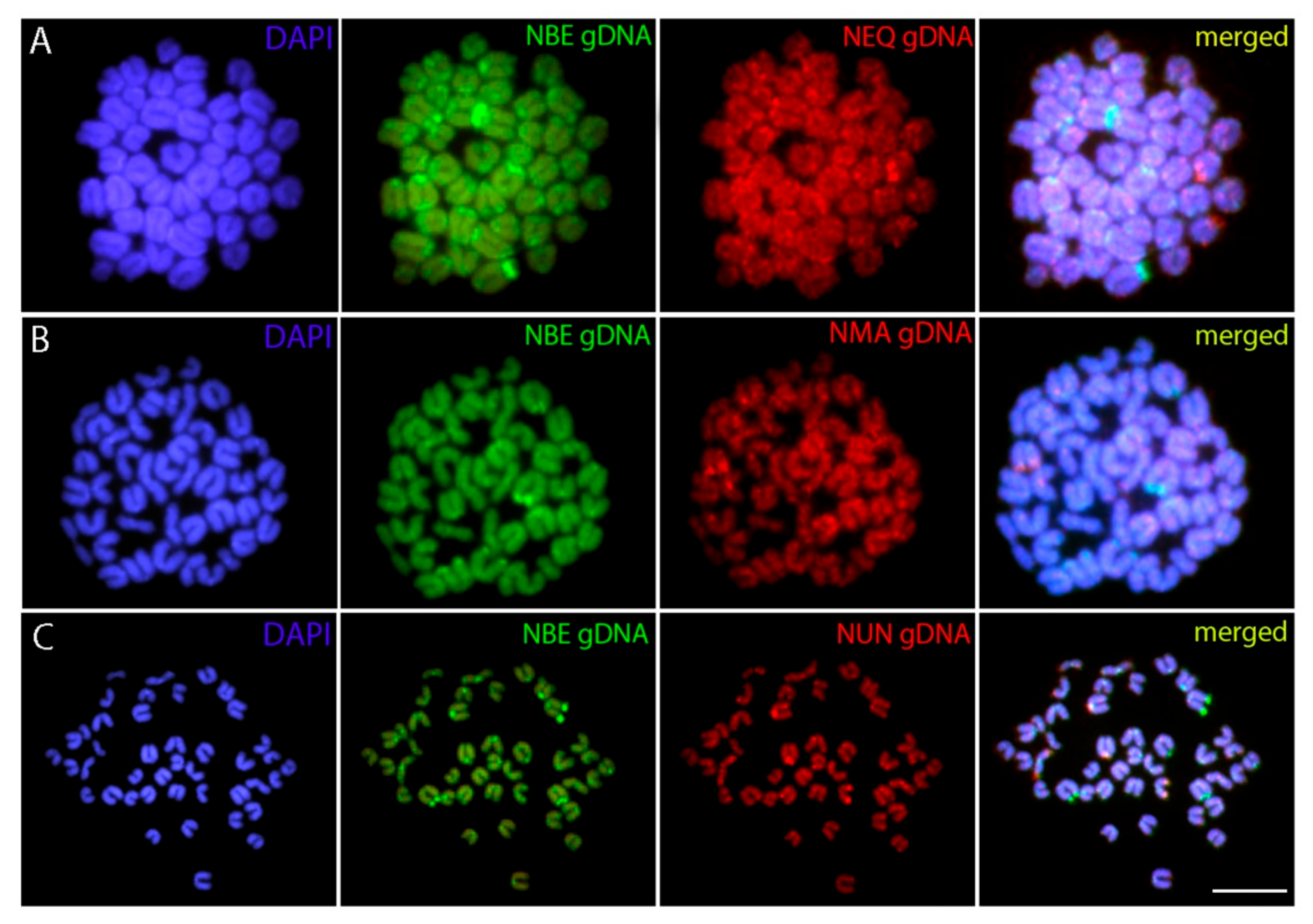
3.3. Patterns of Interspecific Genome Divergence as Revealed by CGH
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Albert, J.S.; Reis, R.E. Historical Biogeography of Neotropical Freshwater Fishes, 1st ed.; University of California Press: Berkeley, CA, USA, 2011. [Google Scholar]
- Nelson, J.S.; Grande, T.C.; Wilson, M.V.H. Fishes of the World, 5th ed.; John Wiley & Sons: Hoboken, NJ, USA, 2016. [Google Scholar]
- Reis, R.E.; Albert, J.S.; Di Dario, F.; Mincarone, M.M.; Petry, P.; Rocha, L.A. Fish biodiversity and conservation in South America. J. Fish Biol. 2016, 89, 12–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, L.H.G.; Hanner, R.; Foresti, F.; Oliveira, C. Can DNA barcoding accurately discriminate megadiverse Neotropical freshwater fish fauna? BMC Genet. 2013, 14, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, M.; Kavalco, K.F.; de Almeida-Toledo, L.F.; Garcia, C. Cryptic diversity between two Imparfinis species (Siluriformes, Heptapteridae) by cytogenetic analysis and DNA barcoding. Zebrafish 2014, 11, 306–317. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, M.; Garcia, C.; Matoso, D.A.; de Jesus, I.S.; Cioffi, M.B.; Bertollo, L.A.C.; Zuanon, J.; Feldberg, E. The Bunocephalus coracoideus species complex (Siluriformes, Aspredinidae). Signs of a speciation process through chromosomal, genetic and ecological diversity. Front. Genet. 2017, 8, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Ramirez, J.L.; Birindelli, J.L.; Carvalho, D.C.; Affonso, P.R.A.M.; Venere, P.C.; Ortega, H.; Carrillo-Avila, M.; Rodríguez-Pulido, J.A.; Galetti, P.M., Jr. Revealing hidden diversity of the underestimated neotropical ichthyofauna: DNA barcoding in the recently described genus Megaleporinus (Characiformes: Anostomidae). Front. Genet. 2017, 8, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Prizon, A.C.; Bruschi, D.P.; Borin-Carvalho, L.A.; Cius, A.; Barbosa, L.M.; Ruiz, H.B.; Zawadzki, C.H.; Fenocchio, A.S.; Portela-Castro, A.L.B. Hidden diversity in the populations of the armored catfish Ancistrus Kner, 1854 (Loricariidae, Hypostominae) from the Paraná River Basin revealed by molecular and cytogenetic data. Front. Genet. 2017, 8, 185. [Google Scholar] [CrossRef] [Green Version]
- Cioffi, M.B.; Moreira-Filho, O.; Ráb, P.; Sember, A.; Molina, W.F.; Bertollo, L.A.C. Conventional cytogenetic approaches—Useful and indispensable tools in discovering fish biodiversity. Curr. Genet. Med. Rep. 2018, 6, 176–186. [Google Scholar] [CrossRef]
- Cioffi, M.B.; Molina, W.F.; Artoni, R.F.; Bertollo, L.A.C. Chromosomes as tools for discovering biodiversity—The case of Erythrinidae fish family. In Recent Trends in Cytogenetic Studies—Methodologies Applications, 1st ed.; Tirunilai, P., Ed.; INTECH: London, UK, 2012; Volume 1, pp. 125–146. [Google Scholar]
- Cioffi, M.B.; Yano, C.F.; Sember, A.; Bertollo, L.A.C. Chromosomal evolution in lower vertebrates: Sex chromosomes in Neotropical fishes. Genes 2017, 8, 258. [Google Scholar] [CrossRef]
- Bertollo, L.A.C. Chromosome evolution in the neotropical Erythrinidae fish family: An overview. In Fish Cytogenetics, 1st ed.; Pisano, E., Ed.; CRC Press: Boca Raton, FL, USA, 2007; pp. 195–211. [Google Scholar]
- Catalog of Fishes: Genera, Species, References. Available online: http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp (accessed on 1 October 2019).
- Netto-Ferreira, A.L. Revisão taxonômica e relações interespecíficas de Lebiasininae (Ostariophysi: Characiformes: Lebiasinidae). Ph.D. Thesis, Universidade de São Paulo, São Paulo, Brazil, 2010. [Google Scholar]
- Benzaquem, D.C.; Oliveira, C.; da Silva Batista, J.; Zuanon, J.; Porto, J.I.R. DNA barcoding in pencilfishes (Lebiasinidae: Nannostomus) reveals cryptic diversity across the brazilian Amazon. PLoS ONE 2015, 10, e0112217. [Google Scholar] [CrossRef] [Green Version]
- Scheel, J.J. Fish Chromosomes and Their Evolution; Danmarks Akvarium: Charlottenlund, Denmark, 1973. [Google Scholar]
- Oliveira, C.; Andreata, A.; Almeida-Toledo, L.F.; Toledo Filho, S.A. Karyotype and nucleolus organizer regions of Pyrrhulina cf. australis (Pisces, Characiformes, Lebiasinidae). Rev. Bras. Genet. 1991, 14, 685–690. [Google Scholar]
- Arai, R. Fish Karyotypes: A Check List, 1st ed.; Springer: Tokyo, Japan, 2011. [Google Scholar]
- Moraes, R.L.R.; Bertollo, L.A.C.; Marinho, M.M.F.; Yano, C.F.; Hatanaka, T.; Barby, F.F.; Troy, W.P.; Cioffi, M.B. Evolutionary relationships and cytotaxonomy considerations in the genus Pyrrhulina (Characiformes, Lebiasinidae). Zebrafish 2017, 14, 536–546. [Google Scholar] [CrossRef] [PubMed]
- Moraes, R.L.; Sember, A.; Bertollo, L.A.C.; de Oliveira, E.A.; Ráb, P.; Hatanaka, T.; Marinho, M.M.F.; Liehr, T.; Al-Rikabi, A.B.H.; Feldberg, E.; et al. Comparative cytogenetics and neo-Y formation in small-sized fish species of the genus Pyrrhulina (Characiformes, Lebiasinidae). Front. Genet. 2019, 10, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sassi, F.M.C.; de Oliveira, E.A.; Bertollo, L.A.C.; Nirchio, M.; Hatanaka, T.; Marinho, M.M.F.; Moreira-Filho, O.; Aroutiounian, R.; Liehr, T.; Al-Rikabi, A.B.H.; et al. Chromosomal evolution and evolutionary relationships of Lebiasina species (Characiformes, Lebiasinidae). Int. J. Mol. Sci. 2019, 20, 2944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toma, G.A.; Moraes, R.L.R.; Sassi, F.M.C.; Bertollo, L.A.C.; de Oliveira, E.A.; Ráb, P.; Sember, A.; Liehr, T.; Hatanaka, T.; Viana, P.F.; et al. Cytogenetics of the small-sized fish, Copeina guttata (Characiformes, Lebiasinidae): Novel insights into the karyotype differentiation of the family. PLoS ONE 2019, 14, e0226746. [Google Scholar] [CrossRef] [PubMed]
- Arefjev, V.A. Karyotypic diversity of characid families (Pisces, Characidae). Caryologia 1990, 43, 291–304. [Google Scholar] [CrossRef]
- Bertollo, L.A.C.; Cioffi, M.B.; Moreira-Filho, O. Direct chromosome preparation from Freshwater teleost fishes. In Fish Cytogenetic Techniques (Ray-Fin Fishes and Chondrichthyans), 1st ed.; Ozouf-Costaz, C., Pisano, E., Foresti, F., Toledo, L.F.A., Eds.; CRC Press: Boca Raton, FL, USA, 2015; Volume 1, pp. 21–26. [Google Scholar]
- Sumner, A.T. A simple technique for demonstrating centromeric heterochromatin. Exp. Cell Res. 1972, 75, 304–306. [Google Scholar] [CrossRef]
- Martins, C.; Ferreira, I.A.; Oliveira, C.; Foresti, F.; Galetti, P.M., Jr. A tandemly repetitive centromeric DNA sequence of the fish Hoplias malabaricus (Characiformes: Erythrinidae) is derived from 5S rDNA. Genetica 2006, 127, 133–141. [Google Scholar] [CrossRef]
- Cioffi, M.B.; Martins, C.; Centofante, L.; Jacobina, U.; Bertollo, L.A.C. Chromosomal variability among allopatric populations of Erythrinidae fish Hoplias malabaricus: Mapping of three classes of repetitive DNAs. Cytogenet. Genome Res. 2009, 125, 132–141. [Google Scholar] [CrossRef]
- Yano, C.F.; Bertollo, L.A.C.; Cioffi, M.B. Fish-FISH: Molecular cytogenetics in fish species. In Fluorescence in Situ Hybridization (FISH)—Application Guide, 2nd ed.; Liehr, T., Ed.; Springer: Berlin, Germany, 2017; pp. 429–444. [Google Scholar]
- Sambrook, J.; Russell, D.W. Molecular Cloning: A Laboratory Manual, 3rd ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2001. [Google Scholar]
- Zwick, M.S.; Hanson, R.E.; Islam-Faridi, M.N.; Stelly, D.M.; Wing, R.A.; Price, H.J.; McKnight, T.D. A rapid procedure for the isolation of C0t-1 DNA from plants. Genome 1997, 40, 138–142. [Google Scholar] [CrossRef] [Green Version]
- Sember, A.; Bertollo, L.A.C.; Ráb, P.; Yano, C.F.; Hatanaka, T.; de Oliveira, E.A.; Cioffi, M.B. Sex chromosome evolution and genomic divergence in the fish Hoplias malabaricus (Characiformes, Erythrinidae). Front. Genet. 2018, 9, 1–12. [Google Scholar] [CrossRef]
- Levan, A.; Fredga, K.; Sandberg, A.A. Nomenclature for centromeric position on chromosomes. Hereditas 1964, 52, 201–220. [Google Scholar] [CrossRef]
- Matthey, R. L’ evolution de la formule chromosomiale chez les vertebrees. Experientia 1945, 1, 78–86. [Google Scholar] [CrossRef]
- King, M. Species Evolution: The Role of Chromosome Change, 1st ed.; Cambridge University Press: Cambridge, UK, 1993. [Google Scholar]
- Dobigny, G.; Ducroz, J.-F.; Robinson, T.J.; Volobouev, V. Cytogenetics and cladistics. Syst. Biol. 2004, 53, 470–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, C.; Almeida-Toledo, L.F.; Foresti, F. Karyotypic evolution in Neotropical fishes. In Fish Cytogenetics, 1st ed.; Pisano, E., Ozouf-Costaz, C., Foresti, F., Kapoor, B.G., Eds.; Science Publishers: Enfield, CT, USA, 2007; pp. 111–164. [Google Scholar]
- Souza e Sousa, J.F.; Viana, P.F.; Bertollo, L.A.C.; Cioffi, M.B.; Feldberg, E. Evolutionary relationships among Boulengerella species (Ctenoluciidae, Characiformes): Genomic organization of repetitive DNAs and highly conserved karyotypes. Cytogenet. Genome Res. 2017, 152, 194–203. [Google Scholar] [CrossRef]
- Betancur-R, R.; Arcila, D.; Vari, R.P.; Hughes, L.C.; Oliveira, C.; Sabaj, M.H.; Ortí, G. Phylogenomic incongruence, hypothesis testing, and taxonomic sampling: The monophyly of characiform fishes. Evolution 2019, 73, 329–345. [Google Scholar] [CrossRef]
- De Barros, A.V.; Wolski, M.A.V.; Nogaroto, V.; Almeida, M.C.; Moreira-Filho, O.; Vicari, M.R. Fragile sites, dysfunctional telomere and chromosome fusions: What is 5S rDNA role? Gene 2017, 608, 20–27. [Google Scholar] [CrossRef]
- Cavalcante, M.G.; Eduardo, C.; Carvalho, M.; Nagamachi, Y.; Pieczarka, J.C.; Vicari, M.R.; Noronha, R.C.R. Physical mapping of repetitive DNA suggests 2n reduction in Amazon turtles Podocnemis (Testudines: Podocnemididae). PLoS ONE 2018, 13, e0197536. [Google Scholar] [CrossRef] [Green Version]
- Glugoski, L.; Giuliano-Caetano, L.; Moreira-Filho, O.; Vicari, M.R.; Nogaroto, V. Co-located hAT transposable element and 5S rDNA in an interstitial telomeric sequence suggest the formation of Robertsonian fusion in armored catfish. Gene 2018, 650, 49–54. [Google Scholar] [CrossRef]
- Schweizer, D.; Loidl, J. A model for heterochromatin dispersion and the evolution of C-band patterns. In Chromosomes Today, 1st ed.; Stahl, A., Luciani, J.M., Vagner-Capodano, A.M., Eds.; Springer: Paris, France, 1987; Volume 9, pp. 61–74. [Google Scholar] [CrossRef]
- Ráb, P.; Crossman, E.J.; Reed, K.M.; Rábová, M. Chromosomal characteristics of ribosomal DNA in two extant species of North American mudminows Umbra pygmaea and U. limi (Euteleostei: Umbridae). Cytogenet. Genome Res. 2002, 98, 194–198. [Google Scholar] [CrossRef]
- Cazaux, B.; Catalan, J.; Veyrunes, F.; Douzery, E.J.; Britton-Davidian, J. Are ribosomal DNA clusters rearrangement hotspots? A case study in the genus Mus (Rodentia, Muridae). BMC Evol. Biol. 2011, 11, 124. [Google Scholar] [CrossRef] [Green Version]
- Santos-Pereira, J.M.; Aguilera, A. R loops: New modulators of genome dynamics and function. Nat. Rev. Genet. 2015, 16, 583–597. [Google Scholar] [CrossRef] [PubMed]
- Sawyer, I.A.; Dundr, M. Chromatin loops and causality loops: The influence of RNA upon spatial nuclear architecture. Chromosoma 2017, 126, 541–557. [Google Scholar] [CrossRef]
- Blokhina, Y.P.; Nguyen, A.D.; Draper, B.W.; Burgess, S.M. The telomere bouquet is a hub where meiotic double-strand breaks, synapsis, and stable homolog juxtaposition are coordinated in the zebrafish, Danio rerio. PLoS Genet. 2019, 15, e1007730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Potapova, T.A.; Gerton, J.L. Ribosomal DNA and the nucleolus in the context of genome organization. Chromosome Res. 2019, 27, 109–127. [Google Scholar] [CrossRef] [PubMed]
- Giles, V.; Thode, G.; Alvarez, M.C. A new Robertsonian fusion in the multiple chromosome polymorphism of a mediterranean population of Gobius paganellus (Gobiidae, Perciformes). Heredity 1985, 55, 255–260. [Google Scholar] [CrossRef] [Green Version]
- Molina, W.F.; Galetti, P.M., Jr. Robertsonian rearrangements in the reef fish Chromis (Perciformes, Pomacentridae) involving chromosomes bearing 5S rRNA genes. Genet. Mol. Biol. 2002, 25, 373–377. [Google Scholar] [CrossRef] [Green Version]
- Rosa, K.O.; Ziemniczak, K.; de Barros, A.V.; Nogaroto, V.; Almeida, M.C.; Cestari, M.M.; Artoni, R.F.; Vicari, M.R. Numeric and structural chromosome polymorphism in Rineloricaria lima (Siluriformes: Loricariidae): Fusion points carrying 5S rDNA or telomere sequence vestiges. Rev. Fish Biol. Fish. 2012, 22, 739–749. [Google Scholar] [CrossRef]
- Sember, A.; Bohlen, J.; Šlechtová, V.; Altmanová, M.; Symonová, R.; Ráb, P. Karyotype differentiation in 19 species of river loach fishes (Nemacheilidae, Teleostei): Extensive variability associated with rDNA and heterochromatin distribution and its phylogenetic and ecological interpretation. BMC Evol. Biol. 2015, 15, 251. [Google Scholar] [CrossRef] [Green Version]
- Getlekha, N.; Molina, W.F.; Cioffi, M.B.; Yano, C.F.; Maneechot, N.; Bertollo, L.A.C.; Supiwong, W.; Tanomtong, A. Repetitive DNAs highlight the role of chromosomal fusions in the karyotype evolution of Dascyllus species (Pomacentridae, Perciformes). Genetica 2016, 144, 203–211. [Google Scholar] [CrossRef]
- Salvadori, S.; Deiana, A.M.; Deidda, F.; Lobina, C.; Mulas, A.; Coluccia, E. XX/XY sex chromosome system and chromosome markers in the snake eel Ophisurus serpens (Anguilliformes: Ophichtidae). Mar. Biol. Res. 2018, 14, 158–164. [Google Scholar] [CrossRef]
- Schmid, M.; Steinlein, C.; Bogart, J.P.; Feichtinger, W.; León, P.; La Marca, E.; Díaz, L.M.; Sanz, A.; Chen, S.H.; Hedges, S.B. The chromosomes of terraran frogs: Insights into vertebrate cytogenetics. Cytogenet. Genome Res. 2016, 130–131, 1–568. [Google Scholar] [CrossRef]
- Da Costa, M.J.R.; do Amaral, P.J.S.; Pieczarka, J.C.; Sampaio, M.I.; Rossi, R.V.; Mendes-Oliveira, A.C.; Noronha, R.C.R.; Nagamachi, C.Y. Cryptic species in Proechimys goeldii (Rodentia, Echimyidae)? A case of molecular and chromosomal differentiation in allopatric populations. Cytogenet. Genome Res. 2016, 148, 199–210. [Google Scholar] [CrossRef] [PubMed]
- Houck, M.L.; Teri, L.; Lear, T.L.; Charter, S.J. Animal cytogenetics. In The AGT Cytogenetics Laboratory Manual, 4th ed.; Arsham, M.S., Barch, M.J., Lawce, H.J., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2017; pp. 1055–1102. [Google Scholar]
- Sousa, R.P.C.; Oliveira-Filho, A.B.; Vallinoto, M.; Cioffi, M.B.; Molina, W.F.; de Oliveira, E.H.; Silva-Oliveira, G.C. Cytogenetics description in Batrachoides surinamensis, (Batrachoididae: Batrachoidiformes): What does the estuary have to say? Estuar. Coast. Shelf Sci. 2018, 213, 253–259. [Google Scholar] [CrossRef]
- Molina, W.F.; Martinez, P.A.; Bertollo, L.A.C.; Bidau, C.J. Evidence for meiotic drive as an explanation for karyotype changes in fishes. Mar. Genom. 2014, 15, 29–34. [Google Scholar] [CrossRef]
- Krysanov, E.; Demidova, T. Extensive karyotype variability of African fish genus Nothobranchius (Cyprinodontiformes). Comp. Cytogenet. 2018, 12, 387–402. [Google Scholar] [CrossRef]
- Völker, M.; Sonnenberg, R.; Ráb, P.; Kullmann, H. Karyotype differentiation in Chromaphyosemion killifishes (Cyprinodontiformes, Nothobranchiidae) II: Cytogenetic and mitochondrial DNA analyses demonstrate karyotype differentiation and its evolutionary direction in C. riggenbachi. Cytogenet. Genome Res. 2006, 115, 70–83. [Google Scholar] [CrossRef]
- Völker, M.; Ráb, P.; Kullmann, H. Karyotype differentiation in Chromaphyosemion killifishes (Cyprinodontiformes, Nothobranchiidae): Patterns, mechanisms, and evolutionary implications. Biol. J. Linn. Soc. 2008, 94, 143–153. [Google Scholar] [CrossRef]
- Ene, A.-C. Chromosomal polymorphism in the goby Neogobius eurycephalus (Perciformes: Gobiidae). Mar. Biol. 2002, 142, 583–588. [Google Scholar] [CrossRef]
- Amores, A.; Wilson, C.A.; Allard, C.A.H.; Detrich, H.W.; Postlethwait, J.H. Cold fusion: Massive karyotype evolution in the Antarctic bullhead notothen Notothenia coriiceps. G3 (Bethesda) 2017, 7, 2195–2207. [Google Scholar] [CrossRef] [Green Version]
- Crossman, E.J.; Ráb, P. Chromosome-banding study of the Alaska blackfish, Dallia pectoralis (Euteleostei: Esocae) with implications for karyotype evolution and relationships of esocoid fishes. Can. J. Zool. 1996, 74, 147–156. [Google Scholar] [CrossRef]
- Crossman, E.J.; Ráb, P. Chromosomal NOR phenotype and C-banded karyotype of Olympic mudminnow, Novumbra hubbsi (Euteleostei: Umbridae). Copeia 2001, 3, 860–865. [Google Scholar] [CrossRef]
- Phillips, R.; Ráb, P. Chromosome evolution in the Salmonidae (Pisces): An update. Biol. Rev. Camb. Philos. Soc. 2001, 76, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.D.; O’Neill, R.J. Chromosomes, conflict, and epigenetics: Chromosomal speciation revisited. Annu. Rev. Genom. Hum. Genet. 2010, 11, 291–316. [Google Scholar] [CrossRef] [PubMed]
- Guerrero, R.F.; Kirkpatrick, M. Local adaptation and the evolution of chromosome fusions. Evolution 2014, 68, 2747–2756. [Google Scholar] [CrossRef]
- Lanctôt, C.; Cheutin, T.; Cremer, M.; Cavalli, G.; Cremer, T. Dynamic genome architecture in the nuclear space: Regulation of gene expression in three dimensions. Nat. Rev. Genet. 2007, 8, 104–115. [Google Scholar] [CrossRef]
- Meaburn, K.J.; Misteli, T.; Soutoglou, E. Spatial genome organization in the formation of chromosomal translocations. Semin. Cancer Biol. 2007, 17, 80–90. [Google Scholar] [CrossRef] [Green Version]
- Meaburn, K.J.; Misteli, T. Cell biology: Chromosome territories. Nature 2007, 445, 379–781. [Google Scholar] [CrossRef]
- Cremer, T.; Cremer, M. Chromosome territories. Cold Spring Harb. Perspect. Biol. 2010, 2, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Roukos, V.; Misteli, T. The biogenesis of chromosome translocations. Nat. Cell Biol. 2014, 16, 293–300. [Google Scholar] [CrossRef]
- Fraser, J.; Williamson, I.; Bickmore, W.A.; Dostie, J. An overview of genome organization and how we got there: From FISH to Hi-C. Microbiol. Mol. Biol. Rev. 2015, 79, 347–372. [Google Scholar] [CrossRef] [Green Version]
- Razin, S.V.; Gavrilov, A.A.; Vassetzky, Y.S.; Ulianov, S.V. Topologically-associating domains: Gene warehouses adapted to serve transcriptional regulation. Transcription 2016, 7, 84–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosin, L.F.; Crocker, O.; Isenhart, R.L.; Nguyen, S.C.; Xu, Z.; Joyce, E.F. Chromosome territory formation attenuates the translocation potential of cells. eLife 2019, 8, e49553. [Google Scholar] [CrossRef] [PubMed]
- Ghavi-Helm, Y.; Jankowski, A.; Meiers, S.; Viales, R.R.; Korbel, J.O.; Furlong, E.E.M. Highly rearranged chromosomes reveal uncoupling between genome topology and gene expression. Nat. Genet. 2019, 51, 1272–1282. [Google Scholar] [CrossRef] [PubMed]
- Mank, J.E.; Avise, J.C. Phylogenetic conservation of chromosome numbers in Actinopterygiian fishes. Genetica 2006, 127, 321–327. [Google Scholar] [CrossRef] [PubMed]
- Cioffi, M.B.; Bertollo, L.A.C. Chromosomal distribution and evolution of repetitive DNAs in fish. Genome Dyn. 2012, 7, 197–221. [Google Scholar] [CrossRef]
- García-Souto, D.; Qarkaxhija, V.; Pasantes, J.J. Resolving the taxonomic status of Chamelea gallina and C. striatula (Veneridae, Bivalvia): A combined molecular cytogenetic and phylogenetic approach. Biomed. Res. Int. 2017, 2017, 7638790. [Google Scholar] [CrossRef] [Green Version]
- Weeks, A.R.; Marec, F.; Breeuwer, J.A.J. A mite species that consists entirely of haploid females. Science 2001, 292, 2479–2482. [Google Scholar] [CrossRef]
- Lukhtanov, V.A.; Dincă, V.; Friberg, M.; Šíchová, J.; Olofsson, M.; Vila, R.; Marec, F.; Wiklund, C. Versatility of multivalent orientation, inverted meiosis, and rescued fitness in holocentric chromosomal hybrids. PNAS 2018, 115, E9610–E9619. [Google Scholar] [CrossRef] [Green Version]
- Do Nascimento, V.D.; Coelho, K.A.; Nogaroto, V.; Almeida, R.B.; Ziemniczak, K.; Centofante, L.; Pavanelli, C.S.; Torres, R.A.; Moreira-Filho, O.; Vicari, M.R. Do multiple karyomorphs and population genetics of freshwater darter characines (Apareiodon affinis) indicate chromosomal speciation? Zool. Anz. 2018, 272, 93–103. [Google Scholar] [CrossRef]
- Zhu, H.P.; Ma, D.M.; Gui, J.F. Triploid origin of the gibel carp as revealed by 5S rDNA localization and chromosome painting. Chromosome Res. 2006, 14, 767–776. [Google Scholar] [CrossRef]
- Zhang, C.; Ye, L.; Chen, Y.; Xiao, J.; Wu, Y.; Tao, M.; Xiao, Y.; Liu, S. The chromosomal constitution of fish hybrid lineage revealed by 5S rDNA FISH. BMC Genet. 2015, 16, 140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soto, M.Á.; Castro, J.P.; Walker, L.I.; Malabarba, L.R.; Santos, M.H.; Almeida, M.C.; Moreira-Filho, O.; Artoni, R.F. Evolution of trans-Andean endemic fishes of the genus Cheirodon (Teleostei: Characidae) are associated with chromosomal rearrangements. Rev. Chil. Hist. Nat. 2018, 91, 8. [Google Scholar] [CrossRef] [Green Version]
- Yano, C.F.; Bertollo, L.A.C.; Ezaz, T.; Trifonov, V.; Sember, A.; Liehr, T.; Cioffi, M.B. Highly conserved Z and molecularly diverged W chromosomes in the fish genus Triportheus (Characiformes, Triportheidae). Heredity 2017, 118, 276–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Oliveira, E.A.; Sember, A.; Bertollo, L.A.C.; Yano, C.F.; Ezaz, T.; Moreira-Filho, O.; Hatanaka, T.; Trifonov, V.; Liehr, T.; Al-Rikabi, A.B.H.; et al. Tracking the evolutionary pathway of sex chromosomes among fishes: Characterizing the unique XX/XY1Y2 system in Hoplias malabaricus (Teleostei, Characiformes). Chromosoma 2018, 127, 115–128. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Sember, A.; Zhu, Q.; de Oliveira, E.A.; Liehr, T.; Al-Rikabi, A.B.H.; Xiao, Z.; Song, H.; Cioffi, M.B. Deciphering the origin and evolution of the X1X2Y system in two closely-related Oplegnathus species (Oplegnathidae and Centrarchiformes). J. Mol. Sci. 2019, 20, 3571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Symonová, R.; Majtánová, Z.; Sember, A.; Staaks, G.B.; Bohlen, J.; Freyhof, J.; Rábová, M.; Ráb, P. Genome differentiation in a species pair of coregonine fishes: An extremely rapid speciation driven by stress-activated retrotransposons mediating extensive ribosomal DNA multiplications. BMC Evol. Biol. 2013, 13, 42. [Google Scholar] [CrossRef] [Green Version]
- Da Silva, A.F.; Feldberg, E.; Carvalho, N.D.M.; Rangel, S.M.H.; Schneider, C.H.; Carvalho-Zilse, G.A.; da Silva, V.F.; Gross, M.C. Effects of environmental pollution on the rDNAomics of Amazonian fish. Environ. Pollut. 2019, 252, 180–187. [Google Scholar] [CrossRef]
- Gornung, E. Twenty years of physical mapping of major ribosomal RNA genes across the teleosts: A review of research. Cytogenet. Genome Res. 2013, 141, 90–102. [Google Scholar] [CrossRef]
- Sochorová, J.; Garcia, S.; Gálvez, F.; Symonová, R.; Kovařík, A. Evolutionary trends in animal ribosomal DNA loci: Introduction to a new online database. Chromosoma 2018, 127, 141–150. [Google Scholar] [CrossRef] [Green Version]
- Milhomem, S.S.R.; Scacchetti, P.C.; Pieczarka, J.C.; Ferguson-Smith, M.A.; Pansonato-Alves, J.C.; O’Brien, P.C.M.; Foresti, F.; Nagamachi, C.Y. Are NORs always located on homeologous chromosomes? A FISH investigation with rDNA and whole chromosome probes in Gymnotus fishes (Gymnotiformes). PLoS ONE 2013, 8, e55608. [Google Scholar] [CrossRef]
- Lim, K.Y.; Kovařík, A.; Matyášek, R.; Chase, M.W.; Clarkson, J.J.; Grandbastien, M.A.; Leitch, A.R. Sequence of events leading to near-complete genome turnover in allopolyploid Nicotiana within five million years. New Phytol. 2007, 175, 756–763. [Google Scholar] [CrossRef] [PubMed]
- Majka, J.; Majka, M.; Kwiatek, M.; Wiśniewska, H. Similarities and differences in the nuclear genome organization within Pooideae species revealed by comparative genomic in situ hybridization (GISH). J. Appl. Genet. 2017, 58, 151–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barby, F.F.; Bertollo, L.A.C.; de Oliveira, E.A.; Yano, C.F.; Hatanaka, T.; Ráb, P.; Sember, A.; Ezaz, T.; Artoni, R.F.; Liehr, T.; et al. Emerging patterns of genome organization in Notopteridae species (Teleostei, Osteoglossiformes) as revealed by Zoo-FISH and Comparative Genomic Hybridization (CGH). Sci. Rep. 2019, 9, 1112. [Google Scholar] [CrossRef] [PubMed]
- Kandul, N.P.; Lukhtanov, V.A.; Pierce, N.E. Karyotypic diversity and speciation in Agrodiaetus butterflies. Evolution 2007, 61, 546–559. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Sun, X.; Cormack, B.P.; Boeke, J.D. Karyotype engineering by chromosome fusion leads to reproductive isolation in yeast. Nature 2018, 560, 392–396. [Google Scholar] [CrossRef]
- Ortiz-Barrientos, D.; Engelstädter, J.; Rieseberg, L.H. Recombination rate evolution and the origin of species. Trends Ecol. Evol. 2016, 31, 226–236. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | 2n | Sampling Site | N |
|---|---|---|---|
| Nannostomus beckfordi | 44 | Agenor Stream (Amazon River), AM | (09♀ 17♂) |
| Nannostomus eques | 36 | Cuieiras River, AM | (15♀ 12♂) |
| Nannostomus marginatus | 42 | Adolpho Ducke reserve (Negro River), AM | (08♀ 12♂) |
| Nannostomus unifasciatus | 22 | Cuieiras River, AM | (09♀ 13♂) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sember, A.; de Oliveira, E.A.; Ráb, P.; Bertollo, L.A.C.; de Freitas, N.L.; Viana, P.F.; Yano, C.F.; Hatanaka, T.; Marinho, M.M.F.; de Moraes, R.L.R.; et al. Centric Fusions behind the Karyotype Evolution of Neotropical Nannostomus Pencilfishes (Characiforme, Lebiasinidae): First Insights from a Molecular Cytogenetic Perspective. Genes 2020, 11, 91. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11010091
Sember A, de Oliveira EA, Ráb P, Bertollo LAC, de Freitas NL, Viana PF, Yano CF, Hatanaka T, Marinho MMF, de Moraes RLR, et al. Centric Fusions behind the Karyotype Evolution of Neotropical Nannostomus Pencilfishes (Characiforme, Lebiasinidae): First Insights from a Molecular Cytogenetic Perspective. Genes. 2020; 11(1):91. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11010091
Chicago/Turabian StyleSember, Alexandr, Ezequiel Aguiar de Oliveira, Petr Ráb, Luiz Antonio Carlos Bertollo, Natália Lourenço de Freitas, Patrik Ferreira Viana, Cassia Fernanda Yano, Terumi Hatanaka, Manoela Maria Ferreira Marinho, Renata Luiza Rosa de Moraes, and et al. 2020. "Centric Fusions behind the Karyotype Evolution of Neotropical Nannostomus Pencilfishes (Characiforme, Lebiasinidae): First Insights from a Molecular Cytogenetic Perspective" Genes 11, no. 1: 91. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11010091