Trypanosoma Cruzi Genome: Organization, Multi-Gene Families, Transcription, and Biological Implications

 , ,
, ,
Abstract
:1. General Aspects of T. cruzi Biology
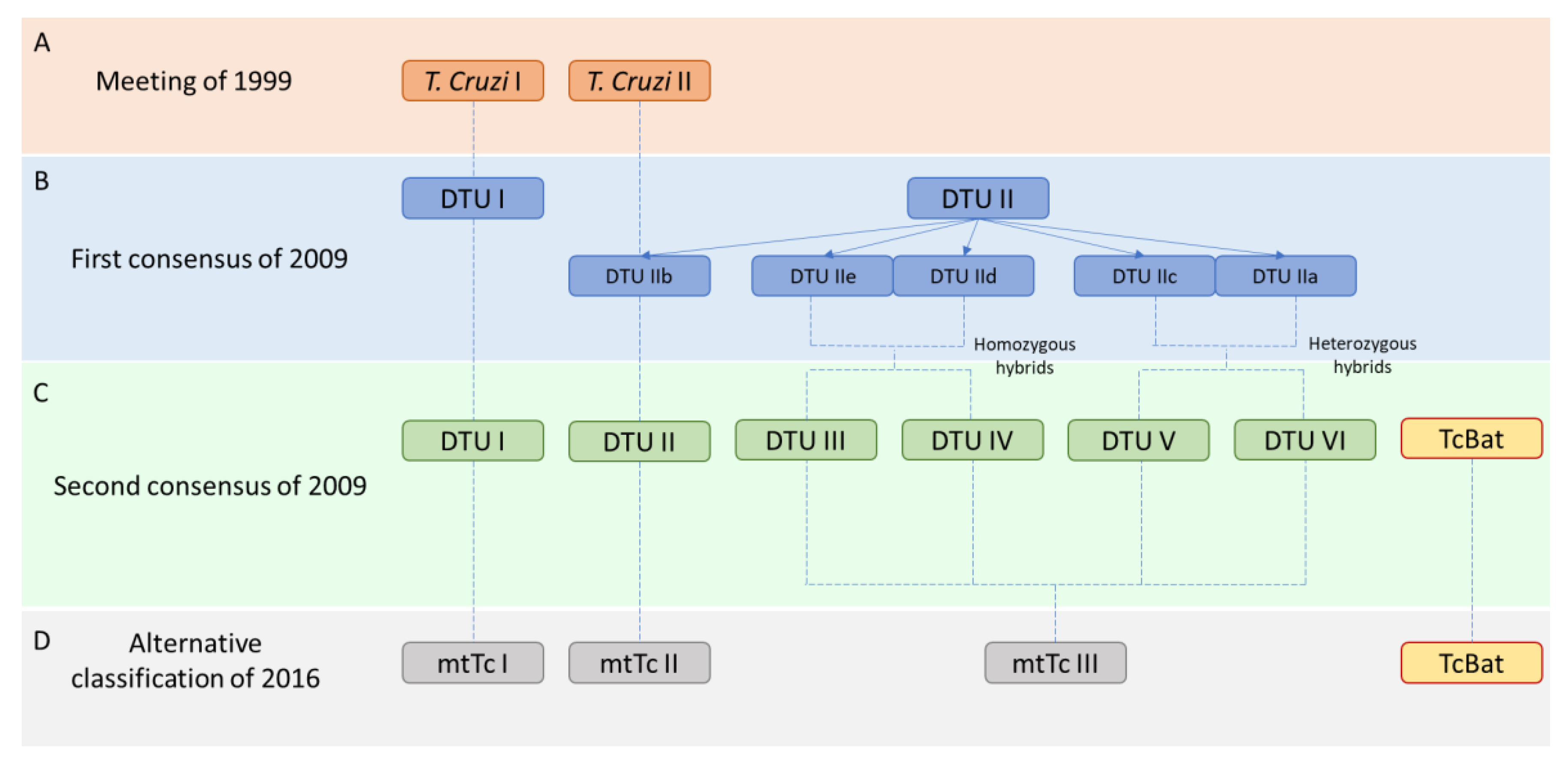
2. Classification of T. cruzi Strains
3. The Genomes of T. cruzi: A New Update
4. Genetic Diversity and Genome Structure of T. cruzi
4.1. Ploidy
4.2. Genome Composition
4.3. Telomeric Regions
4.4. G + C Content
4.5. Replication Origin
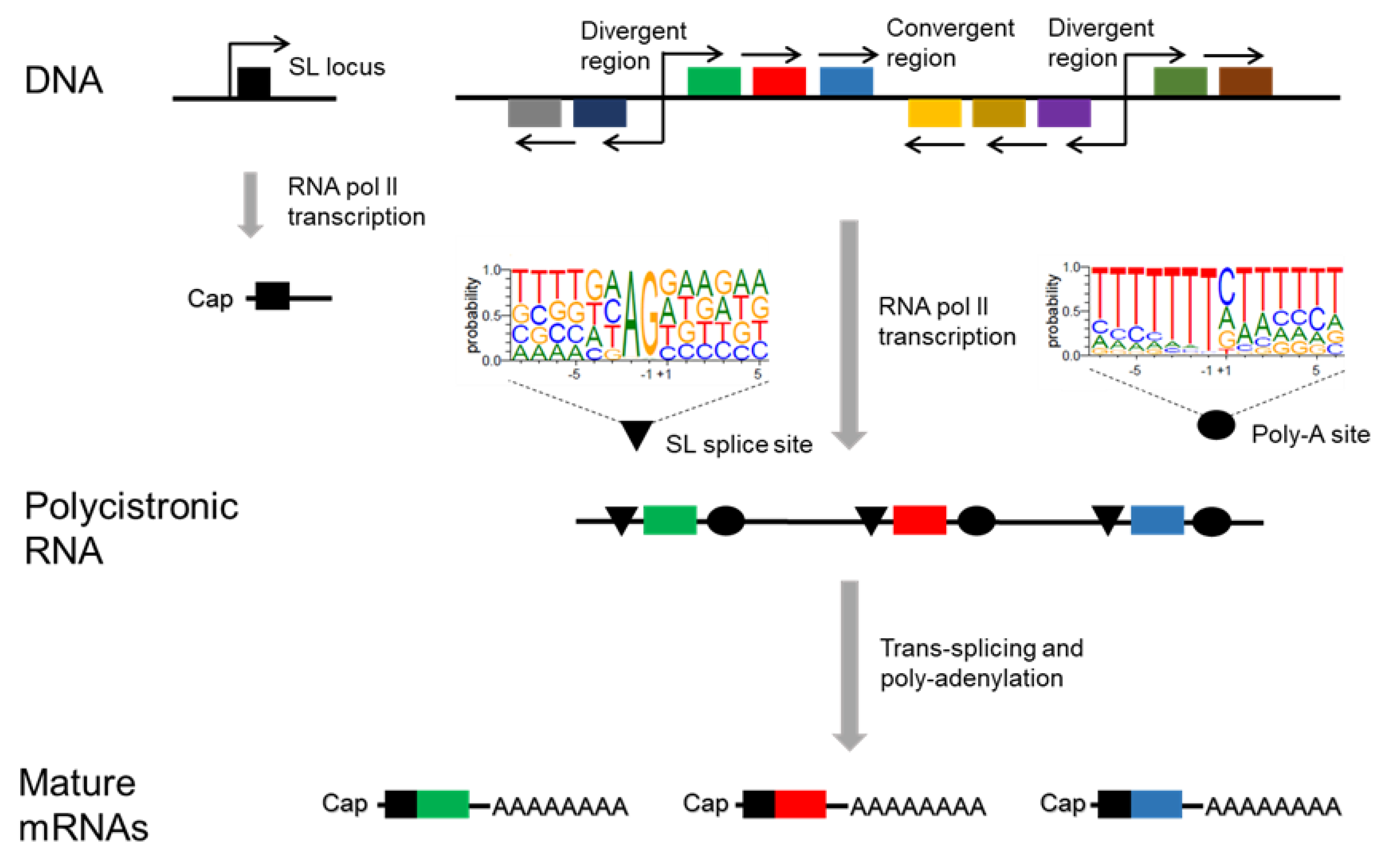
5. Transcription of T. cruzi
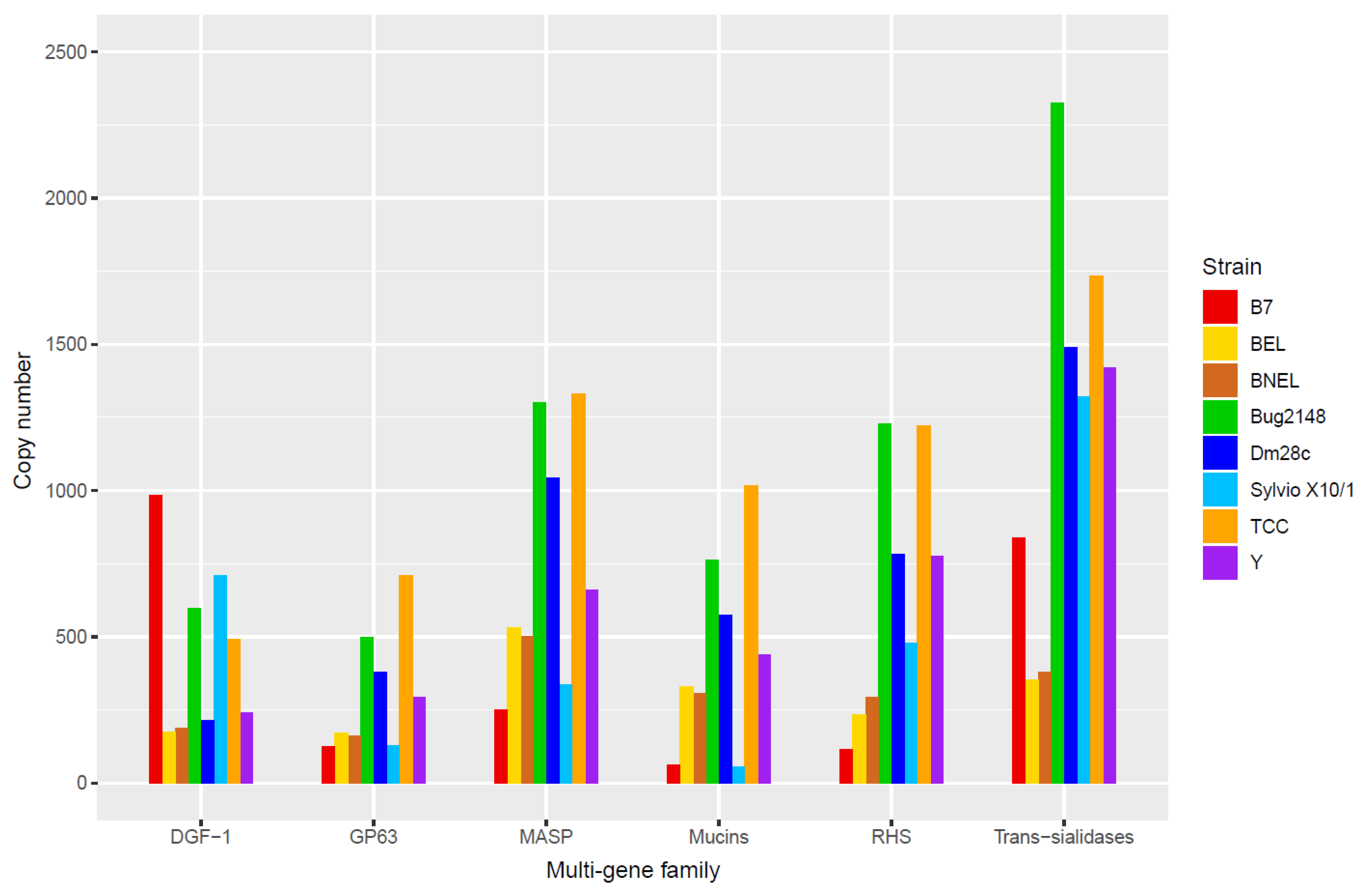
6. Principal Multi-Gene Families of T. cruzi
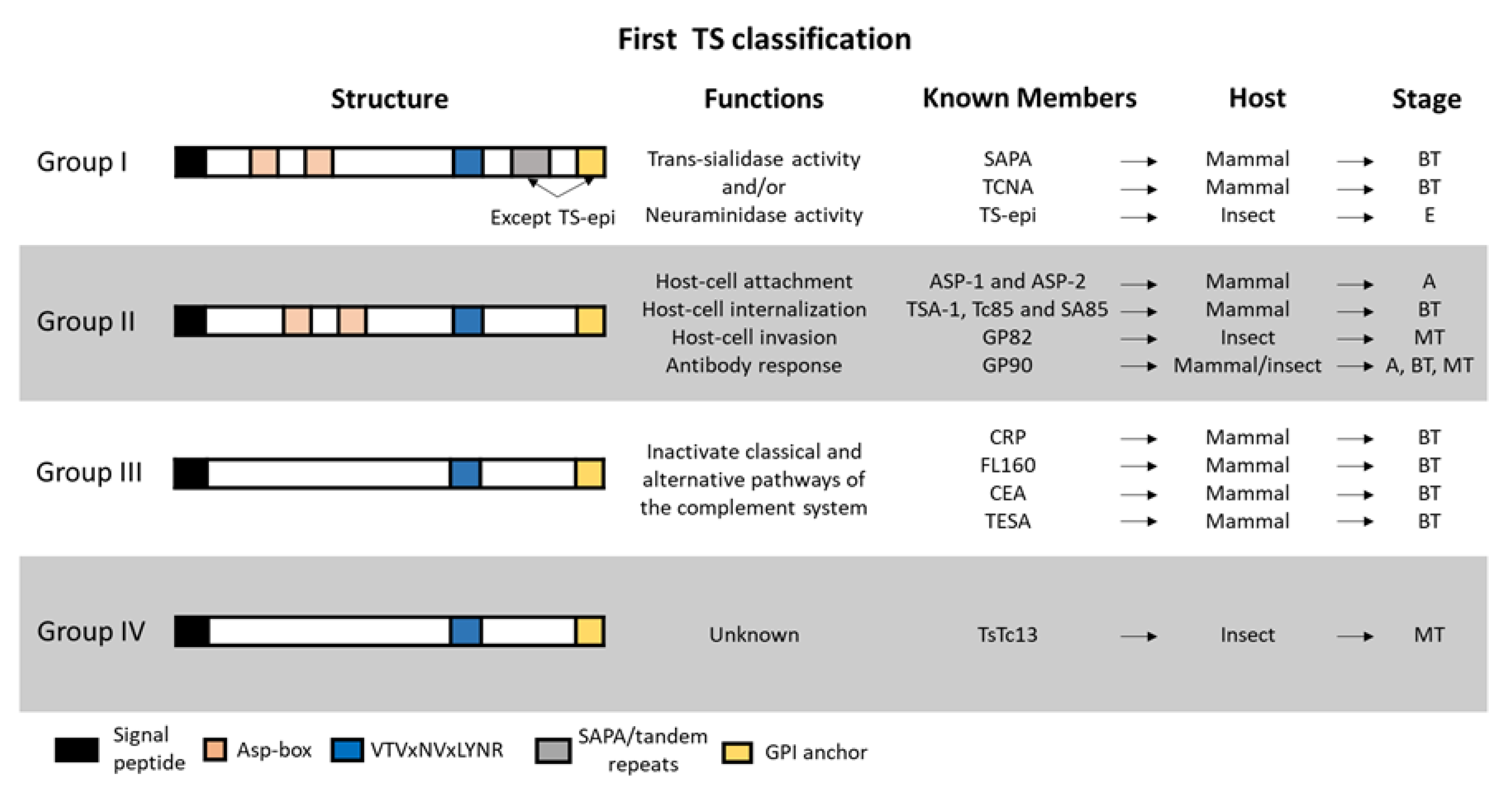
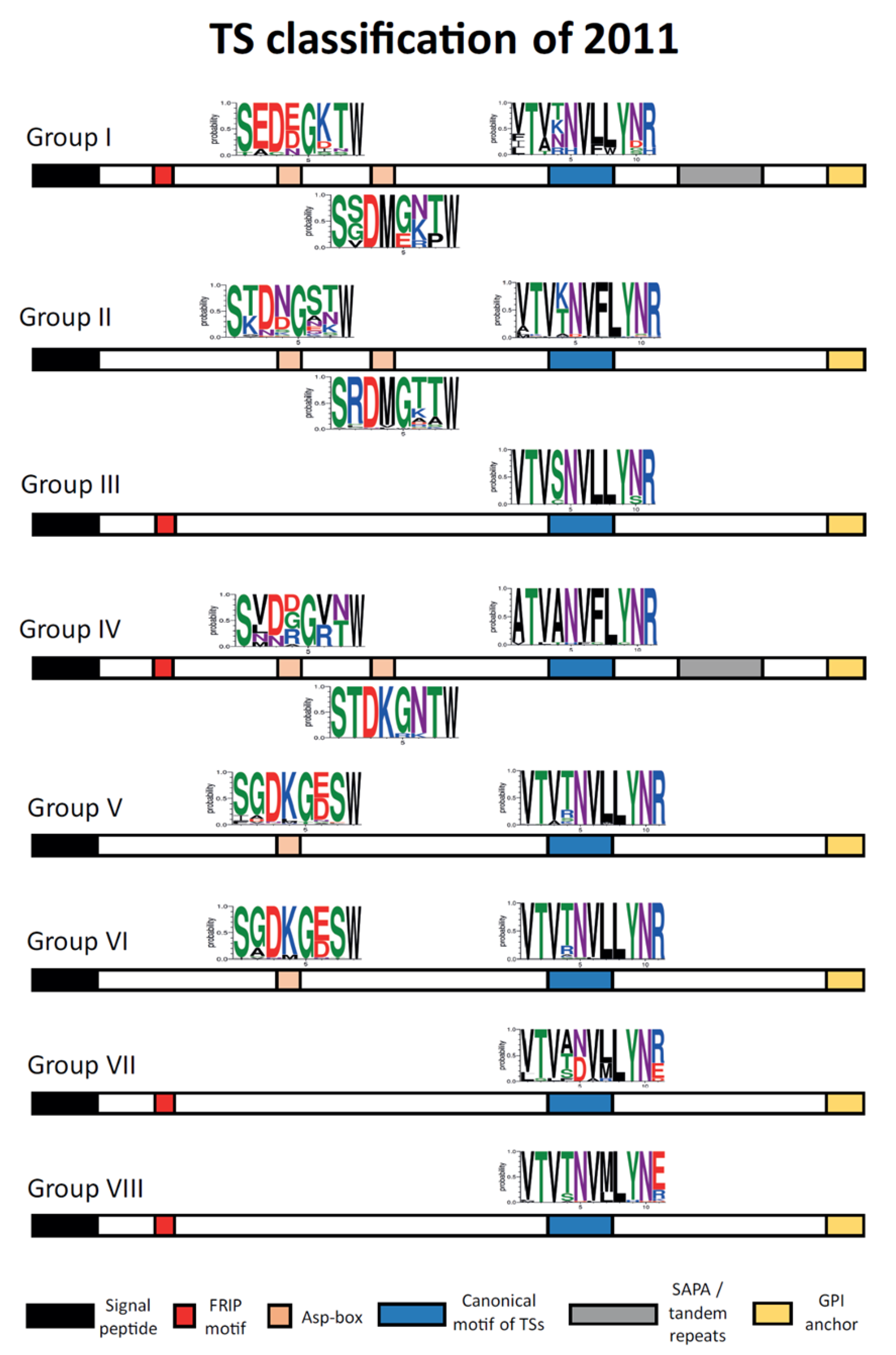
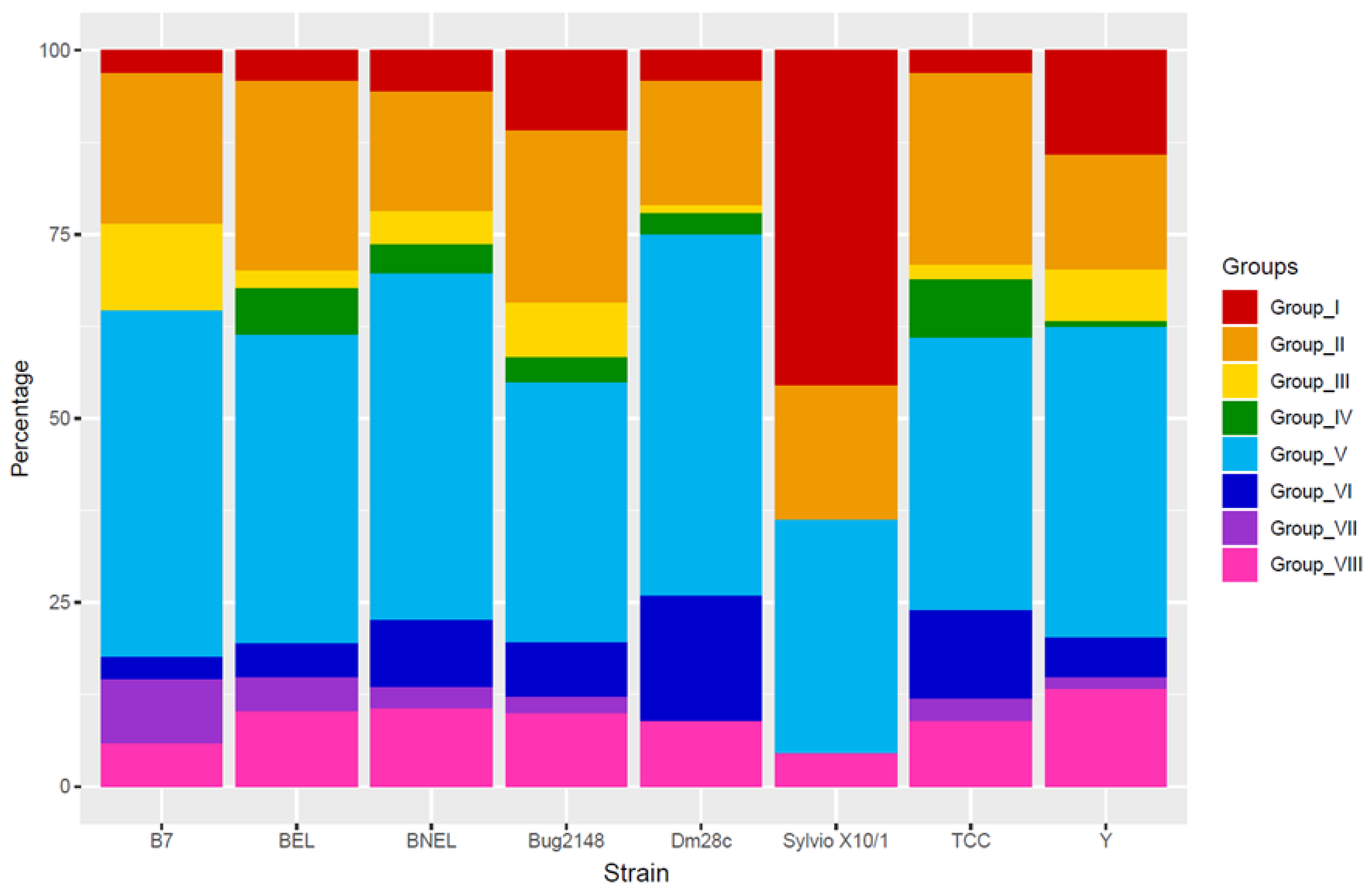
6.1. Trans-Sialidase (TS) Family
6.2. Mucins
6.3. MASPs
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Maslov, D.A.; Opperdoes, F.R.; Kostygov, A.Y.; Hashimi, H.; Lukeš, J.; Yurchenko, V. Recent advances in trypanosomatid research: Genome organization, expression, metabolism, taxonomy and evolution. Parasitology 2019, 146, 1–27. [Google Scholar] [CrossRef] [Green Version]
- Lukeš, J.; Butenko, A.; Hashimi, H.; Maslov, D.A.; Votýpka, J.; Yurchenko, V. Trypanosomatids Are Much More than Just Trypanosomes: Clues from the Expanded Family Tree. Trends Parasitol. 2018, 34, 466–480. [Google Scholar] [CrossRef] [Green Version]
- Rassi, A.; Rassi, A.; Marcondes de Rezende, J. American trypanosomiasis (Chagas disease). Infect. Dis. Clin. N. Am. 2012, 26, 275–291. [Google Scholar] [CrossRef] [PubMed]
- Andrade, L.O.; Machado, C.R.S.; Chiari, E.; Pena, S.D.J.; Macedo, A.M. Trypanosoma cruzi: Role of host genetic background in the differential tissue distribution of parasite clonal populations. Exp. Parasitol. 2002, 100, 269–275. [Google Scholar] [CrossRef]
- Manoel-Caetano, F.S.; Silva, A.E. Implications of genetic variability of Trypanosoma cruzi for the pathogenesis of Chagas disease. Cad. Saúde Pública 2007, 23, 2263–2274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez, H.O.; Guerrero, N.A.; Fortes, A.; Santi-Rocca, J.; Gironès, N.; Fresno, M. Trypanosoma cruzi strains cause different myocarditis patterns in infected mice. Acta Trop. 2014, 139, 57–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santi-Rocca, J.; Fernandez-Cortes, F.; Chillón-Marinas, C.; González-Rubio, M.-L.; Martin, D.; Gironès, N.; Fresno, M. A multi-parametric analysis of Trypanosoma cruzi infection: Common pathophysiologic patterns beyond extreme heterogeneity of host responses. Sci. Rep. 2017, 7, 8893. [Google Scholar] [CrossRef]
- Clayton, J. Chagas disease 101. Nature 2010, 465, S4–S5. [Google Scholar] [CrossRef]
- De Souza, W.; de Carvalho, T.M.U.; Barrias, E.S. Review on Trypanosoma cruzi: Host Cell Interaction. Int. J. Cell. Biol. 2010, 2010. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, J.C.F.; Godinho, J.L.P.; de Souza, W. Biology of human pathogenic trypanosomatids: Epidemiology, lifecycle and ultrastructure. Subcell. Biochem. 2014, 74, 1–42. [Google Scholar] [CrossRef]
- Echeverria, L.E.; Morillo, C.A. American Trypanosomiasis (Chagas Disease). Infect. Dis. Clin. N. Am. 2019, 33, 119–134. [Google Scholar] [CrossRef] [PubMed]
- Lukes, J.; Guilbride, D.L.; Votýpka, J.; Zíková, A.; Benne, R.; Englund, P.T. Kinetoplast DNA network: Evolution of an improbable structure. Eukaryot. Cell 2002, 1, 495–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, S.; Martinez, L.L.I.T.; Westenberger, S.J.; Sturm, N.R. A population study of the minicircles in Trypanosoma cruzi: Predicting guide RNAs in the absence of empirical RNA editing. BMC Genom. 2007, 8, 133. [Google Scholar] [CrossRef] [Green Version]
- Gerasimov, E.S.; Zamyatnina, K.A.; Matveeva, N.S.; Rudenskaya, Y.A.; Kraeva, N.; Kolesnikov, A.A.; Yurchenko, V. Common Structural Patterns in the Maxicircle Divergent Region of Trypanosomatidae. Pathogens 2020, 9, 100. [Google Scholar] [CrossRef] [Green Version]
- Aphasizhev, R.; Aphasizheva, I. Mitochondrial RNA editing in trypanosomes: Small RNAs in control. Biochimie 2014, 100, 125–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westenberger, S.J.; Cerqueira, G.C.; El-Sayed, N.M.; Zingales, B.; Campbell, D.A.; Sturm, N.R. Trypanosoma cruzi mitochondrial maxicircles display species- and strain-specific variation and a conserved element in the non-coding region. BMC Genom. 2006, 7, 60. [Google Scholar] [CrossRef] [Green Version]
- Messenger, L.A.; Llewellyn, M.S.; Bhattacharyya, T.; Franzén, O.; Lewis, M.D.; Ramírez, J.D.; Carrasco, H.J.; Andersson, B.; Miles, M.A. Multiple mitochondrial introgression events and heteroplasmy in Trypanosoma cruzi revealed by maxicircle MLST and next generation sequencing. PLoS Negl. Trop. Dis. 2012, 6, e1584. [Google Scholar] [CrossRef] [Green Version]
- Gibson, W.; Bingle, L.; Blendeman, W.; Brown, J.; Wood, J.; Stevens, J. Structure and sequence variation of the trypanosome spliced leader transcript. Mol. Biochem. Parasitol. 2000, 107, 269–277. [Google Scholar] [CrossRef]
- Akopyants, N.S.; Kimblin, N.; Secundino, N.; Patrick, R.; Peters, N.; Lawyer, P.; Dobson, D.E.; Beverley, S.M.; Sacks, D.L. Demonstration of genetic exchange during cyclical development of Leishmania in the sand fly vector. Science 2009, 324, 265–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berry, A.S.F.; Salazar-Sánchez, R.; Castillo-Neyra, R.; Borrini-Mayorí, K.; Chipana-Ramos, C.; Vargas-Maquera, M.; Ancca-Juarez, J.; Náquira-Velarde, C.; Levy, M.Z.; Brisson, D.; et al. Sexual reproduction in a natural Trypanosoma cruzi population. PLoS Negl. Trop. Dis. 2019, 13, e0007392. [Google Scholar] [CrossRef] [Green Version]
- Schwabl, P.; Imamura, H.; Van den Broeck, F.; Costales, J.A.; Maiguashca-Sánchez, J.; Miles, M.A.; Andersson, B.; Grijalva, M.J.; Llewellyn, M.S. Meiotic sex in Chagas disease parasite Trypanosoma cruzi. Nat. Commun. 2019, 10, 3972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tibayrenc, M.; Ayala, F.J. Reproductive clonality of pathogens: A perspective on pathogenic viruses, bacteria, fungi, and parasitic protozoa. Proc. Natl. Acad. Sci. USA 2012, 109, E3305–E3313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tibayrenc, M.; Ayala, F.J. The population genetics of Trypanosoma cruzi revisited in the light of the predominant clonal evolution model. Acta Trop. 2015, 151, 156–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramírez, J.D.; Llewellyn, M.S. Reproductive clonality in protozoan pathogens--truth or artefact? Mol. Ecol. 2014, 23, 4195–4202. [Google Scholar] [CrossRef]
- Souza, R.T.; Lima, F.M.; Barros, R.M.; Cortez, D.R.; Santos, M.F.; Cordero, E.M.; Ruiz, J.C.; Goldenberg, S.; Teixeira, M.M.G.; da Silveira, J.F. Genome Size, Karyotype Polymorphism and Chromosomal Evolution in Trypanosoma cruzi. PLoS ONE 2011, 6, e23042. [Google Scholar] [CrossRef]
- Reis-Cunha, J.L.; Rodrigues-Luiz, G.F.; Valdivia, H.O.; Baptista, R.P.; Mendes, T.A.O.; de Morais, G.L.; Guedes, R.; Macedo, A.M.; Bern, C.; Gilman, R.H.; et al. Chromosomal copy number variation reveals differential levels of genomic plasticity in distinct Trypanosoma cruzi strains. BMC Genom. 2015, 16. [Google Scholar] [CrossRef] [Green Version]
- Lima, F.M.; Souza, R.T.; Santori, F.R.; Santos, M.F.; Cortez, D.R.; Barros, R.M.; Cano, M.I.; Valadares, H.M.S.; Macedo, A.M.; Mortara, R.A.; et al. Interclonal Variations in the Molecular Karyotype of Trypanosoma cruzi: Chromosome Rearrangements in a Single Cell-Derived Clone of the G Strain. PLoS ONE 2013, 8, e63738. [Google Scholar] [CrossRef] [Green Version]
- Henriksson, J.; Dujardin, J.C.; Barnabé, C.; Brisse, S.; Timperman, G.; Venegas, J.; Pettersson, U.; Tibayrenc, M.; Solari, A. Chromosomal size variation in Trypanosoma cruzi is mainly progressive and is evolutionarily informative. Parasitology 2002, 124, 277–286. [Google Scholar] [CrossRef] [Green Version]
- Lewis, M.D.; Llewellyn, M.S.; Gaunt, M.W.; Yeo, M.; Carrasco, H.J.; Miles, M.A. Flow cytometric analysis and microsatellite genotyping reveal extensive DNA content variation in Trypanosoma cruzi populations and expose contrasts between natural and experimental hybrids. Int. J. Parasitol. 2009, 39, 1305–1317. [Google Scholar] [CrossRef] [Green Version]
- Zingales, B.; Andrade, S.G.; Briones, M.R.S.; Campbell, D.A.; Chiari, E.; Fernandes, O.; Guhl, F.; Lages-Silva, E.; Macedo, A.M.; Machado, C.R.; et al. A new consensus for Trypanosoma cruzi intraspecific nomenclature: Second revision meeting recommends TcI to TcVI. Mem. Inst. Oswaldo Cruz 2009, 104, 1051–1054. [Google Scholar] [CrossRef]
- Brenière, S.F.; Waleckx, E.; Barnabé, C. Over Six Thousand Trypanosoma cruzi Strains Classified into Discrete Typing Units (DTUs): Attempt at an Inventory. PLoS Negl. Trop. Dis. 2016, 10, e0004792. [Google Scholar] [CrossRef] [PubMed]
- Anonymous. Recommendations from a satellite meeting. Mem. Inst. Oswaldo Cruz 1999, 94, 429–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tibayrenc, M. Genetic epidemiology of parasitic protozoa and other infectious agents: The need for an integrated approach. Int. J. Parasitol. 1998, 28, 85–104. [Google Scholar] [CrossRef]
- Brisse, S.; Barnabé, C.; Tibayrenc, M. Identification of six Trypanosoma cruzi phylogenetic lineages by random amplified polymorphic DNA and multilocus enzyme electrophoresis. Int. J. Parasitol. 2000, 30, 35–44. [Google Scholar] [CrossRef]
- Brisse, S.; Verhoef, J.; Tibayrenc, M. Characterisation of large and small subunit rRNA and mini-exon genes further supports the distinction of six Trypanosoma cruzi lineages. Int. J. Parasitol. 2001, 31, 1218–1226. [Google Scholar] [CrossRef]
- Tomazi, L.; Kawashita, S.Y.; Pereira, P.M.; Zingales, B.; Briones, M.R.S. Haplotype distribution of five nuclear genes based on network genealogies and Bayesian inference indicates that Trypanosoma cruzi hybrid strains are polyphyletic. Genet. Mol. Res. 2009, 8, 458–476. [Google Scholar] [CrossRef]
- De Freitas, J.M.; Augusto-Pinto, L.; Pimenta, J.R.; Bastos-Rodrigues, L.; Gonçalves, V.F.; Teixeira, S.M.R.; Chiari, E.; Junqueira, Â.C.V.; Fernandes, O.; Macedo, A.M.; et al. Ancestral Genomes, Sex, and the Population Structure of Trypanosoma cruzi. PLoS Pathog. 2006, 2, e24. [Google Scholar] [CrossRef] [Green Version]
- Westenberger, S.J.; Barnabé, C.; Campbell, D.A.; Sturm, N.R. Two Hybridization Events Define the Population Structure of Trypanosoma cruzi. Genetics 2005, 171, 527–543. [Google Scholar] [CrossRef] [Green Version]
- Marcili, A.; Lima, L.; Cavazzana, M.; Junqueira, A.C.V.; Veludo, H.H.; Maia Da Silva, F.; Campaner, M.; Paiva, F.; Nunes, V.L.B.; Teixeira, M.M.G. A new genotype of Trypanosoma cruzi associated with bats evidenced by phylogenetic analyses using SSU rDNA, cytochrome b and Histone H2B genes and genotyping based on ITS1 rDNA. Parasitology 2009, 136, 641–655. [Google Scholar] [CrossRef]
- Lima, L.; Espinosa-Álvarez, O.; Ortiz, P.A.; Trejo-Varón, J.A.; Carranza, J.C.; Pinto, C.M.; Serrano, M.G.; Buck, G.A.; Camargo, E.P.; Teixeira, M.M.G. Genetic diversity of Trypanosoma cruzi in bats, and multilocus phylogenetic and phylogeographical analyses supporting Tcbat as an independent DTU (discrete typing unit). Acta Trop. 2015, 151, 166–177. [Google Scholar] [CrossRef]
- Barnabé, C.; Mobarec, H.I.; Jurado, M.R.; Cortez, J.A.; Brenière, S.F. Reconsideration of the seven discrete typing units within the species Trypanosoma cruzi, a new proposal of three reliable mitochondrial clades. Infect. Genet. Evol. 2016, 39, 176–186. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed, N.M.; Myler, P.J.; Bartholomeu, D.C.; Nilsson, D.; Aggarwal, G.; Tran, A.-N.; Ghedin, E.; Worthey, E.A.; Delcher, A.L.; Blandin, G.; et al. The Genome Sequence of Trypanosoma cruzi, Etiologic Agent of Chagas Disease. Science 2005, 309, 409–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivens, A.C.; Peacock, C.S.; Worthey, E.A.; Murphy, L.; Aggarwal, G.; Berriman, M.; Sisk, E.; Rajandream, M.-A.; Adlem, E.; Aert, R.; et al. The genome of the kinetoplastid parasite, Leishmania major. Science 2005, 309, 436–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berriman, M.; Ghedin, E.; Hertz-Fowler, C.; Blandin, G.; Renauld, H.; Bartholomeu, D.C.; Lennard, N.J.; Caler, E.; Hamlin, N.E.; Haas, B.; et al. The genome of the African trypanosome Trypanosoma brucei. Science 2005, 309, 416–422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reis-Cunha, J.L.; Bartholomeu, D.C. Trypanosoma cruzi Genome Assemblies: Challenges and Milestones of Assembling a Highly Repetitive and Complex Genome. Methods Mol. Biol. 2019, 1955, 1–22. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed, N.M.; Myler, P.J.; Blandin, G.; Berriman, M.; Crabtree, J.; Aggarwal, G.; Caler, E.; Renauld, H.; Worthey, E.A.; Hertz-Fowler, C.; et al. Comparative genomics of trypanosomatid parasitic protozoa. Science 2005, 309, 404–409. [Google Scholar] [CrossRef] [Green Version]
- Callejas-Hernández, F.; Rastrojo, A.; Poveda, C.; Gironès, N.; Fresno, M. Genomic assemblies of newly sequenced Trypanosoma cruzi strains reveal new genomic expansion and greater complexity. Sci. Rep. 2018, 8, 14631. [Google Scholar] [CrossRef]
- Baptista, R.P.; Reis-Cunha, J.L.; DeBarry, J.D.; Chiari, E.; Kissinger, J.C.; Bartholomeu, D.C.; Macedo, A.M. Assembly of highly repetitive genomes using short reads: The genome of discrete typing unit III Trypanosoma cruzi strain 231. Microb. Genom. 2018, 4. [Google Scholar] [CrossRef] [Green Version]
- Franzén, O.; Ochaya, S.; Sherwood, E.; Lewis, M.D.; Llewellyn, M.S.; Miles, M.A.; Andersson, B. Shotgun Sequencing Analysis of Trypanosoma cruzi I Sylvio X10/1 and Comparison with T. cruzi VI CL Brener. PLoS Negl. Trop. Dis. 2011, 5, e984. [Google Scholar] [CrossRef] [Green Version]
- Bradwell, K.R.; Koparde, V.N.; Matveyev, A.V.; Serrano, M.G.; Alves, J.M.P.; Parikh, H.; Huang, B.; Lee, V.; Espinosa-Alvarez, O.; Ortiz, P.A.; et al. Genomic comparison of Trypanosoma conorhini and Trypanosoma rangeli to Trypanosoma cruzi strains of high and low virulence. BMC Genom. 2018, 19. [Google Scholar] [CrossRef]
- Franzén, O.; Talavera-López, C.; Ochaya, S.; Butler, C.E.; Messenger, L.A.; Lewis, M.D.; Llewellyn, M.S.; Marinkelle, C.J.; Tyler, K.M.; Miles, M.A.; et al. Comparative genomic analysis of human infective Trypanosoma cruzi lineages with the bat-restricted subspecies T. cruzi marinkellei. BMC Genom. 2012, 13, 531. [Google Scholar] [CrossRef] [PubMed]
- Camacho, E.; González-de la Fuente, S.; Rastrojo, A.; Peiró-Pastor, R.; Solana, J.C.; Tabera, L.; Gamarro, F.; Carrasco-Ramiro, F.; Requena, J.M.; Aguado, B. Complete assembly of the Leishmania donovani (HU3 strain) genome and transcriptome annotation. Sci. Rep. 2019, 9, 6127. [Google Scholar] [CrossRef] [PubMed]
- Callejas-Hernández, F.; Gironès, N.; Fresno, M. Genome Sequence of Trypanosoma cruzi Strain Bug2148. Genome Announc. 2018, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Díaz-Viraqué, F.; Pita, S.; Greif, G.; de Souza, R.C.M.; Iraola, G.; Robello, C. Nanopore Sequencing Significantly Improves Genome Assembly of the Protozoan Parasite Trypanosoma cruzi. Genome Biol. Evol. 2019, 11, 1952–1957. [Google Scholar] [CrossRef] [PubMed]
- Berná, L.; Rodriguez, M.; Chiribao, M.L.; Parodi-Talice, A.; Pita, S.; Rijo, G.; Alvarez-Valin, F.; Robello, C. Expanding an expanded genome: Long-read sequencing of Trypanosoma cruzi. Microb. Genom. 2018, 4. [Google Scholar] [CrossRef]
- Arner, E.; Kindlund, E.; Nilsson, D.; Farzana, F.; Ferella, M.; Tammi, M.T.; Andersson, B. Database of Trypanosoma cruzi repeated genes: 20,000 additional gene variants. BMC Genom. 2007, 8, 391. [Google Scholar] [CrossRef] [Green Version]
- Callejas-Hernández, F.; Gutierrez-Nogues, Á.; Rastrojo, A.; Gironès, N.; Fresno, M. Analysis of mRNA processing at whole transcriptome level, transcriptomic profile and genome sequence refinement of Trypanosoma cruzi. Sci. Rep. 2019, 9. [Google Scholar] [CrossRef]
- Weatherly, D.B.; Boehlke, C.; Tarleton, R.L. Chromosome level assembly of the hybrid Trypanosoma cruzi genome. BMC Genom. 2009, 10, 255. [Google Scholar] [CrossRef] [Green Version]
- Berná, L.; Pita, S.; Chiribao, M.L.; Parodi-Talice, A.; Alvarez-Valin, F.; Robello, C. Biology of the Trypanosoma cruzi Genome. In Biology of Trypanosoma cruzi; IntechOpen: London, UK, 2019. [Google Scholar] [CrossRef] [Green Version]
- Downing, T.; Imamura, H.; Decuypere, S.; Clark, T.G.; Coombs, G.H.; Cotton, J.A.; Hilley, J.D.; de Doncker, S.; Maes, I.; Mottram, J.C.; et al. Whole genome sequencing of multiple Leishmania donovani clinical isolates provides insights into population structure and mechanisms of drug resistance. Genome Res. 2011, 21, 2143–2156. [Google Scholar] [CrossRef] [Green Version]
- Mannaert, A.; Downing, T.; Imamura, H.; Dujardin, J.-C. Adaptive mechanisms in pathogens: Universal aneuploidy in Leishmania. Trends Parasitol. 2012, 28, 370–376. [Google Scholar] [CrossRef] [Green Version]
- Dujardin, J.-C.; Mannaert, A.; Durrant, C.; Cotton, J.A. Mosaic aneuploidy in Leishmania: The perspective of whole genome sequencing. Trends Parasitol. 2014, 30, 554–555. [Google Scholar] [CrossRef]
- Almeida, L.V.; Coqueiro-Dos-Santos, A.; Rodriguez-Luiz, G.F.; McCulloch, R.; Bartholomeu, D.C.; Reis-Cunha, J.L. Chromosomal copy number variation analysis by next generation sequencing confirms ploidy stability in Trypanosoma brucei subspecies. Microb. Genom. 2018, 4. [Google Scholar] [CrossRef] [PubMed]
- Reis-Cunha, J.L.; Baptista, R.P.; Rodrigues-Luiz, G.F.; Coqueiro-Dos-Santos, A.; Valdivia, H.O.; de Almeida, L.V.; Cardoso, M.S.; D’Ávila, D.A.; Dias, F.H.C.; Fujiwara, R.T.; et al. Whole genome sequencing of Trypanosoma cruzi field isolates reveals extensive genomic variability and complex aneuploidy patterns within TcII DTU. BMC Genom. 2018, 19, 816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reis-Cunha, J.L.; Valdivia, H.O.; Bartholomeu, D.C. Gene and Chromosomal Copy Number Variations as an Adaptive Mechanism Towards a Parasitic Lifestyle in Trypanosomatids. Curr. Genom. 2018, 19, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Araújo, P.R.; Teixeira, S.M. Regulatory elements involved in the post-transcriptional control of stage-specific gene expression in Trypanosoma cruzi: A review. Memórias Do Inst. Oswaldo Cruz 2011, 106, 257–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pastro, L.; Smircich, P.; Di Paolo, A.; Becco, L.; Duhagon, M.A.; Sotelo-Silveira, J.; Garat, B. Nuclear Compartmentalization Contributes to Stage-Specific Gene Expression Control in Trypanosoma cruzi. Front. Cell Dev. Biol. 2017, 5. [Google Scholar] [CrossRef] [Green Version]
- Vargas, N.; Pedroso, A.; Zingales, B. Chromosomal polymorphism, gene synteny and genome size in T. cruzi I and T. cruzi II groups. Mol. Biochem. Parasitol. 2004, 138, 131–141. [Google Scholar] [CrossRef]
- Herreros-Cabello, A.; Callejas-Hernández, F.; Fresno, M.; Gironès, N. Comparative proteomic analysis of trypomastigotes from Trypanosoma cruzi strains with different pathogenicity. Infect. Genet. Evol. 2019, 76, 104041. [Google Scholar] [CrossRef]
- Avila, C.C.; Mule, S.N.; Rosa-Fernandes, L.; Viner, R.; Barisón, M.J.; Costa-Martins, A.G.; de Oliveira, G.S.; Teixeira, M.M.G.; Marinho, C.R.F.; Silber, A.M.; et al. Proteome-Wide Analysis of Trypanosoma cruzi Exponential and Stationary Growth Phases Reveals a Subcellular Compartment-Specific Regulation. Genes 2018, 9, 413. [Google Scholar] [CrossRef] [Green Version]
- Atwood, J.A.; Weatherly, D.B.; Minning, T.A.; Bundy, B.; Cavola, C.; Opperdoes, F.R.; Orlando, R.; Tarleton, R.L. The Trypanosoma cruzi Proteome. Science 2005, 309, 473–476. [Google Scholar] [CrossRef]
- Godoy, L.M.F.; de Marchini, F.K.; Pavoni, D.P.; Rampazzo, R.C.P.; Probst, C.M.; Goldenberg, S.; Krieger, M.A. Quantitative proteomics of Trypanosoma cruzi during metacyclogenesis. Proteomics 2012, 12, 2694–2703. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, J.L. An Evolutionary View of Trypanosoma cruzi Telomeres. Front. Cell. Infect. Microbiol. 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Moraes Barros, R.R.; Marini, M.M.; Antônio, C.R.; Cortez, D.R.; Miyake, A.M.; Lima, F.M.; Ruiz, J.C.; Bartholomeu, D.C.; Chiurillo, M.A.; Ramirez, J.L.; et al. Anatomy and evolution of telomeric and subtelomeric regions in the human protozoan parasite Trypanosoma cruzi. BMC Genom. 2012, 13, 229. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Chiurillo, M.A.; El-Sayed, N.; Jones, K.; Santos, M.R.M.; Porcile, P.E.; Andersson, B.; Myler, P.; da Silveira, J.F.; Ramírez, J.L. Telomere and subtelomere of Trypanosoma cruzi chromosomes are enriched in (pseudo)genes of retrotransposon hot spot and trans-sialidase-like gene families: The origins of T. cruzi telomeres. Gene 2005, 346, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Lira, C.B.B.; Giardini, M.A.; Neto, J.L.S.; Conte, F.F.; Cano, M.I.N. Telomere biology of trypanosomatids: Beginning to answer some questions. Trends Parasitol. 2007, 23, 357–362. [Google Scholar] [CrossRef] [PubMed]
- Freitas-Junior, L.H.; Bottius, E.; Pirrit, L.A.; Deitsch, K.W.; Scheidig, C.; Guinet, F.; Nehrbass, U.; Wellems, T.E.; Scherf, A. Frequent ectopic recombination of virulence factor genes in telomeric chromosome clusters of P. falciparum. Nature 2000, 407, 1018–1022. [Google Scholar] [CrossRef] [PubMed]
- Kudla, G.; Helwak, A.; Lipinski, L. Gene Conversion and GC-Content Evolution in Mammalian Hsp70. Mol. Biol. Evol. 2004, 21, 1438–1444. [Google Scholar] [CrossRef]
- Galtier, N. Gene conversion drives GC content evolution in mammalian histones. Trends Genet. 2003, 19, 65–68. [Google Scholar] [CrossRef]
- Chiurillo, M.A.; Cano, I.; Da Silveira, J.F.; Ramirez, J.L. Organization of telomeric and sub-telomeric regions of chromosomes from the protozoan parasite Trypanosoma cruzi. Mol. Biochem. Parasitol. 1999, 100, 173–183. [Google Scholar] [CrossRef]
- Kudla, G.; Lipinski, L.; Caffin, F.; Helwak, A.; Zylicz, M. High Guanine and Cytosine Content Increases mRNA Levels in Mammalian Cells. PLoS Biol. 2006, 4, e180. [Google Scholar] [CrossRef]
- Tiengwe, C.; Marcello, L.; Farr, H.; Dickens, N.; Kelly, S.; Swiderski, M.; Vaughan, D.; Gull, K.; Barry, J.D.; Bell, S.D.; et al. Genome-wide Analysis Reveals Extensive Functional Interaction between DNA Replication Initiation and Transcription in the Genome of Trypanosoma brucei. Cell Rep. 2012, 2, 185–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lombraña, R.; Álvarez, A.; Fernández-Justel, J.M.; Almeida, R.; Poza-Carrión, C.; Gomes, F.; Calzada, A.; Requena, J.M.; Gómez, M. Transcriptionally Driven DNA Replication Program of the Human Parasite Leishmania major. Cell Rep. 2016, 16, 1774–1786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marques, C.A.; Dickens, N.J.; Paape, D.; Campbell, S.J.; McCulloch, R. Genome-wide mapping reveals single-origin chromosome replication in Leishmania, a eukaryotic microbe. Genome Biol. 2015, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Da Silva, M.S.; Pavani, R.S.; Damasceno, J.D.; Marques, C.A.; McCulloch, R.; Tosi, L.R.O.; Elias, M.C. Nuclear DNA Replication in Trypanosomatids: There Are No Easy Methods for Solving Difficult Problems. Trends Parasitol. 2017, 33, 858–874. [Google Scholar] [CrossRef] [Green Version]
- De Araujo, C.B.; da Cunha, J.P.C.; Inada, D.T.; Damasceno, J.; Lima, A.R.J.; Hiraiwa, P.; Marques, C.; Gonçalves, E.; Nishiyama-Junior, M.Y.; McCulloch, R.; et al. Replication origin location might contribute to genetic variability in Trypanosoma cruzi. BMC Genom. 2020, 21, 414. [Google Scholar] [CrossRef]
- De Araujo, C.B.; Calderano, S.G.; Elias, M.C. The Dynamics of Replication in Trypanosoma cruzi Parasites by Single-Molecule Analysis. J. Eukaryot. Microbiol. 2019, 66, 514–518. [Google Scholar] [CrossRef]
- De Gaudenzi, J.G.; Noé, G.; Campo, V.A.; Frasch, A.C.; Cassola, A. Gene expression regulation in trypanosomatids. Essays Biochem. 2011, 51, 31–46. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Calvillo, S.; Nguyen, D.; Stuart, K.; Myler, P.J. Transcription Initiation and Termination on Leishmania major Chromosome 3. Eukaryot. Cell. 2004, 3, 506–517. [Google Scholar] [CrossRef] [Green Version]
- Smircich, P.; El-Sayed, N.M.; Garat, B. Intrinsic DNA curvature in trypanosomes. BMC Res. Notes 2017, 10, 585. [Google Scholar] [CrossRef]
- Gilinger, G.; Bellofatto, V. Trypanosome spliced leader RNA genes contain the first identified RNA polymerase II gene promoter in these organisms. Nucleic Acids Res. 2001, 29, 1556–1564. [Google Scholar] [CrossRef] [Green Version]
- Siegel, T.N.; Hekstra, D.R.; Kemp, L.E.; Figueiredo, L.M.; Lowell, J.E.; Fenyo, D.; Wang, X.; Dewell, S.; Cross, G.A.M. Four histone variants mark the boundaries of polycistronic transcription units in Trypanosoma brucei. Genes Dev. 2009, 23, 1063–1076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Respuela, P.; Ferella, M.; Rada-Iglesias, A.; Aslund, L. Histone acetylation and methylation at sites initiating divergent polycistronic transcription in Trypanosoma cruzi. J. Biol. Chem. 2008, 283, 15884–15892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Padilla-Mejía, N.E.; Florencio-Martínez, L.E.; Figueroa-Angulo, E.E.; Manning-Cela, R.G.; Hernández-Rivas, R.; Myler, P.J.; Martínez-Calvillo, S. Gene organization and sequence analyses of transfer RNA genes in Trypanosomatid parasites. BMC Genom. 2009, 10, 232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Günzl, A. The Pre-mRNA Splicing Machinery of Trypanosomes: Complex or Simplified? Eukaryot Cell 2010, 9, 1159–1170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palenchar, J.B.; Bellofatto, V. Gene transcription in trypanosomes. Mol. Biochem. Parasitol. 2006, 146, 135–141. [Google Scholar] [CrossRef]
- Rastrojo, A.; Carrasco-Ramiro, F.; Martín, D.; Crespillo, A.; Reguera, R.M.; Aguado, B.; Requena, J.M. The transcriptome of Leishmania major in the axenic promastigote stage: Transcript annotation and relative expression levels by RNA-seq. BMC Genom. 2013, 14, 223. [Google Scholar] [CrossRef] [Green Version]
- Kolev, N.G.; Franklin, J.B.; Carmi, S.; Shi, H.; Michaeli, S.; Tschudi, C. The Transcriptome of the Human Pathogen Trypanosoma brucei at Single-Nucleotide Resolution. PLoS Pathog. 2010, 6, e1001090. [Google Scholar] [CrossRef]
- Thomas, S.; Green, A.; Sturm, N.R.; Campbell, D.A.; Myler, P.J. Histone acetylations mark origins of polycistronic transcription in Leishmania major. BMC Genom. 2009, 10, 152. [Google Scholar] [CrossRef] [Green Version]
- Minning, T.A.; Weatherly, D.B.; Atwood, J.; Orlando, R.; Tarleton, R.L. The steady-state transcriptome of the four major life-cycle stages of Trypanosoma cruzi. BMC Genom. 2009, 10, 370. [Google Scholar] [CrossRef] [Green Version]
- Minning, T.A.; Weatherly, D.B.; Flibotte, S.; Tarleton, R.L. Widespread, focal copy number variations (CNV) and whole chromosome aneuploidies in Trypanosoma cruzi strains revealed by array comparative genomic hybridization. BMC Genom. 2011, 12, 139. [Google Scholar] [CrossRef] [Green Version]
- Bartholomeu, D.C.; de Paiva, R.M.C.; Mendes, T.A.O.; DaRocha, W.D.; Teixeira, S.M.R. Unveiling the Intracellular Survival Gene Kit of Trypanosomatid Parasites. PLoS Pathog. 2014, 10, e1004399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorch, Y.; Maier-Davis, B.; Kornberg, R.D. Role of DNA sequence in chromatin remodeling and the formation of nucleosome-free regions. Genes Dev. 2014, 28, 2492–2497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patino, L.H.; Ramírez, J.D. RNA-seq in kinetoplastids: A powerful tool for the understanding of the biology and host-pathogen interactions. Infect. Genet. Evol. 2017, 49, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Clayton, C. The Regulation of Trypanosome Gene Expression by RNA-Binding Proteins. PLoS Pathog. 2013, 9, e1003680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelly, S.; Kramer, S.; Schwede, A.; Maini, P.K.; Gull, K.; Carrington, M. Genome organization is a major component of gene expression control in response to stress and during the cell division cycle in trypanosomes. Open Biol. 2012, 2. [Google Scholar] [CrossRef] [Green Version]
- Pech-Canul, Á.D.L.C.; Monteón, V.; Solís-Oviedo, R.-L. A Brief View of the Surface Membrane Proteins from Trypanosoma cruzi. J. Parasitol. Res. 2017, 2017. [Google Scholar] [CrossRef] [Green Version]
- Poveda, C.; Herreros-Cabello, A.; Callejas-Hernández, F.; Osuna-Pérez, J.; Maza, M.C.; Chillón-Marinas, C.; Calderón, J.; Stamatakis, K.; Fresno, M.; Gironès, N. Interaction of Signaling Lymphocytic Activation Molecule Family 1 (SLAMF1) receptor with Trypanosoma cruzi is strain-dependent and affects NADPH oxidase expression and activity. PLoS Negl. Trop. Dis. 2020, 14, e0008608. [Google Scholar] [CrossRef]
- Medina-Acosta, E.; Franco, A.M.R.; Jansen, A.M.; Sampol, M.; Nevés, N.; Pontes-De-Carvalho, L.; Grimaldi, G.; Nussenzweig, V. Trans-sialidase and Sialidase Activities Discriminate between Morphologically Indistinguishable Trypanosomatids. Eur. J. Biochem. 1994, 225, 333–339. [Google Scholar] [CrossRef]
- Freitas, L.M.; dos Santos, S.L.; Rodrigues-Luiz, G.F.; Mendes, T.A.O.; Rodrigues, T.S.; Gazzinelli, R.T.; Teixeira, S.M.R.; Fujiwara, R.T.; Bartholomeu, D.C. Genomic Analyses, Gene Expression and Antigenic Profile of the Trans-Sialidase Superfamily of Trypanosoma cruzi Reveal an Undetected Level of Complexity. PLoS ONE 2011, 6, e25914. [Google Scholar] [CrossRef] [Green Version]
- De Pablos, L.M.; Osuna, A. Multigene Families in Trypanosoma cruzi and Their Role in Infectivity. Infect. Immun. 2012, 80, 2258–2264. [Google Scholar] [CrossRef] [Green Version]
- Frasch, A.C.C. Functional Diversity in the Trans-sialidase and Mucin Families in Trypanosoma cruzi. Parasitol. Today 2000, 16, 282–286. [Google Scholar] [CrossRef]
- Chiurillo, M.A.; Cortez, D.R.; Lima, F.M.; Cortez, C.; Ramírez, J.L.; Martins, A.G.; Serrano, M.G.; Teixeira, M.M.G.; da Silveira, J.F. The diversity and expansion of the trans-sialidase gene family is a common feature in Trypanosoma cruzi clade members. Infect. Genet. Evol. 2016, 37, 266–274. [Google Scholar] [CrossRef] [PubMed]
- Frevert, U.; Schenkman, S.; Nussenzweig, V. Stage-specific expression and intracellular shedding of the cell surface trans-sialidase of Trypanosoma cruzi. Infect. Immun. 1992, 60, 2349–2360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lantos, A.B.; Carlevaro, G.; Araoz, B.; Ruiz Diaz, P.; Camara, M.d.L.M.; Buscaglia, C.A.; Bossi, M.; Yu, H.; Chen, X.; Bertozzi, C.R.; et al. Sialic Acid Glycobiology Unveils Trypanosoma cruzi Trypomastigote Membrane Physiology. PLoS Pathog. 2016, 12, e1005559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schenkman, S.; Eichinger, D.; Pereira, M.E.A.; Nussenzweig, V. Structural and Functional Properties of Trypanosoma Trans-Sialidase. Annu. Rev. Microbiol. 1994, 48, 499–523. [Google Scholar] [CrossRef]
- Oliveira, I.A.; Freire-de-Lima, L.; Penha, L.L.; Dias, W.B.; Todeschini, A.R. Trypanosoma cruzi Trans-Sialidase: Structural Features and Biological Implications. In Proteins and Proteomics of Leishmania and Trypanosoma; Santos, A.L.S., Branquinha, M.H., d’Avila-Levy, C.M., Kneipp, L.F., Sodré, C.L., Eds.; Subcellular Biochemistry; Springer: Dordrecht, The Netherlands, 2014; pp. 181–201. ISBN 978-94-007-7305-9. [Google Scholar]
- Pereira, M.E.; Loures, M.A.; Villalta, F.; Andrade, A.F. Lectin receptors as markers for Trypanosoma cruzi. Developmental stages and a study of the interaction of wheat germ agglutinin with sialic acid residues on epimastigote cells. J. Exp. Med. 1980, 152, 1375–1392. [Google Scholar] [CrossRef] [Green Version]
- Parodi, A.J.; Pollevick, G.D.; Mautner, M.; Buschiazzo, A.; Sanchez, D.O.; Frasch, A.C. Identification of the gene(s) coding for the trans-sialidase of Trypanosoma cruzi. EMBO J. 1992, 11, 1705–1710. [Google Scholar] [CrossRef]
- Previato, J.; Andrade, A.F.B.; Pessolani, M.C.V.; Mendonça-Previato, L. Incorporation of sialic acid into Trypanosoma cruzi macromolecules. A proposal for a new metabolic route. Mol. Biochem. Parasitol. 1985, 16, 85–96. [Google Scholar] [CrossRef]
- Schauer, R.; Reuter, G.; Mühlpfordt, H.; Andrade, A.F.; Pereira, M.E. The occurrence of N-acetyl- and N-glycoloylneuraminic acid in Trypanosoma cruzi. Hoppe-Seyler’s Z Physiol. Chem. 1983, 364, 1053–1057. [Google Scholar] [CrossRef]
- Schenkman, S.; Mortara, R.A. HeLa cells extend and internalize pseudopodia during active invasion by Trypanosoma cruzi trypomastigotes. J. Cell. Sci. 1992, 101, 895–905. [Google Scholar]
- Scudder, P.; Doom, J.P.; Chuenkova, M.; Manger, I.D.; Pereira, M.E. Enzymatic characterization of β-D-galactoside α 2,3-trans-sialidase from Trypanosoma cruzi. J. Biol. Chem. 1993, 268, 9886–9891. [Google Scholar] [PubMed]
- Zingales, B.; Carniol, C.; de Lederkremer, R.M.; Colli, W. Direct sialic acid transfer from a protein donor to glycolipids of trypomastigote forms of Trypanosoma cruzi. Mol. Biochem. Parasitol. 1987, 26, 135–144. [Google Scholar] [CrossRef]
- Pereira-Chioccola, V.L.; Acosta-Serrano, A.; Correia de Almeida, I.; Ferguson, M.A.; Souto-Padron, T.; Rodrigues, M.M.; Travassos, L.R.; Schenkman, S. Mucin-like molecules form a negatively charged coat that protects Trypanosoma cruzi trypomastigotes from killing by human anti-α-galactosyl antibodies. J. Cell. Sci. 2000, 113, 1299–1307. [Google Scholar] [PubMed]
- Pereira, M.E. A developmentally regulated neuraminidase activity in Trypanosoma cruzi. Science 1983, 219, 1444–1446. [Google Scholar] [CrossRef]
- Nardy, A.F.F.R.; Freire-de-Lima, C.G.; Pérez, A.R.; Morrot, A. Role of Trypanosoma cruzi Trans-sialidase on the Escape from Host Immune Surveillance. Front. Microbiol. 2016, 7, 348. [Google Scholar] [CrossRef] [Green Version]
- Colli, W. Trans-sialidase: A unique enzyme activity discovered in the protozoan Trypanosoma cruzi. FASEB J. 1993, 7, 1257–1264. [Google Scholar] [CrossRef] [PubMed]
- Cross, G.A.; Takle, G.B. The surface trans-sialidase family of Trypanosoma cruzi. Annu. Rev. Microbiol. 1993, 47, 385–411. [Google Scholar] [CrossRef] [PubMed]
- Schenkman, S.; Jiang, M.S.; Hart, G.W.; Nussenzweig, V. A novel cell surface trans-sialidase of Trypanosoma cruzi generates a stage-specific epitope required for invasion of mammalian cells. Cell 1991, 65, 1117–1125. [Google Scholar] [CrossRef]
- Buschiazzo, A.; Campetella, O.; Frasch, A.C.C. Trypanosoma rangeli sialidase: Cloning, expression and similarity to T. cruzi trans-sialidase. Glycobiology 1997, 7, 1167–1173. [Google Scholar] [CrossRef] [Green Version]
- Amaya, M.F.; Buschiazzo, A.; Nguyen, T.; Alzari, P.M. The high resolution structures of free and inhibitor-bound Trypanosoma rangeli sialidase and its comparison with T. cruzi trans-sialidase. J. Mol. Biol. 2003, 325, 773–784. [Google Scholar] [CrossRef]
- Schenkman, S.; Pontes de Carvalho, L.; Nussenzweig, V. Trypanosoma cruzi trans-sialidase and neuraminidase activities can be mediated by the same enzymes. J. Exp. Med. 1992, 175, 567–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pollevick, G.D.; Affranchino, J.L.; Frasch, A.C.; Sánchez, D.O. The complete sequence of a shed acute-phase antigen of Trypanosoma cruzi. Mol. Biochem. Parasitol. 1991, 47, 247–250. [Google Scholar] [CrossRef]
- Briones, M.R.; Egima, C.M.; Schenkman, S. Trypanosoma cruzi trans-sialidase gene lacking C-terminal repeats and expressed in epimastigote forms. Mol. Biochem. Parasitol. 1995, 70, 9–17. [Google Scholar] [CrossRef]
- Añez-Rojas, N.; Peralta, A.; Crisante, G.; Rojas, A.; Añez, N.; Ramírez, J.L.; Chiurillo, M.A. Trypanosoma rangeli expresses a gene of the group II trans-sialidase superfamily. Mol. Biochem. Parasitol. 2005, 142, 133–136. [Google Scholar] [CrossRef] [Green Version]
- Mattos, E.C.; Tonelli, R.R.; Colli, W.; Alves, M.J.M. The Gp85 surface glycoproteins from Trypanosoma cruzi. Subcell. Biochem. 2014, 74, 151–180. [Google Scholar] [CrossRef]
- Colli, W.; Alves, M.J. Relevant glycoconjugates on the surface of Trypanosoma cruzi. Mem. Inst. Oswaldo Cruz 1999, 94, 37–49. [Google Scholar] [CrossRef] [Green Version]
- Claser, C.; Espíndola, N.M.; Sasso, G.; Vaz, A.J.; Boscardin, S.B.; Rodrigues, M.M. Immunologically relevant strain polymorphism in the Amastigote Surface Protein 2 of Trypanosoma cruzi. Microbes Infect. 2007, 9, 1011–1019. [Google Scholar] [CrossRef]
- Giordano, R.; Fouts, D.L.; Tewari, D.; Colli, W.; Manning, J.E.; Alves, M.J. Cloning of a surface membrane glycoprotein specific for the infective form of Trypanosoma cruzi having adhesive properties to laminin. J. Biol. Chem. 1999, 274, 3461–3468. [Google Scholar] [CrossRef] [Green Version]
- Magdesian, M.H.; Giordano, R.; Ulrich, H.; Juliano, M.A.; Juliano, L.; Schumacher, R.I.; Colli, W.; Alves, M.J. Infection by Trypanosoma cruzi. Identification of a parasite ligand and its host cell receptor. J. Biol. Chem. 2001, 276, 19382–19389. [Google Scholar] [CrossRef] [Green Version]
- Tonelli, R.R.; Giordano, R.J.; Barbu, E.M.; Torrecilhas, A.C.; Kobayashi, G.S.; Langley, R.R.; Arap, W.; Pasqualini, R.; Colli, W.; Alves, M.J.M. Role of the gp85/trans-sialidases in Trypanosoma cruzi tissue tropism: Preferential binding of a conserved peptide motif to the vasculature in vivo. PLoS Negl. Trop. Dis. 2010, 4, e864. [Google Scholar] [CrossRef] [Green Version]
- Santos, M.A.; Garg, N.; Tarleton, R.L. The identification and molecular characterization of Trypanosoma cruzi amastigote surface protein-1, a member of the trans-sialidase gene super-family. Mol. Biochem. Parasitol. 1997, 86, 1–11. [Google Scholar] [PubMed]
- Wizel, B.; Nunes, M.; Tarleton, R.L. Identification of Trypanosoma cruzi trans-sialidase family members as targets of protective CD8+ TC1 responses. J. Immunol. 1997, 159, 6120–6130. [Google Scholar] [PubMed]
- Wizel, B.; Palmieri, M.; Mendoza, C.; Arana, B.; Sidney, J.; Sette, A.; Tarleton, R. Human infection with Trypanosoma cruzi induces parasite antigen-specific cytotoxic T lymphocyte responses. J. Clin. Investig. 1998, 102, 1062–1071. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, N. Molecular basis of mammalian cell invasion by Trypanosoma cruzi. Acad. Bras. Cienc. 2006, 78, 87–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Favoreto, S.; Dorta, M.L.; Yoshida, N. Trypanosoma cruzi 175-kDa protein tyrosine phosphorylation is associated with host cell invasion. Exp. Parasitol. 1998, 89, 188–194. [Google Scholar] [CrossRef]
- Nogueira, N. Host and parasite factors affecting the invasion of mononuclear phagocytes by Trypanosoma cruzi. Ciba Found. Symp. 1983, 99, 52–73. [Google Scholar] [CrossRef]
- Matsumoto, T.K.; Cotrim, P.C.; da Silveira, J.F.; Stolf, A.M.S.; Umezawa, E.S. Trypanosoma cruzi: Isolation of an immunodominant peptide of TESA (Trypomastigote Excreted-Secreted Antigens) by gene cloning. Diagn. Microbiol. Infect. Dis. 2002, 42, 187–192. [Google Scholar] [CrossRef]
- Beucher, M.; Norris, K.A. Sequence diversity of the Trypanosoma cruzi complement regulatory protein family. Infect. Immun. 2008, 76, 750–758. [Google Scholar] [CrossRef] [Green Version]
- Kipnis, T.L.; David, J.R.; Alper, C.A.; Sher, A.; da Silva, W.D. Enzymatic treatment transforms trypomastigotes of Trypanosoma cruzi into activators of alternative complement pathway and potentiates their uptake by macrophages. Proc. Natl. Acad. Sci. USA 1981, 78, 602–605. [Google Scholar] [CrossRef] [Green Version]
- Grisard, E.C.; Stoco, P.H.; Wagner, G.; Sincero, T.C.M.; Rotava, G.; Rodrigues, J.B.; Snoeijer, C.Q.; Koerich, L.B.; Sperandio, M.M.; Bayer-Santos, E.; et al. Transcriptomic analyses of the avirulent protozoan parasite Trypanosoma rangeli. Mol. Biochem. Parasitol. 2010, 174, 18–25. [Google Scholar] [CrossRef]
- Stoco, P.H.; Wagner, G.; Talavera-Lopez, C.; Gerber, A.; Zaha, A.; Thompson, C.E.; Bartholomeu, D.C.; Lückemeyer, D.D.; Bahia, D.; Loreto, E.; et al. Genome of the avirulent human-infective trypanosome—Trypanosoma rangeli. PLoS Negl. Trop. Dis. 2014, 8, e3176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tonelli, R.R.; Torrecilhas, A.C.; Jacysyn, J.F.; Juliano, M.A.; Colli, W.; Alves, M.J.M. In vivo infection by Trypanosoma cruzi: The conserved FLY domain of the gp85/trans-sialidase family potentiates host infection. Parasitology 2011, 138, 481–492. [Google Scholar] [CrossRef] [PubMed]
- Roggentin, P.; Rothe, B.; Kaper, J.B.; Galen, J.; Lawrisuk, L.; Vimr, E.R.; Schauer, R. Conserved sequences in bacterial and viral sialidases. Glycoconj. J. 1989, 6, 349–353. [Google Scholar] [CrossRef] [PubMed]
- Gaskell, A.; Crennell, S.; Taylor, G. The three domains of a bacterial sialidase: A β-propeller, an immunoglobulin module and a galactose-binding jelly-roll. Structure 1995, 3, 1197–1205. [Google Scholar] [CrossRef] [Green Version]
- Cremona, M.L.; Campetella, O.; Sánchez, D.O.; Frasch, A.C. Enzymically inactive members of the trans-sialidase family from Trypanosoma cruzi display β-galactose binding activity. Glycobiology 1999, 9, 581–587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Todeschini, A.R.; Dias, W.B.; Girard, M.F.; Wieruszeski, J.-M.; Mendonça-Previato, L.; Previato, J.O. Enzymatically inactive trans-sialidase from Trypanosoma cruzi binds sialyl and β-galactopyranosyl residues in a sequential ordered mechanism. J. Biol. Chem. 2004, 279, 5323–5328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acosta-Serrano, A.; Almeida, I.C.; Freitas-Junior, L.H.; Yoshida, N.; Schenkman, S. The mucin-like glycoprotein super-family of Trypanosoma cruzi: Structure and biological roles. Mol. Biochem. Parasitol. 2001, 114, 143–150. [Google Scholar] [CrossRef]
- Buscaglia, C.A.; Campo, V.A.; Frasch, A.C.C.; Di Noia, J.M. Trypanosoma cruzi surface mucins: Host-dependent coat diversity. Nat. Rev. Microbiol. 2006, 4, 229–236. [Google Scholar] [CrossRef]
- Schenkman, S.; Ferguson, M.A.; Heise, N.; de Almeida, M.L.; Mortara, R.A.; Yoshida, N. Mucin-like glycoproteins linked to the membrane by glycosylphosphatidylinositol anchor are the major acceptors of sialic acid in a reaction catalyzed by trans-sialidase in metacyclic forms of Trypanosoma cruzi. Mol. Biochem. Parasitol. 1993, 59, 293–303. [Google Scholar] [CrossRef]
- Mucci, J.; Lantos, A.B.; Buscaglia, C.A.; Leguizamón, M.S.; Campetella, O. The Trypanosoma cruzi Surface, a Nanoscale Patchwork Quilt. Trends Parasitol. 2017, 33, 102–112. [Google Scholar] [CrossRef] [Green Version]
- Campo, V.; Di Noia, J.M.; Buscaglia, C.A.; Agüero, F.; Sánchez, D.O.; Frasch, A.C.C. Differential accumulation of mutations localized in particular domains of the mucin genes expressed in the vertebrate host stage of Trypanosoma cruzi. Mol. Biochem. Parasitol. 2004, 133, 81–91. [Google Scholar] [CrossRef] [PubMed]
- Buscaglia, C.A.; Campo, V.A.; Di Noia, J.M.; Torrecilhas, A.C.T.; De Marchi, C.R.; Ferguson, M.A.J.; Frasch, A.C.C.; Almeida, I.C. The surface coat of the mammal-dwelling infective trypomastigote stage of Trypanosoma cruzi is formed by highly diverse immunogenic mucins. J. Biol. Chem. 2004, 279, 15860–15869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urban, I.; Boiani Santurio, L.; Chidichimo, A.; Yu, H.; Chen, X.; Mucci, J.; Agüero, F.; Buscaglia, C.A. Molecular diversity of the Trypanosoma cruzi TcSMUG family of mucin genes and proteins. Biochem. J. 2011, 438, 303–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Noia, J.M.; D’Orso, I.; Sánchez, D.O.; Frasch, A.C. AU-rich elements in the 3′-untranslated region of a new mucin-type gene family of Trypanosoma cruzi confers mRNA instability and modulates translation efficiency. J. Biol. Chem. 2000, 275, 10218–10227. [Google Scholar] [CrossRef] [Green Version]
- Cámara, M.d.l.M.; Balouz, V.; Cameán, C.C.; Cori, C.R.; Kashiwagi, G.A.; Gil, S.A.; Macchiaverna, N.P.; Cardinal, M.V.; Guaimas, F.; Lobo, M.M.; et al. Trypanosoma cruzi surface mucins are involved in the attachment to the Triatoma infestans rectal ampoule. PLoS Negl. Trop. Dis. 2019, 13, e0007418. [Google Scholar] [CrossRef]
- Mortara, R.A.; da Silva, S.; Araguth, M.F.; Blanco, S.A.; Yoshida, N. Polymorphism of the 35- and 50-kilodalton surface glycoconjugates of Trypanosoma cruzi metacyclic trypomastigotes. Infect. Immun. 1992, 60, 4673–4678. [Google Scholar] [CrossRef] [Green Version]
- Nogueira, N.F.S.; Gonzalez, M.S.; Gomes, J.E.; de Souza, W.; Garcia, E.S.; Azambuja, P.; Nohara, L.L.; Almeida, I.C.; Zingales, B.; Colli, W. Trypanosoma cruzi: Involvement of glycoinositolphospholipids in the attachment to the luminal midgut surface of Rhodnius prolixus. Exp. Parasitol. 2007, 116, 120–128. [Google Scholar] [CrossRef]
- D’Orso, I.; Frasch, A.C. TcUBP-1, a developmentally regulated U-rich RNA-binding protein involved in selective mRNA destabilization in trypanosomes. J. Biol. Chem. 2001, 276, 34801–34809. [Google Scholar] [CrossRef] [Green Version]
- Freitas-Junior, L.H.; Briones, M.R.; Schenkman, S. Two distinct groups of mucin-like genes are differentially expressed in the developmental stages of Trypanosoma cruzi. Mol. Biochem. Parasitol. 1998, 93, 101–114. [Google Scholar] [CrossRef]
- Cánepa, G.E.; Degese, M.S.; Budu, A.; Garcia, C.R.S.; Buscaglia, C.A. Involvement of TSSA (trypomastigote small surface antigen) in Trypanosoma cruzi invasion of mammalian cells. Biochem. J. 2012, 444, 211–218. [Google Scholar] [CrossRef] [Green Version]
- Di Noia, J.M.; D’Orso, I.; Aslund, L.; Sánchez, D.O.; Frasch, A.C. The Trypanosoma cruzi mucin family is transcribed from hundreds of genes having hypervariable regions. J. Biol. Chem. 1998, 273, 10843–10850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Noia, J.M.; Buscaglia, C.A.; De Marchi, C.R.; Almeida, I.C.; Frasch, A.C.C. A Trypanosoma cruzi small surface molecule provides the first immunological evidence that Chagas’ disease is due to a single parasite lineage. J. Exp. Med. 2002, 195, 401–413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cámara, M.d.l.M.; Cánepa, G.E.; Lantos, A.B.; Balouz, V.; Yu, H.; Chen, X.; Campetella, O.; Mucci, J.; Buscaglia, C.A. The Trypomastigote Small Surface Antigen (TSSA) regulates Trypanosoma cruzi infectivity and differentiation. PLoS Negl. Trop. Dis. 2017, 11, e0005856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartholomeu, D.C.; Cerqueira, G.C.; Leão, A.C.A.; daRocha, W.D.; Pais, F.S.; Macedo, C.; Djikeng, A.; Teixeira, S.M.R.; El-Sayed, N.M. Genomic organization and expression profile of the mucin-associated surface protein (masp) family of the human pathogen Trypanosoma cruzi. Nucleic Acids Res. 2009, 37, 3407–3417. [Google Scholar] [CrossRef] [Green Version]
- Souza, R.T.; Santos, M.R.M.; Lima, F.M.; El-Sayed, N.M.; Myler, P.J.; Ruiz, J.C.; da Silveira, J.F. New Trypanosoma cruzi Repeated Element That Shows Site Specificity for Insertion. Eukaryot. Cell 2007, 6, 1228–1238. [Google Scholar] [CrossRef] [Green Version]
- Atwood, J.A.; Minning, T.; Ludolf, F.; Nuccio, A.; Weatherly, D.B.; Alvarez-Manilla, G.; Tarleton, R.; Orlando, R. Glycoproteomics of Trypanosoma cruzi trypomastigotes using subcellular fractionation, lectin affinity, and stable isotope labeling. J. Proteome Res. 2006, 5, 3376–3384. [Google Scholar] [CrossRef]
- dos Santos, S.L.; Freitas, L.M.; Lobo, F.P.; Rodrigues-Luiz, G.F.; Mendes, T.A.d.O.; Oliveira, A.C.S.; Andrade, L.O.; Chiari, É.; Gazzinelli, R.T.; Teixeira, S.M.R.; et al. The MASP Family of Trypanosoma cruzi: Changes in Gene Expression and Antigenic Profile during the Acute Phase of Experimental Infection. PLoS Negl. Trop. Dis. 2012, 6, e1779. [Google Scholar] [CrossRef]
- De Pablos, L.M.; González, G.G.; Solano Parada, J.; Seco Hidalgo, V.; Díaz Lozano, I.M.; Gómez Samblás, M.M.; Cruz Bustos, T.; Osuna, A. Differential Expression and Characterization of a Member of the Mucin-Associated Surface Protein Family Secreted by Trypanosoma cruzi. Infect. Immun. 2011, 79, 3993–4001. [Google Scholar] [CrossRef] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | DTU | Size (Mbp) | Contigs | Contig N50 | %GC | Date of Version | Sequencing Method | References |
|---|---|---|---|---|---|---|---|---|
| G | I | 25.17 | 1450 | 74,655 | 47.40 | November 2018 | Roche 454 | [50] |
| Dm28c | I | 53.27 | 636 | 317,638 | 51.60 | May 2018 | Illumina + PacBio | [55] |
| Sylvio X10/1 | I | 38.59 | 27,019 | 2307 | 51.20 | October 2012 | Roche 454 + Illumina | [49,57] |
| Berenice | II | 40.80 | 934 | 148,957 | 51.20 | June 2020 | Illumina + Nanopore | [54] |
| Y | II | 39.34 | 10,127 | 11,782 | 51.43 | October 2017 | Illumina | [47] |
| 231 | III | 35.36 | 8469 | 14,202 | 48.60 | January 2018 | Illumina | [48] |
| Bug2148 | V | 55.22 | 934 | 196,760 | 51.63 | October 2017 | PacBio | [53] |
| CL | VI | 65.00 | 7764 | 73,547 | 39.80 | November 2018 | Roche 454 | [50] |
| TCC | VI | 87.06 | 1236 | 264,196 | 51.70 | May 2018 | Illumina + PacBio | [55] |
| CL Brener | VI | 89.94 | 32,746 | 14,669 | 51.70 | July 2005 | Sanger | [42] |
| BNEL | VI | 32.53 | 41 | 870,934 | 43.94 | December 2015 | Sanger | [58] |
| BEL | VI | 32.53 | 41 | 870,934 | 40.35 | December 2015 | Sanger | [58] |
| T. c. marinkellei B7 strain | --- | 38.65 | 23,154 | 2846 | 50.90 | October 2012 | Roche 454 + Illumina | [51] |
| TS Groups of T. cruzi | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Group I | Group II | Group III | Group IV | Group V | Group VI | Group VII | Group VIII | ||
| Trypanosoma species with TS sequence similarity | T. c. marinkellei | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ |
| T. rangeli | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | |
| T. conorhini | ✓ | ✓ | ✓ | ||||||
| T. dionisii | ✓ | ✓ | |||||||
| T. evansi | ✓ | ||||||||
| T. congolense | ✓ | ||||||||
| T. vivax | ✓ | ||||||||
| T. grayi | ✓ | ||||||||
| T. carassii | ✓ | ||||||||
| T. brucei | ✓ | ||||||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Herreros-Cabello, A.; Callejas-Hernández, F.; Gironès, N.; Fresno, M. Trypanosoma Cruzi Genome: Organization, Multi-Gene Families, Transcription, and Biological Implications. Genes 2020, 11, 1196. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11101196
Herreros-Cabello A, Callejas-Hernández F, Gironès N, Fresno M. Trypanosoma Cruzi Genome: Organization, Multi-Gene Families, Transcription, and Biological Implications. Genes. 2020; 11(10):1196. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11101196
Chicago/Turabian StyleHerreros-Cabello, Alfonso, Francisco Callejas-Hernández, Núria Gironès, and Manuel Fresno. 2020. "Trypanosoma Cruzi Genome: Organization, Multi-Gene Families, Transcription, and Biological Implications" Genes 11, no. 10: 1196. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11101196