Hsa-miR-375/RASD1 Signaling May Predict Local Control in Early Breast Cancer


 ,
,
Abstract
:1. Introduction
2. Methods
2.1. Patients
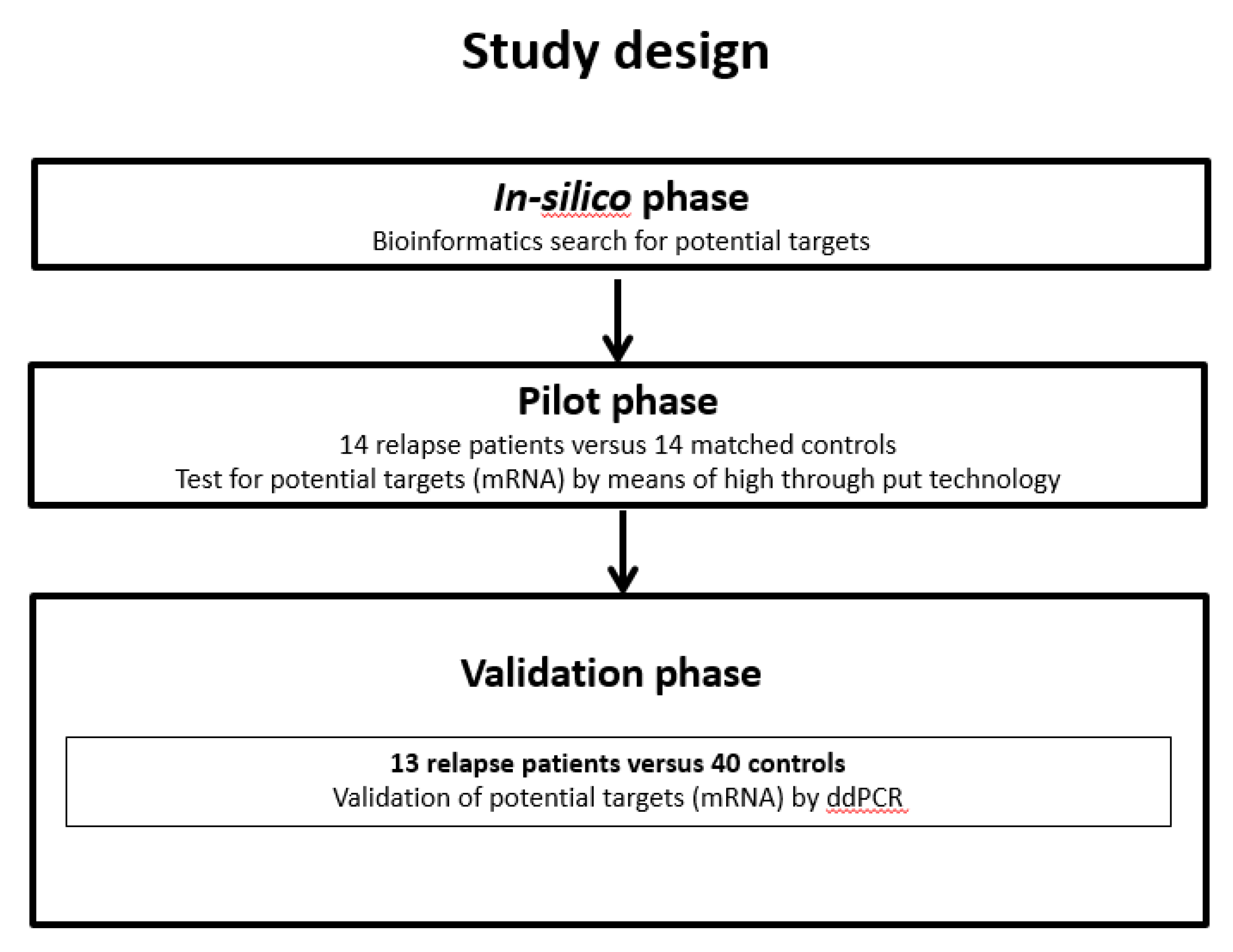
2.2. Experimental Design
2.3. Tissue Samples
2.4. Pilot Phase: Microarray
2.5. Validation Phase: Droplet Digital PCR
2.5.1. Tissue Processing
2.5.2. Droplet Digital PCR
2.5.3. Data Analysis
2.6. Statistics
3. Results
3.1. Patients
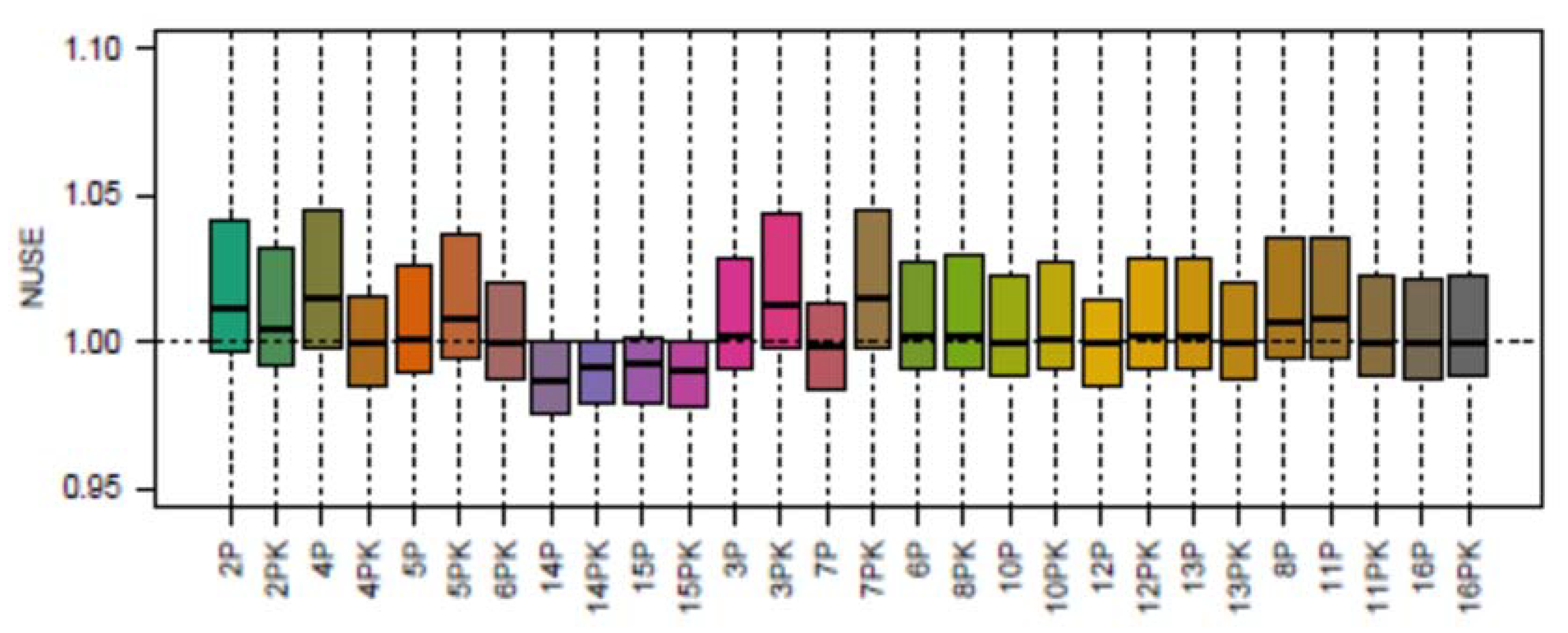
3.2. Pilot Phase
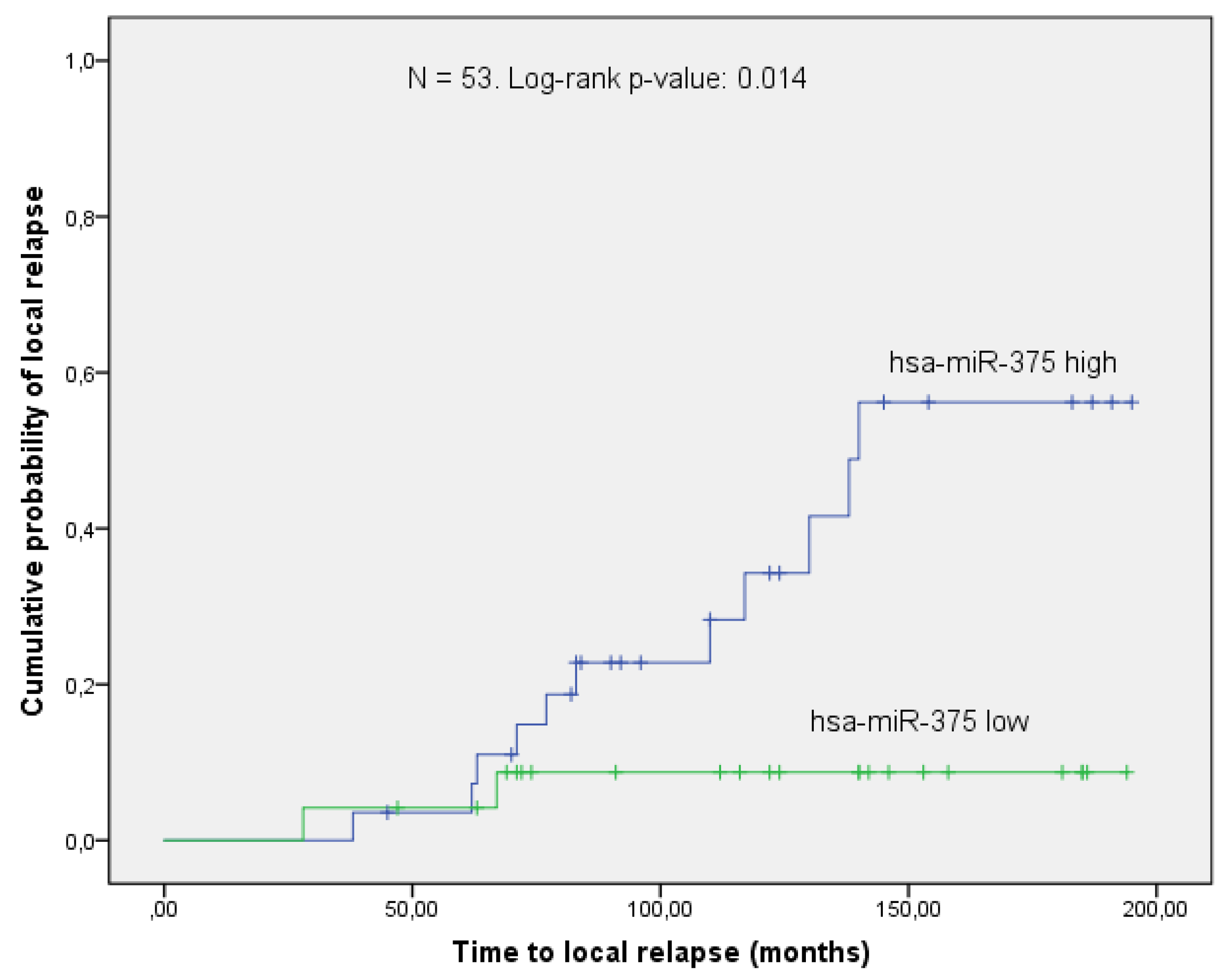
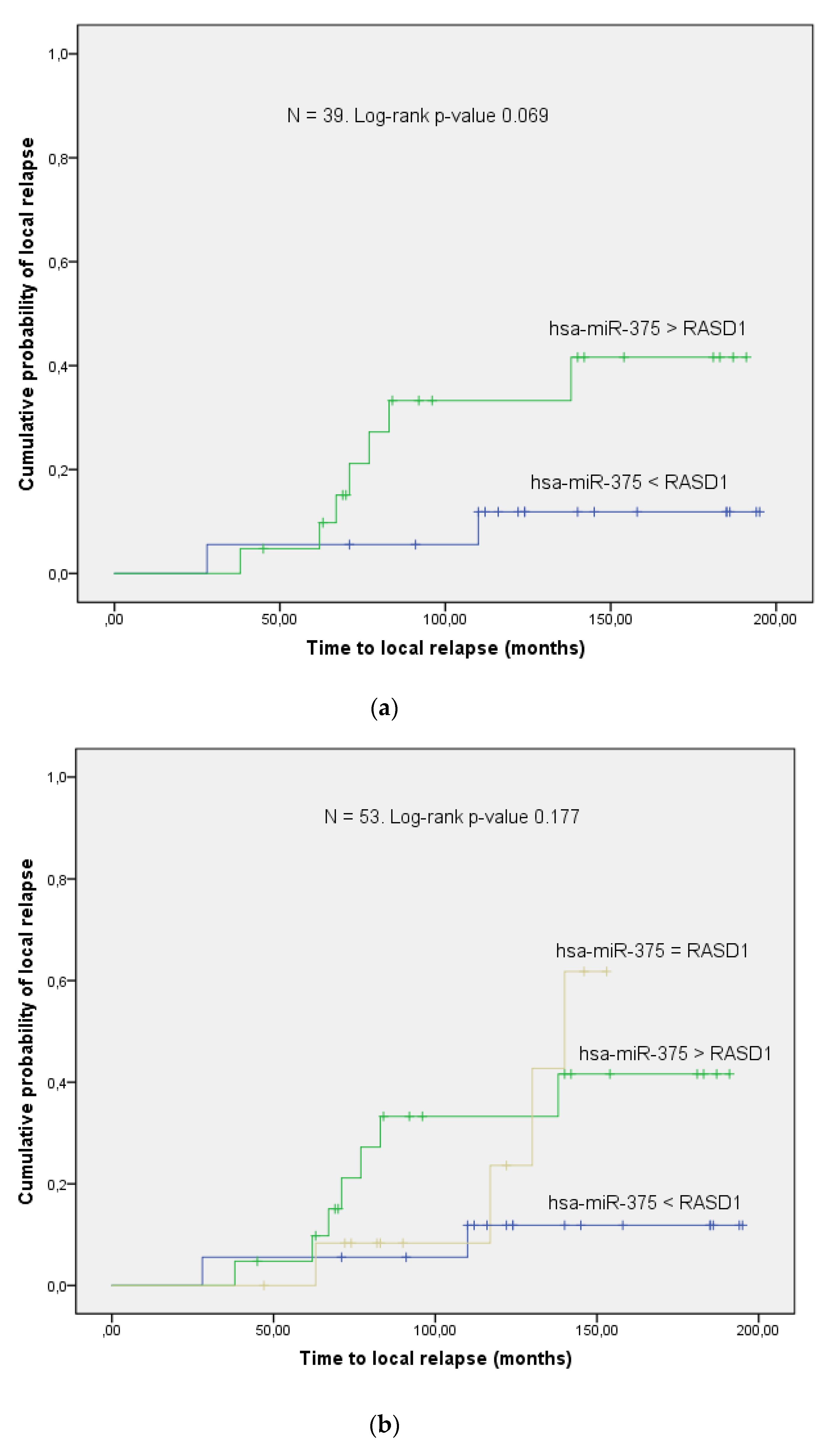
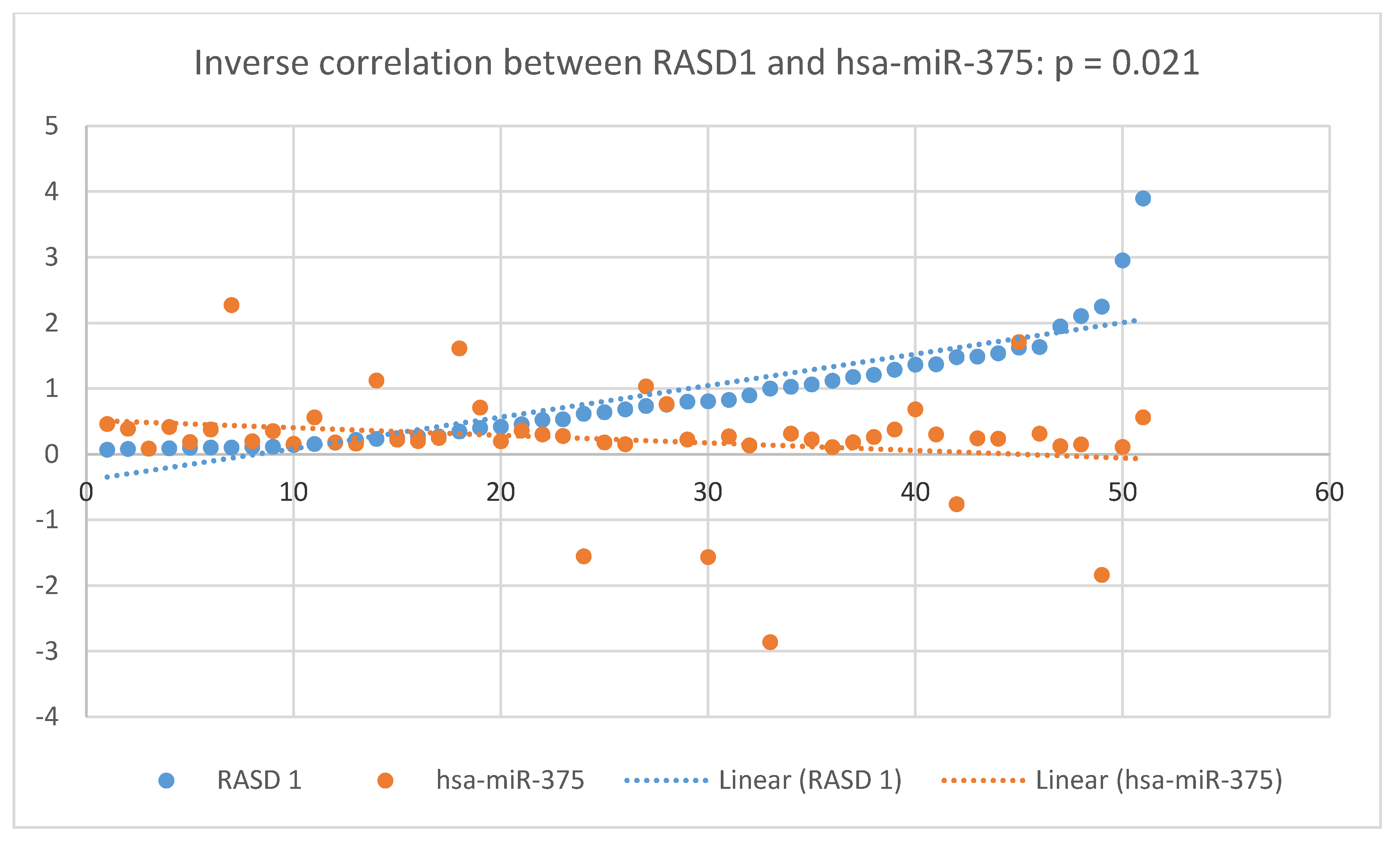
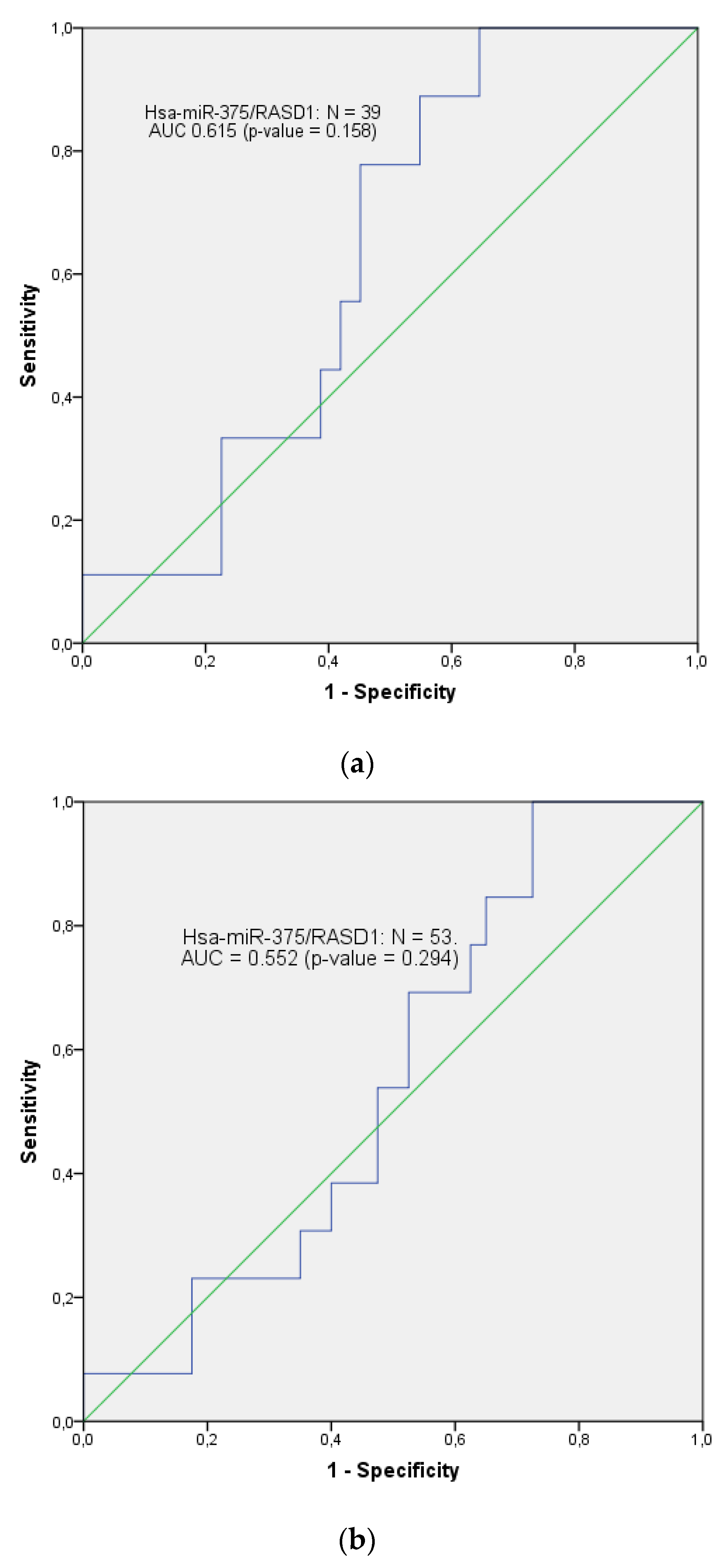
3.3. Validation Phase
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AKT | protein kinase B |
| AUC | area under the curve |
| BCT | breast conserving therapy |
| ddPCR | droplet digital PCR |
| EBCTB | early breast cancer trialist group |
| EEF | eukaryotic elongation factor 2 |
| ER-α | estrogen receptor α |
| FFPE | formalin fixed paraffin embedded |
| HUVEC | human umbilical vein endothelial cells |
| LC | local control |
| LIMMA | linear models for microarray analysis |
| MIQE | minimum information for publication of quantitative digital PCR experiments |
| miR | microRNA, miRNA |
| OS | overall survival |
| p-S6 | phosphorylated ribosomal protein S6 |
| p-GSK-3β | phosphorylated glycogen synthase kinase 3 β |
| PFS | progression free survival |
| PI3K | phosphatidylinositol 3 kinase |
| PSVM | potential support vector machine |
| RASD1 | dexamethasone induced ras-related protein 1 |
| RISC | RNA induced silencing complex |
| ROC | receiver operating curve |
| RT | radiotherapy |
| Sec23A | saccharomyces cerevisiae 23 homolog A |
Appendix A







References
- Carioli, G.; Malvezzi, M.; Rodriguez, T.; Bertuccio, P.; Negri, E.; La Vecchia, C. Trends and predictions to 2020 in breast cancer mortality in Europe. Breast 2017, 36, 89–95. [Google Scholar] [CrossRef]
- Malvezzi, M.; Carioli, G.; Bertuccio, P.; Boffetta, P.; Levi, F.; La Vecchia, C.; Negri, E. European cancer mortality predictions for the year 2019 with focus on breast cancer. Ann. Oncol. 2019, 30, 781–787. [Google Scholar] [CrossRef] [Green Version]
- Darby, S.; McGale, P.; Correa, C.; Taylor, C.; Arriagada, R.; Clarke, M.; Cutter, D.; Davies, C.; Ewertz, M.; Godwin, J.; et al. Effect of radiotherapy after breast-conserving surgery on 10-year recurrence and 15-year breast cancer death: Meta-analysis of individual patient data for 10,801 women in 17 randomised trials. Lancet 2011, 378, 1707–1716. [Google Scholar] [PubMed] [Green Version]
- Sautter-Bihl, M.L.; Sedlmayer, F.; Budach, W.; Dunst, J.; Feyer, P.; Fietkau, R.; Haase, W.; Harms, W.; Rodel, C.; Souchon, R.; et al. One life saved by four prevented recurrences? Update of the Early Breast Cancer Trialists confirms: Postoperative radiotherapy improves survival after breast conserving surgery. Strahlenther Onkol. 2012, 188, 461–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartelink, H.; Horiot, J.C.; Poortmans, P.; Struikmans, H.; Van den Bogaert, W.; Barillot, I.; Fourquet, A.; Borger, J.; Jager, J.; Hoogenraad, W.; et al. Recurrence rates after treatment of breast cancer with standard radiotherapy with or without additional radiation. N. Engl. J. Med. 2001, 345, 1378–1387. [Google Scholar] [CrossRef] [PubMed]
- Whelan, T.; MacKenzie, R.; Julian, J.; Levine, M.; Shelley, W.; Grimard, L.; Lada, B.; Lukka, H.; Perera, F.; Fyles, A.; et al. Randomized trial of breast irradiation schedules after lumpectomy for women with lymph node-negative breast cancer. J. Natl. Cancer Inst. 2002, 94, 1143–1150. [Google Scholar] [CrossRef] [Green Version]
- Bentzen, S.M.; Agrawal, R.K.; Aird, E.G.; Barrett, J.M.; Barrett-Lee, P.J.; Bliss, J.M.; Brown, J.; Dewar, J.A.; Dobbs, H.J.; Haviland, J.S.; et al. The UK Standardisation of Breast Radiotherapy (START) Trial A of radiotherapy hypofractionation for treatment of early breast cancer: A randomised trial. Lancet Oncol. 2008, 9, 331–341. [Google Scholar]
- Goldhirsch, A.; Wood, W.C.; Coates, A.S.; Gelber, R.D.; Thurlimann, B.; Senn, H.J. Strategies for subtypes--dealing with the diversity of breast cancer: Highlights of the St. Gallen International Expert Consensus on the Primary Therapy of Early Breast Cancer 2011. Ann. Oncol. 2011, 22, 1736–1747. [Google Scholar] [CrossRef]
- Iorio, M.V.; Ferracin, M.; Liu, C.G.; Veronese, A.; Spizzo, R.; Sabbioni, S.; Magri, E.; Pedriali, M.; Fabbri, M.; Campiglio, M.; et al. MicroRNA gene expression deregulation in human breast cancer. Cancer Res. 2005, 65, 7065–7070. [Google Scholar] [CrossRef] [Green Version]
- Tang, W.; Li, G.S.; Li, J.D.; Pan, W.Y.; Shi, Q.; Xiong, D.D.; Mo, C.H.; Zeng, J.J.; Chen, G.; Feng, Z.B.; et al. The role of upregulated miR-375 expression in breast cancer: An in vitro and in silico study. Pathol. Res. Pract. 2020, 216, 152754. [Google Scholar] [CrossRef]
- Zehentmayr, F.; Hauser-Kronberger, C.; Zellinger, B.; Hlubek, F.; Schuster, C.; Bodenhofer, U.; Fastner, G.; Deutschmann, H.; Steininger, P.; Reitsamer, R.; et al. Hsa-miR-375 is a predictor of local control in early stage breast cancer. Clin. Epigenet. 2016, 8, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simonini, P.D.S.R.; Breiling, A.; Gupta, N.; Malekpour, M.; Youns, M.; Omranipour, R.; Malekpour, F.; Volinia, S.; Croce, C.M.; Najmabadi, H.; et al. Epigenetically deregulated microRNA-375 is involved in a positive feedback loop with estrogen receptor α in breast cancer cells. Cancer Res. 2010, 70, 9175–9184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Zhang, X.; Wang, Y.; Ye, Y.; Huang, Z.Q. Differential ability of formononetin to stimulate proliferation of endothelial cells and breast cancer cells via a feedback loop involving MicroRNA-375, RASD1, and ER α. Mol. Carcinog. 2018, 57, 817–830. [Google Scholar] [CrossRef]
- Wang, Y.; Dong, X.H.; Li, Z.Z.; Wang, W.B.; Tian, J.; Chen, J. Downregulated RASD1 and upregulated miR-375 are involved in protective effects of calycosin on cerebral ischemia/reperfusion rats. J. Neurol. Sci. 2014, 339, 144–148. [Google Scholar] [CrossRef] [PubMed]
- Munagala, R.; Aqil, F.; Vadhanam, M.V.; Gupta, R.C. MicroRNA ‘signature’ during estrogen-mediated mammary carcinogenesis and its reversal by ellagic acid intervention. Cancer Lett. 2013, 339, 175–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, S.F.; Jin, L.; Liu, G.P.; Wang, P.; Sun, Z.H.; Cao, Y.J.; Shi, H.L.; Liu, X.J.; Shi, Q.; Zhou, X.P.; et al. Overexpression of RASD1 inhibits glioma cell migration/invasion and inactivates the AKT/mTOR signaling pathway. Sci. Rep. 2017, 7, 3202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, D.; Wilson, J.M.; Harvel, N.; Liu, J.; Pei, L.; Huang, S.; Hawthorn, L.; Shi, H. A systematic evaluation of miRNA:mRNA interactions involved in the migration and invasion of breast cancer cells. J. Transl. Med. 2013, 11, 57. [Google Scholar] [CrossRef] [Green Version]
- Szczyrba, J.; Nolte, E.; Wach, S.; Kremmer, E.; Stohr, R.; Hartmann, A.; Wieland, W.; Wullich, B.; Grasser, F.A. Downregulation of Sec23A protein by miRNA-375 in prostate carcinoma. Mol. Cancer Res. 2011, 9, 791–800. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Marian, C.; Makambi, K.H.; Kosti, O.; Kallakury, B.V.; Loffredo, C.A.; Zheng, Y.L. MicroRNA-9 as potential biomarker for breast cancer local recurrence and tumor estrogen receptor status. PLoS ONE 2012, 7, e39011. [Google Scholar] [CrossRef]
- Jonsdottir, K.; Janssen, S.R.; Da Rosa, F.C.; Gudlaugsson, E.; Skaland, I.; Baak, J.P.; Janssen, E.A. Validation of expression patterns for nine miRNAs in 204 lymph-node negative breast cancers. PLoS ONE 2012, 7, e48692. [Google Scholar] [CrossRef] [Green Version]
- Hoppe, R.; Achinger-Kawecka, J.; Winter, S.; Fritz, P.; Lo, W.Y.; Schroth, W.; Brauch, H. Increased expression of miR-126 and miR-10a predict prolonged relapse-free time of primary oestrogen receptor-positive breast cancer following tamoxifen treatment. Eur. J. Cancer 2013, 49, 3598–3608. [Google Scholar] [CrossRef] [PubMed]
- Lyng, M.B.; Laenkholm, A.V.; Sokilde, R.; Gravgaard, K.H.; Litman, T.; Ditzel, H.J. Global microRNA expression profiling of high-risk ER+ breast cancers from patients receiving adjuvant tamoxifen mono-therapy: A DBCG study. PLoS ONE 2012, 7, e36170. [Google Scholar] [CrossRef] [PubMed]
- Perez-Rivas, L.G.; Jerez, J.M.; Carmona, R.; de Luque, V.; Vicioso, L.; Claros, M.G.; Viguera, E.; Pajares, B.; Sanchez, A.; Ribelles, N.; et al. A microRNA signature associated with early recurrence in breast cancer. PLoS ONE 2014, 9, e91884. [Google Scholar] [CrossRef] [PubMed]
- Tuomarila, M.; Luostari, K.; Soini, Y.; Kataja, V.; Kosma, V.M.; Mannermaa, A. Overexpression of microRNA-200c predicts poor outcome in patients with PR-negative breast cancer. PLoS ONE 2014, 9, e109508. [Google Scholar] [CrossRef] [PubMed]
- Svoboda, M.; Sana, J.; Redova, M.; Navratil, J.; Palacova, M.; Fabian, P.; Slaby, O.; Vyzula, R. MiR-34b is associated with clinical outcome in triple-negative breast cancer patients. Diagn. Pathol. 2012, 7, 31. [Google Scholar] [CrossRef] [Green Version]
- Huang, E.; Buchholz, T.A.; Meric, F.; Krishnamurthy, S.; Mirza, N.Q.; Ames, F.C.; Feig, B.W.; Kuerer, H.M.; Ross, M.I.; Singletary, S.E.; et al. Classifying local disease recurrences after breast conservation therapy based on location and histology: New primary tumors have more favorable outcomes than true local disease recurrences. Cancer 2002, 95, 2059–2067. [Google Scholar] [CrossRef]
- Irizarry, R.A.; Bolstad, B.M.; Collin, F.; Cope, L.M.; Hobbs, B.; Speed, T.P. Summaries of affymetrix GeneChip probe level data. Nucleic Acids Res. 2003, 31, e15. [Google Scholar] [CrossRef]
- Irizarry, R.A.; Hobbs, B.; Collin, F.; Beazer-Barclay, Y.D.; Antonellis, K.J.; Scherf, U.; Speed, T.P. Exploration, normalization, and summaries of high density oligonucleotide array probe level data. Biostatistics 2003, 4, 249–264. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, B.S.; Irizarry, R.A. A framework for oligonucleotide microarray preprocessing. Bioinformatics 2010, 26, 2363–2367. [Google Scholar] [CrossRef]
- Huggett, J.F.; Foy, C.A.; Benes, V.; Emslie, K.; Garson, J.A.; Haynes, R.; Hellemans, J.; Kubista, M.; Nolan, R.D.M.T.; Pfaffl, M.W.; et al. The Digital MIQE Guidelines: Minimum Information for Publication of Quantitative Digital PCR Experiments. Clin. Chem. 2013, 59, 892–902. [Google Scholar] [CrossRef]
- Smyth, G.K. Linear models and empirical bayes methods for assessing differential expression in microarray experiments. Stat. Appl. Genet. Mol. Biol. 2004, 3. [Google Scholar] [CrossRef] [PubMed]
- Dhawan, A.; Scott, J.G.; Harris, A.L.; Buffa, F.M. Pan-cancer characterisation of microRNA across cancer hallmarks reveals microRNA-mediated downregulation of tumour suppressors. Nat. Commun. 2018, 9, 5228. [Google Scholar] [CrossRef]
- Hochreiter, S.; Obermayer, K. Support vector machines for dyadic data. Neural. Comput. 2006, 18, 1472–1510. [Google Scholar] [CrossRef] [PubMed]
- Palma, D.A.; Olson, R.; Harrow, S.; Gaede, S.; Louie, A.V.; Haasbeek, C.; Mulroy, L.; Lock, M.; Rodrigues, G.B.; Yaremko, B.P.; et al. Stereotactic ablative radiotherapy versus standard of care palliative treatment in patients with oligometastatic cancers (SABR-COMET): A randomised, phase 2, open-label trial. Lancet 2019, 393, 2051–2058. [Google Scholar] [CrossRef]
- Rubinstein, L.V.; Korn, E.L.; Freidlin, B.; Hunsberger, S.; Ivy, S.P.; Smith, M.A. Design issues of randomized phase II trials and a proposal for phase II screening trials. J. Clin. Oncol. 2005, 23, 7199–7206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Witkos, T.M.; Koscianska, E.; Krzyzosiak, W.J. Practical Aspects of microRNA Target Prediction. Curr. Mol. Med. 2011, 11, 93–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, B.P.; Burge, C.B.; Bartel, D.P. Conserved seed pairing, often flanked by adenosines, indicates that thousands of human genes are microRNA targets. Cell 2005, 120, 15–20. [Google Scholar] [CrossRef] [Green Version]
- Kertesz, M.; Iovino, N.; Unnerstall, U.; Gaul, U.; Segal, E. The role of site accessibility in microRNA target recognition. Nat. Genet. 2007, 39, 1278–1284. [Google Scholar] [CrossRef]
- Vaidyanathan, G.; Cismowski, M.J.; Wang, G.; Vincent, T.S.; Brown, K.D.; Lanier, S.M. The Ras-related protein AGS1/RASD1 suppresses cell growth. Oncogene 2004, 23, 5858–5863. [Google Scholar] [CrossRef] [Green Version]
- Koga, T.; Iwasaki, H.; Ishiguro, M.; Matsuzaki, A.; Kikuchi, M. Losses in chromosomes 17, 19, and 22q in neurofibromatosis type 1 and sporadic neurofibromas: A comparative genomic hybridization analysis. Cancer Genet. Cytogenet. 2002, 136, 113–120. [Google Scholar] [CrossRef]
- Tang, Y.J.; Huang, J.; Tsushima, H.; Ban, G.I.; Zhang, H.; Oristian, K.M.; Puviindran, V.; Williams, N.; Ding, X.; Ou, J.; et al. Tracing Tumor Evolution in Sarcoma Reveals Clonal Origin of Advanced Metastasis. Cell Rep. 2019, 28, 2837–2850.e5. [Google Scholar] [CrossRef] [PubMed]
- Both, J.; Wu, T.; Ten Asbroek, A.L.; Baas, F.; Hulsebos, T.J. Oncogenic Properties of Candidate Oncogenes in Chromosome Region 17p11.2p12 in Human Osteosarcoma. Cytogenet. Genome. Res. 2016, 150, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Wong, K.Y.; Yao, Q.; Yuan, L.Q.; Li, Z.; Ma, E.S.K.; Chim, C.S. Frequent functional activation of RAS signalling not explained by RAS/RAF mutations in relapsed/refractory multiple myeloma. Sci. Rep. 2018, 8, 13522. [Google Scholar] [CrossRef] [PubMed]
- Ward, A.; Balwierz, A.; Zhang, J.D.; Kublbeck, M.; Pawitan, Y.; Hielscher, T.; Wiemann, S.; Sahin, O. Re-expression of microRNA-375 reverses both tamoxifen resistance and accompanying EMT-like properties in breast cancer. Oncogene 2013, 32, 1173–1182. [Google Scholar] [CrossRef] [Green Version]
- Tabas-Madrid, D.; Nogales-Cadenas, R.; Pascual-Montano, A. GeneCodis3: A non-redundant and modular enrichment analysis tool for functional genomics. Nucleic Acids Res. 2012, 40, W478–W483. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient Characteristics and Treatment | Pilot Phase n = 28 | Validation Phase n = 53 | ||||||
|---|---|---|---|---|---|---|---|---|
| Parameters | Relapse n = 14 | Control n = 14 | p-Value | Relapse n = 13 | Control n = 40 | p-Value | ||
| Patient characteristics | Age at diagnosis (years) | Median | 53.5 | 56 | 0.84 | 57 | 54 | 0.06 |
| Range | 40–71 | 41–74 | 40–79 | 41–78 | ||||
| Menopause (n) | No | 3 (21%) | 3 (21%) | 0.75 | 2 (15%) | 16 (40%) | 0.05 | |
| Yes | 7 (50%) | 8 (57%) | 10 (77%) | 18 (45%) | ||||
| Unclear | 4 (29%) | 3 (22%) | 1 (8%) | 6 (15%) | ||||
| T (n) | T1 | 9 (64%) | 8 (57%) | 1 | 11 (85%) | 33 (83%) | 0.86 | |
| T2 | 5 (36%) | 6 (43%) | 2 (15%) | 7 (17%) | ||||
| n (n) | N0 | 13 (93%) | 12 (86%) | 0.77 | 9 (69%) | 30 (75%) | 0.55 | |
| N1 | 1 (7%) | 1 (7%) | 4 (31%) | 10 (25%) | ||||
| N2 | 0 (0%) | 1 (7%) | 0 (0%) | 0 (0%) | ||||
| M (n) | M0 | 14 (100%) | 14 (100%) | 1 | 13 (100%) | 40 (100%) | 1 | |
| Grading (n) | G1 | 0 (0%) | 0 (0%) | 0.87 | 0 (0%) | 4 (10%) | 0.56 | |
| G2 | 7 (50%) | 8 (57%) | 8 (62%) | 22 (55%) | ||||
| G3 | 7 (50%) | 6 (43%) | 5 (38%) | 14 (35%) | ||||
| Histology (n) | IDC 1 | 12 (86%) | 13 (93%) | 0.77 | 8 (62%) | 33 (83%) | 0.39 | |
| ILC 2 | 2 (14%) | 1 (7%) | 3 (23%) | 2 (5%) | ||||
| Other | 0 (0%) | 0 (0%) | 2 (15%) | 5 (13%) | ||||
| In situ component (n) | Yes | 12 (86%) | 8 (57%) | 0.11 | 7 (54%) | 20 (50%) | 0.81 | |
| No | 2 (14%) | 6 (43%) | 6 (46%) | 20 (50%) | ||||
| Receptors (n) | ER positive 3 | 6 (43%) | 8 (57%) | 0.54 | 10 (77%) | 30 (75%) | 0.89 | |
| ER negative | 8 (57%) | 6 (43%) | 3 (23%) | 10 (25%) | ||||
| PR positive 4 | 5 (36%) | 7 (50%) | 0.54 | 8 (62%) | 18 (70%) | 0.57 | ||
| PR negative | 9 (64%) | 7 (50%) | 5 (38%) | 12 (30%) | ||||
| her2neu (n) | Positive | 8 (57%) | 2 (14%) | 0.06 | 4 (31%) | 21 (53%) | 0.56 | |
| Negative | 4 (29%) | 9 (64%) | 9 (69%) | 13 (32%) | ||||
| Not assessable | 2 (14%) | 3 (21%) | 0 (0%) | 6 (15%) | ||||
| Proliferation index (n) | ki67 < 20% | 9 (64%) | 6 (43%) | 0.54 | 4 (31%) | 24 (60%) | 0.91 | |
| ki67 > 20% | 5 (36%) | 7 (50%) | 7 (54%) | 13 (32%) | ||||
| Not assessable | 0 (0%) | 1 (7%) | 2 (15%) | 3 (8%) | ||||
| Treatment | Boost | Intraoperative (n) | 8 (57%) | 7 (50%) | 0.8 | 7 (54%) | 21 (53%) | 0.22 |
| Percutaneous (n) | 6 (43%) | 7 (50%) | 4 (31%) | 19 (47%) | ||||
| None | 0 (0%) | 0 (0%) | 2 (15%) | 0 (0%) | ||||
| Intraoperative dose (Gy) | 10 | 10 | 10 | 10 | ||||
| Percutaneous dose (Gy) | 12 | 12 | 12 | 12 | ||||
| WBRT 5 dose (Gy) | Median | 54 | 54 | 1 | 54 | 54 | 0.2 | |
| Range | 52.5–61.2 | 51.0–57.8 | 51–57.8 | 50–54 | ||||
| Surgery (n) | BCT 6 | 14 (100%) | 14 (100%) | 1 | 13 (100%) | 40 (100%) | 1 | |
| Mastectomy | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) | ||||
| Re-excision (n) | Yes | 6 (43%) | 3 (21%) | 0.54 | 5 (38%) | 18 (45%) | 0.69 | |
| No | 8 (57%) | 11 (79%) | 8 (62%) | 22 (55%) | ||||
| Year of surgery (n) | Before 1998 | 3 (21%) | 4 (29%) | 1 | 5 (38%) | 15 (38%) | 0.434 | |
| Since 1998 | 11 (79%) | 10 (61%) | 8 (62%) | 25 (62%) | ||||
| Chemotherapy (n) | Yes | 7 (50%) | 5 (36%) | 0.21 | 4 (31%) | 11 (28%) | 0.86 | |
| No | 7 (50%) | 9 (64%) | 9 (69%) | 29 (72%) | ||||
| Antihormonal treatment (n) | Yes | 7 (50%) | 6 (43%) | 1 | 9 (69%) | 22 (55%) | 0.46 | |
| No | 7 (50%) | 8 (57%) | 4 (31%) | 16 (40%) | ||||
| Unclear | 0 (0%) | 0 (0%) | 0 (0%) | 2 (5%) | ||||
| Tumor burden in biopsy (%) | Median | 70 | 60 | 0.4 | 70 | 70 | 0.76 | |
| Range | 10–90 | 10–90 | 10–90 | 30–90 | ||||
| Patient Characteristics and Treatment | Pilot Phase n = 28 | Validation Phase n = 54 | |||
|---|---|---|---|---|---|
| Parameters | Relapse n = 14 | Control n = 14 | Relapse n = 13 | Control n = 41 | |
| Time to local failure (months) | Median | 44 | x | 77 | x |
| Range | 15–123 | x | 28–140 | x | |
| Time to distant metastasis (months) | Median | 61 | x | 71 | 109 |
| Range | 28–94 | x | 21–78 | x | |
| Follow-up (months) | Median | 121.5 | 130.5 | 95 | 122 |
| Range | 40–186 | 72–200 | 44–214 | 45–194 | |
| Cancer related deaths (n) | 3 | 0 | 3 | 1 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zellinger, B.; Bodenhofer, U.; Engländer, I.A.; Kronberger, C.; Strasser, P.; Grambozov, B.; Fastner, G.; Stana, M.; Reitsamer, R.; Sotlar, K.; et al. Hsa-miR-375/RASD1 Signaling May Predict Local Control in Early Breast Cancer. Genes 2020, 11, 1404. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11121404
Zellinger B, Bodenhofer U, Engländer IA, Kronberger C, Strasser P, Grambozov B, Fastner G, Stana M, Reitsamer R, Sotlar K, et al. Hsa-miR-375/RASD1 Signaling May Predict Local Control in Early Breast Cancer. Genes. 2020; 11(12):1404. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11121404
Chicago/Turabian StyleZellinger, Barbara, Ulrich Bodenhofer, Immanuela A. Engländer, Cornelia Kronberger, Peter Strasser, Brane Grambozov, Gerd Fastner, Markus Stana, Roland Reitsamer, Karl Sotlar, and et al. 2020. "Hsa-miR-375/RASD1 Signaling May Predict Local Control in Early Breast Cancer" Genes 11, no. 12: 1404. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11121404