Asymmetric Isolation and the Evolution of Behaviors Influencing Dispersal: Rheotaxis of Guppies above Waterfalls


Abstract
:1. Introduction
2. Materials and Methods
2.1. History of an Introduction and Its Effects
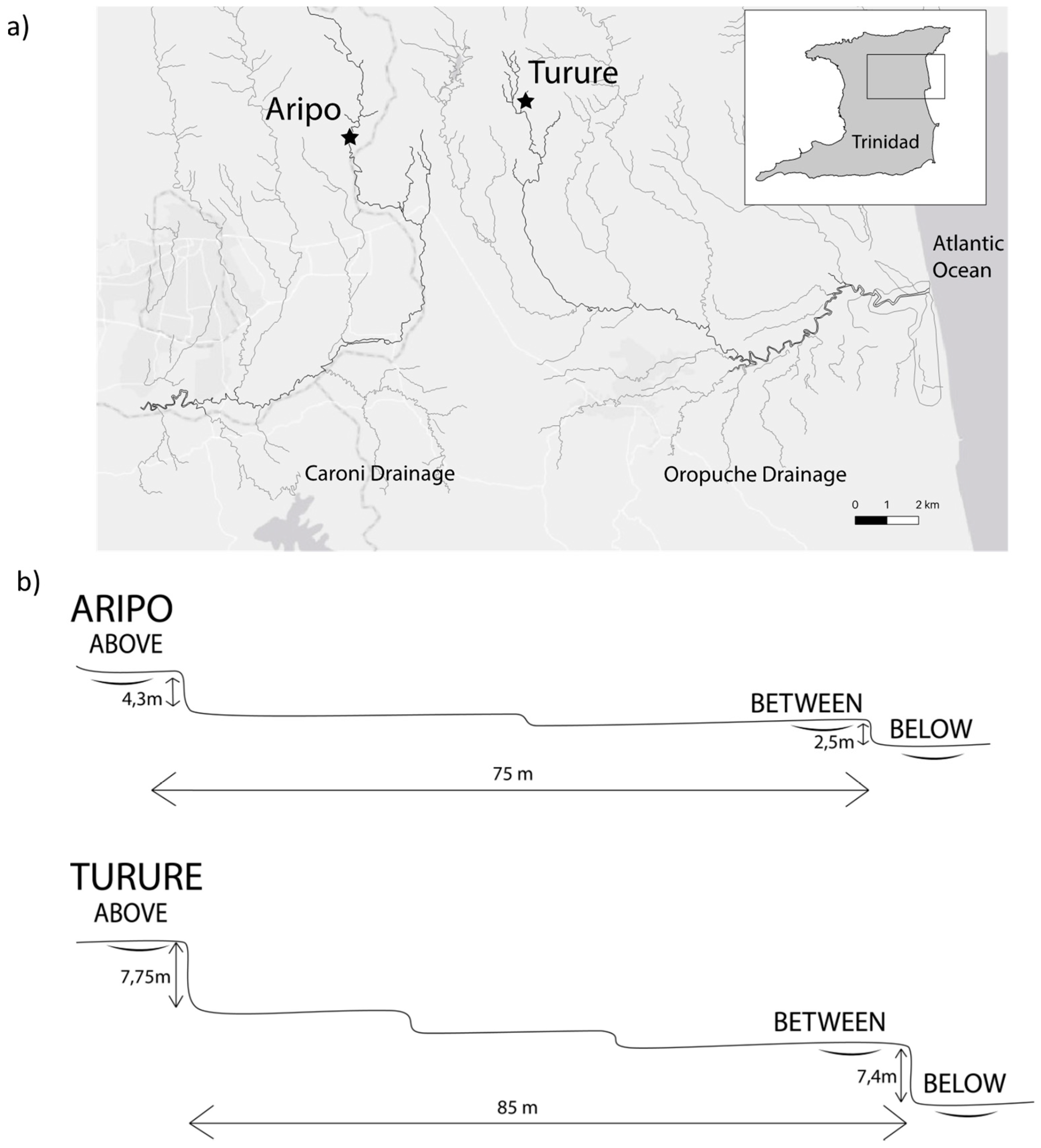
2.2. Fish Sampling
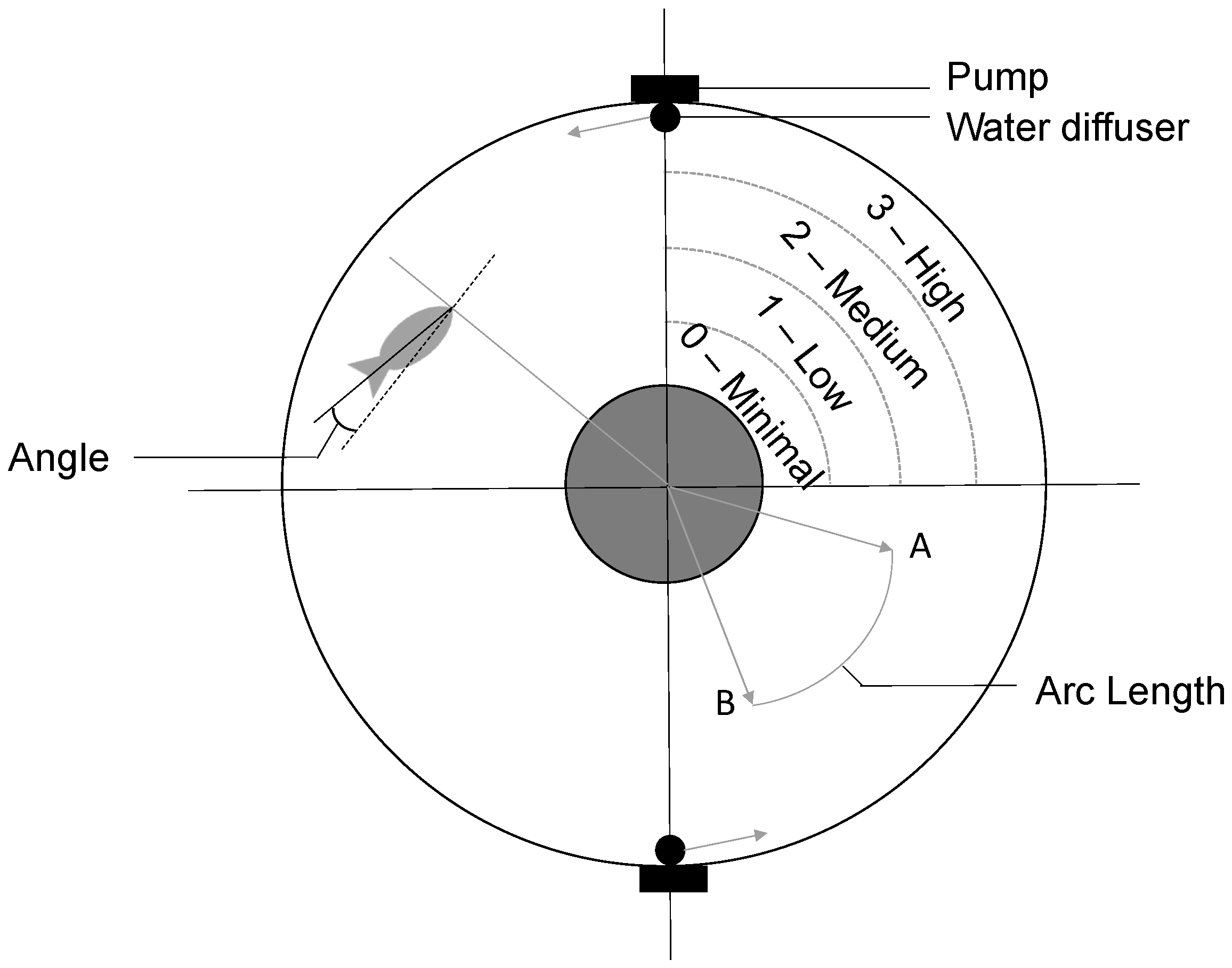
2.3. Rheotaxis Apparatus
2.4. Rheotaxis Trials
2.5. Video Analysis
2.6. Statistical Analyses
3. Results
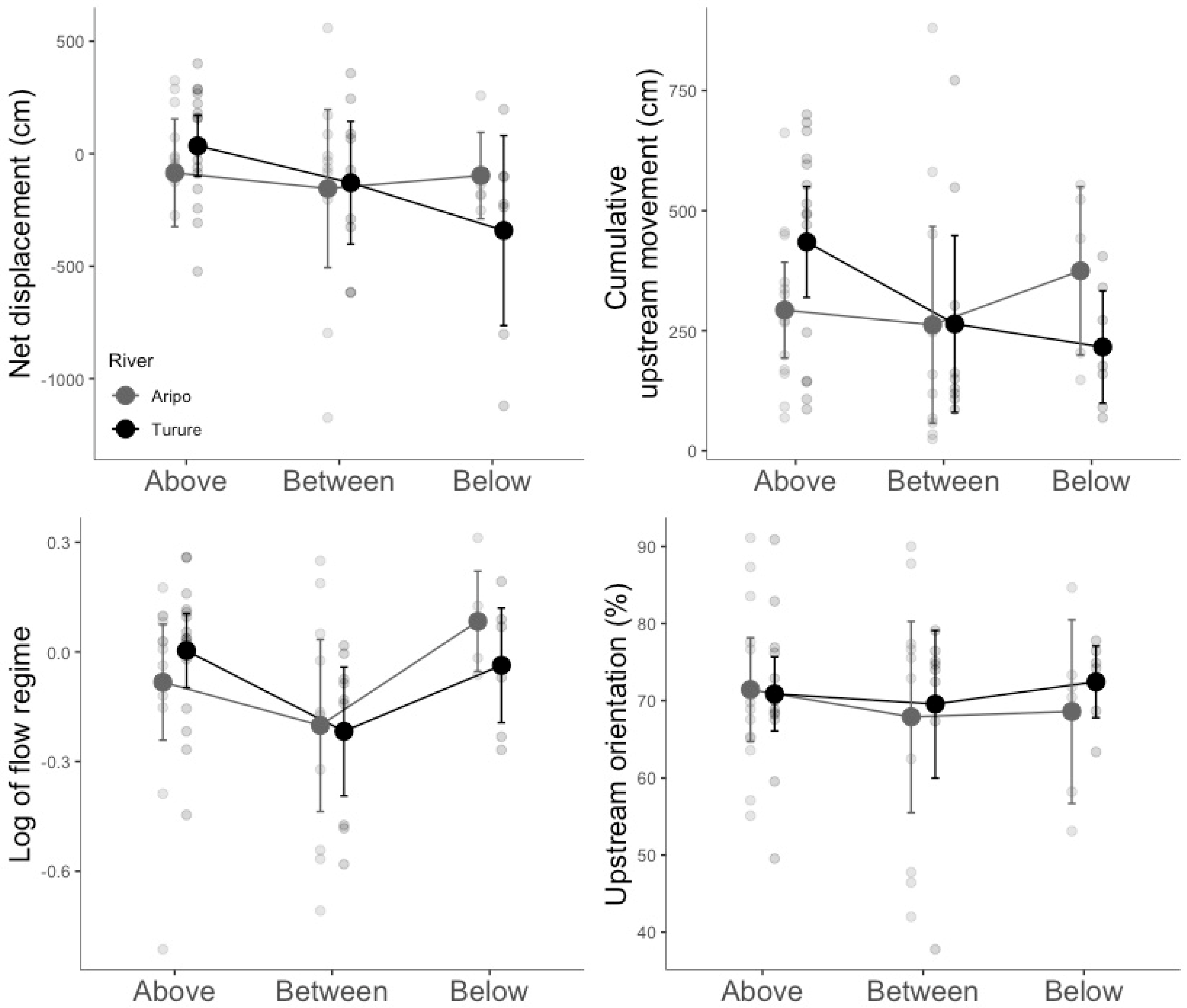
3.1. Summary Statistics
3.2. Linear Models for Rheotactic Behavior
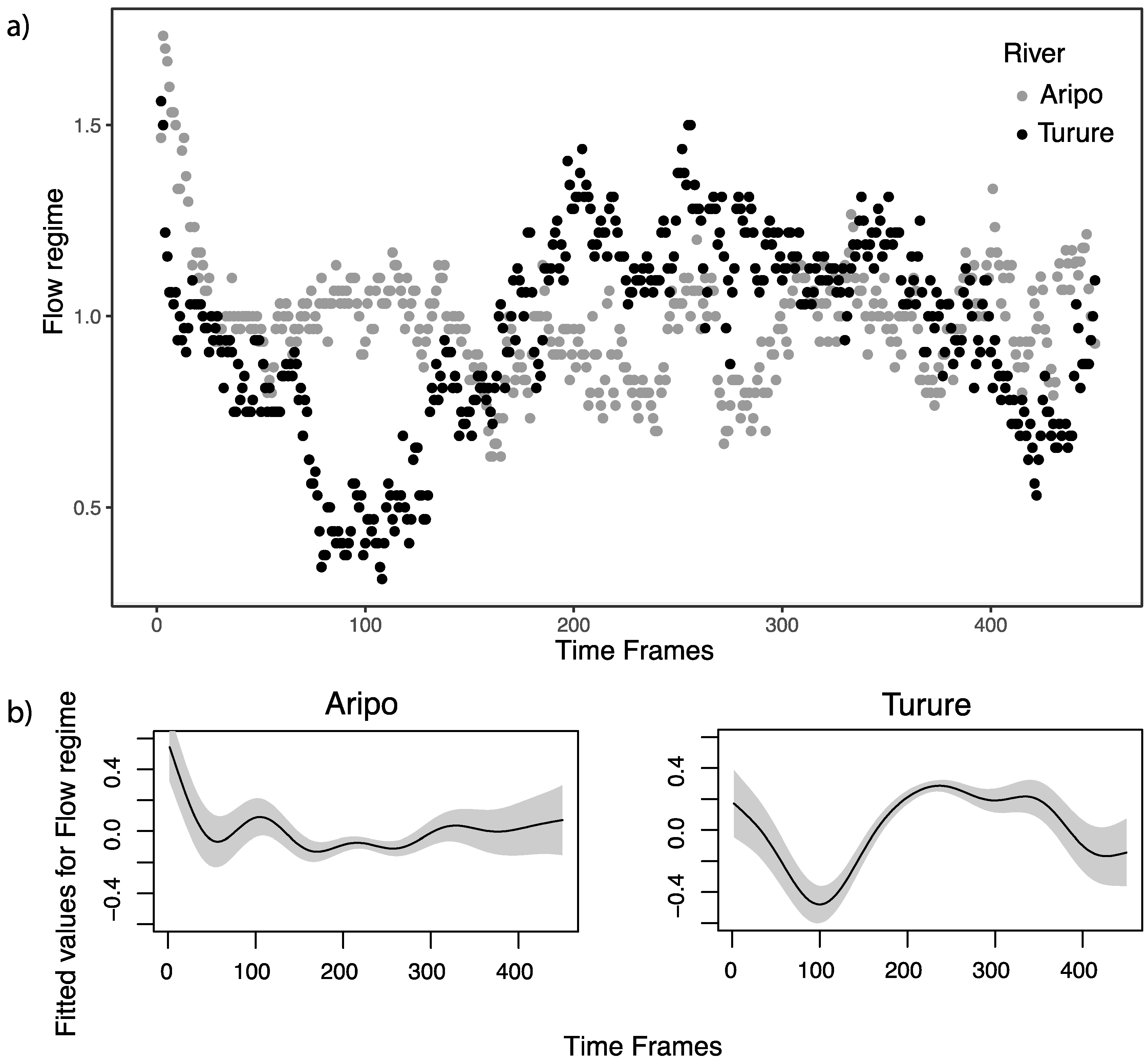
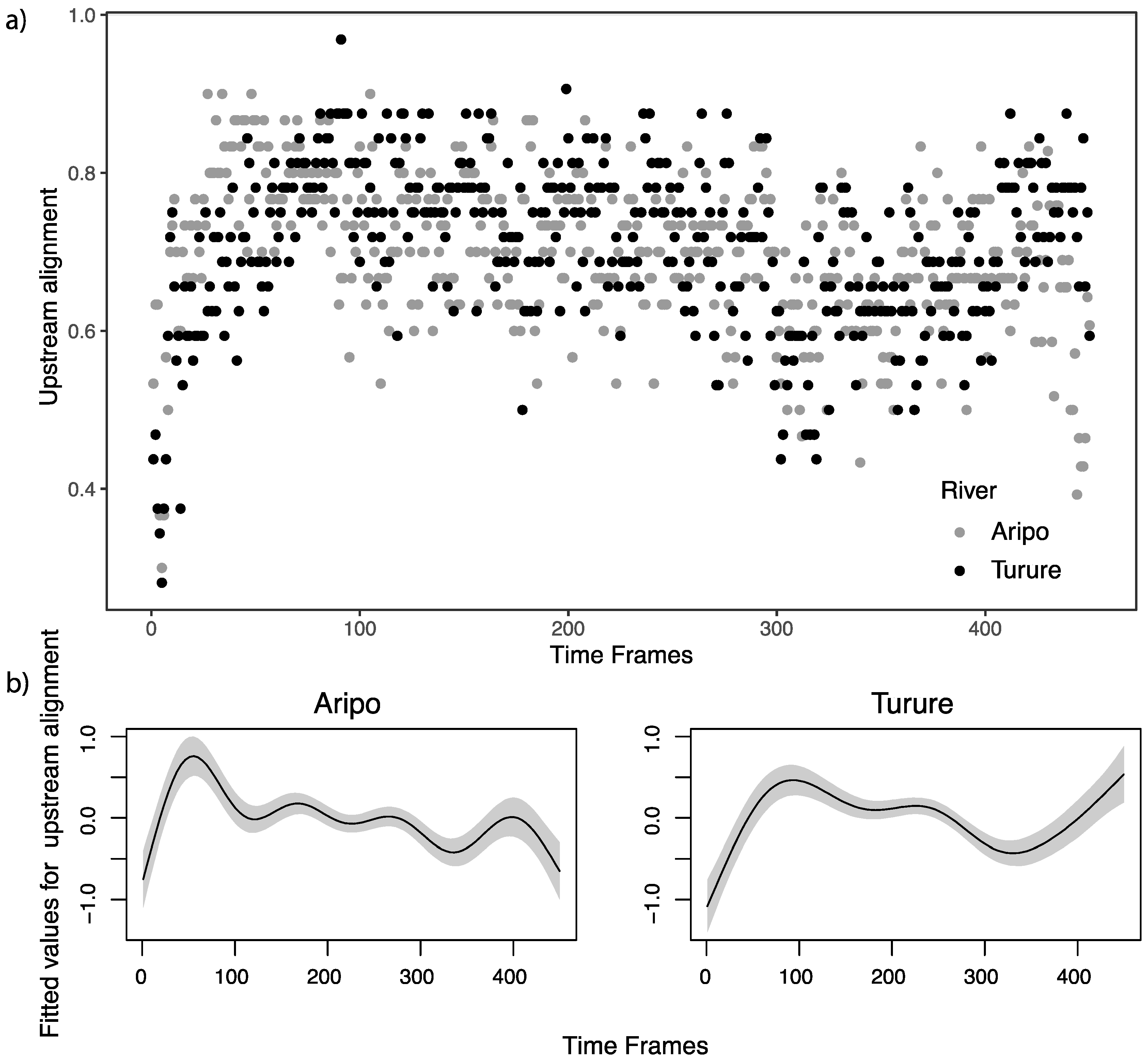
3.3. Generalized Additive Model for Temporal Patterns of Flow Regime and Alignment
4. Discussion
4.1. Objective 1. Rheotaxis in Two Asymmetrically Isolated Guppy Populations
4.2. Objective 2. Multiple Solutions to the Same Problem
4.3. Objective 3. Rheotaxis along A River Gradient
4.4. But Is It Evolution?
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mayr, E. The Challenge of Island Faunas. Aust. Nat. Hist. 1967, 15, 369–374. [Google Scholar]
- Hendry, A.P.; Kinnison, M.T. Perspective: The pace of modern life: Measuring rates of contemporary microevolution. Evolution 1999, 53, 1637–1653. [Google Scholar] [CrossRef] [PubMed]
- Reznick, D.N.; Ghalambor, C.K. The population ecology of contemporary adaptations: What empirical studies reveal about the conditions that promote adaptive evolution. Genetica 2001, 112–113, 183–198. [Google Scholar] [CrossRef]
- Lescak, E.A.; Bassham, S.L.; Catchen, J.; Gelmond, O.; Sherbick, M.L.; von Hippel, F.A.; Cresko, W.A. Evolution of stickleback in 50 years on earthquake-uplifted islands. Proc. Natl. Acad. Sci. USA 2015, 112, E7204–E7212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minteer, B.A.; Collins, J.P. Move it or lose it? The ecological ethics of relocating species under climate change. Ecol. Appl. 2010, 20, 1801–1804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carvalho, G.R.; Shaw, P.W.; Hauser, L.; Seghers, B.H.; Magurran, A.E. Artificial introductions, evolutionary change and population differentiation in Trinidadian guppies (Poecilia reticulata: Poeciliidae). Biol. J. Linn. Soc. 1996, 57, 219–234. [Google Scholar] [CrossRef]
- Losos, J.B.; Warheitt, K.I.; Schoener, T.W. Adaptive differentiation following experimental island colonization in Anolis lizards. Nature 1997, 387, 70–73. [Google Scholar] [CrossRef]
- Huey, R.B.; Gilchrist, G.W.; Hendry, A.P. Using Invasive Species to Study Evolution. In Species Invasions Insights into Ecology, Evolution, and Biogeography; Sinauer Associates: Sunderland, MA, USA, 2005; pp. 139–164. [Google Scholar]
- Ghalambor, C.K.; Hoke, K.L.; Ruell, E.W.; Fischer, E.K.; Reznick, D.N.; Hughes, K.A. Non-adaptive plasticity potentiates rapid adaptive evolution of gene expression in nature. Nature 2015, 525, 372–375. [Google Scholar] [CrossRef] [PubMed]
- Endler, J.A. Natural Selection on Color Patterns in Poecilia reticulata. Evolution 1980, 34, 76–91. [Google Scholar] [CrossRef]
- Reznick, D.N.; Ghalambor, C.K. Selection in nature: Experimental manipulations of natural populations. Integr. Comp. Biol. 2005, 45, 456–462. [Google Scholar] [CrossRef]
- Krueger, C.C.; May, B. Ecological and genetic effects of salmonid introductions in North America. Can. J. Fish. Aquat. Sci. 1991, 48, 66–77. [Google Scholar] [CrossRef]
- Adams, S.B.; Frissell, C.A.; Rieman, B.E. Geography of invasion in mountain streams: Consequences of headwater lake fish introductions. Ecosystems 2001, 4, 296–307. [Google Scholar] [CrossRef]
- Bolnick, D.I.; Caldera, E.J.; Matthews, B. Evidence for asymmetric migration load in a pair of ecologically divergent stickleback populations. Biol. J. Linn. Soc. 2008, 94, 273–287. [Google Scholar] [CrossRef] [Green Version]
- Fitzpatrick, S.W.; Gerberich, J.C.; Kronenberger, J.A.; Angeloni, L.M.; Funk, W.C. Locally adapted traits maintained in the face of high gene flow. Ecol. Lett. 2015, 18, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Fitzpatrick, S.W.; Handelsman, C.; Torres-Dowdall, J.; Ruell, E.; Broder, E.D.; Kronenberger, J.A.; Reznick, D.N.; Ghalambor, C.K.; Angeloni, L.M.; Funk, W.C. Gene flow constrains and facilitates genetically based divergence in quantitative traits. Copeia 2017, 105, 462–474. [Google Scholar] [CrossRef]
- Weese, D.J.; Schwartz, A.K.; Bentzen, P.; Hendry, A.P.; Kinnison, M.T. Eco-evolutionary effects on population recovery following catastrophic disturbance. Evol. Appl. 2011, 4, 354–366. [Google Scholar] [CrossRef]
- Arnold, G.P. Rheotropism in fishes. Biol. Rev. Camb. Philos. Soc. 1974, 49, 515–576. [Google Scholar] [CrossRef]
- Montgomery, J.C.; Baker, C.F.; Carton, A.G. The lateral line can mediate rheotaxis in fish. Nature 1997, 389, 960–963. [Google Scholar] [CrossRef]
- Northcutt, G.R. Swimming against the current. Nature 1997, 389, 915–916. [Google Scholar] [CrossRef]
- Northcote, T.G. Juvenile current response, growth and maturity of above and below waterfall stocks of rainbow trout, Salmo gairdneri. J. Fish Biol. 1981, 18, 741–751. [Google Scholar] [CrossRef]
- Northcote, T.G. Controls for trout and char migratory/resident behaviour mainly in stream systems above and below waterfalls/barriers: A multidecadal and broad geographical review. Ecol. Freshw. Fish 2010, 19, 487–509. [Google Scholar] [CrossRef]
- Jonsson, B. Diadromous and Resident Trout Salmo trutta: Is Their Difference Due to Genetics? Oikos 1982, 38, 297. [Google Scholar] [CrossRef]
- Morita, K.; Yamamoto, S. Contrasts in movement behavior of juvenile white-spotted charr between stocks above and below a dam. Fish. Sci. 2001, 67, 179–181. [Google Scholar] [CrossRef]
- Thompson, C.J.; Ahmed, N.I.; Veen, T.; Peichel, C.L.; Hendry, A.P.; Bolnick, D.I.; Stuart, Y.E. Many-to-one form-to-function mapping weakens parallel morphological evolution. Evolution 2017, 71, 2738–2749. [Google Scholar] [CrossRef] [PubMed]
- Hairston, N.G.; Ellner, S.P.; Geber, M.A.; Yoshida, T.; Fox, J.A. Rapid evolution and the convergence of ecological and evolutionary time. Ecol. Lett. 2005, 8, 1114–1127. [Google Scholar] [CrossRef]
- Thompson, J.N. Rapid evolution as an ecological process. Trends Ecol. Evol. 1998, 13, 329–332. [Google Scholar] [CrossRef]
- Reznick, D.N. Evaluation of the Rate of Evolution in Natural Populations of Guppies (Poecilia reticulata). Science 1997, 275, 1934–1937. [Google Scholar] [CrossRef] [Green Version]
- Magurran, A.E. Evolutionary Ecology: The Trinidadian Guppy; Oxford University Press: Oxford, UK, 2005; Volume 19, ISBN 0198527861. [Google Scholar]
- Shaw, P.W.; Carvalho, G.R.; Magurran, A.E.; Seghers, B.H. Population differentiation in Trinidadian guppies (Poecilia reticulata): Patterns and problems. J. Fish Biol. 1991, 39, 203–209. [Google Scholar] [CrossRef]
- Shaw, P.W.; Carvalho, G.R.; Seghers, B.H.; Magurran, A.E. Genetic consequences of an artificial introduction of guppies (Poecilia reticulata) in N. Trinidad. Proc. R. Soc. B Biol. Sci. 1992, 248, 111–116. [Google Scholar]
- Magurran, A.E.; Seghers, B.H.; Carvalho, G.R.; Shaw, P.W. Behavioural consequences of an artificial introduction of guppies (Poecilia reticulata) in N. Trinidad: Evidence for the evolution of anti-predator behaviour in the wild. Proc. R. Soc. B Biol. Sci. 1992, 248, 117–122. [Google Scholar]
- Becher, S.A.; Magurran, A.E. Gene flow in Trinidadian guppies. J. Fish Biol. 2000, 56, 241–249. [Google Scholar] [CrossRef]
- Carvalho, G.R.; Shaw, P.W.; Magurran, A.E.; Seghers, B.H. Marked genetic divergence revealed by allozymes among populations of the guppy Poecilia reticulata (Poeciliidae), in Trinidad. Biol. J. Linn. Soc. 1991, 42, 389–405. [Google Scholar] [CrossRef]
- Fajen, A.; Breden, F. Mitochondrial DNA Sequence Variation among Natural Populations of the Trinidad Guppy, Poecilia reticulata. Evolution 1992, 46, 1457–1465. [Google Scholar] [CrossRef] [PubMed]
- Reznick, D.; Butler Iv, M.J.; Rodd, H. Life-history evolution in guppies. VII. The comparative ecology of high- and low-predation environments. Am. Nat. 2001, 157, 126–140. [Google Scholar] [CrossRef] [PubMed]
- Croft, D.P.; Albanese, B.; Arrowsmith, B.J.; Botham, M.; Webster, M.; Krause, J. Sex-biased movement in the guppy (Poecilia reticulata). Oecologia 2003, 137, 62–68. [Google Scholar] [CrossRef] [PubMed]
- Archard, G.; Cuthill, I.; Partridge, J.; van Oosterhout, C. Female guppies (Poecilia reticulata) show no preference for conspecific chemosensory cues in the field or an artificial flow chamber. Behaviour 2008, 145, 1329–1346. [Google Scholar]
- Rasband, W. ImageJ; National Institutes of Health: Bethesda, MD, USA, 2012.
- Meijering, E. MTrackJ: A Java Program for Manual Object Tracking; University Medical Center Rotterdam: Rotterdam, The Netherlands, 2006. [Google Scholar]
- Jiang, Y.; Torrance, L.; Peichel, C.L.; Bolnick, D.I. Differences in rheotactic responses contribute to divergent habitat use between parapatric lake and stream threespine stickleback. Evolution 2015, 69, 2517–2524. [Google Scholar] [CrossRef] [PubMed]
- RStudio Team. RStudio: Integrated Development for R 2016; RStudio, Inc.: Boston, MA, USA, 2016. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2018. [Google Scholar]
- Chatham, K. Planned Contrasts: An Overview of Comparison Methods. In Proceedings of the Annual Meeting of the SouthwestEducational Research Association, San Antonio, TX, USA, 21–23 January 1999; pp. 1–19. [Google Scholar]
- Wood, S.; Scheipl, F. gamm4: Generalized Additive Mixed Models Using “Mgcv” and “Lme4” 2017; R Package: Bristol, UK, 2017. [Google Scholar]
- Wood, S.N. Fast stable restricted maximum likelihood and marginal likelihood estimation of semiparametric generalized linear models. J. R. Stat. Soc. Ser. B Stat. Methodol. 2011, 73, 3–36. [Google Scholar] [CrossRef] [Green Version]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; R Core Team. nlme: Linear and Nonlinear Mixed Effects Models 2015; R Package: Madison, WI, USA, 2015. [Google Scholar]
- Mohammed, R.; Oosterhout, C.; Van Schelkle, B.; Cable, J.; McMullan, M. Upstream guppies (Poecilia reticulata, Peters, 1859) go against the flow. Biota Neotrop. 2012, 12, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Raleigh, R.F. Genetic Control in the Lakeward Migrations of Sockeye Salmon (Oncorhynchus nerka) Fry. J. Fish. Res. Board Canada 1967, 24, 2613–2622. [Google Scholar] [CrossRef]
- Jiang, Y.; Peichel, C.L.; Torrance, L.; Rizvi, Z.; Thompson, S.; Palivela, V.V.; Pham, H.; Ling, F.; Bolnick, D.I. Sensory trait variation contributes to biased dispersal of threespine stickleback in flowing water. J. Evol. Biol. 2017, 30, 681–695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fitzpatrick, S.W.; Torres-Dowdall, J.; Reznick, D.N.; Ghalambor, C.K.; Funk, W.C. Parallelism isn’t perfect: Could disease and flooding drive a life-history anomaly in Trinidadian guppies? Am. Nat. 2014, 183, 290–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwartz, A.K.; Hendry, A.P. A test for the parallel co-evolution of male colour and female preference in Trinidadian guppies (Poecilia reticulata). Evol. Ecol. Res. 2007, 9, 71–90. [Google Scholar]
- Millar, N.P.; Hendry, A.P. Population divergence of private and non-private signals in wild guppies. Environ. Biol. Fishes 2012, 94, 513–525. [Google Scholar] [CrossRef]
- Bolnick, D.I.; Barrett, R.D.H.; Oke, K.B.; Rennison, D.J.; Stuart, Y.E. (Non)Parallel Evolution. Annu. Rev. Ecol. Evol. Syst. 2018, 49, 303–330. [Google Scholar] [CrossRef]
- Crispo, E.; Bentzen, P.; Reznick, D.N.; Kinnison, M.T.; Hendry, A.P. The relative influence of natural selection and geography on gene flow in guppies. Mol. Ecol. 2006, 15, 49–62. [Google Scholar] [CrossRef]
- Blondel, L.; Baillie, L.; Quinton, J.; Alemu, J.B.; Paterson, I.; Hendry, A.P.; Bentzen, P. Evidence for contemporary and historical gene flow between guppy populations in different watersheds, with a test for associations with adaptive traits. Ecol. Evol. 2019, 9, 4504–4517. [Google Scholar] [CrossRef]
- Montgomery, J.; Coombs, S.; Halstead, M. Biology of the mechanosensory lateral line in fishes. Rev. Fish Biol. Fish. 1995, 5, 399–416. [Google Scholar] [CrossRef]
- Fischer, E.K.; Soares, D.; Archer, K.R.; Ghalambor, C.K.; Hoke, K.L. Genetically and environmentally mediated divergence in lateral line morphology in the Trinidadian guppy (Poecilia reticulata). J. Exp. Biol. 2013, 216, 3132–3142. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site Locations Relative to Waterfalls | Aripo | Turure | ||
|---|---|---|---|---|
| F0 | F1 | F0 | F1 | |
| Above | 11 | 2 | 14 | 2 |
| Between | 5 | 5 | 6 | 3 |
| Below | 6 | 0 | 5 | 2 |
| Total | 22 | 7 | 25 | 7 |
| Response Variable | Adj. R2 | F | P |
|---|---|---|---|
| Net displacement | 0.00 | ||
| Mean temperature | 0.771 | 0.384 | |
| River | 0.882 | 0.352 | |
| Pool | 0.152 | 0.860 | |
| River × Pool | 1.214 | 0.305 | |
| Log of cumulative upstream movement | 0.28 | ||
| Mass | 0.992 | 0.324 | |
| Mean temperature | 2.028 | 0.160 | |
| Generation | 6.004 | 0.018 | |
| River | 1.885 | 0.176 | |
| Pool | 0.728 | 0.488 | |
| River × Pool | 1.268 | 0.290 | |
| Log of flow regime | 0.54 | ||
| Mean temperature | 0.926 | 0.340 | |
| Generation | 32.020 | 6.29 × 10−7 | |
| River | 1.250 | 0.269 | |
| Pool | 0.849 | 0.433 | |
| River × Pool | 1.364 | 0.265 | |
| Upstream orientation | −0.07 | ||
| River | 0.017 | 0.900 | |
| Pool | 0.293 | 0.748 | |
| River × Pool | 0.169 | 0.845 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blondel, L.; Klemet-N’Guessan, S.; Scott, M.E.; Hendry, A.P. Asymmetric Isolation and the Evolution of Behaviors Influencing Dispersal: Rheotaxis of Guppies above Waterfalls. Genes 2020, 11, 180. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11020180
Blondel L, Klemet-N’Guessan S, Scott ME, Hendry AP. Asymmetric Isolation and the Evolution of Behaviors Influencing Dispersal: Rheotaxis of Guppies above Waterfalls. Genes. 2020; 11(2):180. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11020180
Chicago/Turabian StyleBlondel, Léa, Sandra Klemet-N’Guessan, Marilyn E. Scott, and Andrew P. Hendry. 2020. "Asymmetric Isolation and the Evolution of Behaviors Influencing Dispersal: Rheotaxis of Guppies above Waterfalls" Genes 11, no. 2: 180. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11020180