Evolution of the Small Family of Alternative Splicing Modulators Nuclear Speckle RNA-Binding Proteins in Plants

 , and
, and 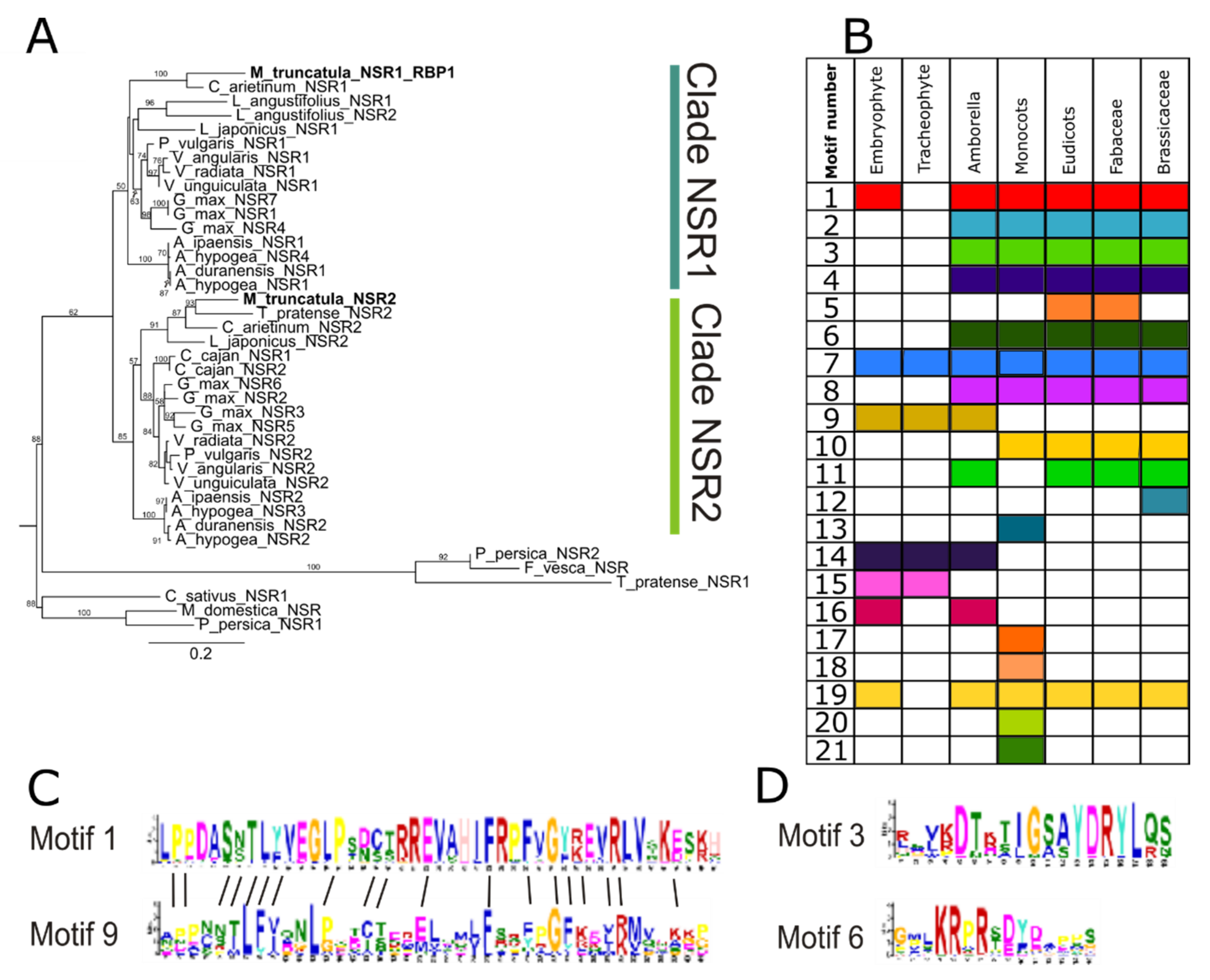{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification of NSR in the Plant Kingdom, Phylogenetic Analysis, and Conserved Protein Motif Characterization
2.2. RNA Expression Analysis of NSR Genes in M. Truncatula and L. Japonicus Nodules
3. Results
3.1. The NSR Protein Family Predates the Origin of Vascular Plants
3.2. Changes in Protein Domain Composition Suggest Distinct Roles for NSRs in Different Plant Lineages
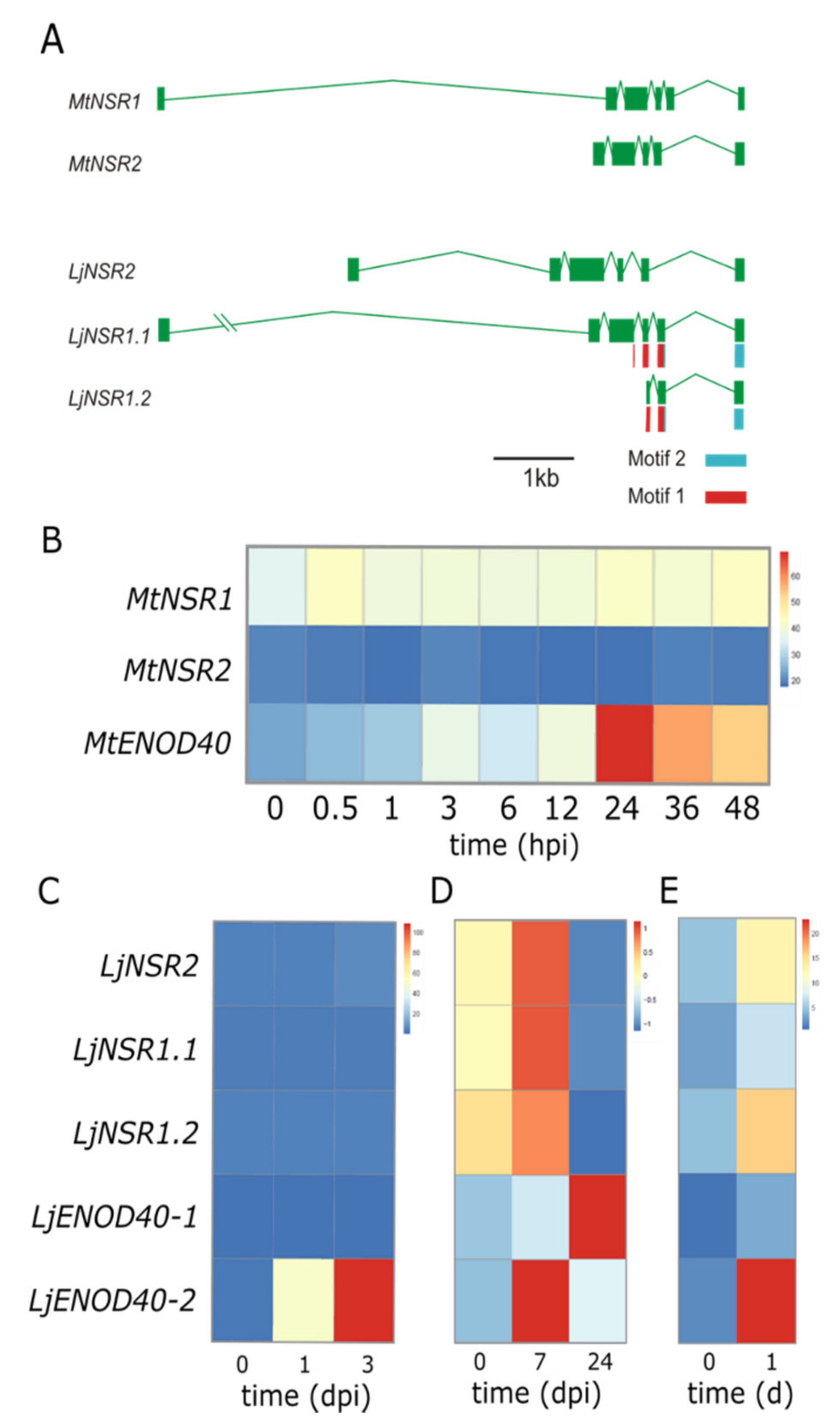
3.3. Alternatively Spliced Variants of NSR Genes May Compensate Lack of Gene Family Expansion
3.4. Transcriptional Behavior of NSR Genes in Legumes Hints at Distinct Roles During Nodulation
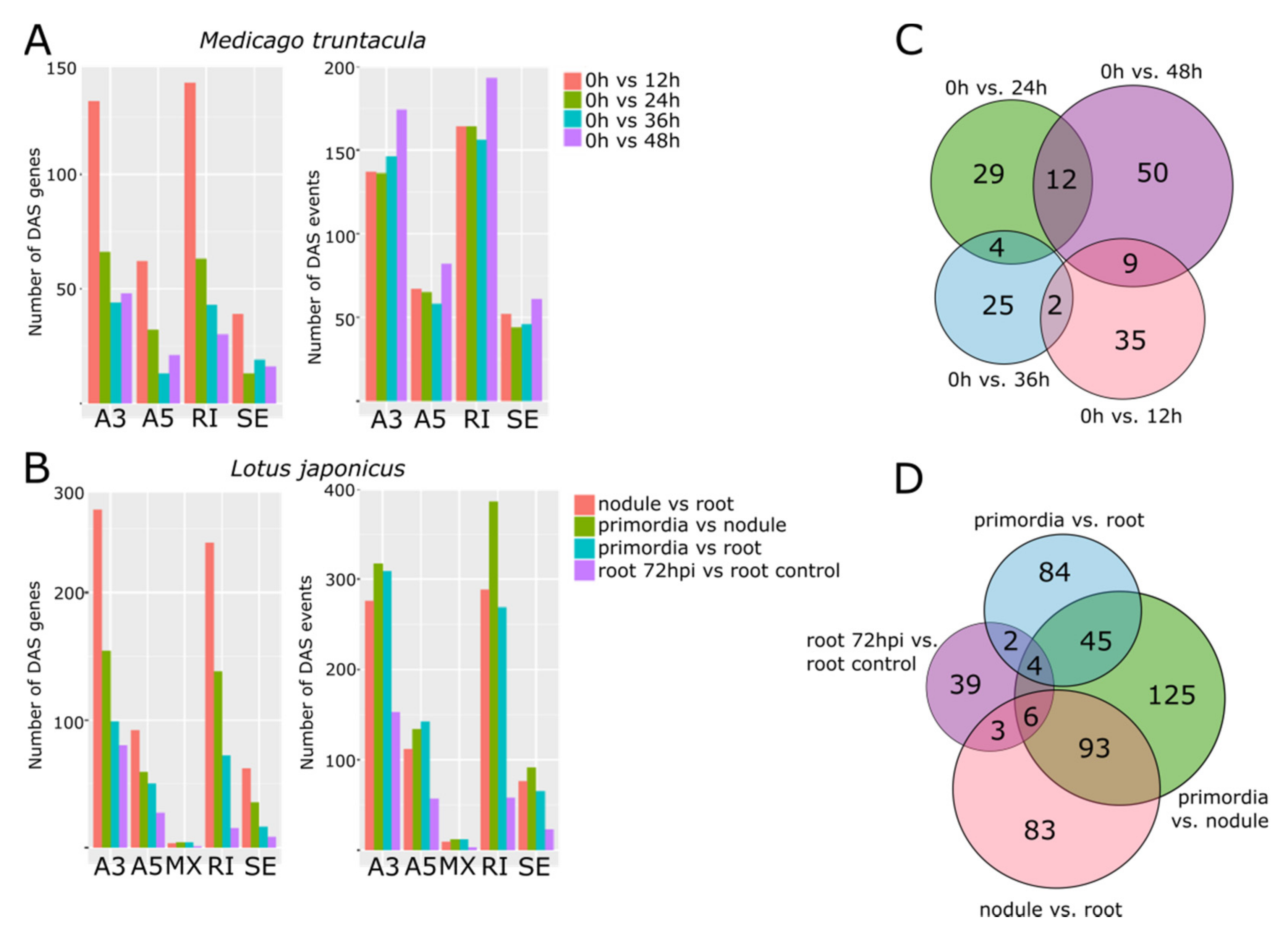
3.5. Distinctive Populations of Alternatively Spliced mRNAs Characterize the Progression of Nodule Development
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Sorenson, R.; Juntawong, P. Getting the message across: Cytoplasmic ribonucleoprotein complexes. Trends Plant. Sci. 2009, 14, 443–453. [Google Scholar]
- Marondedze, C.; Thomas, L.; Serrano, N.L.; Lilley, K.S.; Gehring, C. The RNA-binding protein repertoire of Arabidopsis thaliana. Sci. Rep. 2016, 11, 29766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bardou, F.; Ariel, F.; Simpson, C.G.; Romero-Barrios, N.; Laporte, P.; Balzergue, S.; Brown, J.W.S.; Crespi, M. Long Noncoding RNA Modulates Alternative Splicing Regulators in Arabidopsis. Dev. Cell 2014, 30, 166–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romero-Barrios, N.; Legascue, M.F.; Benhamed, M.; Ariel, F.; Crespi, M. Splicing regulation by long noncoding RNAs. Nucl. Acids Res. 2018, 16, 2169–2184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bazin, J.; Romero, N.; Rigo, R.; Charon, C.; Blein, T.; Ariel, F.; Crespi, M. Nuclear Speckle RNA Binding Proteins Remodel Alternative Splicing and the Non-coding Arabidopsis Transcriptome to Regulate a Cross-Talk Between Auxin and Immune Responses. Front. Plant. Sci. 2018, 9, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Crespi, M.; Jurkevitchl, E.; Poiret, M.; Aubenton-carafa, Y.; Petrovics, G.; Kondorosi, E.; Kondorosi, A. enod4O, a gene expressed during nodule organogenesis, codes for a non-translatable RNA involved in plant growth. EMBO J. 1994, 13, 5099–5112. [Google Scholar] [CrossRef]
- Charon, C.; Sousa, C.; Crespi, M.; Kondorosi, A. Alteration of enod40 expression modifies medicago truncatula root nodule development induced by sinorhizobium meliloti. Plant. Cell 1999, 11, 1953–1966. [Google Scholar]
- Campalans, A.; Kondorosi, A.; Crespi, M. Enod40, a Short Open Reading Frame—Containing mRNA, Induces Cytoplasmic Localization of a Nuclear RNA Binding Protein in Medicago truncatula. Plant. Cell 2004, 16, 1047–1059. [Google Scholar] [CrossRef] [Green Version]
- Goodstein, D.M.; Shu, S.; Howson, R.; Neupane, R.; Hayes, R.D.; Fazo, J.; Mitros, T.; Dirks, W.; Hellsten, U.; Putnam, N.; et al. Phytozome: A comparative platform for green plant genomics. Nucl. Acids Res. 2012, 40, 1178–1186. [Google Scholar] [CrossRef]
- Dash, S.; Campbell, J.D.; Cannon, E.K.; Cleary, A.M.; Huang, W.; Kalberer, S.R.; Karingula, V.; Rice, A.G.; Singh, J.; Umale, P.E.; et al. Legume information system (LegumeInfo.org): A key component of a set of federated data resources for the legume family. Nucl. Acids Res. 2016, 44, D1181–D1188. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C.; Drive, R.M.; Valley, M. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucl. Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mesquite: A Modular System for Evolutionary Analysis. Available online: https://www.mesquiteproject.org/ (accessed on 3 October 2019).
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Figtree Software v1.4.4. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 6 October 2019).
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME Suite: Tools for motif discovery and searching. Nucl. Acids Res. 2009, 37, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Roux, B.; Rodde, N. An integrated analysis of plant and bacterial gene expression in symbiotic root nodules using laser-capture microdissection coupled to RNA sequencing. Plant. J. 2014, 77, 817–837. [Google Scholar] [CrossRef] [PubMed]
- Larrainzar, E.; Riely, B.K.; Kim, S.C.; Carrasquilla-garcia, N.; Yu, H.; Hwang, H.; Oh, M.; Kim, G.B.; Surendrarao, A.K.; Chasman, D.; et al. Deep Sequencing of the Medicago truncatula Root Transcriptome Reveals a Massive and Early Interaction between Nodulation Factor and Ethylene Signals. Plant. Physiol. 2015, 169, 233–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelly, S.; Mun, T.; Stougaard, J.; Ben, C.; Andersen, S.U.; Murray, J.D. Distinct Lotus japonicus Transcriptomic Responses to a Spectrum of Bacteria Ranging From Symbiotic to Pathogenic. Front. Plant. Sci. 2018, 9, 1–14. [Google Scholar] [CrossRef]
- European Nucleotide Archive. Available online: https://www.ebi.ac.uk/ena (accessed on 20 September 2019).
- Bray, N.L.; Pimentel, H.; Melsted, P.; Pachter, L. Near-optimal probabilistic RNA-seq quantification. Nat Biotechnol. 2016, 34, 525–527. [Google Scholar] [CrossRef]
- Pecrix, Y.; Staton, S.E.; Sallet, E.; Lelandais-Brière, C.; Moreau, S.; Carrère, S.; Blein, T.; Jardinaud, M.-F.; Latrasse, D.; Zouine, M.; et al. Whole-genome landscape of Medicago truncatula symbiotic genes. Nat. Plants 2018, 4, 1017–1025. [Google Scholar] [CrossRef]
- Lotus Japonicus Genome Assembly Build 3.0. Available online: http://www.kazusa.or.jp/lotus/summary3.0.html (accessed on 3 February 2020).
- Alamancos, G.P.; Pagès, A.; Trincado, J.L.; Bellora, N.; Eyras, E. Leveraging transcript quantification for fast computation of alternative splicing profiles. RNA 2015, 21, 1521–1531. [Google Scholar] [CrossRef] [Green Version]
- Felsenstein, J. Cases in which Parsimony or Compatibility Methods Will be Positively Misleading Cases In Which Parsimony Or Compatibility Methods Will Be Positively Misleading1. Soc. Syst. Biol. 1978, 27, 401–410. [Google Scholar] [CrossRef] [Green Version]
- Oldroyd, G.E.D. Speak, friend and enter: Signalling systems that promote beneficial symbiotic associations in plants. Nat. Publ. Gr. 2013, 11, 252–263. [Google Scholar] [CrossRef] [PubMed]
- Peal, L.; Jambunathan, N.; Mahalingam, R. Phylogenetic and Expression Analysis of RNA-binding Proteins with Triple RNA Recognition Motifs in Plants. Mol. and Cells 2011, 31, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Morris, J.L.; Puttick, M.N.; Clark, J.W.; Edwards, D.; Kenrick, P.; Pressel, S.; Wellman, C.H.; Yang, Z.; Schneider, H.; Donoghue, P.C.J. The timescale of early land plant evolution. Proc. Natl. Acad. Sci. USA 2018, 115, 2274–2283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panchy, N.; Lehti-shiu, M.; Shiu, S. Evolution of Gene Duplication in Plants. Plant. Physiol. 2016, 171, 2294–2316. [Google Scholar] [CrossRef] [Green Version]
- Seijo, G.; Fernandez, A. Physical mapping of the 5S and 18S–25S rRNA genes by FISH as evidence that. Am. J. Bot. 2004, 91, 1294–1303. [Google Scholar] [CrossRef]
- Schmutz, J.; Cannon, S.B.; Schlueter, J.; Ma, J.; Mitros, T.; Nelson, W.; Hyten, D.L.; Song, Q.; Thelen, J.J.; Cheng, J.; et al. Genome sequence of the palaeopolyploid soybean. Nature 2010, 463, 178–183. [Google Scholar] [CrossRef] [Green Version]
- Gultyaev, A.P.; Roussis, A. Identification of conserved secondary structures and expansion segments in enod40 RNAs reveals new enod40 homologues in plants. Nucl. Acids Res. 2007, 35, 3144–3152. [Google Scholar] [CrossRef]
- Griesmann, M.; Chang, Y.; Liu, X.; Song, Y.; Haberer, G.; Crook, M.B.; Billault-penneteau, B.; Lauressergues, D.; Keller, J.; Imanishi, L.; et al. Phylogenomics reveals multiple losses of nitrogen-fixing root nodule symbiosis. Science 2018, 144, 1–11. [Google Scholar] [CrossRef] [Green Version]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lucero, L.; Bazin, J.; Rodriguez Melo, J.; Ibañez, F.; Crespi, M.D.; Ariel, F. Evolution of the Small Family of Alternative Splicing Modulators Nuclear Speckle RNA-Binding Proteins in Plants. Genes 2020, 11, 207. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11020207
Lucero L, Bazin J, Rodriguez Melo J, Ibañez F, Crespi MD, Ariel F. Evolution of the Small Family of Alternative Splicing Modulators Nuclear Speckle RNA-Binding Proteins in Plants. Genes. 2020; 11(2):207. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11020207
Chicago/Turabian StyleLucero, Leandro, Jeremie Bazin, Johan Rodriguez Melo, Fernando Ibañez, Martín D. Crespi, and Federico Ariel. 2020. "Evolution of the Small Family of Alternative Splicing Modulators Nuclear Speckle RNA-Binding Proteins in Plants" Genes 11, no. 2: 207. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11020207