1. Introduction
Edelweiss (
Leontopodium nivale subsp.
Alpinum (Cass.) Greuter) in the family
Asteraceae is a wildflower that grows in rocky limestone places at high altitude, such as the Swiss Alps [
1,
2]. Due to the rarity of its short-lived white flowers, edelweiss represents beauty and purity related to the Alps and Carpathians. In addition, many countries, including Austria, Bulgaria, Romania, Slovenia, and Switzerland, regard edelweiss as a national symbol.
For a long time, edelweiss has been used as traditional medicine against abdominal aches, bronchitis, diarrhea, dysentery, and fever [
3,
4]. Recently, several studies have shown the efficacy of edelweiss extracts for anti-inflammation in mice and rats [
3] and human keratinocytes and endothelial cells [
4]. In addition, the root extracts of edelweiss contain constituents that enhance cholinergic neurotransmission, indicating its potential for antidementia agents [
5] and antioxidants, such as leontopodic acid A and 3,5-dicaffeoylquinic acid, which can be used as anti-aging agents [
6]. Moreover, the edelweiss extracts showed antibacterial activity against
Enterococcus faecium,
Escherichia coli,
Pseudomonas aeruginosa,
Staphylococcus aureus,
Streptococcus pneumoniae, and
Streptococcus pyogenes, suggesting their possible ethnomedicinal use for respiratory and abdominal disorders [
7].
Several previous studies have analyzed the compounds of edelweiss extracts. For instance, the capillary chromatography method revealed 12 pharmacologically-important compounds, including flavonoids, caffeic acids, and leontopodic acid, from the aerial parts of edelweiss [
8]. To extract antioxidants from edelweiss plants, centrifugal partition chromatography (CPC) and high-performance liquid chromatography (HPLC) methods have been developed [
6]. It is known that the active compounds of edelweiss extracts between the aerial parts and roots are diverse [
3]. For instance, the hairy roots of edelweiss produce pharmacologically-active lignans, such as leoligin and 5-methoxy-leoligin, which can be stimulated by several other components, such as silver nitrate, sucrose, methyl jasmonate, and yeast extract [
9]. Furthermore, the metabolic patterns of 11 different
Leontopodium species have been revealed by nuclear magnetic resonance (1H NMR) spectroscopy and liquid chromatography–mass spectrometry (LC–MS) in their taxonomy relationship [
10].
Plant callus can be defined as unorganized or undifferentiated cell masses, which are easily induced by wounding to cover a plant wound or artificially conducting an in vitro system. Plant callus is artificially cultured by adding nutrients and plant growth regulators in antiseptic growth conditions. Currently, it is possible to produce a high number of specific plant cells with equal quality in a bioreactor. Moreover, a plant cell has an ability referred to as totipotency, which is the genetic potential of a plant cell to produce the entire mature plant [
11]. Thus, it is possible to generate a mature plant from the specific plant callus. Furthermore, callus can be widely used for plant regeneration to conserve rare and endangered plants [
12,
13].
Like animal cells, plant cells have abilities to facilitate the stimulation and regeneration of plants after injury [
14]. Although plant stem cells are regarded as emerging materials in the cosmetic industry, the available materials are limited. It is important to identify new plant sources and evaluate their functional components associated with cosmetics.
Edelweiss extracts are well known for their use in pharmaceutical agents; however, the effects of edelweiss extract as a natural cosmetic source are rarely reported. Here, we investigated the efficacy of edelweiss callus extract (Leontopodium Alpinum callus culture extract; LACCE) derived from edelweiss leaves using multiple assays from in vitro to in vivo and revealed the molecular mechanism caused by LACCE using transcriptome profiling.
2. Materials and Methods
2.1. Production of Edelweiss Callus
Edelweiss seeds were commercially purchased. The edelweiss seeds were soaked in 70% ethanol for 30 s followed by washing with distilled water. Again, the seeds were shaken in 0.3% sodium hypochlorite (Waco, Osaka, Japan) for 20 min and washed with distilled water. The sterilized seeds were germinated on basic Murashige and Skoog (MS) medium (Duchefa Biochemie, Haarlem, The Netherlands). The edelweiss leaves in the aseptic condition were cut into small pieces (0.5 to 1 cm). We induced the early stage of plant cells on MS medium containing 0.5–3 mg/mL of 6-Benzylaminopurine (6-BAP) (Duchefa Biochemie) and 0.3–1 mg/mL of 2,4-Dichlorophenoxyacetic acid (2,4-D) (Duchefa Biochemie) in darkness at 25 ± 2 °C [
15]. The pH of the MS medium was adjusted to 5.8 using 1N of NaOH (Duchefa Biochemie). The induced callus was propagated in the petri dish. The selected callus line was cultured in a bio-reactor in the Anti-Aging Research Institute of BIO-FD&C Co., Ltd., Incheon, Korea. The cultured callus was harvested and washed three times with distilled water. The callus was dehydrated using the freeze drier (IlShinbioBase, Dongducheonsi, Korea) according to the manufacturer’s instructions. The dried edelweiss callus was stirred in distilled water at 50 °C for 8 h. Callus extracts were obtained by heat extraction at 98 °C for 10 min.
2.2. Culture of Human Skin Cells
The evaluated effects of edelweiss extracts on human skin cells, keratinocyte (HaCaT) cells, and normal human Detroit 551 fibroblast (ATCC, Manassas, VA, USA) were cultivated in Dulbecco’s Modified Eagle Medium (DMEM) (Welgene, Gyeongsan-si, Korea) supplemented with 10% fetal bovine serum (FBS) (Thermo Fisher Scientific, Waltham, MA, USA) and 1% antibiotic-antimycotic (Thermo Fisher Scientific, Waltham, MA, USA) at 37 °C with a 5% CO2 condition.
2.3. Assessment of Cell Metabolic Activity by MTT Assay
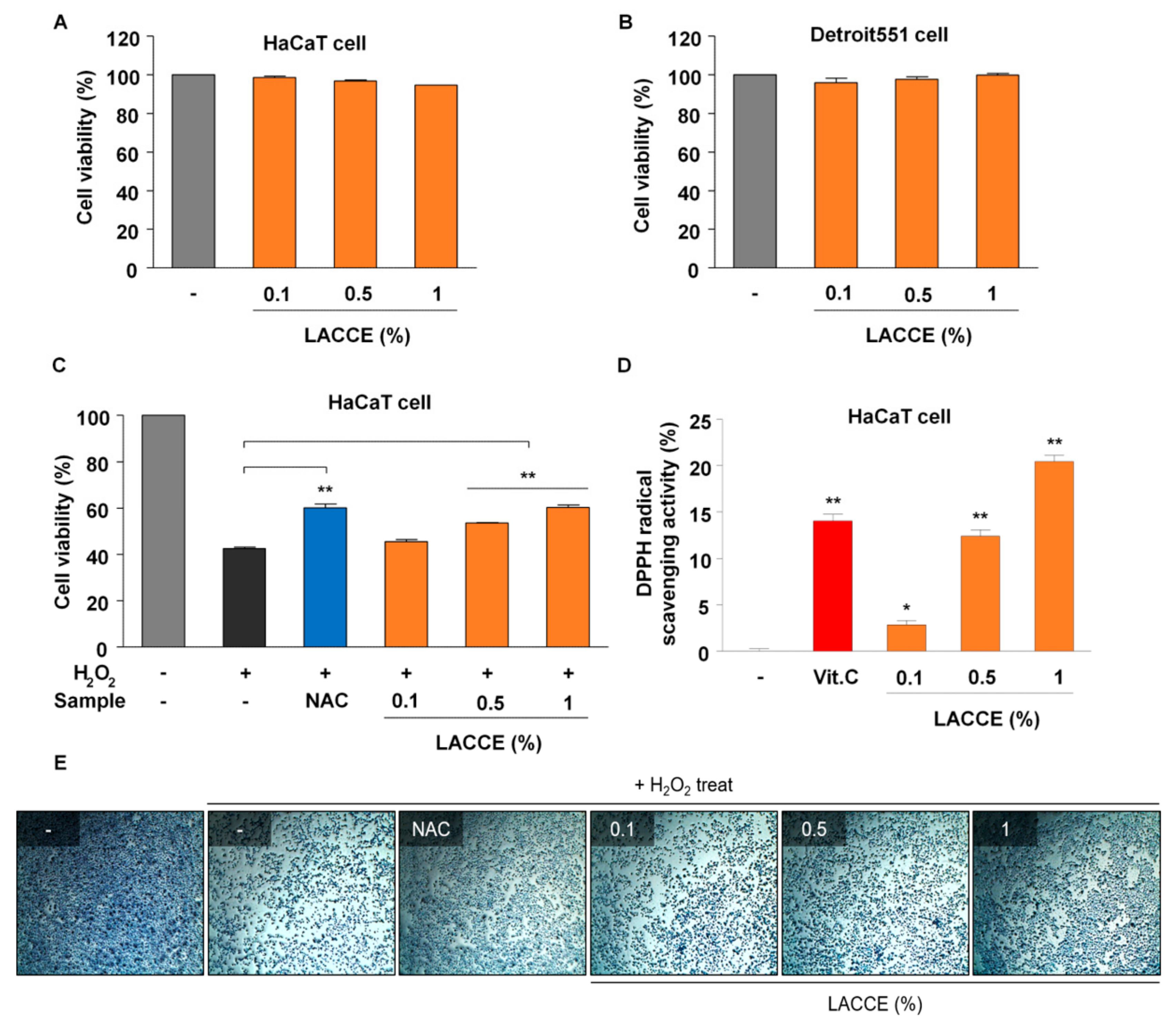
To assess the cytotoxicity of LACCE on the cellular growth, propagation, and survival of human skin cells, an MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide) assay was carried out as described previously [
16,
17]. In brief, HaCaT and Detroit 551 cells at a density of 5 × 10
4 cells per well, respectively, were incubated in a 96-well plate for 24 h. After that, final concentrations of 0.1%, 0.5%, and 1% LACCE were treated for 24 h. Distilled water was used as a control. After the treatment of LACCE, the medium was removed followed by the addition of 4 µL of 5 mg/mL MTT (Sigma-Aldrich, St. Louis, MO, USA) and incubated for 4 h. After removing the medium, 100 µL of dimethylsulfoxide (DMSO) (Sigma) was added and dissolved for 10 min. The wavelength absorbance was measured at 570 nm using a Thermo Scientific Multiskan GO Microplate Spectrophotometer (Fisher Scientific Ltd., Vantaa, Finland). Cell viability was obtained using the following formula. Cell viability (%) = (the amount of absorbance for treated cells/the amount of absorbance of control cells) × 100.
2.4. Assessment of Antioxidant Activity by DPPH Assay
The antioxidant activity of LACCE was measured by a DPPH (2,2-diphenyl-1-picryl-hydrazyl-hydrate) assay as described previously [
18]. In brief, 0.1 mL of final concentrations of 0.1%, 0.5%, and 1% LACCE was treated in 0.1 mL of 0.1 mM of DPPH (Sigma-Aldrich) in the presence of 0.4 mL of ethanol. We used 0.001% ascorbic acid (vitamin C) (Sigma-Aldrich) as a positive control. The samples were mixed well for 10 s and incubated at room temperature in dark conditions for 30 min. The wavelength absorbance was measured at 517 nm using a Thermo Scientific Multiskan GO Microplate Spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA). The radical scavenging activity was calculated using the following formula. Scavenging activity (%) = [1-(absorbance of test sample/absorbance of control)] × 100.
2.5. Assessment of Antioxidant Activity by Hydrogen Peroxide (H2O2) Assay
Hydrogen peroxide produces oxygen-free radicals in cells, resulting in cell death. We tested the cell mortality rate caused by hydrogen peroxide in LACCE-treated samples. HaCaT cells at a density of 5 × 105 cells per well were incubated in a 24-well plate for 24 h. After that, final concentrations of 0.1%, 0.5%, and 1% LACCE were treated in the presence of 1 mM of H2O2 for 8 h. As a positive control, 0.033% NAC (Sigma-Aldrich) was used. An MTT assay was used to measure the cell survival rate of LACCE-treated samples caused by H2O2. In addition, we observed cell morphology by methylene blue staining (Sigma-Aldrich, St. Louis, MO, USA).
2.6. Real-Time Reverse Transcription (RT)-PCR
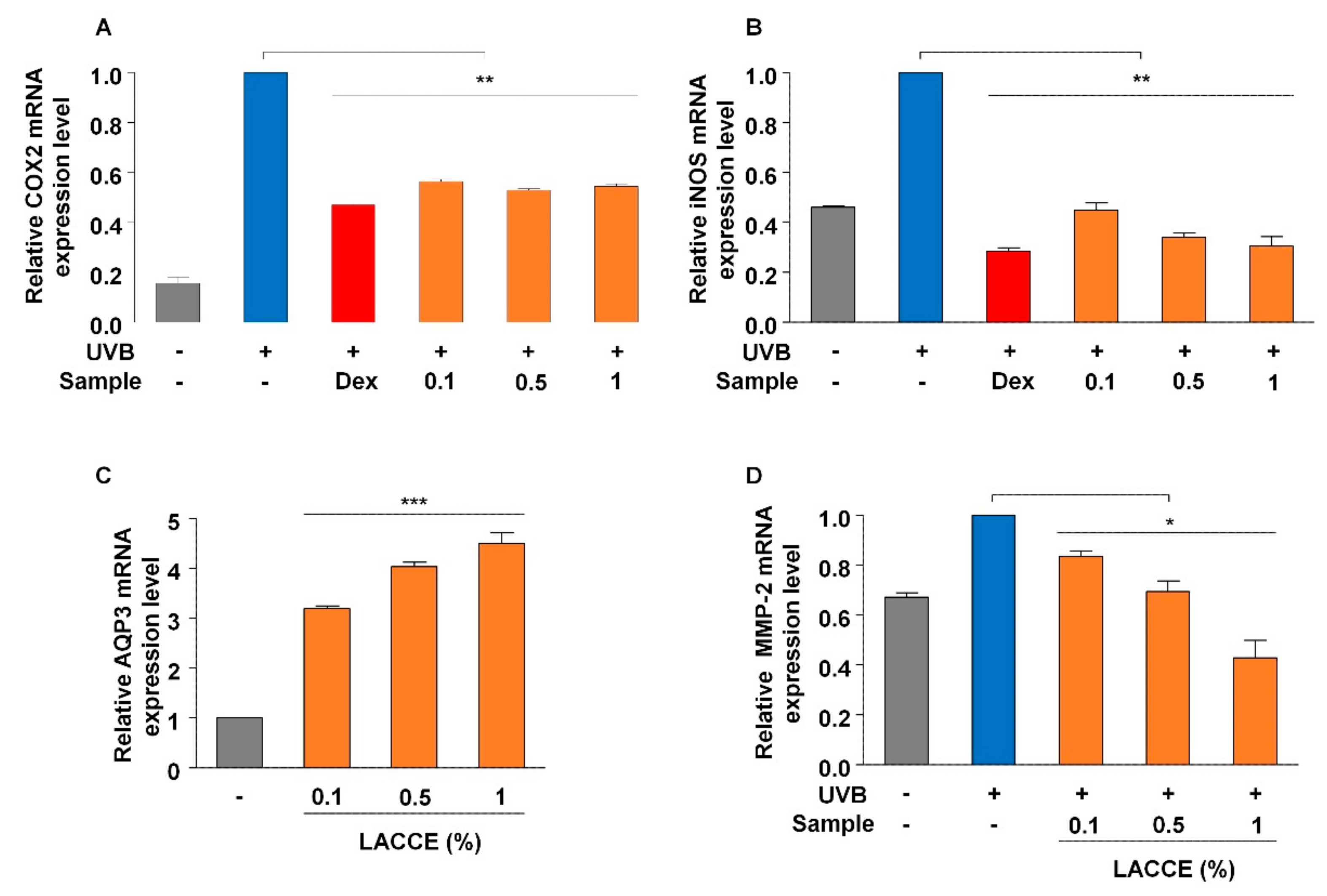
To examine the effect of LACCE on anti-wrinkles, moisturizing, and anti-inflammation, we carried out real-time RT-PCR with known primers amplifying marker genes using QuantiTect Primer Assays (Qiagen, Hilden, Germany) according to the manufacturer’s instructions. Detroit 551 cells at a density of 5 × 104 cells per well were incubated in a 96-well plate for 24 h. After that, final concentrations of 0.1%, 0.5%, and 1% LACCE was treated for 24 h. cDNA was synthesized from the respective Detroit 551 cells treated with different LACCE concentrations using a SuperPrep Cell Lysis & RT Kit for qPCR (Toyobo, Osaka, Japan) containing lysis reagents and RT reagents according to the manufacturer’s instructions. For the anti-wrinkle effect upon LACCE treatment, the expression of the gene encoding Matrix Metalloproteinase-2 (MMP-2) was quantified by real-time RT-PCR using a Thunderbird SYBR qPCR Mix kit (Toyobo) based on the manufacturer’s instructions. For the evaluation of moisturizing through LACCE treatment, the expression of the gene encoding Aquaporin 3 (AQP3) was examined by real-time RT-PCR. In the case of the anti-inflammatory assay, HaCaT cells at a density of 5 × 104 cells per well were incubated in a 96-well plate for 24 h. After removing the medium, UVB was irradiated on the HaCaT cells. For UVB irradiation, 5 mJ/cm2 was used by CL-1000 UV box (Analytik Jena AG, Jena, Germany). After that, final concentrations of 0.1%, 0.5%, and 1% LACCE were treated for 4 h. We used dexamethasone (Dex) (Sigma-Aldrich) as a positive control. For the anti-inflammatory effect from LACCE treatment, the expression of genes encoding COX2 and iNOS, respectively, were quantified by real-time RT-PCR. The expression of individual genes was normalized to GAPDH gene expression.
2.7. Clinical Evaluation of LACCE as an Agent for Cosmetics in Vivo
Clinical study for the evaluation of the efficacy of LACCE on facial lifting and improving periorbital wrinkle, skin elasticity, dermal density and skin thickness was conducted by Ellead company after the approval based on standard operating procedures (Ver. 3.0) (Ellead, Seongnam, Korea) of Ellead IRB in accordance with Korea Good Clinical Practices guideline (B1-2015-4-002) described by the Ministry of Food and Drug Safety (MFDS) and Ellead Standard Operating Procedures (EL-P-7400).
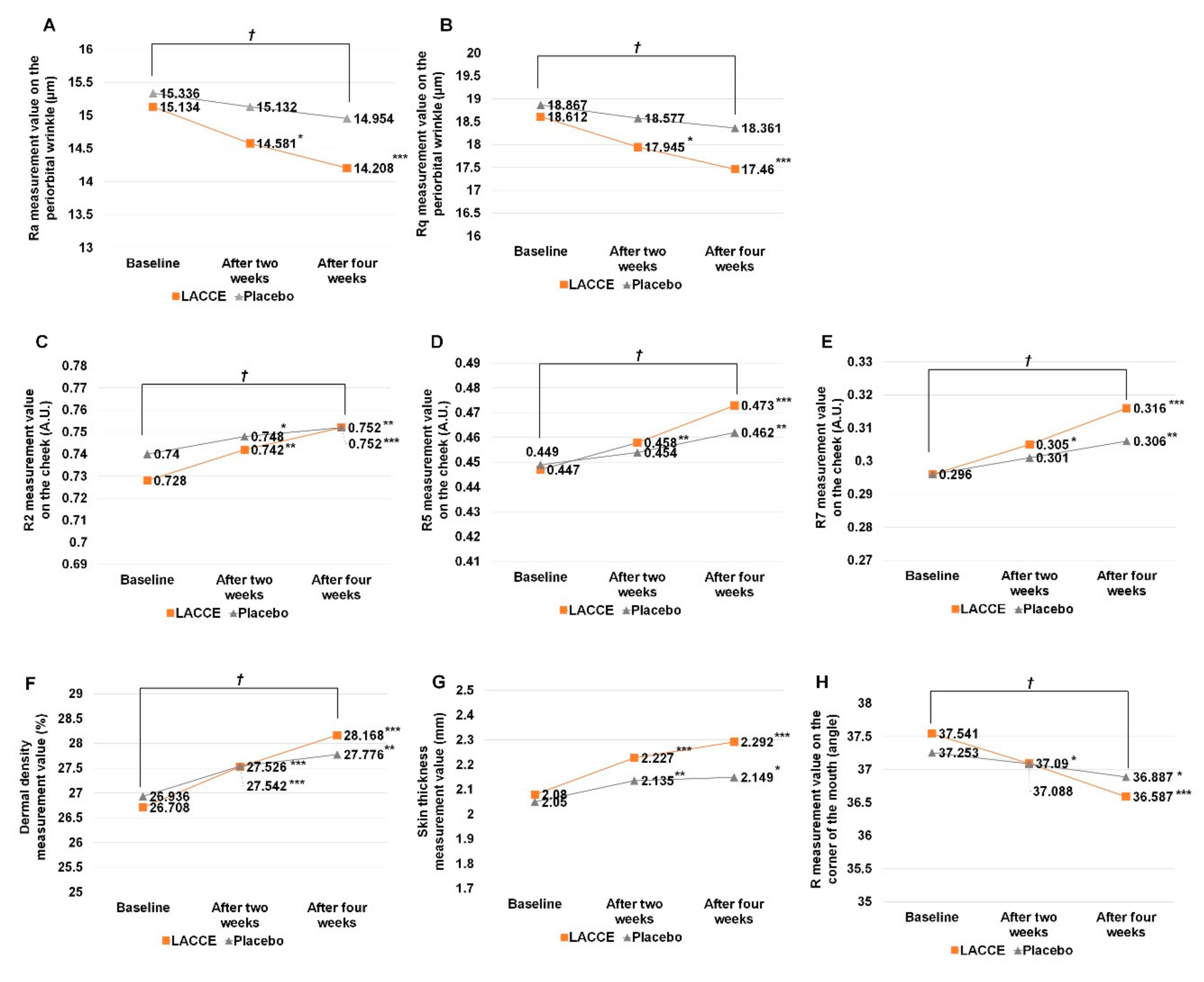
To examine the effect of LACCE in vivo, we carried out a clinical evaluation of LACCE for four different factors: facial lifting and improving periorbital wrinkles, skin elasticity, dermal density, and skin thickness. For this, a total of 21 female volunteers with average 48.04 ± 4.28 years old, who were further divided into 12 volunteers aged in their 40s and nine volunteers aged in their 50s, participated in the clinical evaluation over four weeks.
The clinical test was conducted at three different time points, the baseline, 2 weeks, and 4 weeks, for all volunteers. From a week before the beginning of the study until the end, all volunteers did not undergo additional treatments to improve skin, cosmetics, and sanitary aid, which could have affected the results. After washing the faces of the volunteers using the cleansing foam, the volunteers stood for at least 30 min in a controlled room with a constant temperature (20–24 °C) and humidity and (40%–60% relative humidity). We performed the measurement randomly on the left or right side of the faces of volunteers. Our test was a double-blind test in which neither the subjects nor the researchers knew which product was a test sample. Formulations to calculate the decreased and increased rates were described in
Table 1.
2.8. Measurement of Facial Lifting and Improving Periorbital Wrinkles
At the baseline, periorbital wrinkles, skin elasticity, dermal density, and skin thickness on the cheek, and facial lifting at the corner of the mouth were measured for all volunteers. After that, two different samples, LACCE and a placebo (control), were applied to the designated area of the face twice a day (morning and night). Volunteers were divided into two different groups, group I (LACCE was applied to the left facial area while the placebo was applied to the right facial area) and group II (the placebo was applied to the left facial area while LACCE was applied to the right facial area). Two and four weeks after treatment, the same measurements were conducted to evaluate the effect of LACCE compared with that of the placebo.
Periorbital wrinkles were analyzed using the optical 3D (dimension) skin measurement system PRIMOS High Resolution (Canfield Scientific, Parsippany, NJ, USA), which is ideal for the investigation of the skin microstructure and wrinkles. Two different roughness parameters, Ra (average roughness), which is the average of the absolute values of the profile heights of the roughness profile, and Rq (root mean square roughness), which is the root mean square average of the profile heights of the roughness profile, were measured.
2.9. Measurement of Skin Elasticity
Skin elasticity was analyzed using a Cutometer MPA 580 (Courage & Khazaka, Köln, Germany). A probe with a diameter of 2 mm was mounted on this equipment for the test. Mode 1 or the time-strain mode was used as the measurement condition. On-time 2.0 s and off-time 2.0 s were then applied to the specified negative pressure of the mode 1 condition (450 mbar), and measurements were taken three consecutive times. Three different parameter values, R2 (gross elasticity), R5 (net elasticity), and R7 (biological elasticity), were measured.
2.10. Measurement of Dermal Density and Skin Thickness
The dermal density and skin thickness on the cheek were analyzed using Dermascan-C (Cortex Technology, Hadsund, Denmark), a high-resolution imaging machine using 20-MHz supersonic waves, which is very useful in non-invasively observing changes to the inner layers of the skin. In our study, the B scanning image was used. First, we set the speed of supersonic waves to 20 MHz, spread the contact jelly for the test, placed the probe at a right angle to the skin, and pressed it slightly to measure the cheek area. The skin thickness and dermal density were calculated by the Dermascan-C software system.
The images of the face were photographed with identical photography conditions, such as the same light, high-resolution digital camera, and photographer. After that, facial lifting at the corner of the mouth was examined using Moiré analysis based on the images of the face. We selected the corner of the mouth, where skin sagging is prominent, as the test area. Using the images, the angle (R) between the horizontal line and the contour line drawn at the corner of the mouth was calculated using image analysis software (ImagePro Plus, Rockville, MD, USA).
2.11. Statistical Test
We conducted a one-way ANOVA test for the comparison between the control and test samples. The results were shown with mean and standard deviation (Mean ± SEM). The p-values p < 0.05 (*), p < 0.01 (**), and p < 0.001 were considered statistically significant. All statistical tests were declared statistically significant at the 0.05 level. We used IBM SPSS Statistics version 21.0 (SPSS, Chicago, IL, USA) for the statistical analysis.
2.12. Preparation of Libraries for RNA-Seq and Next-Generation Sequencing
For transcriptome analysis, HaCaT cells at a density of 1 × 106 cells per well were incubated in a six-well plate for 24 h. After that, human keratinocyte cells (treatment) were treated with a final concentration of 1% LACCE extract for 24 h while the mock condition (control) was treated with sterile water. For each condition, three different biological samples were harvested. Total RNA was extracted using an RNeasy Mini Kit (Qiagen, Hilden, Germany) according to the manufacturer’s instructions. After the extraction of total RNAs from each sample, six different libraries for RNA-Seq were prepared by the TruSeq Stranded mRNA LT Sample Prep Kit according to the manufacturer’s instructions. The six different libraries were paired-end sequenced by Illumina’s NovaSeq 6000 system (Macrogen, Seoul, Korea). The obtained raw sequence data were deposited in the National Center for Biotechnology Information (NCBI) Sequence Read Archive (SRA) database with the following respective accession numbers: SRR9127735, SRR9127736, SRR9127737, SRR9127738, SRR9127733, and SRR9127734.
2.13. Mapping, Normalization, and Identification of Differentially-Expressed Genes
We mapped the raw sequence reads from each library to human reference transcripts version GRCh38 (
https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/genome/guide/human/) using the BBMap aligner with default parameters (
https://sourceforge.net/projects/bbmap/). Using the bbmap.sh option, we calculated fragments per kilobase million (FPKM) values for each transcript. The obtained FPKM values from each condition were subjected to DEBrowser for the normalization and analysis of differentially-expressed genes (DEGs) [
19]. FPKM values of less than 1 were deleted. Using EdgeR with the TMM normalization method and exactTest type, we identified differentially-expressed genes according to adjusted
p-values of less than 0.001 and log
2 converted fold changes of more than one.
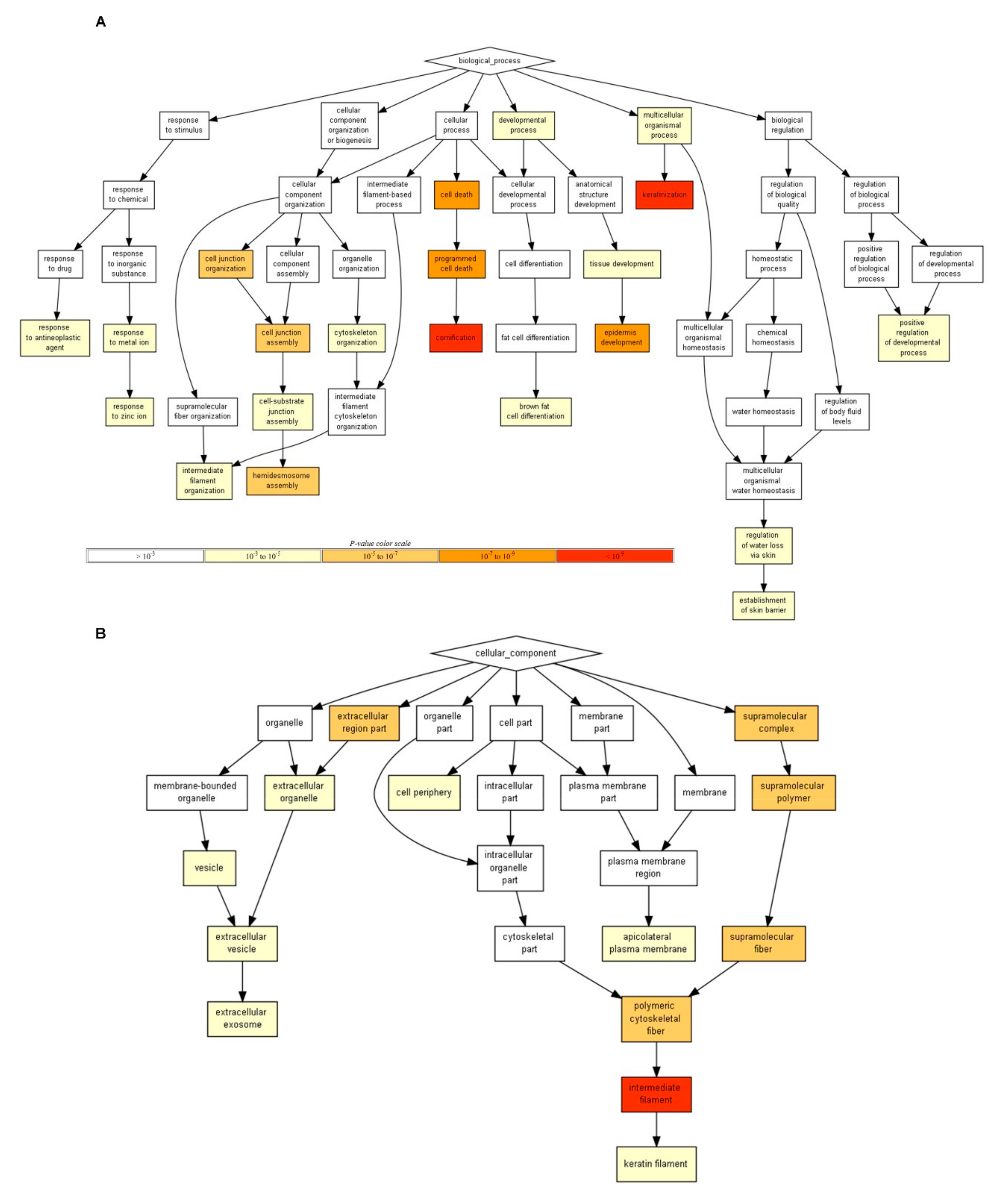
2.14. Gene Ontology (GO) Term Enrichment Analysis
For GO term enrichment analysis, we identified DEGs based on adjusted
p-values of less than 0.05 and log2 converted fold changes of more than 0.5. As a result, 22 up-regulated and 13 down-regulated genes were identified. Each gene set was subjected to GO enrichment analysis using the GORILLA program with default parameters (
http://cbl-gorilla.cs.technion.ac.il/) [
20]. We selected two unranked lists of genes as the running mode using 11,290 expressed genes as the background set.
4. Discussion
Cosmetics are classically defined as any prepared products that can be applied to the human body, including the face, skin, hair, mouth, and eyes, to change or strengthen the appearance of the human body [
21]. In addition, cosmetics are used for cleansing, providing fragrance, and giving protection. Most cosmetics are composed of chemical compounds that can be derived from natural sources or synthetics [
22].
In this study, we examined the possible effects of LACCE as a natural compound for cosmetics through diverse assays in vitro and in vivo. Several in vitro assay results showed the strong antioxidant activity of LACCE in response to UVB treatment. Interestingly, the effect of LACCE as an antioxidant was correlated with the concentration of LACCE. The antioxidant activity of LACCE (1%) was comparable to that of NAC or much higher than vitamin C. LACCE contains a higher amount of chlorogenic acid, 3,5-Dicaffeoylquinic acid, leontopodic acid B, and leontopodic acid A than normal edelweiss callus cultures, as shown in a previous study [
4]. In particular, a previous study identifying two leontopodic acids from edelweiss demonstrated the functional role of LACCE as an antioxidant agent [
23].
The expression of two inflammatory genes was suppressed by LACCE treatment, suggesting the possible role of LACCE in anti-inflammatory activity, as shown previously [
7,
24]. Interestingly, there was no significant difference in the anti-inflammatory effect among different LACCE concentrations, indicating that 1% LACCE might be sufficient for application as an anti-inflammatory agent in cosmetics. By contrast, the expressions of AQP3 and MMP2 required for moisturizing and wrinkling, respectively, were dramatically changed after treatment with different LACCE concentrations.
A clinical test is the most important step for using plant extract as a cosmetic source. In this study, we examined the in vivo effects of LACCE with 21 volunteers. The application of LACCE on face and skin tissues showed significant increases in four different factors (improvements in periorbital wrinkles, skin elasticity, dermal density, and skin thickness) compared with the placebo. In particular, the constant use of LACCE improved the face and skin tissues significantly. Although edelweiss extracts are known as a source for cosmetics, this is the first report demonstrating the successful application of LACCE as a cosmetic material.
To evaluate plant extracts as a cosmetic or medicinal source in vitro, the examination of the expression of marker genes is a popular experimental approach. However, the application of only selected genes for gene expression analysis has several limitations. The recent rapid development of next-generation sequencing facilitates gene expression analysis genome-wide. In this study, we used RNA-Seq to examine transcriptome-wide changes in human keratinocyte cells in response to LACCE. Our results showed that at least 16.56% of human genes were expressed in keratinocyte cells, indicating the tissue-specific expression of human genes. LACCE induced the expression of numerous genes; however, the global change in human transcriptome by LACCE was mild compared with other stress conditions. This result assures the safety of LACCE for application in human tissues.
GO enrichment analysis revealed that up-regulated genes encoding Keratin 5 (KRT5), KRT19, KRT6A, KRT15, KRT14, KRT17, and junction plakoglobin (JUP) were involved in keratinization and cornification. Epidermal keratinocytes play an important role as a barrier against diverse environmental factors [
25]. Specifically, terminal differentiated epidermal cells develop into dead keratinocytes by programmed cell death referred to as cornification, forming a strong epidermal barrier [
26]. After cell death, the cornified skin layer provides many advantages to the face and tissues, including an increase in elasticity, stability, moisturizing, and mechanical resistance [
25]. Of the identified KRT genes, the gene encoding Keratin15 (KRT15), a member of the keratin gene family, is known as a hair follicle stem cell marker that is highly expressed in skin tissues [
27,
28]. A recent study has suggested that KRT15 functions in epithelial regeneration against radiation and wound repair [
29,
30].
In addition to several genes encoding KRT proteins, genes encoding DDIT4, BNIP3, and IGFBP3 function in programmed cell death. For example, DNA damage-inducible transcript 4 (DDIT4) is highly expressed by different stresses and inhibits the mammalian target of rapamycin complex 1 (mTORC1) pathway associated with the treatment of cancer [
31]. Previous studies have identified the DDIT4 gene, which was highly expressed by dexamethasone, which is a chemotherapeutic agent inducing autophagy in lymphocytes, suggesting its possible role in the regulation of cell growth, proliferation, and survival [
32,
33]. LACCE showed similar effects to dexamethasone as an anti-inflammatory agent. Bcl-2/adenovirus E1B 19-kDa-interacting protein (BNIP3) is a member of a pro-apoptosis protein family regulating cellular proliferation [
34]. In addition, BNIP3 is involved in the protection of keratinocytes from UVB-induced apoptosis by up-regulating its gene expression [
35]. Several studies have shown that insulin-like growth-factor-binding protein 3 (IGFBP-3) expression is related to cellular senescence [
36,
37]. The addition of IGFBP-3 induces or inhibits apoptosis depending on cell types [
37]. For example, IGFBP-3 was up-regulated in the human papillomavirus-immortalized cervical cells, resulting in the enhancement of IGF-1-induced mitogenesis [
37]. A recent study has shown the down-regulation of IGFBP-3 in senescent and H
2O
2-induced old cells compared with young cells, suggesting its possible role as an aging marker [
36].
BNIP3, IGFBP3, stratifin (SFN), CA2, FGFBP1, KRT17, and JUP are involved in the positive regulation of the developmental process. Of them, carbonic anhydrases are ubiquitous enzymes present in prokaryotes and eukaryotes, including animals and plants [
38]. In humans, carbonic anhydrase catalyzes the formation of carbonic acid from water and carbon dioxide (CO
2), which are required for brain, kidney, and bone physiology [
39]. A previous study demonstrated that CA4 and CA9 were up-regulated in a mouse during the wound hypoxic period by RNA-Seq [
40]. In addition, the addition of recombinant CA9 enzyme promoted wound re-epithelialization [
40]. Furthermore, a proteome study also revealed the involvement of CA2 protein in aging and neurodegeneration in a mouse [
41]. Here, we demonstrated that LACCE promoted the regeneration of human keratinocyte cells by up-regulating CA2 expression. Fibroblast growth- factor-binding protein 1 (FGFBP1) is an extracellular secreted chaperone binding to FGFs and it modulates FGF signaling [
42]. Several studies have shown that FGFBP1 is highly expressed during angiogenesis and plays important roles in skin carcinogenesis, inflammation, and wound healing [
42,
43,
44]. Moreover, the cellular localization of up-regulated genes in the vesicle, extracellular exosome, intermediate filament, and keratin filament suggests that these proteins are a primary component of the cytoskeleton.
In down-regulated genes, most genes are known to be induced by different stresses. For instance, two genes encoding metallothionein 2a (MT2A) and metallothionein 1e (MT1E) are induced under metal stress conditions, including zinc ion, copper ion, and cadmium ion, and oxidative stress, such as UVB [
45,
46]. Moreover, an inhibitor of DNA-binding 3 (ID3), a member of helix–loop–helix proteins, is an immediate–early gene in response to mitogenic signals and oxidative stress [
47]. Ankyrin repeat domain 1 (ANKRD1), also known as cardiac ankyrin repeat protein (CARP), is a transcriptional cofactor that is up-regulated during wound healing and induces angiogenesis [
48]. Both ID3 and ANKRD1 are involved in the negative regulation of the biological process. ID3 acts as a transcriptional regulator inhibiting stem cell differentiation and promoting cell cycle progression [
47]. In addition, a recent study showed that the loss of ANKRD1 function in mice resulted in delayed wound healing, suggesting its role in wounding and tissue injury [
49]. Furthermore, Rho-related BTB domain-containing 3 (RHOBTB3) functions to promote the proteasomal degradation of hypoxia-inducible factors (HIFs), which are the main regulators of adaptive responses to low oxygen [
50]. In addition, interferon-induced transmembrane protein 1 (IFITM1) inhibits the entry of many viruses in the host cell [
51]. Similarly, the gene encoding interferon alpha-inducible protein 6 (IFI6) is induced by interferon and regulates apoptosis and antiviral innate immunity [
52,
53]. The down-regulation of stress-responsive genes by LACCE treatment suggests that LACCE does not cause stress in human keratinocyte cells.

 ,
, 






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}