The Genetic Basis of Natural Variation in Drosophila melanogaster Immune Defense against Enterococcus faecalis


Abstract
:1. Introduction
2. Materials and Methods
2.1. Drosophila and Bacterial Stocks
2.2. Genome-Wide Association Study
2.3. RNAi Knock-Down of Candidate Genes
2.4. E. faecalis Infection of RNAi Lines with Knock-Down of Candidate Genes
2.5. Effects of Knocking Down Specific Bomanin Genes on E. faecalis Survival
2.6. Statistical Analyses
3. Results
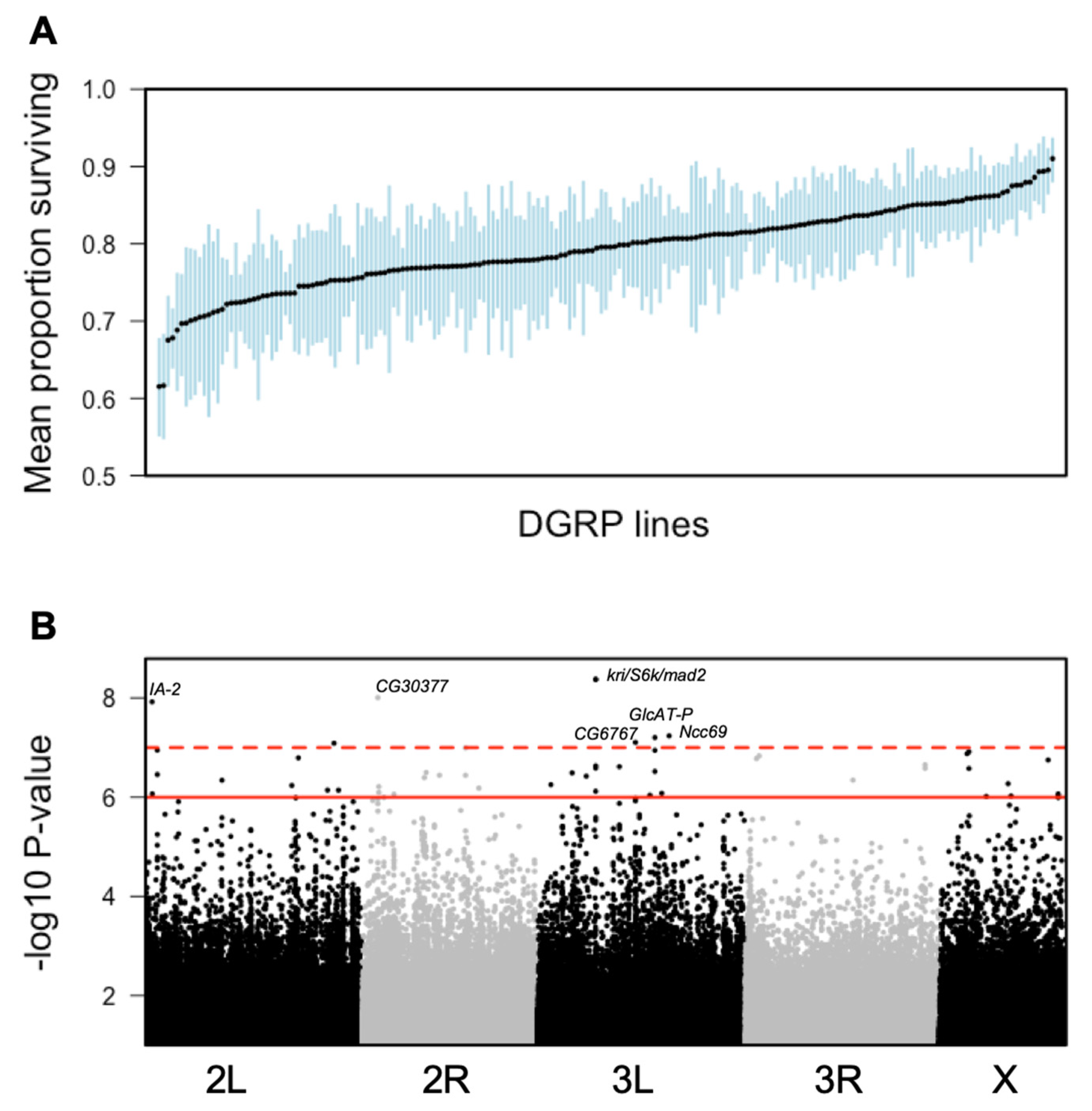
3.1. Variation in Immunity to E. faecalis Infection in the DGRP
3.2. Identification of Candidate Genes
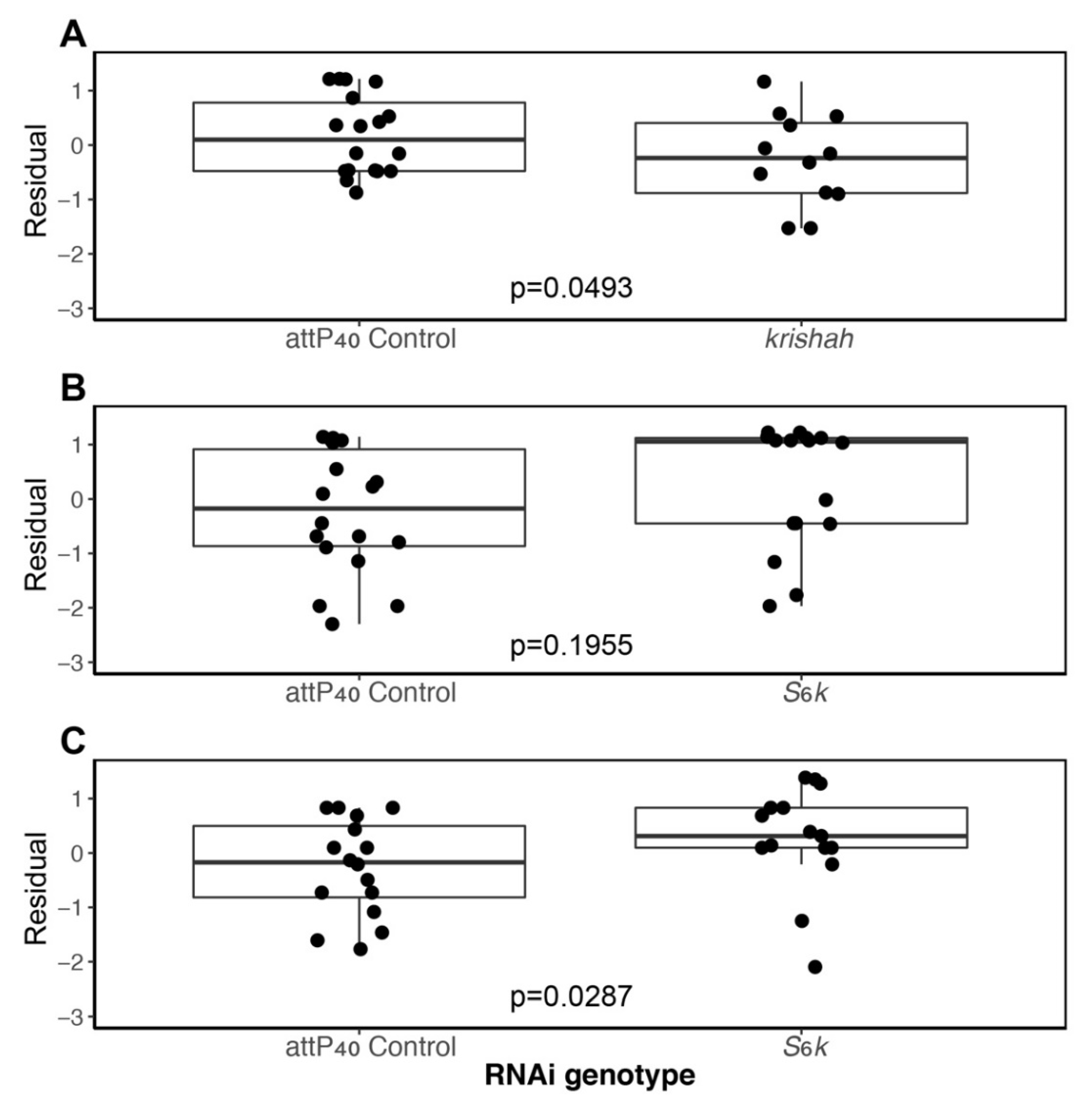
3.3. Candidate Gene Analyses
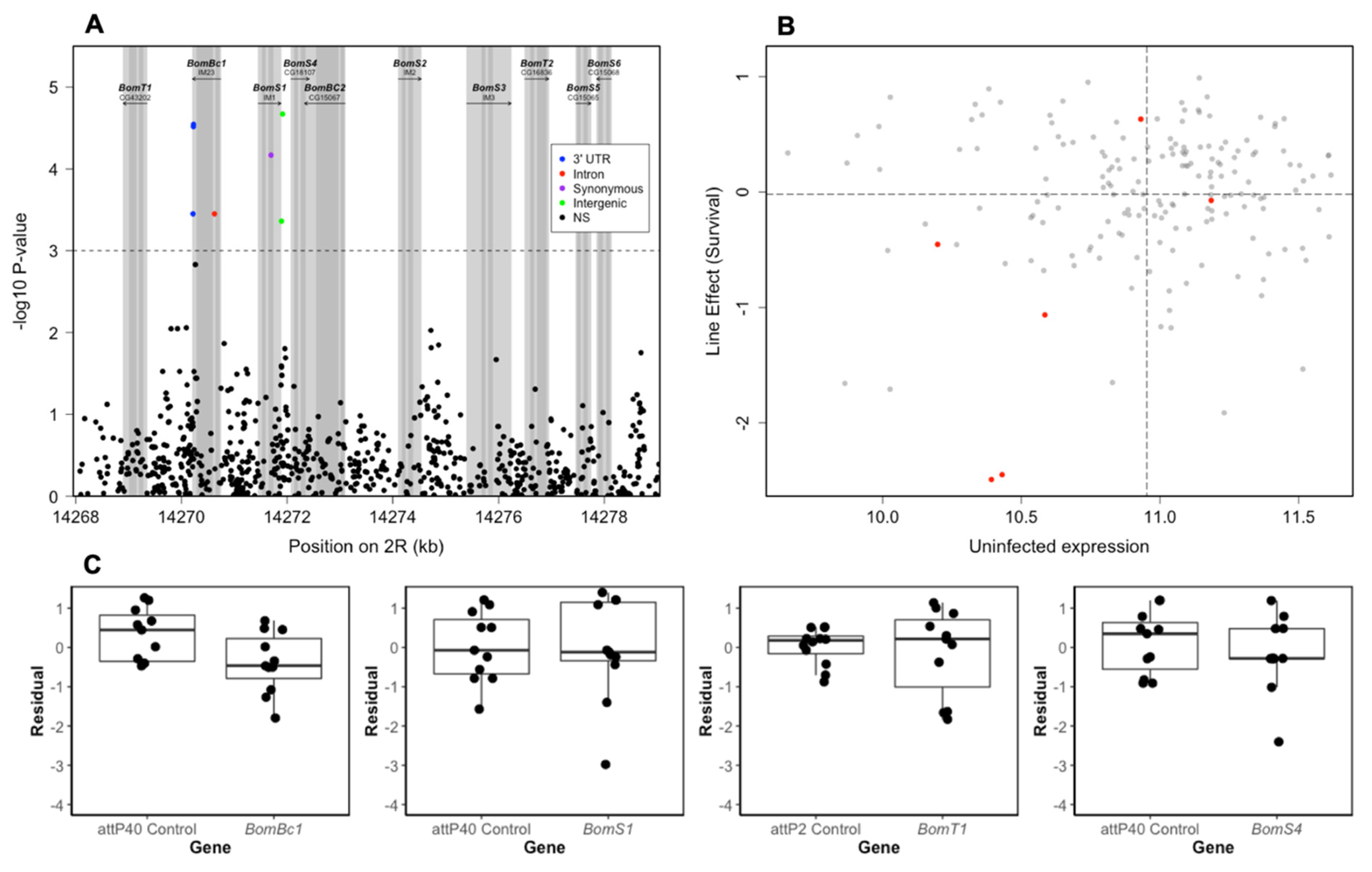
3.4. Role of Bomanin Genes on E. faecalis Survival
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hochachka, W.M.; Dhondt, A.A. Density-dependent decline of host abundance resulting from a new infectious disease. Proc. Natl. Acad. Sci. USA 2000, 97, 5303–5306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daszak, P.; Cunningham, A.A.; Hyatt, A.D. Infectious disease and amphibian population declines. Divers. Distrib. 2003, 9, 141–150. [Google Scholar] [CrossRef] [Green Version]
- Bonneaud, C.; Mazuc, J.; Gonzalez, G.; Haussy, C.; Chastel, O.; Faivre, B.; Sorci, G. Assessing the cost of mounting an immune response. Am. Nat. 2003, 161, 367–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanssen, S.A.; Hasselquist, D.; Folstad, I.; Erikstad, K.E. Costs of immunity: Immune responsiveness reduces survival in a vertebrate. Proc. R. Soc. Lond. B Biol. Sci. 2004, 271, 925–930. [Google Scholar] [CrossRef]
- Minchella, D.J. Host life-history variation in response to parasitism. Parasitology 1985, 90, 205–216. [Google Scholar] [CrossRef]
- Hamilton, W.D. Pathogens as causes of genetic diversity in their host populations. In Population Biology of Infectious Diseases; Anderson, R.M., May, R.M., Eds.; Springer: Berlin, Germany, 1982; pp. 269–296. [Google Scholar]
- Laine, A.-L.; Tellier, A. Heterogeneous selection promotes maintenance of polymorphism in host-parasite interactions. Oikos 2008, 117, 1281–1288. [Google Scholar] [CrossRef]
- Hirschhorn, J.N.; Daly, M.J. Genome-wide association studies for common diseases and complex traits. Nat. Rev. Genet. 2005, 6, 95–108. [Google Scholar] [CrossRef]
- Howick, V.M.; Lazzaro, B.P. The genetic architecture of defence as resistance to and tolerance of bacterial infection in Drosophila melanogaster. Mol. Ecol. 2017, 26, 1533–1546. [Google Scholar] [CrossRef]
- Magwire, M.M.; Fabian, D.K.; Schweyen, H.; Cao, C.; Longdon, B.; Bayer, F.; Jiggins, F.M. Genome-wide association studies reveal a simple genetic basis of resistance to naturally coevolving viruses in Drosophila melanogaster. PLoS Genet. 2012, 8, e1003057. [Google Scholar] [CrossRef]
- Palmer, W.H.; Medd, N.C.; Beard, P.M.; Obbard, D.J. Isolation of a natural DNA virus of Drosophila melanogaster, and characterisation of host resistance and immune responses. PLoS Pathog. 2018, 14, e1007050. [Google Scholar] [CrossRef]
- Unckless, R.L.; Rottschaefer, S.M.; Lazzaro, B.P. The complex contributions of genetics and nutrition to immunity in Drosophila melanogaster. PLoS Genet. 2015, 11, e1005030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.B.; Lu, H.-L.; St Leger, R.J. The genetic basis for variation in resistance to infection in the Drosophila melanogaster genetic reference panel. PLoS Pathog. 2017, 13, e1006260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, C.R.; Gilmore, M.S. Native microbial colonization of Drosophila melanogaster and its use as a model of Enterococcus faecalis pathogenesis. Infect. Immun. 2007, 75, 1565–1576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clemmons, A.W.; Lindsay, S.A.; Wasserman, S.A. An effector peptide family required for Drosophila Toll-mediated immunity. PLoS Pathog. 2015, 11, e1004876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandler, J.A.; Lang, J.M.; Bhatnagar, S.; Eisen, J.A.; Kopp, A. Bacterial communities of diverse Drosophila species: Ecological context of a host-microbe model system. PLoS Genet. 2011, 7, e1002272. [Google Scholar] [CrossRef]
- Lazzaro, B.P. A Population and Quantitative Genetic Analysis of the Drosophila Melanogaster Antibacterial Immune Response. Ph.D. Thesis, Pennsylvania State University, Centre County, PA, USA, 2002. [Google Scholar]
- Lazzaro, B.P.; Sackton, T.B.; Clark, A.G. Genetic variation in Drosophila melanogaster resistance to infection: A comparison across bacteria. Genetics 2006, 174, 1539–1554. [Google Scholar] [CrossRef] [Green Version]
- Duneau, D.F.; Kondolf, H.C.; Im, J.H.; Ortiz, G.A.; Chow, C.; Fox, M.A.; Eugenio, A.T.; Revah, J.; Buchon, N.; Lazzaro, B.P. The Toll pathway underlies host sexual dimorphism in resistance to both Gram-negative and Gram-positive bacteria in mated Drosophila. BMC Biol. 2017, 15, 124. [Google Scholar] [CrossRef] [Green Version]
- Rutschmann, S.; Kilinc, A.; Ferrandon, D. Cutting edge: The Toll pathway is required for resistance to Gram-positive bacterial infections in Drosophila. J. Immunol. 2002, 168, 1542–1546. [Google Scholar] [CrossRef] [Green Version]
- Sackton, T.B.; Lazzaro, B.P.; Clark, A.G. Genotype and gene expression associations with immune function in Drosophila. PLoS Genet. 2010, 6, e1000797. [Google Scholar] [CrossRef] [Green Version]
- Hanson, M.A.; Dostálova, A.; Ceroni, C.; Poidevin, M.; Kondo, S.; Lemaitre, B. Synergy and remarkable specificity of antimicrobial peptides in vivo using a systematic knockout approach. eLife 2019, 8, e44341. [Google Scholar] [CrossRef]
- Lindsay, S.A.; Lin, S.J.H.; Wasserman, S.A. Short-form Bomanins mediate humoral immunity in Drosophila. J. Innate. Immun. 2018, 10, 306–314. [Google Scholar] [CrossRef] [PubMed]
- Mackay, T.F.C.; Richards, S.; Stone, E.A.; Barbadilla, A.; Ayroles, J.F.; Zhu, D.H.; Casillas, S.; Han, Y.; Magwire, M.M.; Cridland, J.M.; et al. The Drosophila melanogaster Genetic Reference Panel. Nature 2012, 482, 173–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, W.; Massouras, A.; Inoue, Y.; Peiffer, J.; Ramia, M.; Tarone, A.M.; Turlapati, L.; Zichner, T.; Zhu, D.H.; Lyman, R.F.; et al. Natural variation in genome architecture among 205 Drosophila melanogaster Genetic Reference Panel lines. Genome Res. 2014, 24, 1193–1208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, J.; Zipperlen, P.; Kubli, E. Drosophila sex-peptide stimulates female innate immune system after mating via the Toll and Imd pathways. Curr. Biol. 2005, 15, 1690–1694. [Google Scholar] [CrossRef] [Green Version]
- McGraw, L.A.; Gibson, G.; Clark, A.G.; Wolfner, M.F. Genes regulated by mating, sperm, or seminal proteins in mated female Drosophila melanogaster. Curr. Biol. 2004, 14, 1509–1514. [Google Scholar] [CrossRef] [Green Version]
- Short, S.M.; Wolfner, M.F.; Lazzaro, B.P. Female Drosophila melanogaster suffer reduced defense against infection due to seminal fluid components. J. Insect. Physiol. 2012, 58, 1192–1201. [Google Scholar] [CrossRef] [Green Version]
- Butterworth, F.M.; Emerson, L.; Rasch, E.M. Maturation and degeneration of the Fat-body in the Drosophila larva and pupa as revealed by morphometric analysis. Tissue Cell 1988, 20, 255–268. [Google Scholar] [CrossRef]
- Felix, T.M.; Hughes, K.A.; Stone, E.A.; Drnevich, J.M.; Leips, J. Age-specific variation in immune response in Drosophila melanogaster has a genetic basis. Genetics 2012, 191, 989–1002. [Google Scholar] [CrossRef] [Green Version]
- Chambers, M.C.; Jacobson, E.; Khalil, S.; Lazzaro, B.P. Consequences of chronic bacterial infection in Drosophila melanogaster. PLoS ONE 2019, 14, e0224440. [Google Scholar] [CrossRef]
- Duneau, D.; Ferdy, J.B.; Revah, J.; Kondolf, H.; Ortiz, G.A.; Lazzaro, B.P.; Buchon, N. Stochastic variation in the initial phase of bacterial infection predicts the probability of survival in D. melanogaster. eLife 2017, 6, e28298. [Google Scholar] [CrossRef] [Green Version]
- Perkins, L.A.; Holderbaum, L.; Tao, R.; Hu, Y.H.; Sopko, R.; McCall, K.; Yang-Zhou, D.; Flockhart, I.; Binari, R.; Shim, H.-S.; et al. The Transgenic RNAi Project at Harvard Medical School: Resources and validation. Genetics 2015, 201, 843–852. [Google Scholar] [CrossRef] [PubMed]
- Abel, T.; Bhatt, R.; Maniatis, T. A Drosophila CREB/ATF transcriptional activator binds to both fat body- and liver-specific regulatory elements. Gene. Dev. 1992, 6, 466–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meister, M.; Lemaitre, B.; Hoffmann, J.A. Antimicrobial peptide defense in Drosophila. Bioessays 1997, 19, 1019–1026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- Bates, D.; Machler, M.; Bolker, B.M.; Walker, S.C. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Wei, T.; Simko, V. R Package “Corrplot”: Visualization of a Correlation Matrix. 2017. Available online: https://github.com/taiyun/corrplot (accessed on 11 November 2019).
- Chapman, J.R.; Hill, T.; Unckless, R.L. Balancing selection drives the maintenance of genetic variation in Drosophila antimicrobial peptides. Genome Biol. Evol. 2019, 11, 2691–2701. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.J.H.; Fulzele, A.; Cohen, L.B.; Bennett, J.; Wasserman, S.A. Bombardier enables delivery of short-form Bomanins in the Drosophila Toll response. Front. Immunol. 2020, 10, 3040. [Google Scholar] [CrossRef]
- Duxbury, E.M.L.; Day, J.P.; Vespasiani, D.M.; Thüringer, Y.; Tolosana, I.; Smith, S.C.L.; Tagliaferri, L.; Kamacioglu, A.; Lindsley, I.; Love, L.; et al. Host-pathogen coevolution increases genetic variation in susceptibility to infection. eLife 2019, 8. [Google Scholar] [CrossRef]
- Schmid-Hempel, P. Evolutionary Parasitology; Oxford University Press: Oxford, UK, 2011; p. 516. [Google Scholar]
- Hill, T.; Koseva, B.S.; Unckless, R.L. The genome of Drosophila innubila reveals lineage-specific patterns of selection in immune genes. Mol. Biol. Evol. 2019, 36, 1405–1417. [Google Scholar] [CrossRef]
- Bulet, P.; Stocklin, R. Insect antimicrobial peptides: Structures, properties and gene regulation. Protein Peptide Lett. 2005, 12, 3–11. [Google Scholar] [CrossRef]
- Hanson, M.A.; Lemaitre, B. New insights on Drosophila antimicrobial peptide function in host defense and beyond. Curr. Opin. Immunol. 2020, 62, 22–30. [Google Scholar] [CrossRef]
- Unckless, R.L.; Howick, V.M.; Lazzaro, B.P. Convergent balancing selection on an antimicrobial peptide in Drosophila. Curr. Biol. 2016, 26, 257–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, T.; Krause, M.W.; Odenwald, W.F.; Toyama, R.; Notkins, A.L. The IA-2 gene family: Homologs in Caenorhabditis elegans, Drosophila and zebrafish. Diabetologia 2001, 44, 81–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.; Bang, H.; Ko, S.; Jung, I.; Hong, H.; Kim-Ha, J. Drosophila ia2 modulates secretion of insulin-like peptide. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2008, 151, 180–184. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.; Wang, J.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; Gwadz, M.; Hurwitz, D.I.; Marchler, G.H.; Song, J.S.; et al. CDD/SPARCLE: The conserved domain database in 2020. Nucleic Acids Res. 2020, 48, D265–D268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Norgate, M.; Southon, A.; Zou, S.G.; Zhan, M.; Sun, Y.; Batterham, P.; Camakaris, J. Copper homeostasis gene discovery in Drosophila melanogaster. Biometals 2007, 20, 683–697. [Google Scholar] [CrossRef] [PubMed]
- Cao, W.; Song, H.J.; Gangi, T.; Kelkar, A.; Antani, I.; Garza, D.; Konsolaki, M. Identification of novel genes that modify phenotypes induced by Alzheimer’s β-amyloid overexpression in Drosophila. Genetics 2008, 178, 1457–1471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egger, B.; Leemans, R.; Loop, T.; Kammermeier, L.; Fan, Y.; Radimerski, T.; Strahm, M.C.; Certa, U.; Reichert, H. Gliogenesis in Drosophila: Genome-wide analysis of downstream genes of glial cells missing in the embryonic nervous system. Development 2002, 129, 3295–3309. [Google Scholar]
- Brown, E.B.; Rayens, E.; Rollmann, S.M. The gene CG6767 afects olfactory behavior in Drosophila melanogaster. Behav. Genet. 2019, 49, 317–326. [Google Scholar] [CrossRef]
- Lince-Faria, M.; Maffini, S.; Orr, B.; Ding, Y.; Florindo, C.; Sunkel, C.E.; Tavares, A.; Johansen, J.; Johansen, K.M.; Maiato, H. Spatiotemporal control of mitosis by the conserved spindle matrix protein Megator. J. Cell Biol. 2009, 184, 647–657. [Google Scholar] [CrossRef] [Green Version]
- Logarinho, E.; Bousbaa, H.; Dias, J.M.; Lopes, C.; Amorim, I.; Antunes-Martins, A.; Sunkel, C.E. Different spindle checkpoint proteins monitor microtubule attachment and tension at kinetochores in Drosophila cells. J. Cell Sci. 2004, 117, 1757–1771. [Google Scholar] [CrossRef] [Green Version]
- Raich, N.; Mahmoudi, S.; Emre, D.; Karess, R.E. Mad1 influences interphase nucleoplasm organization and chromatin regulation in Drosophila. Open Biol. 2018, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghosh, A.C.; Shimell, M.; Leof, E.R.; Haley, M.J.; O’Connor, M.B. UPRT, a suicide-gene therapy candidate in higher eukaryotes, is required for Drosophila larval growth and normal adult lifespan. Sci. Rep. 2015, 5, 13176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watson, K.L.; Chou, M.M.; Blenis, J.; Gelbart, W.M.; Erikson, R.L. A Drosophila gene structurally and functionally homologous to the mammalian 70-kDa S6 kinase gene. Proc. Natl. Acad. Sci. USA 1996, 93, 13694–13698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zahoor, M.-K.; Poidevin, M.; Lecerf, C.; Garrido, D.; Montagne, J. A Drosophila genetic screen for suppressors of S6kinase-dependent growth identifies the F-box subunit Archipelago/FBXW7. Mol. Genet. Genom. 2019, 294, 573–582. [Google Scholar] [CrossRef]
- Cheng, L.; Locke, C.; Davis, G.W. S6 kinase localizes to the presynaptic active zone and functions with PDK1 to control synapse development. J. Cell Biol. 2011, 194, 921–935. [Google Scholar] [CrossRef]
- Wu, Q.; Zhang, Y.; Xu, J.; Shen, P. Regulation of hunger-driven behaviors by neural ribosomal S6 kinase in Drosophila. Proc. Natl. Acad. Sci. USA 2005, 102, 13289–13294. [Google Scholar] [CrossRef] [Green Version]
- Acevedo, S.F.; de Portugal, R.L.P.Y.C.; Gonzalez, D.A.; Rodan, A.R.; Rothenfluh, A. S6 Kinase reflects and regulates ethanol-induced sedation. J. Neurosci. 2015, 35, 15396–15402. [Google Scholar] [CrossRef] [Green Version]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [Green Version]
- Katewa, S.D.; Kapahi, P. Role of TOR signaling in aging and related biological processes in Drosophila melanogaster. Exp. Gerontol. 2011, 46, 382–390. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.E.; Rayyan, M.; Liao, A.; Edery, I.; Pletcher, S.D. Acute dietary restriction acts via TOR, PP2A, and Myc signaling to boost innate immunity in Drosophila. Cell Rep. 2017, 20, 479–490. [Google Scholar] [CrossRef] [Green Version]
- Valanne, S.; Kleino, A.; Myllymäki, H.; Vuoristo, J.; Rämet, M. Iap2 is required for a sustained response in the Drosophila Imd pathway. Dev. Comp. Immunol. 2007, 31, 991–1001. [Google Scholar] [CrossRef] [PubMed]
- De Gregorio, E.; Spellman, P.T.; Tzou, P.; Rubin, G.M.; Lemaitre, B. The Toll and Imd pathways are the major regulators of the immune response in Drosophila. EMBO J. 2002, 21, 2568–2579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffmann, J.A.; Reichhart, J.-M. Drosophila innate immunity: An evolutionary perspective. Nat. Immunol. 2002, 3, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Davoodi, S.; Galenza, A.; Panteluk, A.; Deshpande, R.; Ferguson, M.; Grewa, S.; Foley, E. The Immune Deficiency Pathway regulates metabolic homeostasis in Drosophila. J. Immunol. 2019, 202, 2747–2759. [Google Scholar] [CrossRef]
- Abdel-Nour, M.; Tsalikis, J.; Kleinman, D.; Girardin, S.E. The emerging role of mTOR signalling in antibacterial immunity. Immunol. Cell Biol. 2014, 92, 346–353. [Google Scholar] [CrossRef]
- Ellis, L.L.; Huang, W.; Quinn, A.M.; Ahuja, A.; Alfrejd, B.; Gomez, F.E.; Hjelmen, C.E.; Moore, K.L.; Mackay, T.F.C.; Johnston, J.S.; et al. Intrapopulation genome size variation in D. melanogaster reflects life history variation and plasticity. PLoS Genet. 2014, 10, e1004522. [Google Scholar] [CrossRef] [Green Version]
- Yu, G.Z.; Baeder, D.Y.; Regoes, R.R.; Rolff, J. Combination effects of antimicrobial peptides. Antimicrob. Agents Chemother. 2016, 60, 1717–1724. [Google Scholar] [CrossRef] [Green Version]
- Marxer, M.; Vollenweider, V.; Schmid-Hempel, P. Insect antimicrobial peptides act synergistically to inhibit a trypanosome parasite. Philos. Trans. R. Soc. B 2016, 371. [Google Scholar] [CrossRef]
- Chongsiriwatana, N.P.; Wetzler, M.; Barron, A.E. Functional synergy between antimicrobial peptoids and peptides against Gram-negative bacteria. Antimicrob. Agents Chemother. 2011, 55, 5399–5402. [Google Scholar] [CrossRef] [Green Version]
- Zanchi, C.; Johnston, P.R.; Rolff, J. Evolution of defence cocktails: Antimicrobial peptide combinations reduce mortality and persistent infection. Mol. Ecol. 2017, 26, 5334–5343. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Gene | Type | Factor | Deviance | Df | p-Value |
|---|---|---|---|---|---|
| Krishah | GOI | Line | 3.8655 | 1 | 0.0493 |
| Date | 14.7613 | 3 | 0.0020 | ||
| S6k1 | GOI | Line | 1.6755 | 1 | 0.1955 |
| Date | 7.0270 | 4 | 0.1345 | ||
| S6k2 | GOI | Line | 4.7836 | 1 | 0.0287 |
| Date | 1.0198 | 2 | 0.6006 | ||
| mad2 | GOI | Line | 2.2275 | 1 | 0.1356 |
| Date | 5.9814 | 1 | 0.0145 | ||
| CG42272 | neighbor | Line | 0.5070 | 1 | 0.4765 |
| Date | 6.3657 | 2 | 0.0415 | ||
| CG6767 | GOI | Line | 0.3116 | 1 | 0.5767 |
| Date | 6.0510 | 3 | 0.1092 | ||
| CG6761 | neighbor | Line | 3.1126 | 1 | 0.0777 |
| Date | 3.7781 | 3 | 0.2864 | ||
| IA2 | GOI | Line | 1.5029 | 1 | 0.2202 |
| Date | 1.9210 | 3 | 0.5890 | ||
| Star | neighbor | Line | 3.0205 | 1 | 0.0822 |
| Date | 0.8459 | 2 | 0.6551 | ||
| CG30377 | GOI | Line | 0.1081 | 1 | 0.7423 |
| Date | 0.0669 | 1 | 0.7959 | ||
| Dgk | neighbor | Line | 0.0193 | 1 | 0.8894 |
| Date | 2.9014 | 3 | 0.4071 |
| Gene | Factor | Deviance | Df | p-Value |
|---|---|---|---|---|
| BomBc1 | Line | 5.1995 | 1 | 0.0226 |
| Date | 3.3653 | 2 | 0.1859 | |
| BomS1 | Line | 0.2420 | 1 | 0.6228 |
| Date | 9.8015 | 2 | 0.0074 | |
| BomT1 | Line | 0.0041 | 1 | 0.9491 |
| Date | 10.8707 | 2 | 0.0044 | |
| BomS4 | Line | 0.7496 | 1 | 0.3866 |
| Date | 2.2465 | 2 | 0.3252 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chapman, J.R.; Dowell, M.A.; Chan, R.; Unckless, R.L. The Genetic Basis of Natural Variation in Drosophila melanogaster Immune Defense against Enterococcus faecalis. Genes 2020, 11, 234. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11020234
Chapman JR, Dowell MA, Chan R, Unckless RL. The Genetic Basis of Natural Variation in Drosophila melanogaster Immune Defense against Enterococcus faecalis. Genes. 2020; 11(2):234. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11020234
Chicago/Turabian StyleChapman, Joanne R, Maureen A Dowell, Rosanna Chan, and Robert L Unckless. 2020. "The Genetic Basis of Natural Variation in Drosophila melanogaster Immune Defense against Enterococcus faecalis" Genes 11, no. 2: 234. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11020234