Experimental Evolution of Legume Symbionts: What Have We Learnt?


LIPM, Université de Toulouse, INRAE, CNRS, Castanet-Tolosan 31320, France
*
Author to whom correspondence should be addressed.
Genes 2020, 11(3), 339; https://0-doi-org.brum.beds.ac.uk/10.3390/genes11030339
Submission received: 3 March 2020
/
Revised: 17 March 2020
/
Accepted: 20 March 2020
/
Published: 23 March 2020
(This article belongs to the Special Issue Genetic Evolution of Root Nodule Symbioses)
{kind=link}
{kind=link}
{kind=link}
Abstract
:Rhizobia, the nitrogen-fixing symbionts of legumes, are polyphyletic bacteria distributed in many alpha- and beta-proteobacterial genera. They likely emerged and diversified through independent horizontal transfers of key symbiotic genes. To replay the evolution of a new rhizobium genus under laboratory conditions, the symbiotic plasmid of Cupriavidus taiwanensis was introduced in the plant pathogen Ralstonia solanacearum, and the generated proto-rhizobium was submitted to repeated inoculations to the C. taiwanensis host, Mimosa pudica L. This experiment validated a two-step evolutionary scenario of key symbiotic gene acquisition followed by genome remodeling under plant selection. Nodulation and nodule cell infection were obtained and optimized mainly via the rewiring of regulatory circuits of the recipient bacterium. Symbiotic adaptation was shown to be accelerated by the activity of a mutagenesis cassette conserved in most rhizobia. Investigating mutated genes led us to identify new components of R. solanacearum virulence and C. taiwanensis symbiosis. Nitrogen fixation was not acquired in our short experiment. However, we showed that post-infection sanctions allowed the increase in frequency of nitrogen-fixing variants among a non-fixing population in the M. pudica–C. taiwanensis system and likely allowed the spread of this trait in natura. Experimental evolution thus provided new insights into rhizobium biology and evolution.
1. Introduction
The diversity of current living forms is the result of evolutionary processes that have unfolded over billions of years but are difficult to trace in the absence of fossils. With a few exceptions, where dense sampling of natural populations can be done over short periods [1,2,3], only comparative genomics and phylogeny of extant species allow the prediction of ancestral gene contents and evolutionary scenarios. However, adaptive processes can be analyzed in real time via experimental evolution (EE). EE, the propagation of over tens of thousands of generations of organisms in a controlled environment [4,5,6], allows the investigation of the phenotypic, genomic and molecular adaptation thanks to the analysis of fossil records that are generated and stored during the experiment. When laboratory conditions mimic key conditions of natural environments, EE may allow for the understanding of natural evolutionary processes.
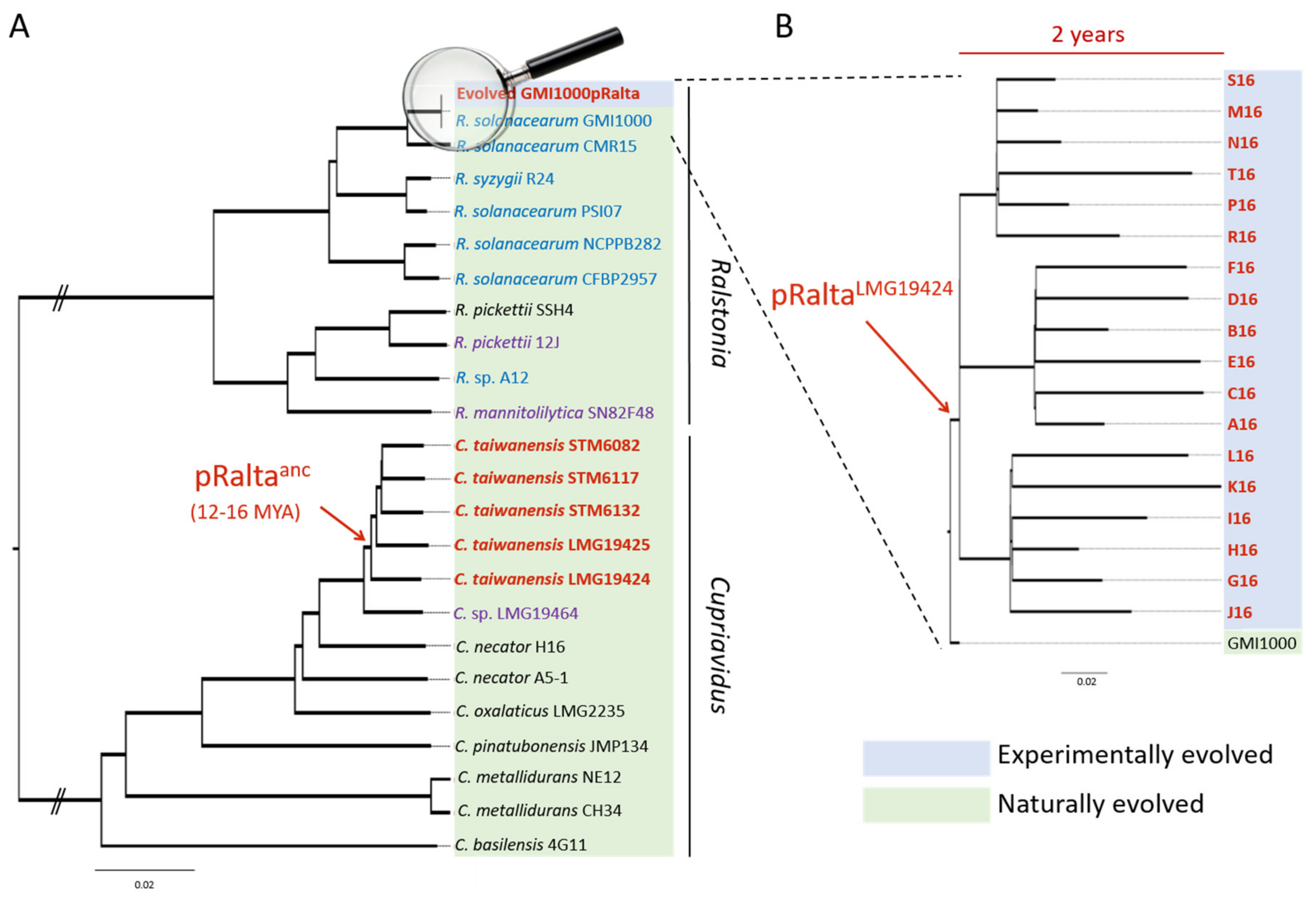
Nitrogen-fixing symbioses have evolved between polyphyletic soil bacteria, called rhizobia, and legumes. In the presence of compatible host plants, rhizobia elicit the formation of novel plant organs, the nodules, which they colonize massively. Inside nodule cells, bacteria reduce large amounts of nitrogen gas into ammonium, which is directly assimilated by the plant and contributes to its growth. On the plant side, this trait is predicted to result from a single gain before the radiation of the Fabales, Fagales, Cucurbitales and Rosales clade (FaFaCuRo), which is followed by multiple independent losses in most descendant lineages [7,8], but maintained in most genera of the Fabales (legumes). On the bacterial side, the trait likely emerged in Frankia [9,10]. The genes that determine the synthesis of rhizobial lipo-chitooligosaccharidic Nod factors triggering nodulation are thought to have been transferred from some Frankia to a diazotrophic proteobacterium, generating the first rhizobium. After this, nodulation (nod) genes and nitrogen fixation (nif-fix) genes cluster together on mobile genetic elements, either plasmids or genomic islands, and spread to hundreds of species of distantly related taxa among alpha- and beta-proteobacteria via horizontal gene transfers (HGT) [11,12,13]. Some rhizobia, e.g., the rhizobial Burkholderia species, are ancient symbionts of legumes, which probably appeared shortly after legumes emerged, ca. 60 million years ago [14,15,16]. Others, such as Cupriavidus taiwanensis, the symbiont of Mimosa pudica, evolved more recently. C. taiwanensis was estimated to have emerged 12–16 million years ago following a single acquisition of a symbiotic plasmid, likely from Burkholderia [15,17,18]. The ancestral rhizobial C. taiwanensis then further diversified into five distinct genospecies with a vertical inheritance of the symbiotic plasmid in most cases (Figure 1).
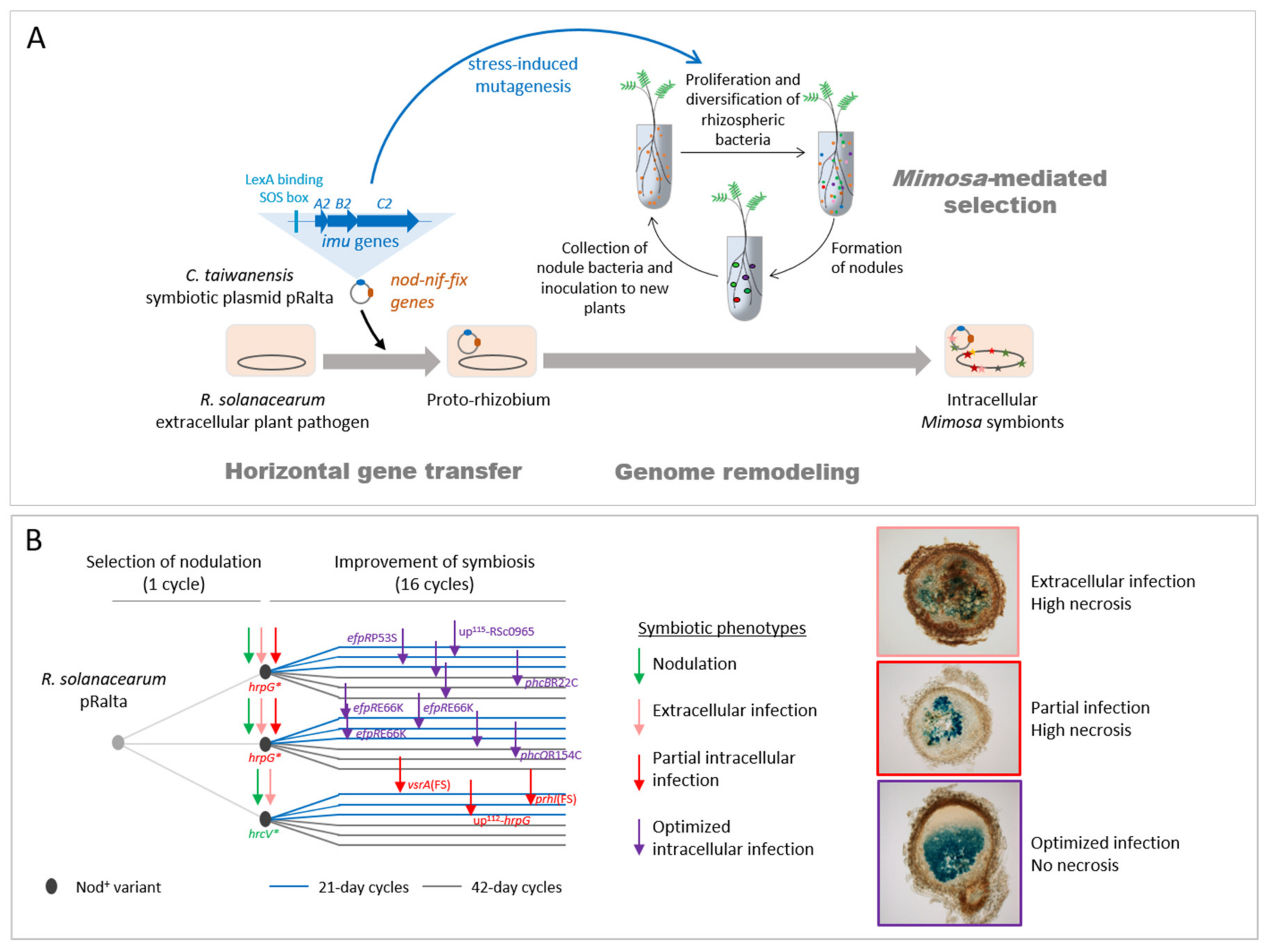
The Cupriavidus branch is a neighbour to Ralstonia, a genus that only contains saprophytic and pathogenic strains. The introduction of the symbiotic plasmid of C. taiwanensis LMG19424 into the plant pathogenic Ralstonia solanacearum GMI1000 strain generated a still pathogenic strain unable to nodulate Mimosa pudica, indicating that the acquisition of essential symbiotic genes may not be sufficient to convert a soil bacterium into a legume symbiont. To replay the emergence of a novel genus of rhizobium, here Ralstonia, we experimentally evolved this chimeric strain using laboratory conditions reproducing environmental conditions that may have favoured symbiosis [19,20]. Bacteria were repeatedly inoculated onto Mimosa plantlets in the absence of other rhizobial competitors (see Figure 2 for the experimental design). At the end of the experiment, 18 final evolved clones were isolated and their evolution analyzed (Figure 1). Despite strong differences between natural and experimental evolution of Mimosa symbionts in terms of genetic backgrounds, time frame and environmental conditions (Figure 1), the laboratory process exhibited striking parallels with natural evolution [17]. In particular, adaptation was characterized by a predominance of purifying selection (a purge of non-synonymous mutations) and associated with positive selection in a set of genes that led to the co-option of the same quorum-sensing system in both processes, while no adaptation was observed in the plasmid carrying the genes responsible for the ecological transition. Moreover, this experiment provided new insights into the evolutionary mechanisms leading to endosymbiosis with legumes and on the biology of rhizobia. Here we review some of the lessons learnt from our evolution experiment.
2. Evidence for a Two-Step Evolutionary Scenario
The pioneering works of Sullivan et al. demonstrated for the first time the conversion of soil bacteria into legume symbionts in natura through the horizontal acquisition of a symbiotic genomic island [25,26]. Since then, it has been well accepted that rhizobia diversified from independent events of horizontal transfer of essential nod and nif-fix genes. This finding was then supported by many genomic and phylogenetic analyses [11,15,27,28] and other field experiments [29,30]. However, the transferred genes are not the only ones involved in symbiosis with legumes. Many genes located outside the mobile symbiotic regions have been described as engaged in symbiosis. Among them, specific surface polysaccharide biosynthesis genes (eps, kps, lps, ndv) [31,32], metabolism genes (glnBK, ilvCD, hemAH, leuCB, phbAC, dme, pckA) [33,34,35], transporters such as mdt, znu (zinc), dctA (dicarboxylic acid), and pstABC (phosphate) genes [36,37,38,39], chaperones (groEL/groES) [40] and secretion systems [41] appear to be crucial for symbiosis in different rhizobia. Contrary to the nod and nif-fix genes, the majority of these symbiotic genes is lineage-specific [33,42]. Using in planta transposon insertion sequencing (Tn-Seq) approaches, Pobigaylo et al. and Flores-Tinoco et al. estimated that 8–15% of Sinorhizobium meliloti genes (500 to 900 genes) are involved in symbiosis [43,44]. These genes are scattered throughout the entire genome and mostly involved in metabolism, gene regulation and other cellular processes, confirming the diversity of genes required to achieve symbiosis. On the other hand, some bacterial functions can be deleterious for symbiosis. For example, type 3 secretion systems (T3SS) were shown to decrease the symbiotic capacity of some rhizobium strains or prevent their interaction with some host plants [41,45,46,47]. Furthermore, transfers of symbiotic genes performed under laboratory conditions did not always turn a non-symbiotic organism into an efficient legume symbiont. Indeed, the transfer of diverse rhizobium symbiotic plasmids into Agrobacterium tumefaciens or Escherichia coli, led to strains that were able to form either pseudo- or non-functional nodules [48,49,50] or nodules fixing low amounts of nitrogen [51]. In line with this, suboptimal symbionts, forming non-fixing nodules, have been observed in the wild following the horizontal acquisition of a symbiosis island [29].
We thus made the hypothesis that the evolution of a new rhizobial species/genus required a post-transfer adaptation step allowing the recruitment and/or the inactivation of specific functions of the recipient genome, especially if the transfer occurred between bacteria with different genomic backgrounds and/or lifestyles. We hypothesized that this optimization step occurred via genome remodeling under plant selection pressure. Indeed, legumes control their microsymbionts through a multistep surveillance system. All along the symbiotic process, plants regularly check the symbiotic status of bacteria and allow, or don’t allow, their proliferation [52]. These checkpoints include the recognition of bacterial Nod-factors and expolysaccharides via specific receptors that convert these signals into a plant developmental program [53,54,55,56,57,58,59], and the production and delivery of ammonium to plant cells, which prevent plant sanctions [60,61,62].
We experimentally validated this two-step evolutionary scenario (Figure 2). First, the transfer of the symbiotic plasmid of the Mimosa symbiont C. taiwanensis into the plant pathogen Ralstonia solanacearum led to a non-nodulating (Nod-) strain, showing that, in this case, the transfer of the main symbiotic genes was not sufficient to convert Ralstonia into Mimosa symbionts. Further evolution of this Nod- Ralstonia chimeric strain through repeated inoculations to Mimosa pudica plantlets and re-isolation of bacteria from nodules allowed the parallel acquisition and progressive improvement of two symbiotic traits (nodulation and nodule cell infection) in independent lineages [19,20,21]. After 16 such cycles, evolved strains of most lineages induced nodules that display features of true nodules: a peripheral vascular system, the production of leghemoglobin, and cells filled with symbiosomes containing bacteria. However, mutualism was not achieved at that stage of the experiment and intracellular bacteria did not persist and prematurely degenerated. It should be noted that in nature rhizobia having suboptimal capabilities of nodulation competitiveness, infection, or nitrogen fixation, or inducing visible plant defense reactions in nodules, have been isolated [29,63,64]. As suggested by our evolution experiment, these natural isolates might correspond to intermediate evolutionary stages before the achievement of symbiosis with their host plant.
When legumes emerged, around 60–100 million years ago [14,65], nearly all extant proteobacterial genera existed and the different rhizobial lineages had already diverged [66,67]. Among α and β proteobacteria, only a few genera (15 α- and 3 β-proteobacterium genera) have been successfully colonized by symbiotic nitrogen fixation traits, suggesting that only these genera were predisposed for symbiosis. In a comparative genomics study, Garrido-Oter et al. proposed that an ancestral state of adaptation to the root environment in the Rhizobiales predisposed these bacteria to symbiosis [68]. Based on a Transposon-Sequencing (Tn-Seq) approach, Salas et al. also came to the same conclusion that rhizobia were adapted to the rhizosphere of their host plants before their specialization in nodulation [69]. The adaptation to rhizosphere colonization may include the capacity to use specific plant metabolites [70,71] and tolerance to biotic and abiotic stresses. Predisposition to legume symbiosis may require, in addition, the production of specific surface polysaccharides (EPS, KPS, LPS), and the capacity to avoid plant defenses [72,73]. Although we did not identify which genetic features predispose a bacterium to legume endosymbiosis, we provided evidence that a strain with a completely different lifestyle (pathogenic and strictly extracellular) can be converted into an intracellular legume symbiont. This revealed that indigenous functions of the recipient genome were recruited and/or suppressed for symbiosis. The capacity of R. solanacearum to thrive in many plant environments and infect more than 250 different plant species [74], including some legume species [75], might have contributed to its predisposition to symbiosis.
Phylogenetic studies predicted that mutualistic bacteria often derived from pathogens whilst the opposite was very rare [76]. C. taiwanensis might indeed have evolved from an opportunistic pathogen, since the closest identified bacterium to symbiotic C. taiwanensis was isolated from a cystic fibrosis patient (C. sp. LMG19464) [77], and these symbionts have an atypical rhizobial T3SS, which is not connected to the regulation of nodulation [45] and whose organization is similar to the T3SS of the human opportunistic bacterium Burkholderia cenocepacia [78]. Transitions from parasitism to mutualism were empirically demonstrated in other evolution experiments, that, for instance, converted E. coli parasitic phages [79], nematode pathogenic bacteria [80] or mice gut fungal pathogens [81] into beneficial symbionts.
3. Regulatory Rewiring of the Recipient Genome as a Main Driver of Symbiotic Adaptation
While the genetic mechanisms of transfer of mobile genetic elements, including symbiosis plasmids and genomic islands, have been widely studied and well described [82,83,84,85], post-transfer adaptation steps are still underexplored. Our evolution experiment allowed us to tackle this issue. To understand how R. solanacearum that acquired a symbiotic plasmid further evolved into Mimosa intracellular symbionts, we resequenced the genome of many intermediate and final evolved clones along parallel lineages. This led us to analyze the genotype/phenotype correlations and identify the main symbiosis-adaptive mutations.
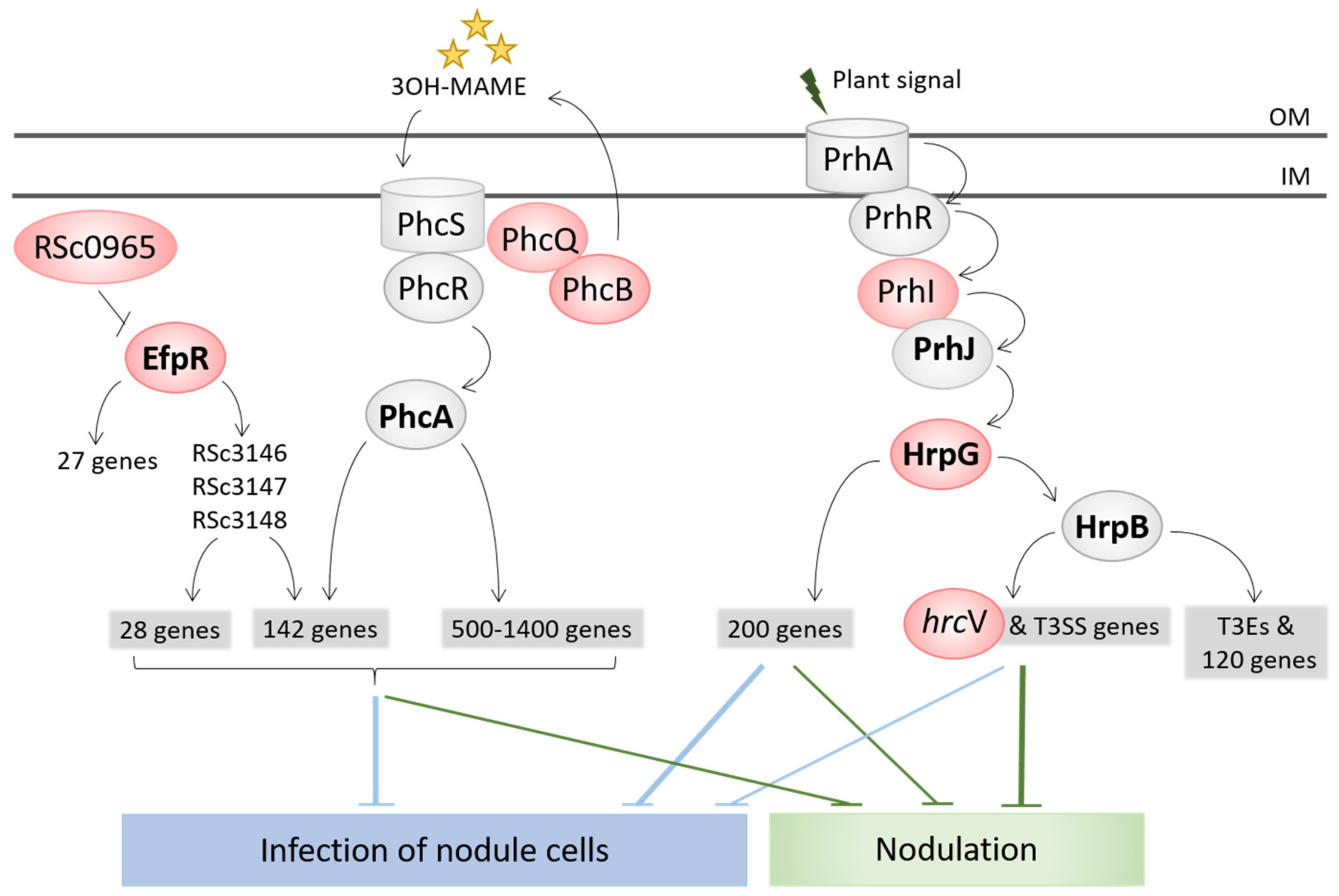
The capacity to enter roots and form nodules was unlocked by the inactivation of the T3SS (hrcV mutation), which is the main virulence factor of R. solanacearum [74]. The sole inactivation of the T3SS led to the formation of necrotic nodules that were only extracellularly invaded. A first level of intracellular infection, i.e., infection of nodule cells, was gained via the inactivation of virulence regulators, either VsrA (frameshift mutation in the vsrA gene) or HrpG (stop mutations in the hrpG gene, or mutation in the hrpG promoter region or in the sigma factor prhI acting upstream from HrpG in the regulatory cascade) [21,22]. The stop mutations in HrpG allowed the concomitant gain of the nodulation capacity since HrpG controls the T3SS and associated effectors [86], while other mutations (in vsrA, hrpG promoter and prhI) arose after the hrcV mutation having unlocked nodulation (Figure 2B). HrpG and VsrA thus likely control functions that positively or negatively interfere with intracellular infection. A small number of infected cells and the presence of necrotic zones characterized this first level of intracellular infection. A second level of infection, characterized by a high number of infected cells and an almost total absence of necrosis, was then reached in several lines. This second level of infection was obtained via mutations affecting global transcription regulators, either EfpR or PhcA. Missense mutations in efpR itself or an intergenic mutation upstream from a gene of unknown function triggered a constitutive repression of EfpR [23], while mutations in two different components of the Phc quorum sensing system (phcB and phcQ) led to a modulation of the quorum sensing threshold activating PhcA [24]. EfpR and PhcA were shown to act as both central players of the R. solanacearum virulence regulatory network and global catabolic repressors [23,87,88,89]. Interestingly, two genes of the phc regulatory pathway, phcB and phcS, display a signature of positive selection in naturally evolved Mimosa symbionts and we showed that PhcA was co-opted for symbiosis in C. taiwanensis LMG19424 [17].
Altogether, our analysis showed that the rewiring of regulatory circuits was the main driver of the symbiotic adaptation of Ralstonia. Strikingly, all identified adaptive mutations for infection affect global regulatory proteins (HrpG, VsrA, EfpR, PhcA) controlling, positively or negatively, the expression of several hundreds of genes (Figure 3) [23,86,89,90,91,92]. This is in accordance with other evolution experiments in which global regulators were frequently targeted [93,94,95,96,97]. Our results confirmed that mutations in global regulators can be highly beneficial by changing the expression of hundreds of genes in a concerted way, particularly during the first steps of adaptation to a new environment [98]. The fact that the conversion of R. solanacearum into intracellular legume symbionts was mainly driven by regulation rewiring means that ancillary functions required for nodule infection are there but need to be expressed at the right time and place. A future challenge will be to identify which functions controlled by the targeted regulators are beneficial or deleterious for intracellular infection in the Ralstonia–Mimosa interaction. We hypothesize that the same functions are affected in the different lines, but the large overlap between the EfpR and PhcA regulons [24] makes this task difficult (Figure 3).
In natural rhizobia, a large number of regulatory systems involved in symbiosis were described. These systems vary a lot among rhizobia, even in close natural taxons. For example, the expression of nodulation genes is regulated by NodD proteins in all rhizobia. However, depending on rhizobium species, the expression of these genes is controlled by either a single NodD regulatory protein [78] or multiple, both positive and negative, regulators organized in intricate networks [99,100,101,102,103]. Likewise, expression of nitrogen fixation (nif-fix) genes is under the control of the conserved NifA protein, but upstream from NifA the regulatory cascade is highly variable among rhizobia, involving different two-component systems (FixLJ, RegSR, NtrBC, ActRS, FixL-FxkR) responding to various signals [104,105,106,107] and specific downstream transcription regulators (FixK1, FixK2, FnrN) [108,109,110]. Surface polysaccharide biosynthesis genes are other striking examples of essential symbiotic genes subjected to very complex regulations involving a multitude of interconnected regulators that largely differ between rhizobium species [111,112]. Integration of symbiotic functions (both endogenously recruited and horizontally acquired) in the regulatory circuits of the recipient bacterium is thus probably a key step in the evolution of new rhizobium species. In our EE, depending on the lineages, different regulatory pathways were modified by bacteria to achieve nodulation and infection, thus making a noticeable parallel between natural and experimental evolutionary processes of rhizobia.
So far, no adaptive mutations were identified on the symbiotic plasmid in the experiment, indicating that adaptation mainly occurred on the recipient genome. This is in accordance with the lack of evidence of positive selection of plasmidic genes in natural Mimosa pudica symbionts [17]. A possible explanation is that in the natural evolution, like in the experimental one, the symbiotic plasmid originated from a more ancient Mimosa symbiont [13,16], and did not require further adaptation to the legume host.
4. The Legume Plant, a Strong Selection Pressure for Shaping Bacterial Endosymbiotic Evolution
Horizontal gene transfer (HGT) of symbiotic genes is frequent in nature [113,114], which is consistent with the observations that rhizosphere and nodule environments promote HGT [85,115]. Given that rhizobia co-exist in the soil, and even in nodules, with many other different bacteria [116,117], the transfer of symbiosis genes to new bacterial genera must be extremely common. The fact that rhizobia are restricted to a relatively small number of bacterial genera [13,118] suggests that exceptional circumstances are needed for the successful actualization of symbiotic potential in strains having received the nod and nif-fix genes. We can infer what these circumstances are by learning from field experiments with Lotus, Biserrula pelecinus and soybean symbionts [23,27,28,116]: (i) introduction of an exotic legume plant with a compatible rhizobium which is mal-adapted to the new environmental conditions, (ii) the presence of endogenous soil bacteria already adapted to the environment where the legume is introduced, and (iii) the absence of competing rhizobia.
Our experiment shows that, once these circumstances are reproduced in the laboratory, the plant exerts a strong selection pressure for the full expression and optimization of symbiotic traits. It was already well known that the plant selects the most nodulation-competitive strains. While large bacterial populations are present in the rhizosphere [119,120,121], each nodule is formed by one or a few bacteria [122]. Thus, bacterial root entry and nodulation represent a strong bottleneck and, in a mixed community, only the most competitive bacteria successfully infect the plant [123]. This property was used as a genetic screen in the past [124] and was evidenced in our EE where nodulation competitiveness strongly improved all along cycles [19]. In most cases, the genetic factors underpinning this phenotype have not been identified in our experiment (with a few exceptions, see below). Adaptive mutations improving nodulation could affect genes involved in diverse functions (growth in the rhizosphere, motility, biosynthesis of cell surface components) since it is known that multiple bacterial traits affect nodulation competitiveness [123,125].
Our evolution experiment showed for the first time that a late symbiotic stage, the infection of nodule cells, could be acquired and further improved by plant-mediated selection through two mechanisms. Firstly, bacteria multiply at much higher densities during intracellular infection (ca. 108–109 bacterial cells per nodule) than in infection threads and extracellular spaces (ca. 106 bacteria per nodule). Secondly, we observed a coupling between infection and nodulation since all mutations that improved intracellular infection also improved nodulation competitiveness [22] (and unpublished data). The genetic link between infection and nodulation is supported by the finding that mutations optimizing infection in our EE modify the expression of bacterial genes during the earliest stages of symbiosis (infected root hairs) [24]. This coupling is also consistent with the known effect of Nod factors on intracellular infection [126,127,128] and the fact that the intracellular release of rhizobia in primodium cells depends on the proper progression of infection threads [129], which otherwise conditions nodule development. From an evolutionary perspective, the nodulation/cell invasion coupling enables the selection at the root entry level of a bacterial trait that is only expressed at the later stages of the symbiotic process. These results also call for a more extensive examination of this question by systematically measuring nodulation competitiveness of non-infectious rhizobial mutants, such as exopolysaccharide deficient mutants, which has been never, or at best extremely rarely, performed.
However, the acquisition of intracellular infection was not universal in our EE as some lines (ancestor CBM356 and derived lineages with 42-day cycles) did not evolve infection (Figure 2B). Thus, depending on chance (i.e., the nature of the first nodulation-adaptive mutation) and selective regime (long (42 day) vs. short (21 day) cycles), extracellular-infecting clones may prevail over intracellular ones, possibly leading to an evolutionary dead-end.
Following their accommodation into plant cells and when oxygen concentration is low enough, rhizobia initiate nitrogen fixation, the critical process required for nutrient exchanges during this mutualistic symbiosis. A number of studies have shown that nitrogen fixation is not selected at the root entry/nodulation level per se [60,130,131,132], unless it is associated with another trait [133,134,135]. Instead, post-infection control mechanisms promote fitness of fixing bacteria over non-fixing ones [61,62]. These mechanisms were hypothesized to contribute to the stability of mutualism over long time-scales [136,137], by limiting the proliferation of non-fixing bacteria that may otherwise outcompete fixing bacteria that invest cellular resources into the energetically costly process of nitrogen fixation. In our system, we were interested in tackling another question: do post-infection control mechanisms allow the evolution of mutualism in natura and in our experimental setup? In other words, would nitrogen-fixing clones be selected in our EE, if they happen to arise? Using the reference C. taiwanensis/M. pudica system and isogenic Fix+ and Fix- bacterial mutants, we followed the dynamics of the two bacterial populations along the symbiotic process and identified the timing and extent of the post-infection control mechanisms specifically targeting Fix- cells [60]. We show that Fix- bacteria multiply similarly to Fix+ bacteria in nodules, but only until 16–21 days post-infection, the time at which their fitness begins to decline compared to Fix+ bacteria due to a premature degeneration. Interestingly, differential survival of Fix- vs. Fix+ bacteria occurred when single nodules were co-infected by the two bacterial genotypes. This, together with similar results obtained with Lotus and Acmispon [138], indicated the existence of cell-autonomous sanctions. Moreover, mathematical modeling predicted the impact of different environmental factors on the kinetics of invasion of rare Fix+ clones in a Fix- population, and these predictions were experimentally validated by witnessing the rapid increase in frequency of Fix+ bacteria along serial inoculation–nodulation cycles [60]. An important topic for future research will be to identify the molecular bases of post-infection control mechanisms. Previous work showed that soybean nodules could restrict oxygen diffusion in non-fixing nodules [61], but the generality and mechanistic details of this process remain to be investigated.
Together, our work has shown that plant-mediated selection mechanisms were efficient in promoting the successive evolution of key symbiotic properties: nodulation, intracellular infection and nitrogen fixation. While we focused here on plant-mediated selection factors observed in our EE, it is worth remembering that additional biotic and abiotic factors (including physical environment, soil bacterial communities, plant life histories) also shape bacterial adaptation in natural populations [139].
5. Discovery of a Hypermutagenesis Mechanism that Accelerates HGT-Based Evolution
The generation of genetic variation through de novo mutations fuels bacterial adaptation to new environments. Mutations in genes involved in DNA repair, such as mutS/L/H, can lead to a hypermutator phenotype, characterized by a constitutive elevation of mutation rate and, consequently, increased evolvability. Hypermutator genotypes are often selected in evolution experiments or during latent host colonization [140,141]. Bacteria can also increase mutation rates transiently in response to stress, a mechanism known as stress-induced mutagenesis (SIM) [142,143]. SIM is due to the expression of error-prone DNA polymerases under the control of major stress-induced regulatory pathways such as SOS or RpoS. Despite the long-term recognition of this phenomenon, the contribution of SIM to bacterial adaptive evolution has received relatively little experimental scrutiny.
Resequencing evolved clones revealed a large number of mutations in our experiment, although the strains were not constitutive hypermutators. Additional sequencing and experiments revealed that bacterial clones underwent transient hypermutagenesis (~5–10× increase in mutation rate) during each exposure to the plant culture medium, with a further increase (~4×) in the presence of Mimosa pudica plantlets. We identified error-prone DNA polymerases from the imuABC family that were located on pRalta and responsible for this transient hypermutagenesis [144]. In our system, SIM accelerates adaptation since, when re-evolved for 5 additional cycles, clones carrying imuABC genes (like our original ancestor) adapted faster than isogenic imuABC mutants evolved in the same conditions [144]. Interestingly, the interrogation of genomic databases revealed than more than half of symbiotic plasmids carried error-prone DNA polymerases. It is tempting to speculate that the co-transfer of symbiosis genes with mutagenesis cassettes may facilitate adaptation to new symbiotic environments following plasmid acquisition by increasing the probability to generate variants with improved symbiotic capabilities that the plant can select. The fact that the natural process, as the experimental one, resulted in rapid genetic diversification dominated by purifying selection, also argues in favor of the role of imuABC-based transient mutagenesis in the natural evolution of rhizobia. Whether dependent on plasmidic or chromosomal genes, SIM may contribute to bursts in mutation rates occurring during host-microbe interactions [145,146]. Moreover, it is worth mentioning that other mechanisms can increase evolvability in rhizobia, such as the activation of HGT by root exudates [85], the transposition of insertion sequence elements [47,147], and the Non-Homologous End-Joining mechanism (NHEJ) [148].
Balancing the benefits during the early steps of adaptation to a new environment, hypermutagenesis can compromise fitness on the long term due to the accumulation of deleterious mutations [140]. This can lead to selection against hypermutagenesis, consistent with the modulation of mutation rates observed in several evolution experiments [149,150,151]. Testing whether SIM can be counter-selected would bring interesting new arguments into the debate over the adaptive nature of SIM. In that respect, it is intriguing to note that SIM does not occur in the original host of pRalta, C. taiwanensis, but the causes, environmental or genetic, are unknown.
6. New Insights into the Biology of Evolutionary Protagonists
EE uses extant strains with specific properties or lifestyles as protagonists. The tracking of mutations underlying phenotypic changes during laboratory evolution may reveal unidentified molecular players of functions otherwise studied by traditional molecular genetics [152,153]. In our experiment, this potential to advance the understanding of the biology of ancestral strains was highlighted by the discovery of new virulence and symbiotic components in R. solanacearum and C. taiwanensis, respectively.
The virulence mechanisms of R. solanacearum have been genetically dissected for many years. This allowed for the identification of a complex regulatory network involving many interconnected regulatory cascades, which controls the different functions involved in pathogenicity [154,155,156,157,158]. In this network, the LysR family transcriptional regulator PhcA was identified as a central regulator, whose activation is controlled by the quorum sensing (QS) PhcBSR(Q) system [159]. However, the role of the last gene of the operon, phcQ, could not be evidenced [160]. In our experiment, we focused on a phcQ mutation that enhanced the capacity of bacteria to infect nodule cells. This led us to show that this mutation delayed the cell density-dependent activation of PhcA and further showed that phcQ is involved in the production of QS molecules [24]. In addition, our experiment and another one conducted in parallel [161] revealed the occurrence of a virulence pathway previously overlooked, the efpR pathway. Mutations in this gene or in one upstream regulatory component allowed a better in planta colonization of R. solanacearum evolved on different host plants [87] and an improved intracellular infection capacity of the chimeric Ralstonia evolved on the M. pudica legume [23]. Mutation in the upstream yet unidentified regulatory component (a SNP in the intergenic region upstream from the Rsc0965 gene) led to a surprising complete loss of virulence.
The nod genes conveyed by symbiotic plasmids and islands determine the biosynthesis of signalling compounds, called Nod factors, which trigger the plant developmental program leading to nodulation and infection in legumes [162,163]. In 2008, Amadou et al. found that the Nod factors produced by C. taiwanensis were consistent with the in silico prediction based on the genome sequence and the identified nod genes [78]. They were found to be pentameric chito-oligomers sulphated at the reducing end, N-acylated by vaccenic acid or palmitic acid and mostly substituted by an N-methyl and a carbamoyl group at the non-reducing end. This was recently challenged from the analysis of one of our evolved lines. In the R line, the capacity to infect nodule cells was not obtained after 16 cycles of evolution despite the detection of an adaptive mutation for intracellular infection (hrpGA179V) in the final R16 clone [20]. Searching for the modification that countered the adaptive effect of the hrpGA179V mutation led us to investigate the role of a gene deleted in the symbiotic plasmid of R16 and whose reintroduction restored nodule cell infection capacity [164]. We found that this gene, now called noeM, is a host-specificity nodulation gene responsible for a Nod factor modification that was previously missed, the opening and oxidation of the reducing end of the molecule [165]. Surprisingly the inactivation of noeM did not affect the nodule cell infection capacity but only the nodulation competitiveness of its natural host C. taiwanensis in interaction with M. pudica, indicating that its impact on symbiosis depends on the genomic background.
7. Conclusions
The evolution of nitrogen-fixing root nodule symbioses was a key innovation that gave access to new ecological niches to both plant (nitrogen-depleted soils) and bacterial (root nodules) symbionts. Comparative genomic and phylogenomic approaches have greatly contributed to understanding the evolution of these complex interactions [7,8]. By using EE on the bacterial partner, we were able to complement these approaches and provide a dynamic view of evolution, although in a somewhat artificial context, since it is impossible to know and reproduce the environmental conditions and protagonists that took part in the emergence of this symbiosis. EE did not address the question of the origin of the first rhizobia, nor that of the co-evolution between plants and rhizobia. Instead, it tackled the issue of the spreading of the N2-fixing capacity in many proteobacterial branches. One key lesson was that a bacterium with a completely different lifestyle (pathogenic, extracellular) can be turned into an intracellular legume symbiont via the acquisition of mutualistic genes and regulatory rewiring under plant selection pressure. This result helps us to understand why so many bacterial branches have successfully been colonized by nitrogen-fixing traits. So far, our work focused on the first symbiotic stages: acquisition and improvement of nodulation and nodule invasion. By continuing the evolution experiment and its genetic analysis, future work should address the evolution of late stages of symbiosis, such as the acquisition of persistence, nitrogen fixation and mutualism and should provide a better understanding of these poorly known symbiotic stages. EE including a single selection cycle also proved to be very useful in understanding the bacterial genetic bases of incompatible symbiotic interactions [47,166,167] and to analyze plant-mediated selection acting on rhizobial communities [168,169]. Complementary to synthetic biology and classical genetic approaches, EE will undoubtedly become an indispensable tool to select efficient, host-adapted microsymbionts in the perspective of optimizing current symbiotic associations or transferring N2-fixation in crops [170,171].
Author Contributions
G.G.D.d.M., P.R., C.M.-B., and D.C. wrote the paper. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the French National Research Agency (ANR-16-CE20-0011-01) and the Laboratoire d’Excellence (LABEX) TULIP (ANR-10-LABX-41). G.G.D.d.M. was supported by a fellowship from the French Ministère de l’Enseignement Supérieur, de la Recherche et de l’Innovation (MESRI). P.R. was funded by the European Union’s Horizon 2020 research and innovation programme under the Marie Skłodowska Curie grant agreement No 845838.
Acknowledgments
We thank Finlay Warsop Thomas for careful reading of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Yang, L.; Jelsbak, L.; Marvig, R.L.; Damkiaer, S.; Workman, C.T.; Rau, M.H.; Hansen, S.K.; Folkesson, A.; Johansen, H.K.; Ciofu, O.; et al. Evolutionary dynamics of bacteria in a human host environment. Proc. Natl. Acad. Sci. USA 2011, 108, 7481–7486. [Google Scholar] [CrossRef] [Green Version]
- Zhao, S.; Lieberman, T.D.; Poyet, M.; Kauffman, K.M.; Gibbons, S.M.; Groussin, M.; Xavier, R.J.; Alm, E.J. Adaptive evolution within gut microbiomes of healthy people. Cell Host Microbe 2019, 25, 656–667. [Google Scholar] [CrossRef]
- Garud, N.R.; Good, B.H.; Hallatschek, O.; Pollard, K.S. Evolutionary dynamics of bacteria in the gut microbiome within and across hosts. PLoS Biol. 2019, 17, e3000102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooper, V.S. Experimental evolution as a high-throughput screen for genetic adaptations. mSphere 2018, 3, e00121-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elena, S.; Lenski, R. Evolution experiments with microorganisms: The dynamics and genetic bases of adaptation. Nat. Rev. Genet. 2003, 4, 457–469. [Google Scholar] [CrossRef] [PubMed]
- Van den Bergh, B.; Swings, T.; Fauvart, M.; Michiels, J. Experimental design, population dynamics, and diversity in microbial experimental evolution. Microbiol. Mol. Biol. Rev. 2018, 82, e00008-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Velzen, R.; Doyle, J.J.; Geurts, R. A resurrected scenario: Single gain and massive loss of nitrogen-fixing nodulation. Trends Plant Sci. 2019, 24, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Griesmann, M.; Chang, Y.; Liu, X.; Song, Y.; Haberer, G.; Crook, M.B.; Billault-Penneteau, B.; Lauressergues, D.; Keller, J.; Imanishi, L.; et al. Phylogenomics reveals multiple losses of nitrogen-fixing root nodule symbiosis. Science 2018, 361, 144. [Google Scholar] [CrossRef] [Green Version]
- Ktari, A.; Nouioui, I.; Furnholm, T.; Swanson, E.; Ghodhbane-Gtari, F.; Tisa, L.S.; Gtari, M. Permanent draft genome sequence of Frankia sp. NRRL B-16219 reveals the presence of canonical nod genes, which are highly homologous to those detected in Candidatus Frankia Dg1 genome. Stand. Genom. Sci. 2017, 12, 51. [Google Scholar] [CrossRef] [Green Version]
- Persson, T.; Battenberg, K.; Demina, I.V.; Vigil-Stenman, T.; Vanden Heuvel, B.; Pujic, P.; Facciotti, M.T.; Wilbanks, E.G.; O’Brien, A.; Fournier, P.; et al. Candidatus Frankia datiscae Dg1, the actinobacterial microsymbiont of Datisca glomerata, expresses the canonical nod genes nodABC in symbiosis with its host plant. PLoS ONE 2015, 10, e0127630. [Google Scholar] [CrossRef] [Green Version]
- Andrews, M.; De Meyer, S.; James, E.K.; Stępkowski, T.; Hodge, S.; Simon, M.F.; Young, J.P.W. Horizontal transfer of symbiosis genes within and between rhizobial genera: Occurrence and importance. Genes (Basel) 2018, 9, 321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacLean, A.M.; Finan, T.M.; Sadowsky, M.J. Genomes of the symbiotic nitrogen-fixing bacteria of legumes. Plant Physiol. 2007, 144, 615–622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Lajudie, P.; Young, J.P.W. International committee on systematics of prokaryotes subcommittee on the taxonomy of rhizobia and agrobacteria minutes of the meeting by video conference, 11 July 2018. Int. J. Syst. Evol. Microbiol. 2019, 69, 1835–1840. [Google Scholar] [CrossRef]
- Sprent, J.I.; James, E.K. Legume evolution: Where do nodules and mycorrhizas fit in? Plant Physiol. 2007, 144, 575–581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bontemps, C.; Elliott, G.N.; Simon, M.F.; Dos Reis Junior, F.B.; Gross, E.; Lawton, R.C.; Neto, N.E.; Loureiro, M.D.F.; De Faria, S.M.; Sprent, J.I.; et al. Burkholderia species are ancient symbionts of legumes. Mol. Ecol. 2010, 19, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Sprent, J.I. 60Ma of legume nodulation. What’s new? What’s changing? J. Exp. Bot. 2008, 59, 1081–1084. [Google Scholar] [CrossRef] [PubMed]
- Clerissi, C.; Touchon, M.; Capela, D.; Tang, M.; Cruveiller, S.; Parker, M.A.; Moulin, L.; Masson-Boivin, C.; Rocha, E.P.C. Parallels between experimental and natural evolution of legume symbionts. Nat. Commun. 2018, 9, 2264. [Google Scholar] [CrossRef]
- Mishra, R.P.N.; Tisseyre, P.; Melkonian, R.; Chaintreuil, C.; Miche, L.; Klonowska, A.; Gonzalez, S.; Bena, G.; Laguerre, G.; Moulin, L. Genetic diversity of Mimosa pudica rhizobial symbionts in soils of French Guiana: Investigating the origin and diversity of Burkholderia phymatum and other beta-rhizobia. FEMS Microbiol. Ecol. 2012, 79, 487–503. [Google Scholar] [CrossRef]
- Marchetti, M.; Jauneau, A.; Capela, D.; Remigi, P.; Gris, C.; Batut, J.; Masson-Boivin, C. Shaping bacterial symbiosis with legumes by experimental evolution. Mol. Plant-Microbe Interact. 2014, 27, 956–964. [Google Scholar] [CrossRef] [Green Version]
- Marchetti, M.; Clerissi, C.; Yousfi, Y.; Gris, C.; Bouchez, O.; Rocha, E.; Cruveiller, S.; Jauneau, A.; Capela, D.; Masson-Boivin, C. Experimental evolution of rhizobia may lead to either extra- or intracellular symbiotic adaptation depending on the selection regime. Mol. Ecol. 2017, 26, 1818–1831. [Google Scholar] [CrossRef] [Green Version]
- Marchetti, M.; Capela, D.; Glew, M.; Cruveiller, S.; Chane-Woon-Ming, B.; Gris, C.; Timmers, T.; Poinsot, V.; Gilbert, L.B.; Heeb, P.; et al. Experimental evolution of a plant pathogen into a legume symbiont. PLoS Biol. 2010, 8, e1000280. [Google Scholar] [CrossRef] [PubMed]
- Guan, S.H.; Gris, C.; Cruveiller, S.; Pouzet, C.; Tasse, L.; Leru, A.; Maillard, A.; Medigue, C.; Batut, J.; Masson-Boivin, C.; et al. Experimental evolution of nodule intracellular infection in legume symbionts. ISME J. 2013, 7, 1367–1377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capela, D.; Marchetti, M.; Clérissi, C.; Perrier, A.; Guetta, D.; Gris, C.; Valls, M.; Jauneau, A.; Cruveiller, S.; Rocha, E.P.C.; et al. Recruitment of a lineage-specific virulence regulatory pathway promotes intracellular infection by a plant pathogen experimentally evolved into a legume symbiont. Mol. Biol. Evol. 2017, 34, 2503–2521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, M.; Bouchez, O.; Cruveiller, S.; Masson-Boivin, C.; Capela, D. Modulation of quorum sensing as an adaptation to nodule cell infection during experimental evolution of legume symbionts. mBio 2020, 11, e03129-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sullivan, J.T.; Patrick, H.N.; Lowther, W.L.; Scott, D.B.; Ronson, C.W. Nodulating strains of Rhizobium loti arise through chromosomal symbiotic gene transfer in the environment. Proc. Natl. Acad. Sci. USA 1995, 92, 8985–8989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sullivan, J.; Ronson, C. Evolution of rhizobia by acquisition of a 500-kb symbiosis island that integrates into a phe-tRNA gene. Proc. Natl. Acad. Sci. USA 1998, 95, 5145–5149. [Google Scholar] [CrossRef] [Green Version]
- Suominen, L.; Roos, C.; Lortet, G.; Paulin, L.; Lindström, K. Identification and structure of the Rhizobium galegae common nodulation genes: Evidence for horizontal gene transfer. Mol. Biol. Evol. 2001, 18, 907–916. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.M.; Moulin, L.; Bontemps, C.; Vandamme, P.; Bena, G.; Boivin-Masson, C. Legume symbiotic nitrogen fixation by beta-proteobacteria is widespread in nature. J. Bacteriol. 2003, 185, 7266–7272. [Google Scholar] [CrossRef] [Green Version]
- Nandasena, K.G.; O’Hara, G.W.; Tiwari, R.P.; Sezmis, E.; Howieson, J.G. In situ lateral transfer of symbiosis islands results in rapid evolution of diverse competitive strains of mesorhizobia suboptimal in symbiotic nitrogen fixation on the pasture legume Biserrula pelecinus L. Environ. Microbiol. 2007, 9, 2496–2511. [Google Scholar] [CrossRef]
- Barcellos, F.G.; Menna, P.; Batista, J.S.D.; Hungria, M. Evidence of horizontal transfer of symbiotic genes from a Bradyrhizobium japonicum inoculant strain to indigenous diazotrophs Sinorhizobium (Ensifer) fredii and Bradyrhizobium elkanii in a Brazilian Savannah soil. Appl. Environ. Microbiol. 2007, 73, 2635–2643. [Google Scholar] [CrossRef] [Green Version]
- Downie, J.A. The roles of extracellular proteins, polysaccharides and signals in the interactions of rhizobia with legume roots. FEMS Microbiol. Rev. 2010, 34, 150–170. [Google Scholar] [CrossRef] [PubMed]
- Fraysse, N.; Couderc, F.; Poinsot, V. Surface polysaccharide involvement in establishing the rhizobium-legume symbiosis. Eur. J. Biochem. 2003, 270, 1365–1380. [Google Scholar] [CrossRef]
- Tian, C.F.; Zhou, Y.J.; Zhang, Y.M.; Li, Q.Q.; Zhang, Y.Z.; Li, D.F.; Wang, S.; Wang, J.; Gilbert, L.B.; Li, Y.R.; et al. Comparative genomics of rhizobia nodulating soybean suggests extensive recruitment of lineage-specific genes in adaptations. Proc. Natl. Acad. Sci. USA 2012, 109, 8629–8634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Aono, T.; Poole, P.; Finan, T.M. NAD(P)+-malic enzyme mutants of Sinorhizobium sp. strain NGR234, but not Azorhizobium caulinodans ORS571, maintain symbiotic N2 fixation capabilities. Appl. Environ. Microbiol. 2012, 78, 2803–2812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yurgel, S.N.; Rice, J.; Kahn, M.L. Transcriptome analysis of the role of GlnD/GlnBK in nitrogen stress adaptation by Sinorhizobium meliloti Rm1021. PLoS ONE 2013, 8, e58028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batista, S.; Catalán, A.I.; Hernández-Lucas, I.; Martínez-Romero, E.; Aguilar, O.M.; Martínez-Drets, G. Identification of a system that allows a Rhizobium tropici dctA mutant to grow on succinate, but not on other C4-dicarboxylates. Can. J. Microbiol. 2001, 47, 509–518. [Google Scholar] [CrossRef]
- Jiao, J.; Wu, L.J.; Zhang, B.; Hu, Y.; Li, Y.; Zhang, X.X.; Guo, H.J.; Liu, L.X.; Chen, W.X.; Zhang, Z.; et al. MucR is required for transcriptional activation of conserved ion transporters to support nitrogen fixation of Sinorhizobium fredii in soybean nodules. Mol. Plant Microbe Interact. 2016, 29, 352–361. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Jiao, J.; Liu, L.X.; Sun, Y.W.; Chen, W.F.; Sui, X.H.; Chen, W.X.; Tian, C.F. Evidence for phosphate starvation of rhizobia without terminal differentiation in legume nodules. Mol. Plant Microbe Interact. 2018, 31, 1060–1068. [Google Scholar] [CrossRef] [Green Version]
- Jiao, J.; Ni, M.; Zhang, B.; Zhang, Z.; Young, J.P.W.; Chan, T.F.; Chen, W.X.; Lam, H.M.; Tian, C.F. Coordinated regulation of core and accessory genes in the multipartite genome of Sinorhizobium fredii. PLoS Genet. 2018, 14, e1007428. [Google Scholar] [CrossRef]
- Bittner, A.N.; Foltz, A.; Oke, V. Only one of five groEL genes is required for viability and successful symbiosis in Sinorhizobium meliloti. J. Bacteriol. 2007, 189, 1884–1889. [Google Scholar] [CrossRef] [Green Version]
- Nelson, M.S.; Sadowsky, M.J. Secretion systems and signal exchange between nitrogen-fixing rhizobia and legumes. Front. Plant Sci. 2015, 6, 491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Black, M.; Moolhuijzen, P.; Chapman, B.; Barrero, R.; Howieson, J.; Hungria, M.; Bellgard, M. The genetics of symbiotic nitrogen fixation: Comparative genomics of 14 rhizobia strains by resolution of protein clusters. Genes (Basel) 2012, 3, 138–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pobigaylo, N.; Szymczak, S.; Nattkemper, T.W.; Becker, A. Identification of genes relevant to symbiosis and competitiveness in Sinorhizobium meliloti using signature-tagged mutants. Mol. Plant Microbe Interact. 2008, 21, 219–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flores-Tinoco, C.E.; Christen, M.; Christen, B. Co-catabolism of arginine and succinate drives symbiotic nitrogen fixation. BioRxiv 2019. [Google Scholar] [CrossRef]
- Saad, M.M.; Crèvecoeur, M.; Masson-Boivin, C.; Perret, X. The type 3 protein secretion system of Cupriavidus taiwanensis strain LMG19424 compromises symbiosis with Leucaena leucocephala. Appl. Environ. Microbiol. 2012, 78, 7476–7479. [Google Scholar] [CrossRef] [Green Version]
- Viprey, V.; Del Greco, A.; Golinowski, W.; Broughton, W.J.; Perret, X. Symbiotic implications of type III protein secretion machinery in Rhizobium. Mol. Microbiol. 1998, 28, 1381–1389. [Google Scholar] [CrossRef]
- Zhao, R.; Liu, L.X.; Zhang, Y.Z.; Jiao, J.; Cui, W.J.; Zhang, B.; Wang, X.L.; Li, M.L.; Chen, Y.; Xiong, Z.Q.; et al. Adaptive evolution of rhizobial symbiotic compatibility mediated by co-evolved insertion sequences. ISME J. 2018, 12, 101–111. [Google Scholar] [CrossRef] [Green Version]
- Hirsch, A.M.; Wilson, K.J.; Jones, J.D.; Bang, M.; Walker, V.V.; Ausubel, F.M. Rhizobium meliloti nodulation genes allow Agrobacterium tumefaciens and Escherichia coli to form pseudonodules on alfalfa. J. Bacteriol. 1984, 158, 1133–1143. [Google Scholar] [CrossRef] [Green Version]
- Abe, M.; Kawamura, R.; Higashi, S.; Mori, S.; Shibata, M.; Uchiumi, T. Transfer of the symbiotic plasmid from Rhizobium leguminosarum biovar trifolii to Agrobacterium tumefaciens. J. Gen. Appl. Microbiol. 1998, 44, 65–74. [Google Scholar] [CrossRef] [Green Version]
- Nakatsukasa, H.; Uchiumi, T.; Kucho, K.; Suzuki, A.; Higashi, S.; Abe, M. Transposon mediation allows a symbiotic plasmid of Rhizobium leguminosarum bv. trifolii to become a symbiosis island in Agrobacterium and Rhizobium. J. Gen. Appl. Microbiol. 2008, 54, 107–118. [Google Scholar] [CrossRef] [Green Version]
- Martinez, E.; Palacios, R.; Sanchez, F. Nitrogen fixing nodules induced by Agrobacterium tumefaciens harboring Rhizobium phaseoli plasmids. J. Bacteriol. 1987, 169, 2828–2834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benezech, C.; Doudement, M.; Gourion, B. Legumes tolerance to rhizobia is not always observed and not always deserved. Cell. Microbiol. 2020, 22, e13124. [Google Scholar] [CrossRef] [PubMed]
- Limpens, E.; Franken, C.; Smit, P.; Willemse, J.; Bisseling, T.; Geurts, R. LysM domain receptor kinases regulating rhizobial Nod factor-induced infection. Science 2003, 302, 630–633. [Google Scholar] [CrossRef] [PubMed]
- Madsen, E.B.; Madsen, L.H.; Radutoiu, S.; Olbryt, M.; Rakwalska, M.; Szczyglowski, K.; Sato, S.; Kaneko, T.; Tabata, S.; Sandal, N.; et al. A receptor kinase gene of the LysM type is involved in legume perception of rhizobial signals. Nature 2003, 425, 637–640. [Google Scholar] [CrossRef]
- Radutoiu, S.; Madsen, L.H.; Madsen, E.B.; Felle, H.H.; Umehara, Y.; Grønlund, M.; Sato, S.; Nakamura, Y.; Tabata, S.; Sandal, N.; et al. Plant recognition of symbiotic bacteria requires two LysM receptor-like kinases. Nature 2003, 425, 585–592. [Google Scholar] [CrossRef]
- Arrighi, J.F.; Barre, A.; Ben Amor, B.; Bersoult, A.; Soriano, L.C.; Mirabella, R.; de Carvalho-Niebel, F.; Journet, E.P.; Ghérardi, M.; Huguet, T.; et al. The Medicago truncatula lysin motif-receptor-like kinase gene family includes NFP and new nodule-expressed genes. Plant Physiol. 2006, 142, 265–279. [Google Scholar] [CrossRef] [Green Version]
- Mulder, L.; Lefebvre, B.; Cullimore, J.; Imberty, A. LysM domains of Medicago truncatula NFP protein involved in Nod factor perception. Glycosylation state, molecular modeling and docking of chitooligosaccharides and Nod factors. Glycobiology 2006, 16, 801–809. [Google Scholar] [CrossRef] [Green Version]
- Smit, P.; Limpens, E.; Geurts, R.; Fedorova, E.; Dolgikh, E.; Gough, C.; Bisseling, T. Medicago LYK3, an entry receptor in rhizobial nodulation factor signaling. Plant Physiol. 2007, 145, 183–191. [Google Scholar] [CrossRef] [Green Version]
- Kawaharada, Y.; Kelly, S.; Nielsen, M.W.; Hjuler, C.T.; Gysel, K.; Muszyński, A.; Carlson, R.W.; Thygesen, M.B.; Sandal, N.; Asmussen, M.H.; et al. Receptor-mediated exopolysaccharide perception controls bacterial infection. Nature 2015, 523, 308–312. [Google Scholar] [CrossRef] [Green Version]
- Daubech, B.; Remigi, P.; Doin de Moura, G.; Marchetti, M.; Pouzet, C.; Auriac, M.C.; Gokhale, C.S.; Masson-Boivin, C.; Capela, D. Spatio-temporal control of mutualism in legumes helps spread symbiotic nitrogen fixation. Elife 2017, 6. [Google Scholar] [CrossRef] [Green Version]
- Kiers, E.; Rousseau, R.; West, S.; Denison, R. Host sanctions and the legume-rhizobium mutualism. Nature 2003, 425, 78–81. [Google Scholar] [CrossRef] [PubMed]
- Oono, R.; Anderson, C.G.; Denison, R.F. Failure to fix nitrogen by non-reproductive symbiotic rhizobia triggers host sanctions that reduce fitness of their reproductive clonemates. Proc. Biol. Sci. 2011, 278, 2698–2703. [Google Scholar] [CrossRef] [Green Version]
- Gehlot, H.S.; Tak, N.; Kaushik, M.; Mitra, S.; Chen, W.-M.; Poweleit, N.; Panwar, D.; Poonar, N.; Parihar, R.; Tak, A.; et al. An invasive Mimosa in India does not adopt the symbiont of its native relatives. Ann. Bot. 2013, 112, 179–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gossmann, J.A.; Markmann, K.; Brachmann, A.; Rose, L.E.; Parniske, M. Polymorphic infection and organogenesis patterns induced by a Rhizobium leguminosarum isolate from Lotus root nodules are determined by the host genotype. New Phytol. 2012, 196, 561–573. [Google Scholar] [CrossRef] [PubMed]
- Werner, G.D.A.; Cornwell, W.K.; Sprent, J.I.; Kattge, J.; Kiers, E.T. A single evolutionary innovation drives the deep evolution of symbiotic N2-fixation in angiosperms. Nat. Commun. 2014, 5, 4087. [Google Scholar] [CrossRef] [Green Version]
- Turner, S.L.; Young, J.P.W. The glutamine synthetases of rhizobia: Phylogenetics and evolutionary implications. Mol. Biol. Evol. 2000, 17, 309–319. [Google Scholar] [CrossRef] [Green Version]
- Marin, J.; Battistuzzi, F.U.; Brown, A.C.; Hedges, S.B. The timetree of prokaryotes: New insights into their evolution and speciation. Mol. Biol. Evol. 2017, 34, 437–446. [Google Scholar] [CrossRef]
- Garrido-Oter, R.; Nakano, R.T.; Dombrowski, N.; Ma, K.W.; McHardy, A.C.; Schulze-Lefert, P.; Team, A. Modular traits of the Rhizobiales root microbiota and their evolutionary relationship with symbiotic rhizobia. Cell Host Microbe 2018, 24, 155–167. [Google Scholar] [CrossRef] [Green Version]
- Salas, M.E.; Lozano, M.J.; López, J.L.; Draghi, W.O.; Serrania, J.; Torres Tejerizo, G.A.; Albicoro, F.J.; Nilsson, J.F.; Pistorio, M.; Del Papa, M.F.; et al. Specificity traits consistent with legume-rhizobia coevolution displayed by Ensifer meliloti rhizosphere colonization. Environ. Microbiol. 2017, 19, 3423–3438. [Google Scholar] [CrossRef] [Green Version]
- Ding, H.; Yip, C.B.; Geddes, B.A.; Oresnik, I.J.; Hynes, M.F. Glycerol utilization by Rhizobium leguminosarum requires an ABC transporter and affects competition for nodulation. Microbiology 2012, 158, 1369–1378. [Google Scholar] [CrossRef]
- Garcia-Fraile, P.; Seaman, J.C.; Karunakaran, R.; Edwards, A.; Poole, P.S.; Downie, J.A. Arabinose and protocatechuate catabolism genes are important for growth of Rhizobium leguminosarum biovar viciae in the pea rhizosphere. Plant Soil 2015, 390, 251–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gourion, B.; Berrabah, F.; Ratet, P.; Stacey, G. Rhizobium-legume symbioses: The crucial role of plant immunity. Trends Plant Sci. 2015, 20, 186–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berrabah, F.; Bourcy, M.; Eschstruth, A.; Cayrel, A.; Guefrachi, I.; Mergaert, P.; Wen, J.; Jean, V.; Mysore, K.S.; Gourion, B.; et al. A nonRD receptor-like kinase prevents nodule early senescence and defense-like reactions during symbiosis. New Phytol. 2014, 203, 1305–1314. [Google Scholar] [CrossRef] [PubMed]
- Genin, S.; Denny, T.P. Pathogenomics of the Ralstonia solanacearum species complex. Ann. Rev. Phytopathol. 2012, 50, 67–89. [Google Scholar] [CrossRef] [PubMed]
- Vailleau, F.; Sartorel, E.; Jardinaud, M.F.; Chardon, F.; Genin, S.; Huguet, T.; Gentzbittel, L.; Petitprez, M. Characterization of the interaction between the bacterial wilt pathogen Ralstonia solanacearum and the model legume plant Medicago truncatula. Mol. Plant Microbe Interact. 2007, 20, 159–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sachs, J.L.; Skophammer, R.G.; Bansal, N.; Stajich, J.E. Evolutionary origins and diversification of proteobacterial mutualists. Proc. Biol. Sci. 2014, 281, 20132146. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.M.; Laevens, S.; Lee, T.M.; Coenye, T.; De Vos, P.; Mergeay, M.; Vandamme, P. Ralstonia taiwanensis sp nov., isolated from root nodules of Mimosa species and sputum of a cystic fibrosis patient. Int. J. Syst. Evol. Microbiol. 2001, 51, 1729–1735. [Google Scholar] [CrossRef] [Green Version]
- Amadou, C.; Pascal, G.; Mangenot, S.; Glew, M.; Bontemps, C.; Capela, D.; Carrere, S.; Cruveiller, S.; Dossat, C.; Lajus, A.; et al. Genome sequence of the beta-rhizobium Cupriavidus taiwanensis and comparative genomics of rhizobia. Genome Res. 2008, 18, 1472–1483. [Google Scholar] [CrossRef] [Green Version]
- Shapiro, J.W.; Turner, P.E. Evolution of mutualism from parasitism in experimental virus populations. Evolution 2018, 72, 707–712. [Google Scholar] [CrossRef]
- King, K.C.; Brockhurst, M.A.; Vasieva, O.; Paterson, S.; Betts, A.; Ford, S.A.; Frost, C.L.; Horsburgh, M.J.; Haldenby, S.; Hurst, G.D. Rapid evolution of microbe-mediated protection against pathogens in a worm host. ISME J. 2016, 10, 1915–1924. [Google Scholar] [CrossRef] [Green Version]
- Tso, G.H.W.; Reales-Calderon, J.A.; Tan, A.S.M.; Sem, X.; Le, G.T.T.; Tan, T.G.; Lai, G.C.; Srinivasan, K.G.; Yurieva, M.; Liao, W.; et al. Experimental evolution of a fungal pathogen into a gut symbiont. Science 2018, 362, 589–595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bañuelos-Vazquez, L.A.; Torres Tejerizo, G.; Brom, S. Regulation of conjugative transfer of plasmids and integrative conjugative elements. Plasmid 2017, 91, 82–89. [Google Scholar] [CrossRef] [PubMed]
- Ding, H.; Hynes, M.F. Plasmid transfer systems in the rhizobia. Can. J. Microbiol. 2009, 55, 917–927. [Google Scholar] [CrossRef] [PubMed]
- Ramsay, J.P.; Tester, L.G.L.; Major, A.S.; Sullivan, J.T.; Edgar, C.D.; Kleffmann, T.; Patterson-House, J.R.; Hall, D.A.; Tate, W.P.; Hynes, M.F.; et al. Ribosomal frameshifting and dual-target antiactivation restrict quorum-sensing-activated transfer of a mobile genetic element. Proc. Natl. Acad. Sci. USA 2015, 112, 4104–4109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ling, J.; Wang, H.; Wu, P.; Li, T.; Tang, Y.; Naseer, N.; Zheng, H.; Masson-Boivin, C.; Zhong, Z.; Zhu, J. Plant nodulation inducers enhance horizontal gene transfer of Azorhizobium caulinodans symbiosis island. Proc. Natl. Acad. Sci. USA 2016, 113, 13875–13880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valls, M.; Genin, S.; Boucher, C. Integrated regulation of the type III secretion system and other virulence determinants in Ralstonia solanacearum. PLoS Pathog. 2006, 2, 798–807. [Google Scholar] [CrossRef] [Green Version]
- Perrier, A.; Peyraud, R.; Rengel, D.; Barlet, X.; Lucasson, E.; Gouzy, J.; Peeters, N.; Genin, S.; Guidot, A. Enhanced in planta fitness through adaptive mutations in efpR, a dual regulator of virulence and metabolic functions in the plant pathogen Ralstonia solanacearum. PLoS Pathog. 2016, 12, e1006044. [Google Scholar] [CrossRef]
- Peyraud, R.; Cottret, L.; Marmiesse, L.; Gouzy, J.; Genin, S. A Resource allocation trade-off between virulence and proliferation drives metabolic versatility in the plant pathogen Ralstonia solanacearum. PLoS Pathog. 2016, 12, e1005939. [Google Scholar] [CrossRef] [Green Version]
- Perrier, A.; Barlet, X.; Peyraud, R.; Rengel, D.; Guidot, A.; Genin, S. Comparative transcriptomic studies identify specific expression patterns of virulence factors under the control of the master regulator PhcA in the Ralstonia solanacearum species complex. Microb. Pathog. 2018, 116, 273–278. [Google Scholar] [CrossRef]
- Mori, Y.; Ishikawa, S.; Ohnishi, H.; Shimatani, M.; Morikawa, Y.; Hayashi, K.; Ohnishi, K.; Kiba, A.; Kai, K.; Hikichi, Y. Involvement of ralfuranones in the quorum sensing signalling pathway and virulence of Ralstonia solanacearum strain OE1-1. Mol. Plant Pathol. 2018, 19, 454–463. [Google Scholar] [CrossRef] [Green Version]
- Perrier, A.; Barlet, X.; Rengel, D.; Prior, P.; Poussier, S.; Genin, S.; Guidot, A. Spontaneous mutations in a regulatory gene induce phenotypic heterogeneity and adaptation of Ralstonia solanacearum to changing environments. Environ. Microbiol. 2019, 21, 3140–3152. [Google Scholar] [CrossRef] [PubMed]
- Khokhani, D.; Lowe-Power, T.M.; Tran, T.M.; Allen, C. A Single regulator mediates strategic switching between attachment/spread and growth/virulence in the plant pathogen. mBio 2017, 8, e00895-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Philippe, N.; Crozat, E.; Lenski, R.E.; Schneider, D. Evolution of global regulatory networks during a long-term experiment with Esherichia coli. Bioessays 2007, 29, 846–860. [Google Scholar] [CrossRef] [PubMed]
- Hindré, T.; Knibbe, C.; Beslon, G.; Schneider, D. New insights into bacterial adaptation through in vivo and in silico experimental evolution. Nat. Rev. Microbiol. 2012, 10, 352–365. [Google Scholar] [CrossRef]
- Pankey, S.M.; Foxall, R.L.; Ster, I.M.; Perry, L.A.; Schuster, B.M.; Donner, R.A.; Coyle, M.; Cooper, V.S.; Whistler, C.A. Host-selected mutations converging on a global regulator drive an adaptive leap towards symbiosis in bacteria. Elife 2017, 6, e24414. [Google Scholar] [CrossRef]
- Carroll, S.M.; Chubiz, L.M.; Agashe, D.; Marx, C.J. Parallel and divergent evolutionary solutions for the optimization of an engineered central metabolism in Methylobacterium extorquens AM1. Microorganisms 2015, 3, 152–174. [Google Scholar] [CrossRef]
- Damkiær, S.; Yang, L.; Molin, S.; Jelsbak, L. Evolutionary remodeling of global regulatory networks during long-term bacterial adaptation to human hosts. Proc. Natl. Acad. Sci. USA 2013, 110, 7766–7771. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Verdugo, A.; Tenaillon, O.; Gaut, B.S. First-Step Mutations during adaptation restore the expression of hundreds of genes. Mol. Biol. Evol. 2016, 33, 25–39. [Google Scholar] [CrossRef] [Green Version]
- Loh, J.; Stacey, G. Nodulation gene regulation in Bradyrhizobium japonicum: A unique integration of global regulatory circuits. Appl. Environ. Microbiol. 2003, 69, 10–17. [Google Scholar] [CrossRef] [Green Version]
- Del Cerro, P.; Rolla-Santos, A.A.; Gomes, D.F.; Marks, B.B.; del Rosario Espuny, M.; Rodríguez-Carvajal, M.; Soria-Díaz, M.E.; Nakatani, A.S.; Hungria, M.; Ollero, F.J.; et al. Opening the black box of nodD3, nodD4 and nodD5 genes of Rhizobium tropici strain CIAT 899. BMC Genom. 2015, 16, 864. [Google Scholar] [CrossRef]
- Kobayashi, H.; Graven, Y.N.; Broughton, W.J.; Perret, X. Flavonoids induce temporal shifts in gene-expression of nod-box controlled loci in Rhizobium sp NGR234. Mol. Microbiol. 2004, 51, 335–347. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Gao, K.; Kondorosi, E.; Kondorosi, A.; Rolfe, B.G. Functional genomic analysis of global regulator NolR in Sinorhizobium meliloti. Mol. Plant Microbe Interact. 2005, 18, 1340–1352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Del Cerro, P.; Rolla-Santos, A.A.; Valderrama-Fernández, R.; Gil-Serrano, A.; Bellogín, R.A.; Gomes, D.F.; Pérez-Montaño, F.; Megías, M.; Hungría, M.; Ollero, F.J. NrcR, a New transcriptional regulator of Rhizobium tropici CIAT 899 involved in the legume root-nodule symbiosis. PLoS ONE 2016, 11, e0154029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bauer, E.; Kaspar, T.; Fischer, H.M.; Hennecke, H. Expression of the fixR-nifA operon in Bradyrhizobium japonicum depends on a new response regulator, RegR. J. Bacteriol. 1998, 180, 3853–3863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaminski, P.A.; Elmerich, C. The control of Azorhizobium caulinodans nifA expression by oxygen, ammonia and by the HF-I-like protein, NrfA. Mol. Microbiol. 1998, 28, 603–613. [Google Scholar] [CrossRef]
- Cosseau, C.; Batut, J. Genomics of the ccoNOQP-encoded cbb(3) oxidase complex in bacteria. Arch. Microbiol. 2004, 181, 89–96. [Google Scholar] [CrossRef]
- Fenner, B.J.; Tiwari, R.P.; Reeve, W.G.; Dilworth, M.J.; Glenn, A.R. Sinorhizobium medicae genes whose regulation involves the ActS and/or ActR signal transduction proteins. FEMS Microbiol. Lett. 2004, 236, 21–31. [Google Scholar] [CrossRef]
- Moris, M.; Dombrecht, B.; Xi, C.; Vanderleyden, J.; Michiels, J. Regulatory role of Rhizobium etli CNPAF512 fnrN during symbiosis. Appl. Environ. Microbiol. 2004, 70, 1287–1296. [Google Scholar] [CrossRef] [Green Version]
- Reyes-González, A.; Talbi, C.; Rodríguez, S.; Rivera, P.; Zamorano-Sánchez, D.; Girard, L. Expanding the regulatory network that controls nitrogen fixation in Sinorhizobium meliloti: Elucidating the role of the two-component system hFixL-FxkR. Microbiology 2016, 162, 979–988. [Google Scholar] [CrossRef]
- Mesa, S.; Hauser, F.; Friberg, M.; Malaguti, E.; Fischer, H.M.; Hennecke, H. Comprehensive assessment of the regulons controlled by the FixLJ-FixK2-FixK1 cascade in Bradyrhizobium japonicum. J. Bacteriol. 2008, 190, 6568–6579. [Google Scholar] [CrossRef] [Green Version]
- Janczarek, M. Environmental signals and regulatory pathways that influence exopolysaccharide production in rhizobia. Int. J. Mol. Sci. 2011, 12, 7898–7933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonomi, H.R.; Posadas, D.M.; Paris, G.; Carrica, M.e.C.; Frederickson, M.; Pietrasanta, L.I.; Bogomolni, R.A.; Zorreguieta, A.; Goldbaum, F.A. Light regulates attachment, exopolysaccharide production, and nodulation in Rhizobium leguminosarum through a LOV-histidine kinase photoreceptor. Proc. Natl. Acad. Sci. USA 2012, 109, 12135–12140. [Google Scholar] [CrossRef] [Green Version]
- Lemaire, B.; Van Cauwenberghe, J.; Chimphango, S.; Stirton, C.; Honnay, O.; Smets, E.; Muasya, A.M. Recombination and horizontal transfer of nodulation and ACC deaminase (acdS) genes within Alpha- and Betaproteobacteria nodulating legumes of the Cape Fynbos biome. FEMS Microbiol. Ecol. 2015, 91, fiv118. [Google Scholar] [CrossRef] [Green Version]
- Pérez Carrascal, O.M.; VanInsberghe, D.; Juárez, S.; Polz, M.F.; Vinuesa, P.; González, V. Population genomics of the symbiotic plasmids of sympatric nitrogen-fixing Rhizobium species associated with Phaseolus vulgaris. Environ. Microbiol. 2016, 18, 2660–2676. [Google Scholar] [CrossRef] [PubMed]
- Bañuelos-Vazquez, L.A.; Torres Tejerizo, G.; Cervantes-De La Luz, L.; Girard, L.; Romero, D.; Brom, S. Conjugative transfer between Rhizobium etli endosymbionts inside the root nodule. Environ. Microbiol. 2019, 21, 3430–3441. [Google Scholar] [CrossRef] [PubMed]
- Zgadzaj, R.; James, E.K.; Kelly, S.; Kawaharada, Y.; de Jonge, N.; Jensen, D.B.; Madsen, L.H.; Radutoiu, S. A Legume genetic framework controls infection of nodules by symbiotic and endophytic bacteria. PLoS Genet. 2015, 11, e1005280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-Hidalgo, P.; Hirsch, A. The nodule microbiome: N2-fixing rhizobia do not live alone. In Phytobiomes Journal; Carolyn, A., Young, C.A., Eds.; The American Phytopathological Society: St. Paul, MN, USA, 2017; Volume 1, pp. 70–82. [Google Scholar] [CrossRef] [Green Version]
- Tang, M.; Capela, D. Rhizobium diversity in the light of evolution. In Advances in Botanic Research: Regulations of Nitrogen-Fixing Symbioses in Legumes; Frendo, P., Frugier, F., Masson-Boivin, C., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; Volume 94, pp. 251–288. [Google Scholar]
- Zgadzaj, R.; Garrido-Oter, R.; Jensen, D.B.; Koprivova, A.; Schulze-Lefert, P.; Radutoiu, S. Root nodule symbiosis in Lotus japonicus drives the establishment of distinctive rhizosphere, root, and nodule bacterial communities. Proc. Natl. Acad. Sci. USA 2016, 113, E7996–E8005. [Google Scholar] [CrossRef] [Green Version]
- Mendes, R.; Garbeva, P.; Raaijmakers, J.M. The rhizosphere microbiome: Significance of plant beneficial, plant pathogenic, and human pathogenic microorganisms. FEMS Microbiol. Rev. 2013, 37, 634–663. [Google Scholar] [CrossRef]
- Xiao, X.; Chen, W.; Zong, L.; Yang, J.; Jiao, S.; Lin, Y.; Wang, E.; Wei, G. Two cultivated legume plants reveal the enrichment process of the microbiome in the rhizocompartments. Mol. Ecol. 2017, 26, 1641–1651. [Google Scholar] [CrossRef]
- Gage, D.J. Infection and invasion of roots by symbiotic, nitrogen-fixing rhizobia during nodulation of temperate legumes. Microbiol. Mol. Biol. Rev. 2004, 68, 280–300. [Google Scholar] [CrossRef] [Green Version]
- Triplett, E.W.; Sadowsky, M.J. Genetics of competition for nodulation of Legumes. Ann. Rev. Microbiol. 1992, 46, 399–428. [Google Scholar] [CrossRef] [PubMed]
- Long, S.R.; Buikema, W.J.; Ausubel, F.M. Cloning of Rhizobium meliloti nodulation genes by direct complementation of Nod- mutants. Nature 1982, 298, 485–488. [Google Scholar] [CrossRef]
- Clúa, J.; Roda, C.; Zanetti, M.E.; Blanco, F.A. Compatibility between legumes and rhizobia for the establishment of a successful nitrogen-fixing symbiosis. Genes (Basel) 2018, 9, 125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capoen, W.; Goormachtig, S.; De Rycke, R.; Schroeyers, K.; Holsters, M. SrSymRK, a plant receptor essential for symbiosome formation. Proc. Natl. Acad. Sci. USA 2005, 102, 10369–10374. [Google Scholar] [CrossRef] [Green Version]
- Ovchinnikova, E.; Journet, E.-P.; Chabaud, M.; Cosson, V.; Ratet, P.; Duc, G.; Fedorova, E.; Liu, W.; den Camp, R.O.; Zhukov, V.; et al. IPD3 controls the formation of nitrogen-fixing symbiosomes in Pea and Medicago Spp. Mol. Plant-Microbe Interact. 2011, 24, 1333–1344. [Google Scholar] [CrossRef] [Green Version]
- Moling, S.; Pietraszewska-Bogiel, A.; Postma, M.; Fedorova, E.; Hink, M.A.; Limpens, E.; Gadella, T.W.; Bisseling, T. Nod factor receptors form heteromeric complexes and are essential for intracellular infection in Medicago nodules. Plant Cell 2014, 26, 4188–4199. [Google Scholar] [CrossRef] [Green Version]
- Xiao, T.T.; Schilderink, S.; Moling, S.; Deinum, E.E.; Kondorosi, E.; Franssen, H.; Kulikova, O.; Niebel, A.; Bisseling, T. Fate map of Medicago truncatula root nodules. Development 2014, 141, 3517–3528. [Google Scholar] [CrossRef] [Green Version]
- Westhoek, A.; Field, E.; Rehling, F.; Mulley, G.; Webb, I.; Poole, P.S.; Turnbull, L.A. Policing the legume-Rhizobium symbiosis: A critical test of partner choice. Sci. Rep. 2017, 7, 1419. [Google Scholar] [CrossRef]
- Hahn, M.; Studer, D. Competitiveness of a nif Bradyrhizobium japonicum mutant against the wild-type strain. FEMS Microbiol. Lett. 1986, 33, 143–148. [Google Scholar] [CrossRef]
- Amarger, N. Competition for nodule formation between effective and ineffective strains of Rhizobium meliloti. Soil Biol. Biochem. 1981, 13, 475–480. [Google Scholar] [CrossRef]
- Gubry-Rangin, C.; Garcia, M.; Bena, G. Partner choice in Medicago Truncatula-Sinorhizobium symbiosis. Proc. Biol. Sci. 2010, 277, 1947–1951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heath, K.D.; Tiffin, P. Stabilizing mechanisms in a legume-rhizobium mutualism. Evolution 2009, 63, 652–662. [Google Scholar] [CrossRef] [PubMed]
- Younginger, B.S.; Friesen, M.L. Connecting signals and benefits through partner choice in plant-microbe interactions. FEMS Microbiol. Lett. 2019, 366, fnz217. [Google Scholar] [CrossRef] [PubMed]
- Kiers, E.T.; Denison, R.F. Sanctions, cooperation, and the stability of plant-rhizosphere mutualisms. Ann. Rev. Ecol. Evol. Syst. 2008, 39, 215–236. [Google Scholar] [CrossRef] [Green Version]
- Friesen, M.L. Widespread fitness alignment in the legume-rhizobium symbiosis. New Phytol. 2012, 194, 1096–1111. [Google Scholar] [CrossRef] [PubMed]
- Regus, J.U.; Quides, K.W.; O’Neill, M.R.; Suzuki, R.; Savory, E.A.; Chang, J.H.; Sachs, J.L. Cell autonomous sanctions in legumes target ineffective rhizobia in nodules with mixed infections. Am. J. Bot. 2017, 104, 1299–1312. [Google Scholar] [CrossRef] [Green Version]
- Burghardt, L.T. Evolving together, evolving apart: Measuring the fitness of rhizobial bacteria in and out of symbiosis with leguminous plants. New Phytol. 2019. [Google Scholar] [CrossRef] [Green Version]
- Denamur, E.; Matic, I. Evolution of mutation rates in bacteria. Mol. Microbiol. 2006, 60, 820–827. [Google Scholar] [CrossRef]
- Raynes, Y.; Sniegowski, P.D. Experimental evolution and the dynamics of genomic mutation rate modifiers. Heredity (Edinb.) 2014, 113, 375–380. [Google Scholar] [CrossRef] [Green Version]
- Foster, P.L. Stress-induced mutagenesis in bacteria. Crit. Rev. Biochem. Mol. Biol. 2007, 42, 373–397. [Google Scholar] [CrossRef]
- Galhardo, R.S.; Hastings, P.J.; Rosenberg, S.M. Mutation as a stress response and the regulation of evolvability. Crit. Rev. Biochem. Mol. Biol. 2007, 42, 399–435. [Google Scholar] [CrossRef] [Green Version]
- Remigi, P.; Capela, D.; Clerissi, C.; Tasse, L.; Torchet, R.; Bouchez, O.; Batut, J.; Cruveiller, S.; Rocha, E.P.; Masson-Boivin, C. Transient hypermutagenesis accelerates the evolution of legume endosymbionts following horizontal gene transfer. PLoS Biol. 2014, 12, e1001942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Didelot, X.; Walker, A.S.; Peto, T.E.; Crook, D.W.; Wilson, D.J. Within-host evolution of bacterial pathogens. Nat. Rev. Microbiol. 2016, 14, 150–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linz, B.; Windsor, H.M.; McGraw, J.J.; Hansen, L.M.; Gajewski, J.P.; Tomsho, L.P.; Hake, C.M.; Solnick, J.V.; Schuster, S.C.; Marshall, B.J. A mutation burst during the acute phase of Helicobacter pylori infection in humans and rhesus macaques. Nat. Commun. 2014, 5, 4165. [Google Scholar] [CrossRef]
- Barros-Carvalho, G.A.; Hungria, M.; Lopes, F.M.; Van Sluys, M.A. Brazilian-adapted soybean Bradyrhizobium strains uncover IS elements with potential impact on biological nitrogen fixation. FEMS Microbiol. Lett. 2019, 366, fnz046. [Google Scholar] [CrossRef] [PubMed]
- Dupuy, P.; Sauviac, L.; Bruand, C. Stress-inducible NHEJ in bacteria: Function in DNA repair and acquisition of heterologous DNA. Nucleic Acids Res. 2019, 47, 1335–1349. [Google Scholar] [CrossRef] [PubMed]
- Wielgoss, S.; Barrick, J.E.; Tenaillon, O.; Wiser, M.J.; Dittmar, W.J.; Cruveiller, S.; Chane-Woon-Ming, B.; Medigue, C.; Lenski, R.E.; Schneider, D. Mutation rate dynamics in a bacterial population reflect tension between adaptation and genetic load. Proc. Natl. Acad. Sci. USA 2013, 110, 222–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDonald, M.J.; Hsieh, Y.Y.; Yu, Y.H.; Chang, S.L.; Leu, J.Y. The evolution of low mutation rates in experimental mutator populations of Saccharomyces cerevisiae. Curr. Biol. 2012, 22, 1235–1240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swings, T.; Van den Bergh, B.; Wuyts, S.; Oeyen, E.; Voordeckers, K.; Verstrepen, K.J.; Fauvart, M.; Verstraeten, N.; Michiels, J. Adaptive tuning of mutation rates allows fast response to lethal stress in Escherichia coli. Elife 2017, 6, e22939. [Google Scholar] [CrossRef]
- Remigi, P.; Masson-Boivin, C.; Rocha, E.P.C. Experimental evolution as a tool to investigate natural processes and molecular functions. Trends Microbiol. 2019, 27, 623–634. [Google Scholar] [CrossRef] [Green Version]
- Yi, X. Experimental evolution and proximate mechanisms in biology. Synth. Syst. Biotechnol. 2017, 2, 253–258. [Google Scholar] [CrossRef] [PubMed]
- Peeters, N.; Guidot, A.; Vailleau, F.; Valls, M. Ralstonia solanacearum, a widespread bacterial plant pathogen in the post-genomic era. Mol. Plant Pathol. 2013, 14, 651–662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ray, S.K.; Kumar, R.; Peeters, N.; Boucher, C.; Genin, S. rpoN1, but not rpoN2, is required for twitching motility, natural competence, growth on nitrate, and virulence of Ralstonia solanacearum. Front. Microbiol. 2015, 6, 229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hikichi, Y.; Mori, Y.; Ishikawa, S.; Hayashi, K.; Ohnishi, K.; Kiba, A.; Kai, K. Regulation involved in colonization of intercellular spaces of host plants. Front. Plant Sci. 2017, 8, 967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Zhang, W.; Han, L.; Li, J.; Shi, X.; Hikichi, Y.; Ohnishi, K. Involvement of a PadR regulator PrhP on virulence of Ralstonia solanacearum by controlling detoxification of phenolic acids and type III secretion system. Mol. Plant Pathol. 2019, 20, 1477–1490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Li, J.; Zhang, W.; Shi, H.; Luo, F.; Hikichi, Y.; Shi, X.; Ohnishi, K. A putative LysR-type transcriptional regulator PrhO positively regulates the type III secretion system and contributes to the virulence of Ralstonia solanacearum. Mol. Plant Pathol. 2018, 19, 1808–1819. [Google Scholar] [CrossRef] [Green Version]
- Schell, M.A. Control of virulence and pathogenicity genes of Ralstonia solanacearum by an elaborate sensory network. Ann. Rev. Phytopathol. 2000, 38, 263–292. [Google Scholar] [CrossRef]
- Clough, S.J.; Lee, K.E.; Schell, M.A.; Denny, T.P. A two-component system in Ralstonia (Pseudomonas) solanacearum modulates production of PhcA-regulated virulence factors in response to 3-hydroxypalmitic acid methyl ester. J. Bacteriol. 1997, 179, 3639–3648. [Google Scholar] [CrossRef] [Green Version]
- Guidot, A.; Jiang, W.; Ferdy, J.-B.; Thebaud, C.; Barberis, P.; Gouzy, J.; Genin, S. Multihost experimental evolution of the pathogen Ralstonia solanacearum unveils genes involved in adaptation to plants. Mol. Biol. Evol. 2014, 31, 2913–2928. [Google Scholar] [CrossRef] [Green Version]
- D’Haeze, W.; Holsters, M. Nod factor structures, responses, and perception during initiation of nodule development. Glycobiology 2002, 12, 79R–105R. [Google Scholar] [CrossRef]
- Oldroyd, G.E.D.; Murray, J.D.; Poole, P.S.; Downie, J.A. The rules of engagement in the legume-rhizobial symbiosis. Ann. Rev. Genet. 2011, 45, 119–144. [Google Scholar] [CrossRef] [PubMed]
- Daubech, B. Évolution Expérimentale d’un Symbiote de Légumineuse: Étude des Facteurs Génétiques et des Forces de Sélection qui Favorisent ou non l’évolution du Mutualisme. Ph.D. Thesis, Université Toulouse 3 Paul Sabatier, Toulouse, France, 2019. [Google Scholar]
- Daubech, B.; Poinsot, V.; Klonowska, A.; Capela, D.; Chaintreuil, C.; Moulin, L.; Marchetti, M.; Masson-Boivin, C. A new nodulation gene involved in the biosynthesis of Nod Factors with an open-chain oxidized terminal residue and in the symbiosis with Mimosa pudica. Mol. Plant Microbe Interact. 2019, 32, 1635–1648. [Google Scholar] [CrossRef] [PubMed]
- Sugawara, M.; Takahashi, S.; Umehara, Y.; Iwano, H.; Tsurumaru, H.; Odake, H.; Suzuki, Y.; Kondo, H.; Konno, Y.; Yamakawa, T.; et al. Variation in bradyrhizobial NopP effector determines symbiotic incompatibility with Rj2-soybeans via effector-triggered immunity. Nat. Commun. 2018, 9, 3139. [Google Scholar] [CrossRef] [PubMed]
- Crook, M.B.; Lindsay, D.P.; Biggs, M.B.; Bentley, J.S.; Price, J.C.; Clement, S.C.; Clement, M.J.; Long, S.R.; Griffitts, J.S. Rhizobial plasmids that cause impaired symbiotic nitrogen fixation and enhanced host invasion. Mol. Plant Microbe Interact. 2012, 25, 1026–1033. [Google Scholar] [CrossRef] [Green Version]
- Burghardt, L.T.; Epstein, B.; Guhlin, J.; Nelson, M.S.; Taylor, M.R.; Young, N.D.; Sadowsky, M.J.; Tiffin, P. Select and resequence reveals relative fitness of bacteria in symbiotic and free-living environments. Proc. Natl. Acad. Sci. USA 2018, 115, 2425–2430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burghardt, L.T.; Trujillo, D.I.; Epstein, B.; Tiffin, P.; Young, N.D. A select and resequence approach reveals strain-specific effects of Medicago nodule-specific PLAT-domain genes. Plant Physiol. 2020, 182, 463–471. [Google Scholar] [CrossRef] [Green Version]
- Rogers, C.; Oldroyd, G.E.D. Synthetic biology approaches to engineering the nitrogen symbiosis in cereals. J. Exp. Bot. 2014, 65, 1939–1946. [Google Scholar] [CrossRef] [Green Version]
- Pankievicz, V.C.S.; Irving, T.B.; Maia, L.G.S.; Ané, J.M. Are we there yet? The long walk towards the development of efficient symbiotic associations between nitrogen-fixing bacteria and non-leguminous crops. BMC Biol. 2019, 17, 99. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Phylogeny of naturally and experimentally evolved Mimosa pudica symbionts in the Cupriavidus and Ralstonia branches. (A) Neighbour-joining phylogeny of selected Ralstonia and Cupriavidus strains based on 1003 gene families defined as core genome by [17]. Evolved “GMI1000pRalta” represents a group of 18 closely related strains derived via experimental evolution from the strain Ralstonia solanacearum GMI1000. (B) Close-up of the phylogenetic relationships between the 18 resequenced clones evolved for 16 cycles (two years) compared to the ancestor (R. solanacearum GMI1000). This neighbour-joining phylogeny is based on single-nucleotide polymorphisms identified in the experimentally evolved strains. (A) and (B) M. pudica symbionts are in red and bold. Plant pathogens, opportunistic human pathogens and saprophytic strains are in blue, purple and black, respectively. Arrows indicate the acquisition of a symbiotic plasmid, either the ancestor of the Cupriavidus taiwanensis one (pRaltaanc) during natural evolution or that of C. taiwanensis LMG19424 (pRaltaLMG19424) during experimental evolution.
Figure 1.
Phylogeny of naturally and experimentally evolved Mimosa pudica symbionts in the Cupriavidus and Ralstonia branches. (A) Neighbour-joining phylogeny of selected Ralstonia and Cupriavidus strains based on 1003 gene families defined as core genome by [17]. Evolved “GMI1000pRalta” represents a group of 18 closely related strains derived via experimental evolution from the strain Ralstonia solanacearum GMI1000. (B) Close-up of the phylogenetic relationships between the 18 resequenced clones evolved for 16 cycles (two years) compared to the ancestor (R. solanacearum GMI1000). This neighbour-joining phylogeny is based on single-nucleotide polymorphisms identified in the experimentally evolved strains. (A) and (B) M. pudica symbionts are in red and bold. Plant pathogens, opportunistic human pathogens and saprophytic strains are in blue, purple and black, respectively. Arrows indicate the acquisition of a symbiotic plasmid, either the ancestor of the Cupriavidus taiwanensis one (pRaltaanc) during natural evolution or that of C. taiwanensis LMG19424 (pRaltaLMG19424) during experimental evolution.

Figure 2.
Experimental evolution of the plant pathogen Ralstonia solanacearum into legume symbionts. (A) The symbiotic plasmid (pRalta) of Cupriavidus taiwanensis, the natural symbiont of Mimosa pudica, was introduced into R. solanacearum [21], generating a chimeric strain unable to nodulate M. pudica. This proto-rhizobium was then evolved through serial cycles of inoculation to M. pudica plantlets and re-isolation of nodule bacteria. In the first cycle, three nodulating variants were entrapped by the plant, and in the following cycles these variants were symbiotically improved (see panel B). In addition to the nod and nif-fix genes involved in Nod Factor synthesis and nitrogen fixation respectively, the pRalta plasmid carries a mutagenesis cassette (imuA2B2C2) that elevates mutation rate when bacteria are free-living in the plant culture medium. This transient hypermutagenesis increases the genetic diversity of the rhizospheric bacterial population among which the plant selects the most beneficial variants. (B) 18 parallel lineages were derived from the three nodulating (Nod+) variants via two different selection regimes (cycles of 21 or 42 days). After acquisition of the nodulation ability, bacteria progressively improved their capacity to infect nodules. Three levels of infection were observed in the evolved lines: extracellular, partially intracellular and nicely intracellular. The main adaptive mutations responsible for the acquisition and improvement of symbiosis were identified [21,22,23,24]. *, stop mutation. FS, frameshift. up112, up115, intergenic mutations located 112 bp and 115 bp upstream from the gene indicated.
Figure 2.
Experimental evolution of the plant pathogen Ralstonia solanacearum into legume symbionts. (A) The symbiotic plasmid (pRalta) of Cupriavidus taiwanensis, the natural symbiont of Mimosa pudica, was introduced into R. solanacearum [21], generating a chimeric strain unable to nodulate M. pudica. This proto-rhizobium was then evolved through serial cycles of inoculation to M. pudica plantlets and re-isolation of nodule bacteria. In the first cycle, three nodulating variants were entrapped by the plant, and in the following cycles these variants were symbiotically improved (see panel B). In addition to the nod and nif-fix genes involved in Nod Factor synthesis and nitrogen fixation respectively, the pRalta plasmid carries a mutagenesis cassette (imuA2B2C2) that elevates mutation rate when bacteria are free-living in the plant culture medium. This transient hypermutagenesis increases the genetic diversity of the rhizospheric bacterial population among which the plant selects the most beneficial variants. (B) 18 parallel lineages were derived from the three nodulating (Nod+) variants via two different selection regimes (cycles of 21 or 42 days). After acquisition of the nodulation ability, bacteria progressively improved their capacity to infect nodules. Three levels of infection were observed in the evolved lines: extracellular, partially intracellular and nicely intracellular. The main adaptive mutations responsible for the acquisition and improvement of symbiosis were identified [21,22,23,24]. *, stop mutation. FS, frameshift. up112, up115, intergenic mutations located 112 bp and 115 bp upstream from the gene indicated.

Figure 3.
Adaptation to endosymbiosis mainly occurred through regulatory rewiring. Genes mutated in the experiment are indicated in red. Nodulation was gained by inactivation of the T3SS via a stop mutation in hrcV [21]. A first level of intracellular infection was obtained by the inactivation of the virulence regulator HrpG [21,22]. Optimized intracellular infection was reached either by inactivation of the EfpR pathway or a modulation of the quorum sensing pathway controlling PhcA [23,24]. EfpR and PhcA controls a common set of genes as well as other specific genes. It is not known which group of genes controlled by EfpR and/or PhcA interfere with intracellular infection. Transcriptional regulators are in bold. 3OH-MAME, (R)-3-hydroxymyristic acid methyl ester, quorum sensing molecules produced by the GMI1000 strain of R. solanacearum.
Figure 3.
Adaptation to endosymbiosis mainly occurred through regulatory rewiring. Genes mutated in the experiment are indicated in red. Nodulation was gained by inactivation of the T3SS via a stop mutation in hrcV [21]. A first level of intracellular infection was obtained by the inactivation of the virulence regulator HrpG [21,22]. Optimized intracellular infection was reached either by inactivation of the EfpR pathway or a modulation of the quorum sensing pathway controlling PhcA [23,24]. EfpR and PhcA controls a common set of genes as well as other specific genes. It is not known which group of genes controlled by EfpR and/or PhcA interfere with intracellular infection. Transcriptional regulators are in bold. 3OH-MAME, (R)-3-hydroxymyristic acid methyl ester, quorum sensing molecules produced by the GMI1000 strain of R. solanacearum.

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Doin de Moura, G.G.; Remigi, P.; Masson-Boivin, C.; Capela, D. Experimental Evolution of Legume Symbionts: What Have We Learnt? Genes 2020, 11, 339. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11030339
AMA Style
Doin de Moura GG, Remigi P, Masson-Boivin C, Capela D. Experimental Evolution of Legume Symbionts: What Have We Learnt? Genes. 2020; 11(3):339. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11030339
Chicago/Turabian StyleDoin de Moura, Ginaini Grazielli, Philippe Remigi, Catherine Masson-Boivin, and Delphine Capela. 2020. "Experimental Evolution of Legume Symbionts: What Have We Learnt?" Genes 11, no. 3: 339. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11030339
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.