Analysis of HBV Genomes Integrated into the Genomes of Human Hepatoma PLC/PRF/5 Cells by HBV Sequence Capture-Based Next-Generation Sequencing

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Cellular DNA Extraction and DNA Fragmentation
2.3. Library Preparation and HBV Sequence Capture-Based Next-Generation Sequencing (NGS)
2.4. Confirmation of HBV Genome Integration by Sanger Sequencing Methods
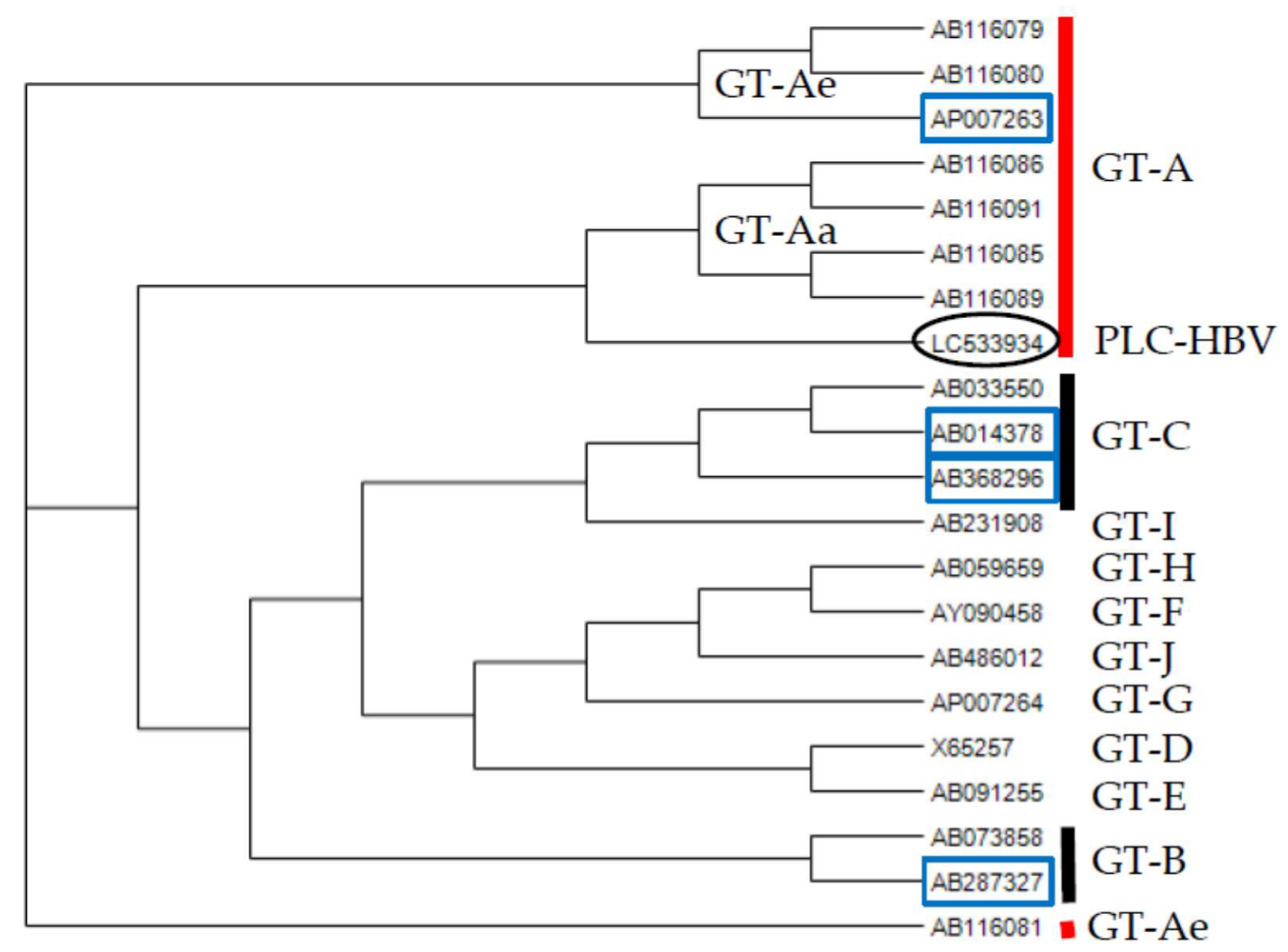
2.5. Phylogenetic Analysis
2.6. Immunofluorescence Study
3. Results
3.1. HBV DNA Sequence Derived from PLC/PRF/5 Cells
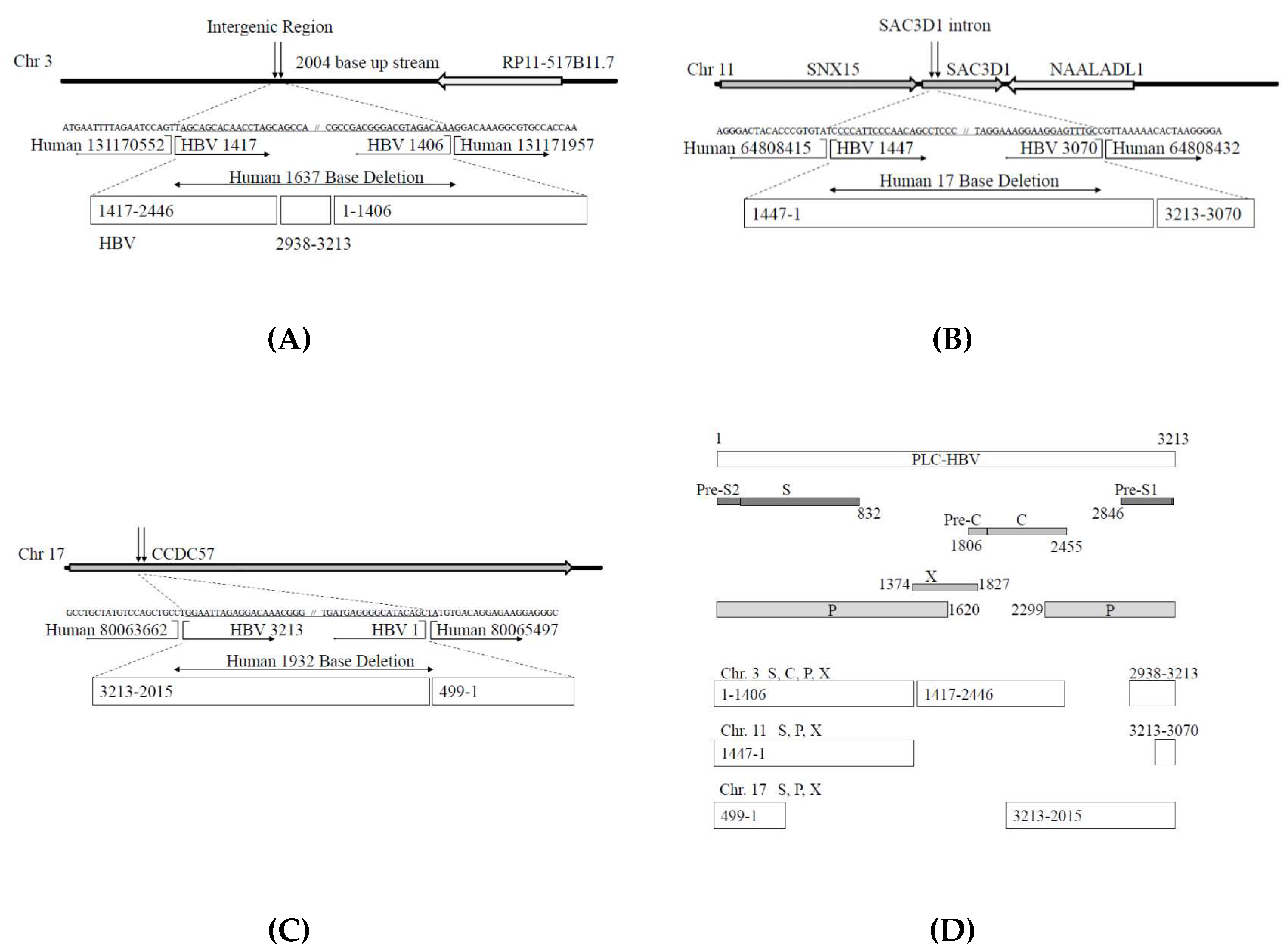
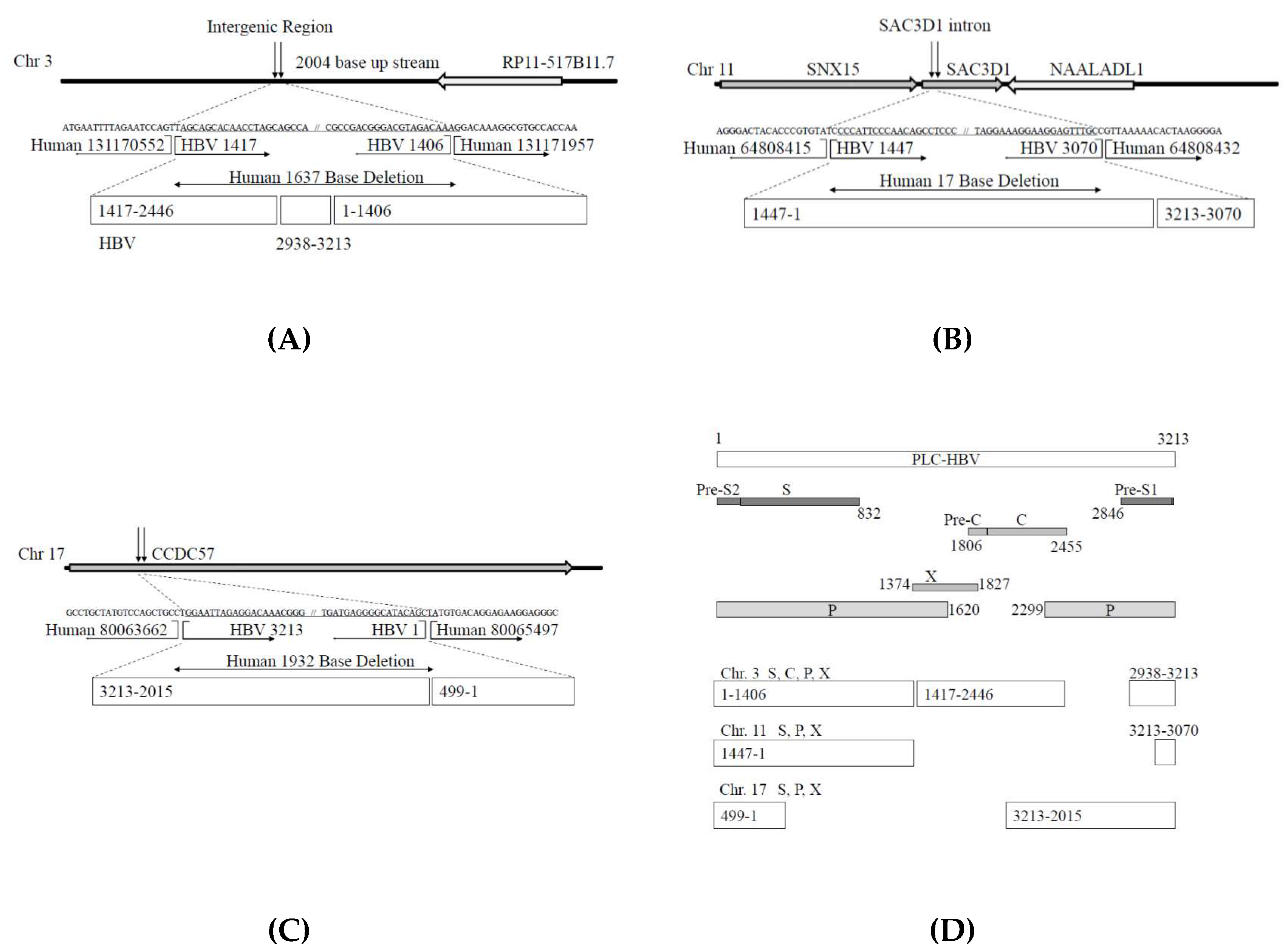
3.2. Analysis of HBV Genome Integrants and Their Locations in the Human Genome
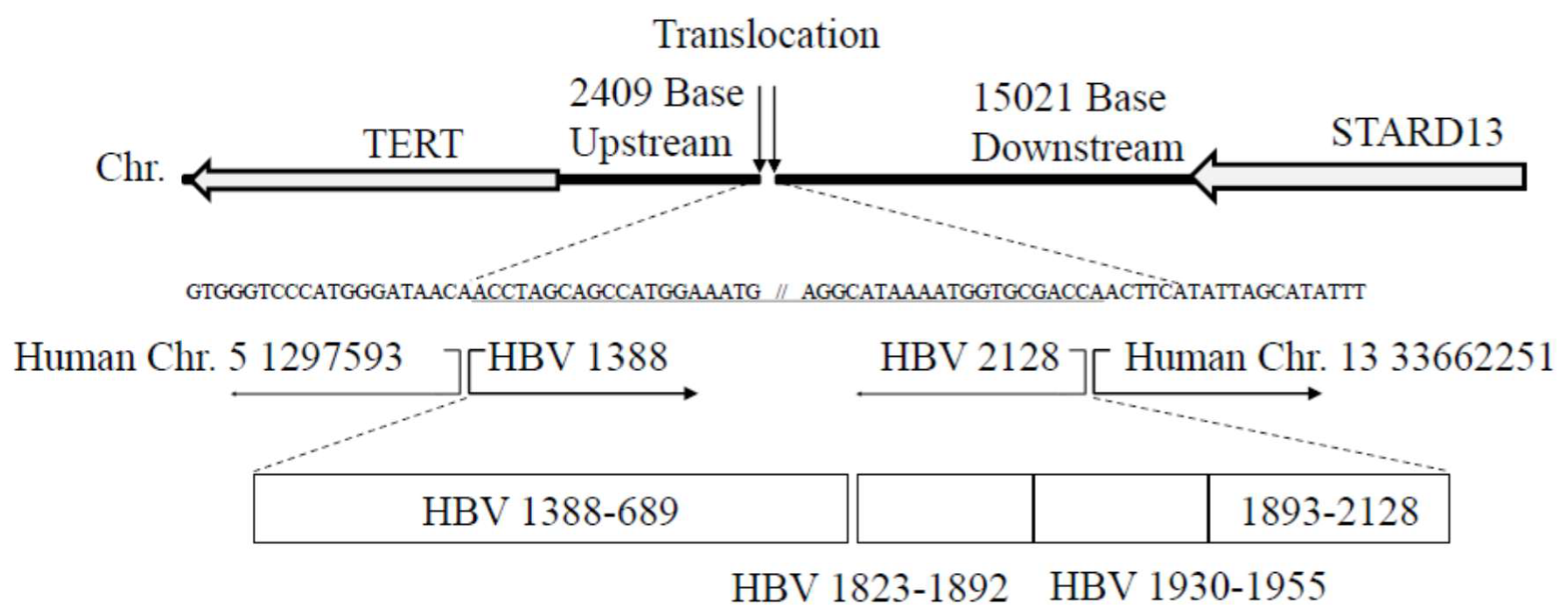
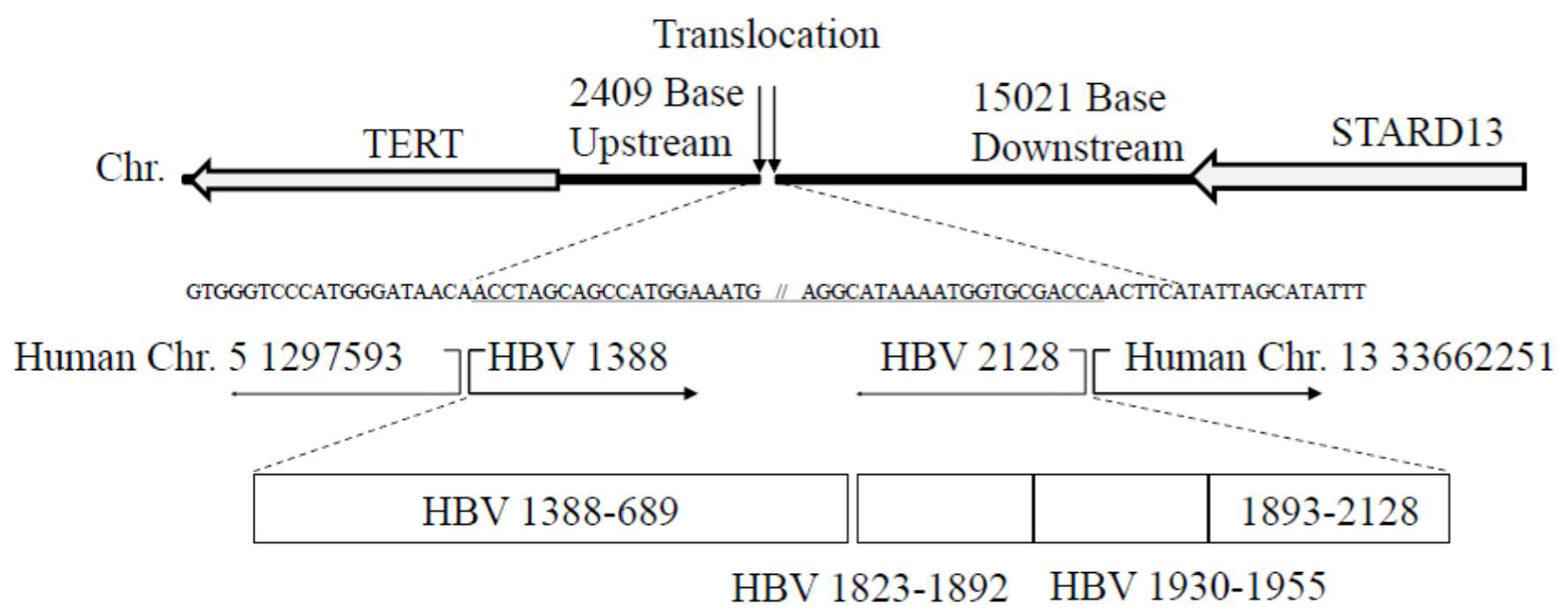
3.3. Translocation of Chromosomes (5; 13) with HBV Integrants
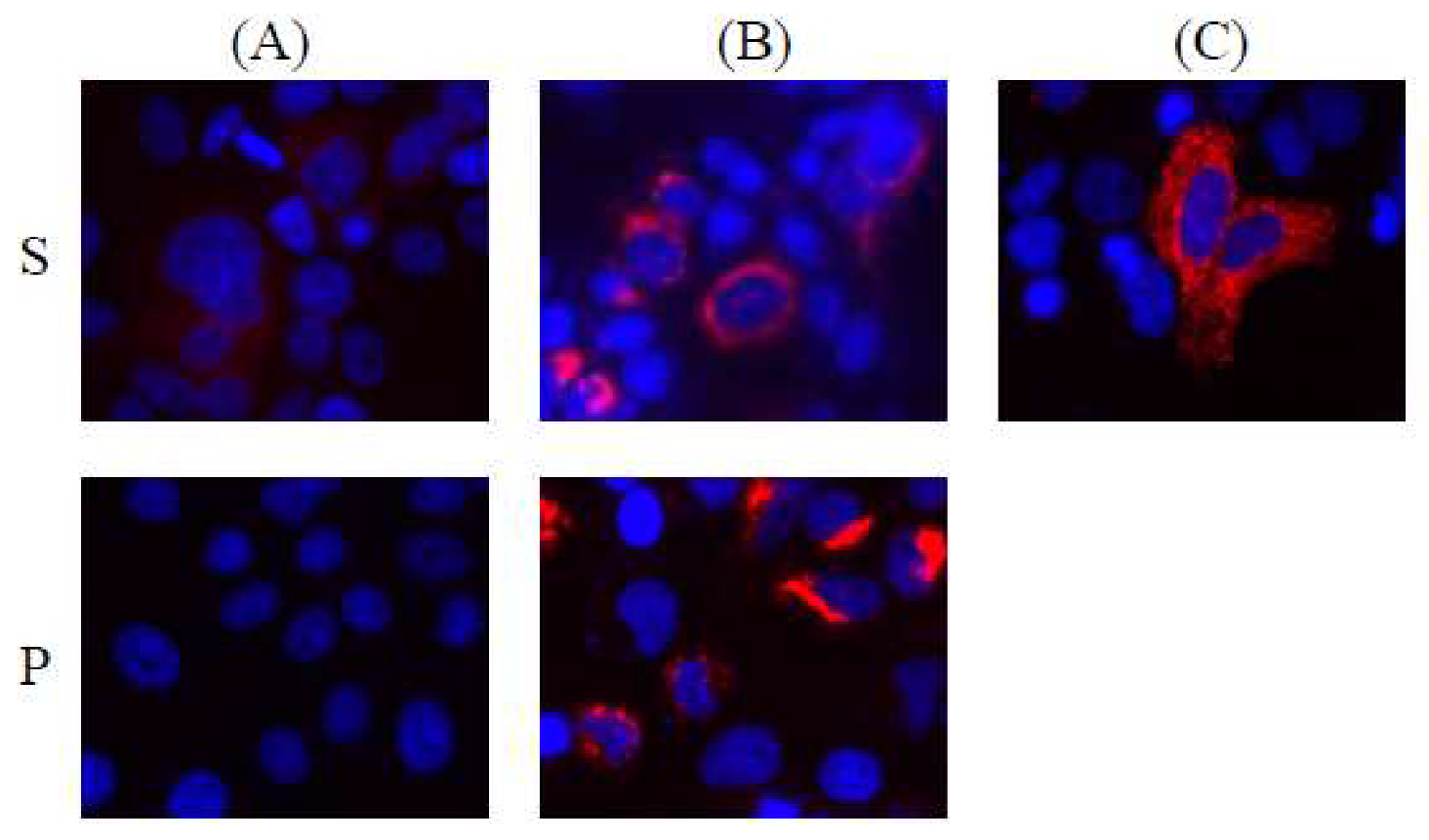
3.4. Automatic Expression of Proteins of HBV Integrants from Chromosomes 3 and 11, from the Vector without Any Promoter Sequences
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Suzuki, K.; Uchida, T.; Shikata, T.; Moriyama, M.; Arakawa, Y.; Mizokami, M.; Mima, F. Expression of pre-S1, pre-S2, S and X peptides in relation to viral replication in livers with chronic hepatitis B. Liver 1990, 10, 355–364. [Google Scholar] [CrossRef] [PubMed]
- Terada, T.; Moriyama, M.; Uchida, T.; Arakawa, Y. Nucleotide sequence of the precore/core gene and X gene of hepatitis B virus DNA in asymptomatic hepatitis B virus carriers who are negative for serum hepatitis B core antibody. Intervirology 2001, 44, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Yokosuka, O.; Omata, M.; Imazeki, F.; Ito, Y.; Okuda, K. Hepatitis B virus RNA transcripts and DNA in chronic liver disease. N. Engl. J. Med. 1986, 315, 1187–1192. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, R.; Churchill, N.D.; Mulrooney-Cousins, P.M.; Michalak, T.I. Initial sites of hepadnavirus integration into host genome in human hepatocytes and in the woodchuck model of hepatitis B-associated hepatocellular carcinoma. Oncogenesis 2017, 6, e317. [Google Scholar] [CrossRef] [PubMed]
- Tu, T.; Budzinska, M.A.; Vondran, F.W.R.; Shackel, N.A.; Urban, S. Hepatitis B Virus DNA Integration Occurs Early in the Viral Life Cycle in an In Vitro Infection Model via Sodium Taurocholate Cotransporting Polypeptide-Dependent Uptake of Enveloped Virus Particles. J. Virol. 2018, 92, e02007–e02017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Totoki, Y.; Tatsuno, K.; Covington, K.R.; Ueda, H.; Creighton, C.J.; Kato, M.; Tsuji, S.; Donehower, L.A.; Slagle, B.L.; Nakamura, H.; et al. Trans-ancestry mutational landscape of hepatocellular carcinoma genomes. Nat. Genet. 2014, 46, 1267–1273. [Google Scholar] [CrossRef] [PubMed]
- Zucman-Rossi, J.; Villanueva, A.; Nault, J.C.; Llovet, J.M. Genetic Landscape and Biomarkers of Hepatocellular Carcinoma. Gastroenterology 2015, 149, 1226–1239.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tatsuno, K.; Midorikawa, Y.; Takayama, T.; Yamamoto, S.; Nagae, G.; Moriyama, M.; Nakagawa, H.; Koike, K.; Moriya, K.; Aburatani, H. Impact of AAV2 and Hepatitis B Virus Integration Into Genome on Development of Hepatocellular Carcinoma in Patients with Prior Hepatitis B Virus Infection. Clin. Cancer Res. 2019, 25, 6217–6227. [Google Scholar] [CrossRef] [PubMed]
- Sung, W.K.; Zheng, H.; Li, S.; Chen, R.; Liu, X.; Li, Y.; Lee, N.P.; Lee, W.H.; Ariyaratne, P.N.; Tennakoon, C.; et al. Genome-wide survey of recurrent HBV integration in hepatocellular carcinoma. Nat. Genet. 2012, 44, 765–769. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.H.; Liu, X.; Yan, H.X.; Li, W.Y.; Zeng, X.; Yang, Y.; Zhao, J.; Liu, S.P.; Zhuang, X.H.; Lin, C.; et al. Genomic and oncogenic preference of HBV integration in hepatocellular carcinoma. Nat. Commun. 2016, 7, 12992. [Google Scholar] [CrossRef] [PubMed]
- No, J.S.; Kim, W.K.; Cho, S.; Lee, S.H.; Kim, J.A.; Lee, D.; Song, D.H.; Gu, S.H.; Jeong, S.T.; Wiley, M.R.; et al. Comparison of targeted next-generation sequencing for whole-genome sequencing of Hantaan orthohantavirus in Apodemus agrarius lung tissues. Sci. Rep. 2019, 9, 16631. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Zeng, X.; Lee, N.P.; Liu, X.; Chen, S.; Guo, B.; Yi, S.; Zhuang, X.; Chen, F.; Wang, G.; et al. HIVID: An efficient method to detect HBV integration using low coverage sequencing. Genomics 2013, 102, 338–344. [Google Scholar] [CrossRef] [PubMed]
- Shukla, R.; Upton, K.R.; Muñoz-Lopez, M.; Gerhardt, D.J.; Fisher, M.E.; Nguyen, T.; Brennan, P.M.; Baillie, J.K.; Collino, A.; Ghisletti, S.; et al. Endogenous retrotransposition activates oncogenic pathways in hepatocellular carcinoma. Cell 2013, 153, 101–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacNab, G.M.; Alexander, J.J.; Lecatsas, G.; Bey, E.M.; Urbanowicz, J.M. Hepatitis B surface antigen produced by a human hepatoma cell line. Br. J. Cancer 1976, 34, 509–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poptsova, M.S.; Il’icheva, I.A.; Nechipurenko, D.Y.; Panchenko, L.A.; Khodikov, M.V.; Oparina, N.Y.; Polozov, R.V.; Nechipurenko, Y.D.; Grokhovsky, S.L. Non-random DNA fragmentation in next-generation sequencing. Sci. Rep. 2014, 4, 4532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimura, M.A. Simple Method for Estimating Evolutionary Rates of Base Substitutions Through Comparative Studies of Nucleotide Sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Yotsumoto, S.; Okamoto, H.; Tsuda, F.; Miyakawa, Y.; Mayumi, M. Subtyping hepatitis B virus DNA in free or integrated forms by amplification of the S-gene sequences by the polymerase chain reaction and single-track sequencing for adenine. J. Virol. Methods 1990, 28, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Freitas, N.; Cunha, C.; Menne, S.; Gudima, S.O. Envelope proteins derived from naturally integrated hepatitis B virus DNA support assembly and release of infectious hepatitis delta virus particles. J. Virol. 2014, 88, 5742–5754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haga, Y.; Kanda, T.; Nakamoto, S.; Nakamura, M.; Sasaki, R.; Wu, S.; Yokosuka, O. Interferon induces interleukin 8 and bone marrow stromal cell antigen 2 expression, inhibiting the production of hepatitis B virus surface antigen from human hepatocytes. Biochem. Biophys. Res. Commun. 2017, 486, 858–863. [Google Scholar] [CrossRef] [PubMed]
- Edman, J.C.; Gray, P.; Valenzuela, P.; Rall, L.B.; Rutter, W.J. Integration of hepatitis B virus sequences and their expression in a human hepatoma cell. Nature 1980, 286, 535–538. [Google Scholar] [CrossRef] [PubMed]
- Sugauchi, F.; Kumada, H.; Acharya, S.A.; Shrestha, S.M.; Gamutan, M.T.A.; Khan, M.; Gish, R.G.; Tanaka, Y.; Kato, T.; Orito, E.; et al. Epidemiological and sequence differences between two subtypes (Ae and Aa) of hepatitis B virus genotype A. J. Gen. Virol. 2004, 85, 811–820. [Google Scholar] [CrossRef] [PubMed]
- Shibayama, T.; Masuda, G.; Ajisawa, A.; Hiruma, K.; Tsuda, F.; Nishizawa, T.; Takahashi, M.; Okamoto, H. Characterization of seven genotypes (A to E, G and H) of hepatitis B virus recovered from Japanese patients infected with human immunodeficiency virus type 1. J. Med. Virol. 2005, 76, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, T.; Tanaka, Y.; Simonetti, J.; Osiowy, C.; Borresen, M.L.; Koch, A.; Kurbanov, F.; Sugiyama, M.; Minuk, G.Y.; McMahon, B.J.; et al. Classification of hepatitis B virus genotype B into 2 major types based on characterization of a novel subgenotype in Arctic indigenous populations. J. Infect. Dis. 2007, 196, 1487–1492. [Google Scholar] [CrossRef] [Green Version]
- Marion, P.L.; Salazar, F.H.; Alexander, J.J.; Robinson, W.S. State of hepatitis B viral DNA in a human hepatoma cell line. J. Virol. 1980, 33, 795–806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daemer, R.J.; Feinstone, S.M.; Alexander, J.J.; Tully, J.G.; London, W.T.; Wong, D.C.; Purcell, R.H. PLC/PRF/5 (Alexander) hepatoma cell line: Further characterization and studies of infectivity. Infect. Immun. 1980, 30, 607–611. [Google Scholar] [PubMed]
- Monjardino, J.P.; Fowler, M.J.; Thomas, H.C. Defective hepatitis B virus DNA molecules detected in a stable integration pattern in a hepatoma cell line, and in induced tumours and derived cell lines. J. Gen. Virol. 1983, 64 Pt 10, 2299–2303. [Google Scholar] [CrossRef] [PubMed]
- Sato, N.; Watanabe, S.; Miura, K.; Goka, R.; Morimoto, N.; Takaoka, Y.; Nomoto, H.; Tsukui, M.; Isoda, N.; Nagashima, S.; et al. Acute Liver Failure Caused by the Transmission of Hepatitis B Virus from the Spouse after 38 Years of Marriage. Intern. Med. 2019, 58, 2963–2968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogawa, M.; Kamimura, S.; Kanda, T.; Takahashi, H.; Mizutani, T.; Matsumoto, N.; Yamamoto, T.; Nirei, K.; Matsuoka, S.; Takei, M.; et al. Long-term follow-up of a Japanese patient with hepatitis B virus genotype H and human immunodeficiency virus coinfection. Future Virol. 2019, 14, 633–639. [Google Scholar] [CrossRef]
- Li, N.; Zhao, L.; Guo, C.; Liu, C.; Liu, Y. Identification of a novel DNA repair-related prognostic signature predicting survival of patients with hepatocellular carcinoma. Cancer Manag. Res. 2019, 11, 7473–7484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, M.E.; Kim, J.Y.; Kim, G.H.; Park, S.Y.; Kim, Y.H.; Oh, S.O. SAC3D1: A novel prognostic marker in hepatocellular carcinoma. Sci. Rep. 2018, 8, 15608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeannot, E.; Harlé, A.; Holmes, A.; Sastre-Garau, X. Nuclear factor I X is a recurrent target for HPV16 insertions in anal carcinomas. Genes Chromosomes Cancer. 2018, 57, 638–644. [Google Scholar] [CrossRef] [PubMed]
- Lesbats, P.; Engelman, A.N.; Cherepanov, P. Retroviral DNA Integration. Chem. Rev. 2016, 116, 12730–12757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bodelon, C.; Untereiner, M.E.; Machiela, M.J.; Vinokurova, S.; Wentzensen, N. Genomic characterization of viral integration sites in HPV-related cancers. Int. J. Cancer. 2016, 139, 2001–2011. [Google Scholar] [CrossRef] [PubMed]
- Aimola, G.; Beythien, G.; Aswad, A.; Kaufer, B.B. Current understanding of human herpesvirus 6 (HHV-6) chromosomal integration. Antiviral Res. 2020, 176, 104720. [Google Scholar] [CrossRef] [PubMed]
- Tuna, M.; Amos, C.I. Next, generation sequencing and its applications in HPV-associated cancers. Oncotarget 2017, 8, 8877–8889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowcock, A.M.; Pinto, M.R.; Bey, E.; Kuyl, J.M.; Dusheiko, G.M.; Bernstein, R. The PLC/PRF/5 human hepatoma cell line. II. Chromosomal assignment of hepatitis B virus integration sites. Cancer Genet. Cytogenet. 1985, 18, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Kojima, H.; Yokosuka, O.; Imazeki, F.; Saisho, H.; Omata, M. Telomerase activity and telomere length in hepatocellular carcinoma and chronic liver disease. Gastroenterology 1997, 112, 493–500. [Google Scholar] [CrossRef] [PubMed]
- Wolosz, D.; Walczak, A.; Szparecki, G.; Dwojak, M.; Winiarska, M.; Wolinska, E.; Gornicka, B. Deleted in Liver Cancer 2 (DLC2) protein expression in hepatocellular carcinoma. Eur. J. Histochem. 2019, 63. [Google Scholar] [CrossRef] [PubMed]
- Ching, Y.P.; Wong, C.M.; Chan, S.F.; Leung, T.H.; Ng, D.C.; Jin, D.Y.; Ng, I.O. Deleted in liver cancer (DLC) 2 encodes a RhoGAP protein with growth suppressor function and is underexpressed in hepatocellular carcinoma. J. Biol. Chem. 2003, 278, 10824–10830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiaorong, L.; Wei, W.; Liyuan, Q.; Kaiyan, Y. Underexpression of deleted in liver cancer 2 (DLC2) is associated with overexpression of RhoA and poor prognosis in hepatocellular carcinoma. BMC Cancer 2008, 8, 205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Wang, F.; Hu, Y. STARD13 promotes hepatocellular carcinoma apoptosis by acting as a ceRNA for Fas. Biotechnol. Lett. 2017, 39, 207–217. [Google Scholar] [CrossRef] [PubMed]
- Wylie, T.N.; Wylie, K.M.; Herter, B.N.; Storch, G.A. Enhanced virome sequencing using targeted sequence capture. Genome Res. 2015, 25, 1910–1920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wylie, K.M.; Wylie, T.N.; Buller, R.; Herter, B.; Cannella, M.T.; Storch, G.A. Detection of Viruses in Clinical Samples by Use of Metagenomic Sequencing and Targeted Sequence Capture. J. Clin. Microbiol. 2018, 56, e01123-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiselev, D.; Matsvay, A.; Abramov, I.; Dedkov, V.; Shipulin, G.; Khafizov, K. Current Trends in Diagnostics of Viral Infections of Unknown Etiology. Viruses 2020, 12, 211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, M.; Hao, J.; Cui, S.; Worthley, D.L.; Zhang, S.; Wang, Z.; Shi, J.; Liu, L.; Wang, X.; Ke, A.; et al. Diverse modes of clonal evolution in HBV-related hepatocellular carcinoma revealed by single-cell genome sequencing. Cell Res. 2018, 28, 359–373. [Google Scholar] [CrossRef] [PubMed]
- Brechot, C.; Pourcel, C.; Louise, A.; Rain, B.; Tiollais, P. Presence of integrated hepatitis B virus DNA sequences in cellular DNA of human hepatocellular carcinoma. Nature 1980, 286, 533–535. [Google Scholar] [CrossRef] [PubMed]
- Dejean, A.; Brechot, C.; Tiollais, P.; Wain-Hobson, S. Characterization of integrated hepatitis B viral DNA cloned from a human hepatoma and the hepatoma-derived cell line PLC/PRF/5. Proc. Natl. Acad. Sci. USA 1983, 80, 2505–2509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koch, S.; Freytag von Loringhoven, A.; Kahmann, R.; Hofschneider, P.H.; Koshy, R. The genetic organization of integrated hepatitis B virus DNA in the human hepatoma cell line PLC/PRF/5. Nucleic Acids Res. 1984, 12, 6871–6886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, Y.; Yamamoto, H.; Oikawa, R.; Toyota, M.; Yamamoto, M.; Kokudo, N.; Tanaka, S.; Arii, S.; Yotsuyanagi, H.; Koike, K.; et al. DNA methylation at hepatitis B viral integrants is associated with methylation at flanking human genomic sequences. Genome Res. 2015, 25, 328–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reference Sequence | Reference Length | Consensus Length | Total Reads | Single Reads | Reads in Pairs | Average Coverage |
|---|---|---|---|---|---|---|
| Chr. 1 | 2.49 × 108 | 1,545,531 | 190,160 | 18,518 | 171,642 | 0.076008 |
| Chr. 2 | 2.43 × 108 | 1,344,151 | 171,652 | 23,708 | 147,944 | 0.06996 |
| Chr. 3 | 1.98 × 108 | 985,756 | 134,430 | 19,312 | 115,118 | 0.067181 |
| Chr. 4 | 1.91 × 108 | 763,084 | 101,024 | 27,548 | 73,476 | 0.051384 |
| Chr. 5 | 1.81 × 108 | 878,501 | 107,805 | 12,905 | 94,900 | 0.059238 |
| Chr. 6 | 1.71 × 108 | 978,370 | 143,424 | 13,186 | 130,238 | 0.083435 |
| Chr. 7 | 1.59 × 108 | 1,044,242 | 136,171 | 13,449 | 122,722 | 0.085094 |
| Chr. 8 | 1.46 × 108 | 762,738 | 93,417 | 14,827 | 78,590 | 0.063299 |
| Chr. 9 | 1.41 × 108 | 599,234 | 65,741 | 9779 | 55,962 | 0.046138 |
| Chr. 10 | 1.36 × 108 | 769,055 | 110,259 | 27,795 | 82,464 | 0.079924 |
| Chr. 11 | 1.35 × 108 | 863,578 | 2,006,583 | 126,939 | 1,879,644 | 1.488781 |
| Chr. 12 | 1.34 × 108 | 736,429 | 115,138 | 21,172 | 93,966 | 0.084807 |
| Chr. 13 | 1.15 × 108 | 412,243 | 58,815 | 10,367 | 48,448 | 0.050038 |
| Chr. 14 | 1.07 × 108 | 455,809 | 53,252 | 4762 | 48,490 | 0.049338 |
| Chr. 15 | 1.03 × 108 | 490,545 | 348,653 | 17,775 | 330,878 | 0.338605 |
| Chr. 16 | 90,354,753 | 563,883 | 114,919 | 48,517 | 66,402 | 0.122922 |
| Chr. 17 | 81,195,210 | 524,407 | 106,805 | 25,583 | 81,222 | 0.128115 |
| Chr. 18 | 78,077,248 | 394,380 | 52,617 | 7057 | 45,560 | 0.067025 |
| Chr. 19 | 59,128,983 | 447,625 | 53,263 | 7119 | 46144 | 0.089332 |
| Chr. 20 | 63,025,520 | 474,048 | 68,184 | 5948 | 62,236 | 0.107619 |
| Chr. 21 | 48,129,895 | 254,738 | 32,117 | 4913 | 27,204 | 0.0658 |
| Chr. 22 | 51,304,566 | 253,101 | 29,371 | 3049 | 26,322 | 0.05685 |
| Chr. X | 1.55 × 108 | 674,067 | 73,717 | 8491 | 65,226 | 0.047146 |
| Chr. Y | 59,373,566 | 100,605 | 8838 | 8018 | 820 | 0.013505 |
| Chr. MT | 16,569 | 14,178 | 4540 | 172 | 4368 | 27.45869 |
| HBV | 3215 | 3213 | 1,784,734 | 448,408 | 1,336,326 | 53,732.01 |
| Human Chromosome (Chr.) | Human Junction Nucleotide Position | Gene Name | HBV Fragment Start Position | HBV Fragment End Position | HBV Genome | Number of Total Reads * |
|---|---|---|---|---|---|---|
| Chr.3 | 131170552 | NA | 1044 | 1406 | P, S, X | 3673 |
| Chr.3 | 131171957 | NA | 1415 | 1914 | C, P, S, X | 3589 |
| Chr.4 | 181507570 | NA | 96 | 432 | P, S | 1548 |
| Chr.4 | 181508764 | NA | 235 | 387 | P, S | 4377 |
| Chr.5 | 1297593 | NA | 1175 | 1364 | P, S | 3680 |
| Chr.8 | 35304663 | UNC5D | 2389 | 2862 | C, P, S | 4625 |
| Chr.11 | 64808415 | SNX15 | 1313 | 1575 | P, S, X | 5911 |
| Chr.11 | 64808432 | SAC3D1 | 2575 | 2851 | P, S | 6616 |
| Chr.12 | 110012332 | MVK | 692 | 1379 | P, S, X | 10,130 |
| Chr.13 | 33662251 | NA | 1897 | 2109 | C, P, S | 4060 |
| Chr.13 | 33662698 | NA | 783 | 1612 | P, S, X | 1494 |
| Chr.17 | 80063662 | CCDC57 | 428 | 2586 | P, S | 7699 |
| Chr.17 | 80065497 | CCDC57 | 2062 | 2420 | C, P, S | 6918 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ishii, T.; Tamura, A.; Shibata, T.; Kuroda, K.; Kanda, T.; Sugiyama, M.; Mizokami, M.; Moriyama, M. Analysis of HBV Genomes Integrated into the Genomes of Human Hepatoma PLC/PRF/5 Cells by HBV Sequence Capture-Based Next-Generation Sequencing. Genes 2020, 11, 661. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11060661
Ishii T, Tamura A, Shibata T, Kuroda K, Kanda T, Sugiyama M, Mizokami M, Moriyama M. Analysis of HBV Genomes Integrated into the Genomes of Human Hepatoma PLC/PRF/5 Cells by HBV Sequence Capture-Based Next-Generation Sequencing. Genes. 2020; 11(6):661. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11060661
Chicago/Turabian StyleIshii, Tomotaka, Akinori Tamura, Toshikatsu Shibata, Kazumichi Kuroda, Tatsuo Kanda, Masaya Sugiyama, Masashi Mizokami, and Mitsuhiko Moriyama. 2020. "Analysis of HBV Genomes Integrated into the Genomes of Human Hepatoma PLC/PRF/5 Cells by HBV Sequence Capture-Based Next-Generation Sequencing" Genes 11, no. 6: 661. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11060661