Omics Approaches Applied to Penicillium chrysogenum and Penicillin Production: Revealing the Secrets of Improved Productivity


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Genomics
2.1. The P. chrysogenum Wisconsin 54-1255 Genome
2.2. The P. chrysogenum P2niaD18 Genome
2.3. Industrial Strains Genomes
2.4. Analysis of the Mutations in the Genome of Penicillin High-Producing Strains
3. Transcriptomics
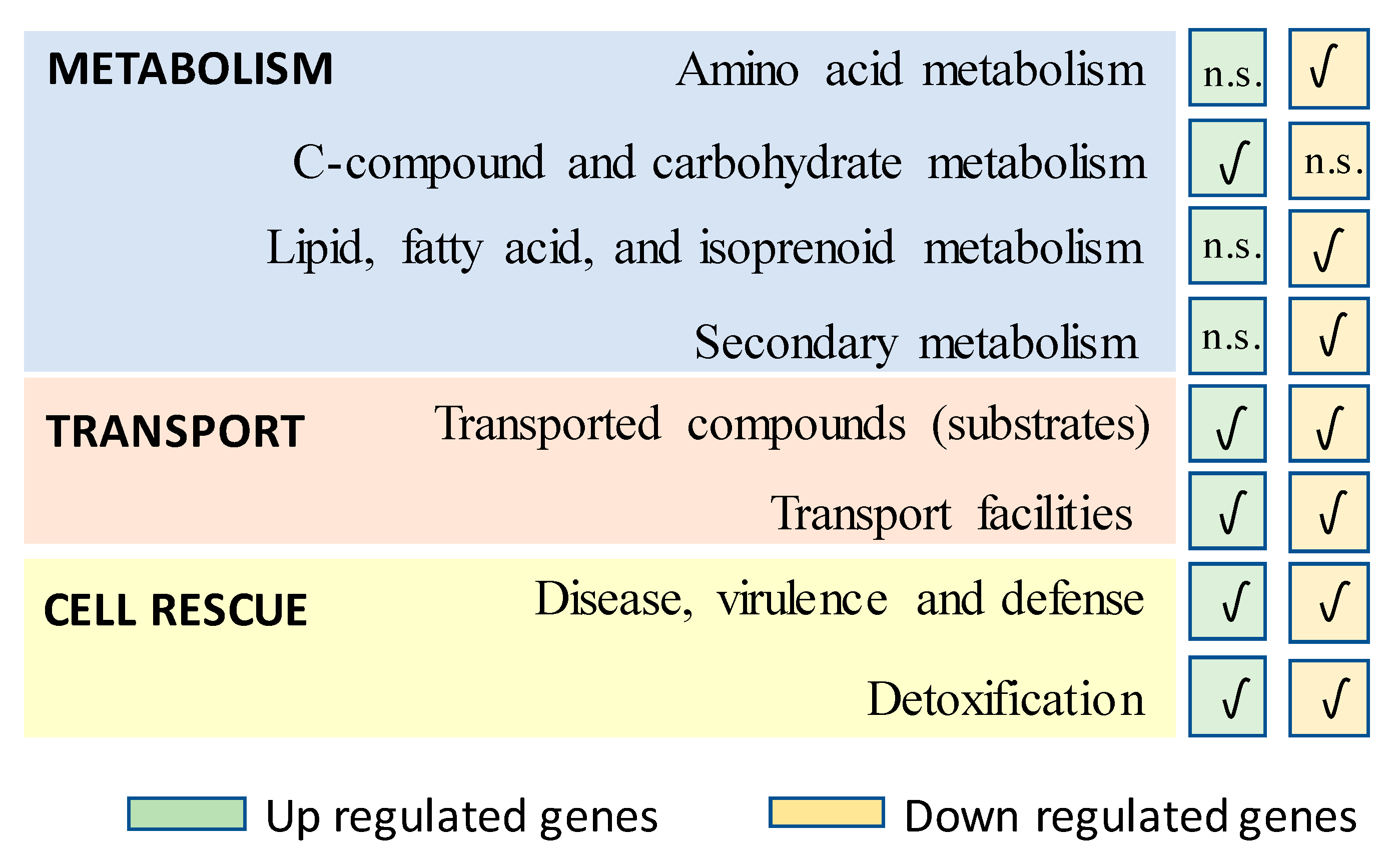
3.1. Transcriptomics and Strain Improvement
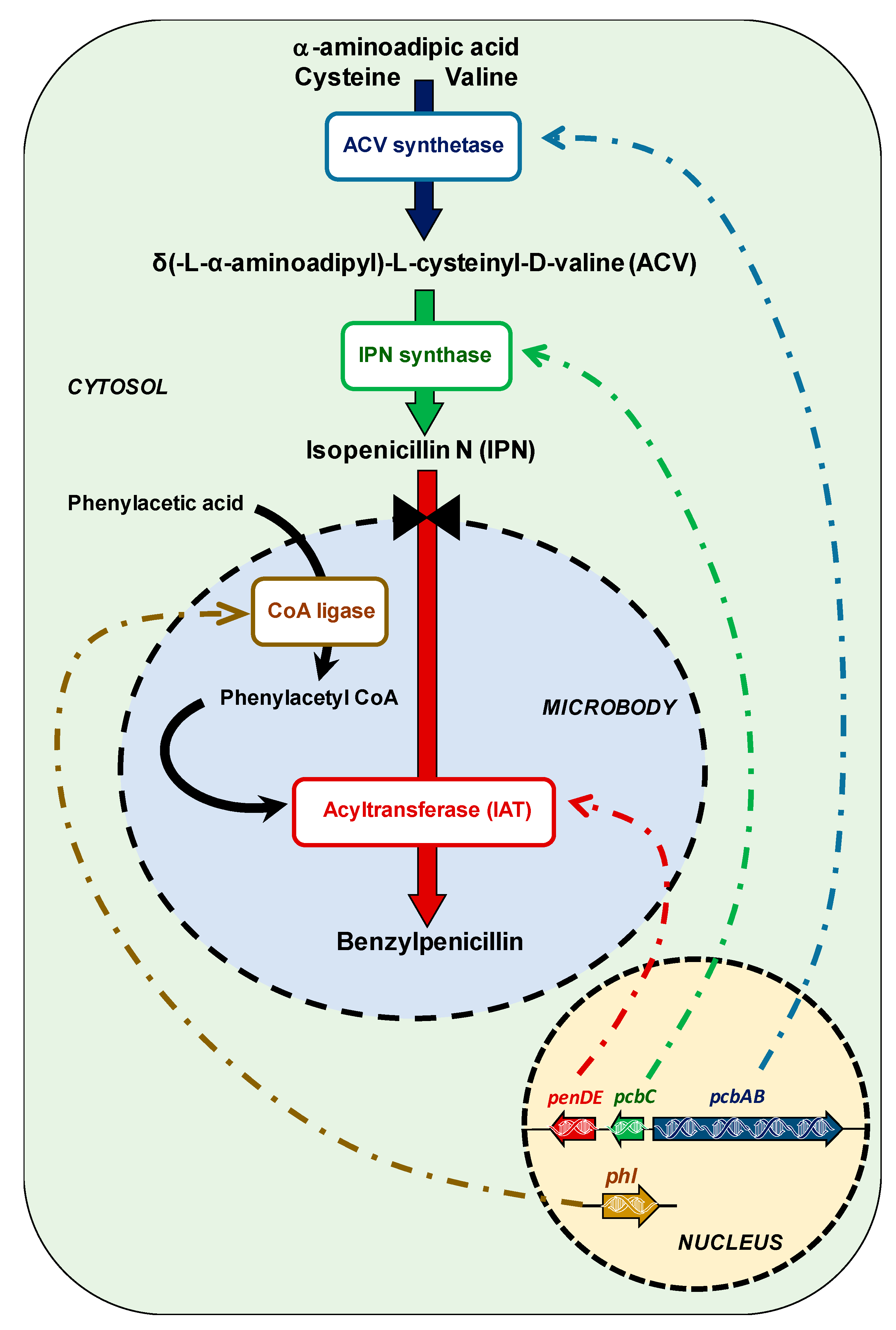
3.2. Transcriptomics and the Metabolism of the Benzylpenicillin Side-Chain Precursor
3.3. Transcriptomics and the Impact of the Velvet Regulatory Complex
3.4. Transcriptomics under Environmental Stress Conditions
3.5. Transcriptomics and Production of Cephem Antibiotics by Engineered Strains of P. chrysogenum
4. Proteomics
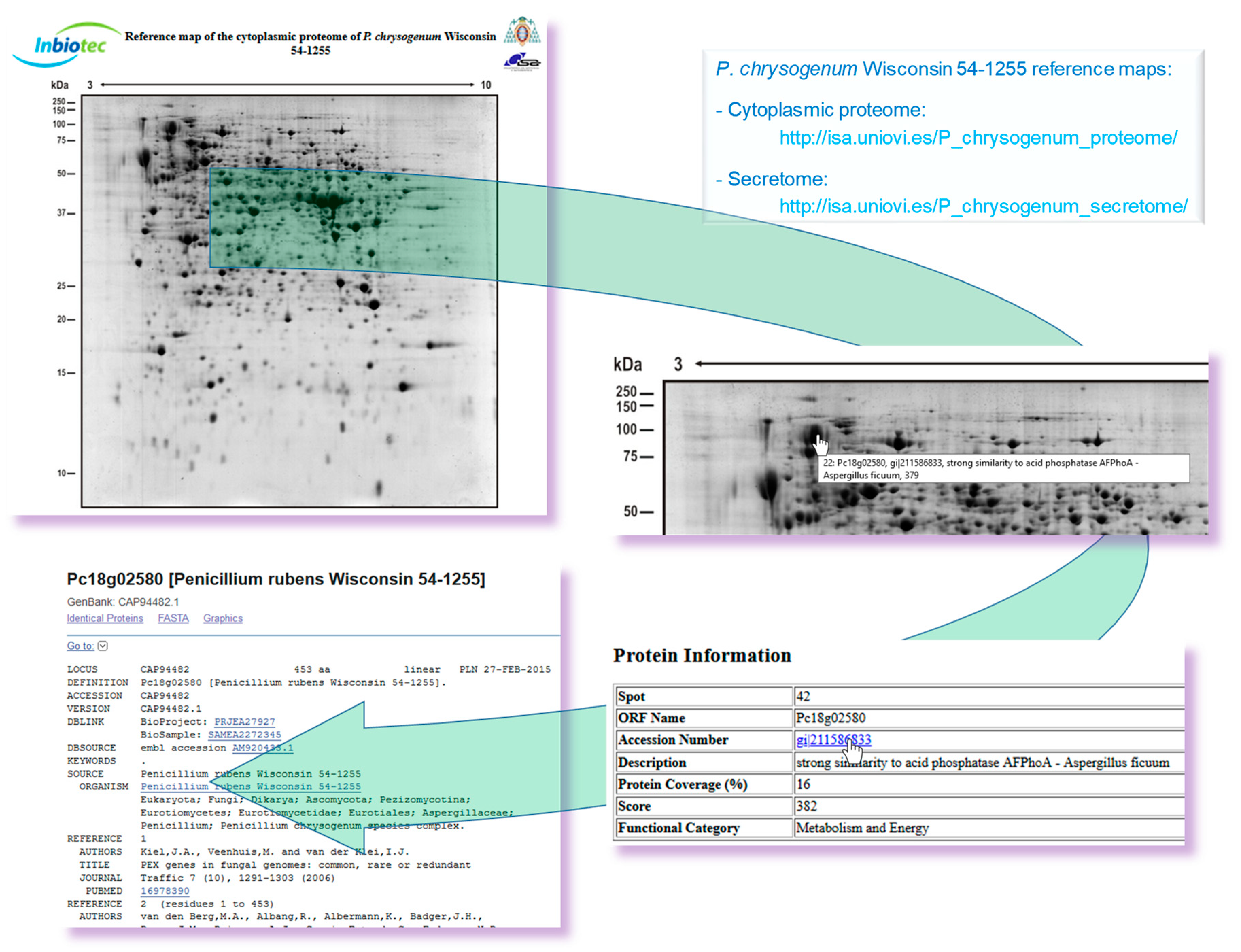
4.1. Reference Maps of Different P. chrysogenum Sub-Proteomes
4.2. Expression Proteomics Analyses
5. Metabolomics
6. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Houbraken, J.; Frisvad, J.C.; Samson, R.A. Fleming’s penicillin producing strain is not Penicillium chrysogenum but P. rubens. IMA Fungus 2011, 2, 87–95. [Google Scholar] [CrossRef] [Green Version]
- Ligon, B.L. Penicillin: Its discovery and early development. Semin. Pediatr. Infect. Dis. 2004, 15, 52–57. [Google Scholar] [CrossRef] [PubMed]
- Barreiro, C.; Garcia-Estrada, C. Proteomics and Penicillium chrysogenum: Unveiling the secrets behind penicillin production. J. Proteom. 2019, 198, 119–131. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Estrada, C.; Martin, J.F. Biosynthetic gene clusters for relevant secondary metabolites produced by Penicillium roqueforti in blue cheeses. Appl. Microbiol. Biotechnol. 2016, 100, 8303–8313. [Google Scholar] [CrossRef] [PubMed]
- Borchert, A. Biotechnology of industrial antibiotics. Hrsg. E. J. Vandamme. 832 S., zahlr. Abb., Marcel Dekker, Inc., New York 1984. Geb. SFr. 299,–. Mater. Corros. 1984, 35, 588. [Google Scholar] [CrossRef]
- Barreiro, C.; Martin, J.F.; Garcia-Estrada, C. Proteomics shows new faces for the old penicillin producer Penicillium chrysogenum. J. Biomed. Biotechnol. 2012, 2012, 105109. [Google Scholar] [CrossRef] [Green Version]
- Martín, J.-F.; García-Estrada, C.; Zeilinger, S. Biosynthesis and Molecular Genetics of Fungal Secondary Metabolites; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar] [CrossRef]
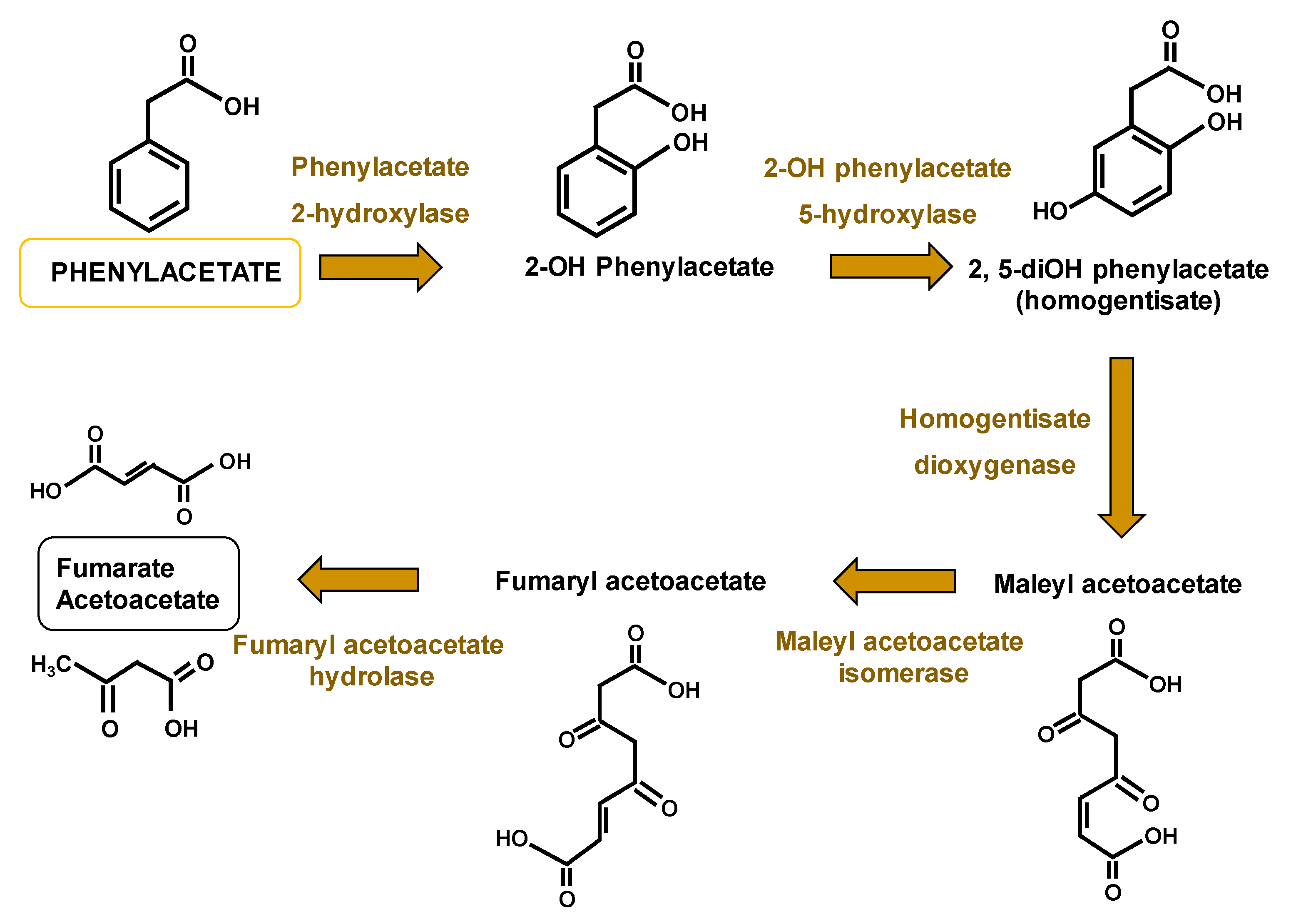
- Fernandez-Canon, J.M.; Penalva, M.A. Molecular characterization of a gene encoding a homogentisate dioxygenase from Aspergillus nidulans and identification of its human and plant homologues. J. Biol. Chem. 1995, 270, 21199–21205. [Google Scholar] [CrossRef] [Green Version]
- Mingot, J.M.; Penalva, M.A.; Fernandez-Canon, J.M. Disruption of phacA, an Aspergillus nidulans gene encoding a novel cytochrome P450 monooxygenase catalyzing phenylacetate 2-hydroxylation, results in penicillin overproduction. J. Biol. Chem. 1999, 274, 14545–14550. [Google Scholar] [CrossRef] [Green Version]
- Arias-Barrau, E.; Olivera, E.R.; Luengo, J.M.; Fernandez, C.; Galan, B.; Garcia, J.L.; Diaz, E.; Minambres, B. The homogentisate pathway: A central catabolic pathway involved in the degradation of L-phenylalanine, L-tyrosine, and 3-hydroxyphenylacetate in Pseudomonas putida. J. Bacteriol. 2004, 186, 5062–5077. [Google Scholar] [CrossRef] [Green Version]
- Brakhage, A.A.; Sprote, P.; Al-Abdallah, Q.; Gehrke, A.; Plattner, H.; Tuncher, A. Regulation of penicillin biosynthesis in filamentous fungi. Adv. Biochem. Eng. Biotechnol. 2004, 88, 45–90. [Google Scholar] [CrossRef]
- Martin, J.F.; Ullan, R.V.; Garcia-Estrada, C. Regulation and compartmentalization of β-lactam biosynthesis. Microb. Biotechnol. 2010, 3, 285–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Estrada, C.; Domínguez, R.; Kosalková, K.; Martin, J. Transcription Factors Controlling Primary and Secondary Metabolism in Filamentous Fungi: The β-Lactam Paradigm. Fermentation 2018, 4, 47. [Google Scholar] [CrossRef] [Green Version]
- Adnan, M.; Zheng, W.; Islam, W.; Arif, M.; Abubakar, Y.S.; Wang, Z.; Lu, G. Carbon Catabolite Repression in Filamentous Fungi. Int. J. Mol. Sci. 2017, 19, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nehlin, J.O.; Ronne, H. Yeast MIG1 repressor is related to the mammalian early growth response and Wilms’ tumour finger proteins. EMBO J. 1990, 9, 2891–2898. [Google Scholar] [CrossRef] [PubMed]
- Martín, J.F.; Casqueiro, J.; Kosalková, K.; Marcos, A.T.; Gutiérrez, S. Penicillin and cephalosporin biosynthesis: Mechanism of carbon catabolite regulation of penicillin production. Antonie Leeuwenhoek 1999, 75, 21–31. [Google Scholar] [CrossRef]
- Cepeda-García, C.; Domínguez-Santos, R.; García-Rico, R.O.; García-Estrada, C.; Cajiao, A.; Fierro, F.; Martín, J.F. Direct involvement of the CreA transcription factor in penicillin biosynthesis and expression of the pcbAB gene in Penicillium chrysogenum. Appl. Microbiol. Biotechnol. 2014, 98, 7113–7124. [Google Scholar] [CrossRef] [Green Version]
- Tilburn, J.; Sarkar, S.; Widdick, D.A.; Espeso, E.A.; Orejas, M.; Mungroo, J.; Peñalva, M.A.; Arst, H.N., Jr. The Aspergillus PacC zinc finger transcription factor mediates regulation of both acid- and alkaline-expressed genes by ambient pH. EMBO J. 1995, 14, 779–790. [Google Scholar] [CrossRef]
- Suárez, T.; Peñalva, M.A. Characterization of a Penicillium chrysogenum gene encoding a PacC transcription factor and its binding sites in the divergent pcbAB-pcbC promoter of the penicillin biosynthetic cluster. Mol. Microbiol. 1996, 20, 529–540. [Google Scholar] [CrossRef]
- Tudzynski, B. Nitrogen regulation of fungal secondary metabolism in fungi. Front. Microbiol. 2014, 5, 656. [Google Scholar] [CrossRef] [Green Version]
- Sánchez, S.; Flores, M.E.; Demain, A.L. Nitrogen regulation of penicillin and cephalosporin fermentation. In Nitrogen Source Control of Microbial Processes; Sanchez-Esquivel, S., Ed.; CRC Press, Inc.: Boca Raton, FL, USA, 1988; pp. 121–136. [Google Scholar]
- Haas, H.; Marzluf, G.A. NRE, the major nitrogen regulatory protein of Penicillium chrysogenum, binds specifically to elements in the intergenic promoter regions of nitrate assimilation and penicillin biosynthetic gene clusters. Curr. Genet. 1995, 28, 177–183. [Google Scholar] [CrossRef]
- Schmitt, E.K.; Kück, U. The fungal CPCR1 protein, which binds specifically to β-lactam biosynthesis genes, is related to human regulatory factor X transcription factors. J. Biol. Chem. 2000, 275, 9348–9357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domínguez-Santos, R.; Martín, J.F.; Kosalková, K.; Prieto, C.; Ullán, R.V.; García-Estrada, C. The regulatory factor PcRFX1 controls the expression of the three genes of β-lactam biosynthesis in Penicillium chrysogenum. Fungal Genet. Biol. 2012, 49, 866–881. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, E.K.; Hoff, B.; Kück, U. AcFKH1, a novel member of the forkhead family, associates with the RFX transcription factor CPCR1 in the cephalosporin C-producing fungus Acremonium chrysogenum. Gene 2004, 342, 269–281. [Google Scholar] [CrossRef] [PubMed]
- Domínguez-Santos, R.; García-Estrada, C.; Kosalková, K.; Prieto, C.; Santamarta, I.; Martín, J.F. PcFKH1, a novel regulatory factor from the forkhead family, controls the biosynthesis of penicillin in Penicillium chrysogenum. Biochimie 2015, 115, 162–176. [Google Scholar] [CrossRef]
- Bok, J.W.; Keller, N.P. LaeA, a Regulator of Secondary Metabolism in Aspergillus spp. Eukaryot. Cell 2004, 3, 527–535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kosalková, K.; García-Estrada, C.; Ullán, R.V.; Godio, R.P.; Feltrer, R.; Teijeira, F.; Mauriz, E.; Martín, J.F. The global regulator LaeA controls penicillin biosynthesis, pigmentation and sporulation, but not roquefortine C synthesis in Penicillium chrysogenum. Biochimie 2009, 91, 214–225. [Google Scholar] [CrossRef]
- Bayram, O.; Krappmann, S.; Ni, M.; Bok, J.W.; Helmstaedt, K.; Valerius, O.; Braus-Stromeyer, S.; Kwon, N.J.; Keller, N.P.; Yu, J.H.; et al. VelB/VeA/LaeA Complex Coordinates Light Signal With Fungal Development and Secondary Metabolism. Science 2008, 320, 1504–1506. [Google Scholar] [CrossRef]
- Bayram, O.; Braus, G.H. Coordination of Secondary Metabolism and Development in Fungi: The Velvet Family of Regulatory Proteins. FEMS Microbiol. Rev. 2012, 36, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Hoff, B.; Kamerewerd, J.; Sigl, C.; Mitterbauer, R.; Zadra, I.; Kürnsteiner, H.; Kück, U. Two components of a velvet-like complex control hyphal morphogenesis, conidiophore development, and penicillin biosynthesis in Penicillium chrysogenum. Eukaryot. Cell 2010, 9, 1236–1250. [Google Scholar] [CrossRef] [Green Version]
- Kopke, K.; Hoff, B.; Bloemendal, S.; Katschorowski, A.; Kamerewerd, J.; Kuck, U. Members of the Penicillium chrysogenum velvet complex play functionally opposing roles in the regulation of penicillin biosynthesis and conidiation. Eukaryot. Cell 2013, 12, 299–310. [Google Scholar] [CrossRef] [Green Version]
- Fierro, F.; Barredo, J.L.; Díez, B.; Gutierrez, S.; Fernández, F.J.; Martín, J.F. The penicillin gene cluster is amplified in tandem repeats linked by conserved hexanucleotide sequences. Proc. Natl. Acad. Sci. USA 1995, 92, 6200–6204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez-Saiz, M.; Barredo, J.L.; Moreno, M.A.; Fernandez-Canon, J.M.; Penalva, M.A.; Diez, B. Reduced function of a phenylacetate-oxidizing cytochrome p450 caused strong genetic improvement in early phylogeny of penicillin-producing strains. J. Bacteriol. 2001, 183, 5465–5471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez-Saiz, M.; Diez, B.; Barredo, J.L. Why did the Fleming strain fail in penicillin industry? Fungal Genet. Biol. 2005, 42, 464–470. [Google Scholar] [CrossRef] [PubMed]
- van den Berg, M.A.; Albang, R.; Albermann, K.; Badger, J.H.; Daran, J.M.; Driessen, A.J.; Garcia-Estrada, C.; Fedorova, N.D.; Harris, D.M.; Heijne, W.H.; et al. Genome sequencing and analysis of the filamentous fungus Penicillium chrysogenum. Nat. Biotechnol. 2008, 26, 1161–1168. [Google Scholar] [CrossRef] [Green Version]
- Martín, J.F. Transport systems, intracellular traffic of intermediates and secretion of b-lactam antibiotics in fungi. Fungal Biol. Biotechnol. 2020, 7, 6. [Google Scholar] [CrossRef] [Green Version]
- Kosalkova, K.; Rodriguez-Saiz, M.; Barredo, J.L.; Martin, J.F. Binding of the PTA1 transcriptional activator to the divergent promoter region of the first two genes of the penicillin pathway in different Penicillium species. Curr. Genet. 2007, 52, 229–237. [Google Scholar] [CrossRef]
- Elander, R.P. Industrial production of β-lactam antibiotics. Appl. Microbiol. Biotechnol. 2003, 61, 385–392. [Google Scholar] [CrossRef]
- Salo, O.V.; Ries, M.; Medema, M.H.; Lankhorst, P.P.; Vreeken, R.J.; Bovenberg, R.A.; Driessen, A.J. Genomic mutational analysis of the impact of the classical strain improvement program on β-lactam producing Penicillium chrysogenum. BMC Genom. 2015, 16, 937. [Google Scholar] [CrossRef] [Green Version]
- Hoff, B.; Pöggeler, S.; Kück, U. Eighty years after its discovery, Fleming’s Penicillium strain discloses the secret of its sex. Eukaryot. Cell 2008, 7, 465–470. [Google Scholar] [CrossRef] [Green Version]
- Lein, J. The Panlabs penicillin strain improvement program. Overproduction Microb. Metab. 1986, 105–139. [Google Scholar]
- Specht, T.; Dahlmann, T.A.; Zadra, I.; Kürnsteiner, H.; Kück, U. Complete Sequencing and Chromosome-Scale Genome Assembly of the Industrial Progenitor Strain P2niaD18 from the Penicillin Producer Penicillium chrysogenum. Genome Announc. 2014, 2, e00577-14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newbert, R.W.; Barton, B.; Greaves, P.; Harper, J.; Turner, G. Analysis of a commercially improved Penicillium chrysogenum strain series: Involvement of recombinogenic regions in amplification and deletion of the penicillin biosynthesis gene cluster. J. Ind. Microbiol. Biotechnol. 1997, 19, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.Q.; Zhong, J.; Zhao, Y.; Xiao, J.; Liu, J.; Dai, M.; Zheng, G.; Zhang, L.; Yu, J.; Wu, J.; et al. Genome sequencing of high-penicillin producing industrial strain of Penicillium chrysogenum. BMC Genom. 2014, 15 (Suppl. 1), S11. [Google Scholar] [CrossRef] [Green Version]
- Fierro, F.; Montenegro, E.; Gutierrez, S.; Martin, J.F. Mutants blocked in penicillin biosynthesis show a deletion of the entire penicillin gene cluster at a specific site within a conserved hexanucleotide sequence. Appl. Microbiol. Biotechnol. 1996, 44, 597–604. [Google Scholar] [CrossRef] [PubMed]
- Veiga, T.; Nijland, J.G.; Driessen, A.J.; Bovenberg, R.A.; Touw, H.; van den Berg, M.A.; Pronk, J.T.; Daran, J.M. Impact of velvet complex on transcriptome and penicillin G production in glucose-limited chemostat cultures of a β-lactam high-producing Penicillium chrysogenum strain. Omics 2012, 16, 320–333. [Google Scholar] [CrossRef] [Green Version]
- Martin, J.F. Key role of LaeA and velvet complex proteins on expression of β-lactam and PR-toxin genes in Penicillium chrysogenum: Cross-talk regulation of secondary metabolite pathways. J. Ind. Microbiol. Biotechnol. 2017, 44, 525–535. [Google Scholar] [CrossRef]
- Peng, Q.; Yuan, Y.; Gao, M.; Chen, X.; Liu, B.; Liu, P.; Wu, Y.; Wu, D. Genomic characteristics and comparative genomics analysis of Penicillium chrysogenum KF-25. BMC Genom. 2014, 15, 144. [Google Scholar] [CrossRef] [Green Version]
- Gujar, V.V.; Fuke, P.; Khardenavis, A.A.; Purohit, H.J. Draft genome sequence of Penicillium chrysogenum strain HKF2, a fungus with potential for production of prebiotic synthesizing enzymes. 3 Biotech 2018, 8, 106. [Google Scholar] [CrossRef]
- Schlickeiser, S.; Pleyer, U. Western, Northern, and Southern Blotting. In Basic Science Techniques in Clinical Practice; Patel, H., Arya, M., Shergill, I., Eds.; Springer: London, UK, 2007; pp. 48–57. [Google Scholar] [CrossRef]
- Southern, E.M. Detection of specific sequences among DNA fragments separated by gel electrophoresis. J. Mol. Biol. 1975, 98, 503–517. [Google Scholar] [CrossRef]
- Alwine, J.C.; Kemp, D.J.; Stark, G.R. Method for detection of specific RNAs in agarose gels by transfer to diazobenzyloxymethyl-paper and hybridization with DNA probes. Proc. Natl. Acad. Sci. USA 1977, 74, 5350–5354. [Google Scholar] [CrossRef] [Green Version]
- Renart, J.; Reiser, J.; Stark, G.R. Transfer of proteins from gels to diazobenzyloxymethyl-paper and detection with antisera: A method for studying antibody specificity and antigen structure. Proc. Natl. Acad. Sci. USA 1979, 76, 3116–3120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Towbin, H.; Staehelin, T.; Gordon, J. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: Procedure and some applications. Proc. Natl. Acad. Sci. USA 1979, 76, 4350–4354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burnette, W.N. “Western blotting”: Electrophoretic transfer of proteins from sodium dodecyl sulfate--polyacrylamide gels to unmodified nitrocellulose and radiographic detection with antibody and radioiodinated protein A. Anal. Biochem. 1981, 112, 195–203. [Google Scholar] [CrossRef]
- DeRisi, J.L.; Iyer, V.R.; Brown, P.O. Exploring the metabolic and genetic control of gene expression on a genomic scale. Science 1997, 278, 680–686. [Google Scholar] [CrossRef] [Green Version]
- Schena, M.; Shalon, D.; Davis, R.W.; Brown, P.O. Quantitative monitoring of gene expression patterns with a complementary DNA microarray. Science 1995, 270, 467–470. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Sepúlveda, B.M.; Hinton, J.C. Functional Transcriptomics for Bacterial Gene Detectives. Microbiol. Spectr. 2018, 6, 1–13. [Google Scholar] [CrossRef]
- Nagalakshmi, U.; Wang, Z.; Waern, K.; Shou, C.; Raha, D.; Gerstein, M.; Snyder, M. The transcriptional landscape of the yeast genome defined by RNA sequencing. Science 2008, 320, 1344–1349. [Google Scholar] [CrossRef] [Green Version]
- Terfehr, D.; Dahlmann, T.A.; Kück, U. Transcriptome analysis of the two unrelated fungal β-lactam producers Acremonium chrysogenum and Penicillium chrysogenum: Velvet-regulated genes are major targets during conventional strain improvement programs. BMC Genom. 2017, 18, 272. [Google Scholar] [CrossRef] [Green Version]
- Priebe, S.; Kreisel, C.; Horn, F.; Guthke, R.; Linde, J. FungiFun2: A comprehensive online resource for systematic analysis of gene lists from fungal species. Bioinformatics 2015, 31, 445–446. [Google Scholar] [CrossRef] [Green Version]
- Ruepp, A.; Zollner, A.; Maier, D.; Albermann, K.; Hani, J.; Mokrejs, M.; Tetko, I.; Güldener, U.; Mannhaupt, G.; Münsterkötter, M.; et al. The FunCat, a functional annotation scheme for systematic classification of proteins from whole genomes. Nucleic Acids Res. 2004, 32, 5539–5545. [Google Scholar] [CrossRef]
- Jami, M.-S.; García-Estrada, C.; Barreiro, C.; Cuadrado, A.-A.; Salehi-Najafabadi, Z.; Martín, J.-F. The Penicillium chrysogenum extracellular proteome. Conversion from a food-rotting strain to a versatile cell factory for white biotechnology. Mol. Cell. Proteom. MCP 2010, 9, 2729–2744. [Google Scholar] [CrossRef] [Green Version]
- Jami, M.-S.; Barreiro, C.; García-Estrada, C.; Martín, J.-F. Proteome analysis of the penicillin producer Penicillium chrysogenum: Characterization of protein changes during the industrial strain improvement. Mol. Cell. Proteom. MCP 2010, 9, 1182–1198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, D.M.; van der Krogt, Z.A.; Klaassen, P.; Raamsdonk, L.M.; Hage, S.; van den Berg, M.A.; Bovenberg, R.A.L.; Pronk, J.T.; Daran, J.-M. Exploring and dissecting genome-wide gene expression responses of Penicillium chrysogenum to phenylacetic acid consumption and penicillinG production. BMC Genom. 2009, 10, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hidalgo, P.I.; Ullan, R.V.; Albillos, S.M.; Montero, O.; Fernandez-Bodega, M.A.; Garcia-Estrada, C.; Fernandez-Aguado, M.; Martin, J.F. Molecular characterization of the PR-toxin gene cluster in Penicillium roqueforti and Penicillium chrysogenum: Cross talk of secondary metabolite pathways. Fungal Genet. Biol. B 2014, 62, 11–24. [Google Scholar] [CrossRef]
- Veiga, T.; Solis-Escalante, D.; Romagnoli, G.; ten Pierick, A.; Hanemaaijer, M.; Deshmukh, A.T.; Wahl, A.; Pronk, J.T.; Daran, J.-M. Resolving phenylalanine metabolism sheds light on natural synthesis of penicillin G in Penicillium chrysogenum. Eukaryot. Cell 2012, 11, 238–249. [Google Scholar] [CrossRef] [Green Version]
- Douma, R.D.; Batista, J.M.; Touw, K.M.; Kiel, J.A.K.W.; Krikken, A.M.; Zhao, Z.; Veiga, T.; Klaassen, P.; Bovenberg, R.A.L.; Daran, J.-M.; et al. Degeneration of penicillin production in ethanol-limited chemostat cultivations of Penicillium chrysogenum: A systems biology approach. BMC Syst. Biol. 2011, 5, 132. [Google Scholar] [CrossRef] [Green Version]
- Novikova, N.D. Review of the Knowledge of Microbial Contamination of the Russian Manned Spacecraft. Microb. Ecol. 2004, 47, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Sathishkumar, Y.; Krishnaraj, C.; Rajagopal, K.; Sen, D.; Lee, Y.S. High throughput de novo RNA sequencing elucidates novel responses in Penicillium chrysogenum under microgravity. Bioprocess Biosyst. Eng. 2016, 39, 223–231. [Google Scholar] [CrossRef]
- Harris, D.M.; Westerlaken, I.; Schipper, D.; van der Krogt, Z.A.; Gombert, A.K.; Sutherland, J.; Raamsdonk, L.M.; van den Berg, M.A.; Bovenberg, R.A.; Pronk, J.T.; et al. Engineering of Penicillium chrysogenum for fermentative production of a novel carbamoylated cephem antibiotic precursor. Metab. Eng. 2009, 11, 125–137. [Google Scholar] [CrossRef]
- Koetsier, M.J.; Gombert, A.K.; Fekken, S.; Bovenberg, R.A.; van den Berg, M.A.; Kiel, J.A.; Jekel, P.A.; Janssen, D.B.; Pronk, J.T.; van der Klei, I.J.; et al. The Penicillium chrysogenum aclA gene encodes a broad-substrate-specificity acyl-coenzyme A ligase involved in activation of adipic acid, a side-chain precursor for cephem antibiotics. Fungal Genet. Biol. 2010, 47, 33–42. [Google Scholar] [CrossRef]
- Veiga, T.; Gombert, A.K.; Landes, N.; Verhoeven, M.D.; Kiel, J.A.; Krikken, A.M.; Nijland, J.G.; Touw, H.; Luttik, M.A.; Van der Toorn, J.C.; et al. Metabolic engineering of β-oxidation in Penicillium chrysogenum for improved semi-synthetic cephalosporin biosynthesis. Metab. Eng. 2012, 14, 437–448. [Google Scholar] [CrossRef] [PubMed]
- Thomson, J.J. XLVII. On rays of positive electricity. Lond. Edinb. Dublin Philos. Mag. J. Sci. 1907, 13, 561–575. [Google Scholar] [CrossRef] [Green Version]
- McLafferty, F.W. A century of progress in molecular mass spectrometry. Annu. Rev. Anal. Chem. 2011, 4, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Iii, J. A century of mass spectrometry: From atoms to proteomes. Nat. Methods Nat. Methods 2011, 8, 633–637. [Google Scholar] [CrossRef]
- James, P. Protein identification in the post-genome era: The rapid rise of proteomics. Q. Rev. Biophys. 1997, 30, 279–331. [Google Scholar] [CrossRef] [Green Version]
- O’Farrell, P.H. The pre-omics era: The early days of two-dimensional gels. Proteomics 2008, 8, 4842–4852. [Google Scholar] [CrossRef] [Green Version]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- O’Farrell, P.H. High resolution two-dimensional electrophoresis of proteins. J. Biol. Chem. 1975, 250, 4007–4021. [Google Scholar]
- Kiel, J.A.; van den Berg, M.A.; Fusetti, F.; Poolman, B.; Bovenberg, R.A.; Veenhuis, M.; van der Klei, I.J. Matching the proteome to the genome: The microbody of penicillin-producing Penicillium chrysogenum cells. Funct. Integr. Genom. 2009, 9, 167–184. [Google Scholar] [CrossRef] [Green Version]
- Cheng, J.S.; Zhao, Y.; Qiao, B.; Lu, H.; Chen, Y.; Yuan, Y.J. Comprehensive Profiling of Proteome Changes Provide Insights of Industrial Penicillium chrysogenum During Pilot and Industrial Penicillin G Fermentation. Appl. Biochem. Biotechnol. 2016, 179, 788–804. [Google Scholar] [CrossRef]
- Wilkins, M.R.; Sanchez, J.C.; Gooley, A.A.; Appel, R.D.; Humphery-Smith, I.; Hochstrasser, D.F.; Williams, K.L. Progress with proteome projects: Why all proteins expressed by a genome should be identified and how to do it. Biotechnol. Genet. Eng. Rev. 1996, 13, 19–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barreiro, C. (Ed.) Methods in Proteomics Applied to Corynebacterium glutamicum. In New Trends in Corynebacterium glutamicum: Beyond the Amino Acids, Chapter: 3; Nova Science Publishers, Inc.: New York, NY, USA, 2015; pp. 45–68. [Google Scholar]
- Chandrasekhar, K.; Dileep, A.; Lebonah, D.; Jasti, P. A Short Review on Proteomics and its Applications. Int. Lett. Nat. Sci. 2014, 17, 77–84. [Google Scholar] [CrossRef]
- Jami, M.S.; Martin, J.F.; Barreiro, C.; Dominguez-Santos, R.; Vasco-Cardenas, M.F.; Pascual, M.; Garcia-Estrada, C. Catabolism of phenylacetic acid in Penicillium rubens. Proteome-wide analysis in response to the benzylpenicillin side chain precursor. J. Proteom. 2018, 187, 243–259. [Google Scholar] [CrossRef] [PubMed]
- Kosalkova, K.; Garcia-Estrada, C.; Barreiro, C.; Florez, M.G.; Jami, M.S.; Paniagua, M.A.; Martin, J.F. Casein phosphopeptides drastically increase the secretion of extracellular proteins in Aspergillus awamori. Proteomics studies reveal changes in the secretory pathway. Microb Cell Fact 2012, 11, 5. [Google Scholar] [CrossRef] [Green Version]
- Dominguez-Santos, R.; Kosalkova, K.; Garcia-Estrada, C.; Barreiro, C.; Ibanez, A.; Morales, A.; Martin, J.F. Casein phosphopeptides and CaCl2 increase penicillin production and cause an increment in microbody/peroxisome proteins in Penicillium Chrysogenum. J. Proteom. 2017, 156, 52–62. [Google Scholar] [CrossRef]
- Garcia-Estrada, C.; Barreiro, C.; Jami, M.S.; Martin-Gonzalez, J.; Martin, J.F. The inducers 1,3-diaminopropane and spermidine cause the reprogramming of metabolism in Penicillium chrysogenum, leading to multiple vesicles and penicillin overproduction. J. Proteom. 2013, 85, 129–159. [Google Scholar] [CrossRef]
- Barreiro, C.; García-Estrada, C.; Martin, J. Proteomics Methodology Applied to the Analysis of Filamentous Fungi - New Trends for an Impressive Diverse Group of Organisms. In Tandem Mass Spectrometry—Applications and Principles; Harn, G.L., Prasain, J., Eds.; Intech-Open Access Publisher: London, UK, 2012; pp. 127–160. [Google Scholar] [CrossRef] [Green Version]
- Barreiro, C.; Morales, A.; Vazquez-Iglesias, I.; Sola-Landa, A. Intra- and Extra-cellular Proteome Analyses of Steroid-Producer Mycobacteria. Methods Mol. Biol. 2017, 1645, 73–92. [Google Scholar] [CrossRef]
- Chang, K.Y.; Georgianna, D.R.; Heber, S.; Payne, G.A.; Muddiman, D.C. Detection of alternative splice variants at the proteome level in Aspergillus flavus. J. Proteome Res. 2010, 9, 1209–1217. [Google Scholar] [CrossRef] [Green Version]
- Wright, J.C.; Sugden, D.; Francis-McIntyre, S.; Riba-Garcia, I.; Gaskell, S.J.; Grigoriev, I.V.; Baker, S.E.; Beynon, R.J.; Hubbard, S.J. Exploiting proteomic data for genome annotation and gene model validation in Aspergillus niger. BMC Genom. 2009, 10, 61. [Google Scholar] [CrossRef] [Green Version]
- García-Estrada, C.; Martín, J.-F. Penicillins. In Biosynthesis and Molecular Genetics of Fungal Secondary Metabolites; Martín, J.-F., García-Estrada, C., Zeilinger, S., Eds.; Springer: New York, NY, USA, 2014; pp. 17–42. [Google Scholar] [CrossRef]
- Baker, S.E. Protein hyperproduction in fungi by design. Appl. Microbiol. Biotechnol. 2018, 102, 8621–8628. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Calvo, L.; Ullan, R.V.; Fernandez-Aguado, M.; Garcia-Lino, A.M.; Balana-Fouce, R.; Barreiro, C. Secreted protein extract analyses present the plant pathogen Alternaria alternata as a suitable industrial enzyme toolbox. J. Proteom. 2018, 177, 48–64. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.-T.; Jeon, J.; Choi, J.; Cheong, K.; Song, H.; Choi, G.; Kang, S.; Lee, Y.-H. Kingdom-Wide Analysis of Fungal Small Secreted Proteins (SSPs) Reveals their Potential Role in Host Association. Front. Plant Sci. 2016, 7, 186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bendtsen, J.D.; Nielsen, H.; von Heijne, G.; Brunak, S. Improved prediction of signal peptides: SignalP 3.0. J. Mol. Biol. 2004, 340, 783–795. [Google Scholar] [CrossRef] [PubMed]
- Bendtsen, J.D.; Jensen, L.J.; Blom, N.; Von Heijne, G.; Brunak, S. Feature-based prediction of non-classical and leaderless protein secretion. Protein Eng. Des. Sel. Peds 2004, 17, 349–356. [Google Scholar] [CrossRef] [Green Version]
- Raper, K.B. THE DEVELOPMENT OF IMPROVED PENICILLIN-PRODUCING MOLDS. Ann. N. Y. Acad. Sci. 1946, 48, 41–56. [Google Scholar] [CrossRef]
- Barreiro, C.; Martínez-Castro, M. Trends in the biosynthesis and production of the immunosuppressant tacrolimus (FK506). Appl. Microbiol. Biotechnol. 2013, 98. [Google Scholar] [CrossRef]
- Society, A.C. Penicillin Production through Deep-tank Fermentation. Available online: https://0-www-acs-org.brum.beds.ac.uk/content/acs/en/education/whatischemistry/landmarks/penicillin.html (accessed on 12 September 2018).
- Barreiro, C.; Martinez-Castro, M. Regulation of the phosphate metabolism in Streptomyces genus: Impact on the secondary metabolites. Appl. Microbiol. Biotechnol. 2019, 103, 1643–1658. [Google Scholar] [CrossRef]
- Macheleidt, J.; Mattern, D.J.; Fischer, J.; Netzker, T.; Weber, J.; Schroeckh, V.; Valiante, V.; Brakhage, A.A. Regulation and Role of Fungal Secondary Metabolites. Annu. Rev. Genet. 2016, 50, 371–392. [Google Scholar] [CrossRef]
- Brodhagen, M.; Keller, N.P. Signalling pathways connecting mycotoxin production and sporulation. Mol. Plant Pathol. 2006, 7, 285–301. [Google Scholar] [CrossRef]
- Garcia-Rico, R.O.; Martin, J.F.; Fierro, F. The pga1 gene of Penicillium chrysogenum NRRL 1951 encodes a heterotrimeric G protein α subunit that controls growth and development. Res. Microbiol. 2007, 158, 437–446. [Google Scholar] [CrossRef]
- Garcia-Rico, R.O.; Fierro, F.; Mauriz, E.; Gomez, A.; Fernandez-Bodega, M.A.; Martin, J.F. The heterotrimeric Galpha protein pga1 regulates biosynthesis of penicillin, chrysogenin and roquefortine in Penicillium chrysogenum. Microbiology 2008, 154, 3567–3578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Rico, R.O.; Martin, J.F.; Fierro, F. Heterotrimeric Galpha protein Pga1 from Penicillium chrysogenum triggers germination in response to carbon sources and affects negatively resistance to different stress conditions. Fungal Genet. Biol. B 2011, 48, 641–649. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Rico, R.O.; Fierro, F.; Martin, J.F. Heterotrimeric Galpha protein Pga1 of Penicillium chrysogenum controls conidiation mainly by a cAMP-independent mechanism. Biochem. Cell Biol. Biochim. Biol. Cell. 2008, 86, 57–69. [Google Scholar] [CrossRef] [PubMed]
- Carrasco-Navarro, U.; Vera-Estrella, R.; Barkla, B.J.; Zuniga-Leon, E.; Reyes-Vivas, H.; Fernandez, F.J.; Fierro, F. Proteomic analysis of the signaling pathway mediated by the heterotrimeric Galpha protein Pga1 of Penicillium Chrysogenum. Microb. Cell Fact. 2016, 15, 173. [Google Scholar] [CrossRef] [Green Version]
- Unlu, M.; Morgan, M.E.; Minden, J.S. Difference gel electrophoresis: A single gel method for detecting changes in protein extracts. Electrophoresis 1997, 18, 2071–2077. [Google Scholar] [CrossRef]
- Timms, J.F.; Cramer, R. Difference gel electrophoresis. Proteomics 2008, 8, 4886–4897. [Google Scholar] [CrossRef]
- Martin, J.; Garcia-Estrada, C.; Rumbero, A.; Recio, E.; Albillos, S.M.; Ullan, R.V.; Martin, J.F. Characterization of an autoinducer of penicillin biosynthesis in Penicillium chrysogenum. Appl. Environ. Microbiol. 2011, 77, 5688–5696. [Google Scholar] [CrossRef] [Green Version]
- Martin, J.; Garcia-Estrada, C.; Kosalkova, K.; Ullan, R.V.; Albillos, S.M.; Martin, J.F. The inducers 1,3-diaminopropane and spermidine produce a drastic increase in the expression of the penicillin biosynthetic genes for prolonged time, mediated by the laeA regulator. Fungal Genet. Biol. B 2012, 49, 1004–1013. [Google Scholar] [CrossRef]
- Roze, L.V.; Chanda, A.; Linz, J.E. Compartmentalization and molecular traffic in secondary metabolism: A new understanding of established cellular processes. Fungal Genet. Biol. B 2011, 48, 35–48. [Google Scholar] [CrossRef] [Green Version]
- Martin, J.F.; Garcia-Estrada, C.; Ullan, R.V. Transport of substrates into peroxisomes: The paradigm of β-lactam biosynthetic intermediates. Biomol. Concepts 2013, 4, 197–211. [Google Scholar] [CrossRef]
- Moyer, A.J.; Coghill, R.D. Penicillin: X. The Effect of Phenylacetic Acid on Penicillin Production. J. Bacteriol. 1947, 53, 329–341. [Google Scholar] [CrossRef] [Green Version]
- Ferrer-Sevillano, F.; Fernandez-Canon, J.M. Novel phacB-encoded cytochrome P450 monooxygenase from Aspergillus nidulans with 3-hydroxyphenylacetate 6-hydroxylase and 3,4-dihydroxyphenylacetate 6-hydroxylase activities. Eukaryot. Cell 2007, 6, 514–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerke, J.; Bayram, O.; Braus, G.H. Fungal S-adenosylmethionine synthetase and the control of development and secondary metabolism in Aspergillus nidulans. Fungal Genet. Biol. B 2012, 49, 443–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thykaer, J.; Rueksomtawin, K.; Noorman, H.; Nielsen, J. NADPH-dependent glutamate dehydrogenase in Penicillium chrysogenum is involved in regulation of β-lactam production. Microbiology 2008, 154, 1242–1250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, J.F. Calcium-containing phosphopeptides pave the secretory pathway for efficient protein traffic and secretion in fungi. Microb. Cell Fact. 2014, 13, 117. [Google Scholar] [CrossRef] [PubMed]
- El-Marsafy, M.; Abdel-Akher, M.; El-Saied, H. Effect of media composition on the penicillin production. Zentralblatt fur Bakteriologie, Parasitenkunde, Infektionskrankheiten und Hygiene. Zweite Nat. Abt. Allg. Landwirtsch. Tech. Mikrobiol. 1977, 132, 117–122. [Google Scholar] [CrossRef]
- Lara, F.; del Carmen Mateos, R.; Vazquez, G.; Sanchez, S. Induction of penicillin biosynthesis by L-glutamate in Penicillium chrysogenum. Biochem. Biophys. Res. Commun. 1982, 105, 172–178. [Google Scholar] [CrossRef]
- Nasution, U.; van Gulik, W.M.; Ras, C.; Proell, A.; Heijnen, J.J. A metabolome study of the steady-state relation between central metabolism, amino acid biosynthesis and penicillin production in Penicillium chrysogenum. Metab. Eng. 2008, 10, 10–23. [Google Scholar] [CrossRef]
- Guo, M.; Zhang, X.; Li, M.; Li, T.; Duan, X.; Zhang, D.; Hu, L.; Huang, R. Label-Free Proteomic Analysis of Molecular Effects of 2-Methoxy-1,4-naphthoquinone on Penicillium italicum. Int. J. Mol. Sci. 2019, 20, 3459. [Google Scholar] [CrossRef] [Green Version]
- Delgado, J.; Nunez, F.; Asensio, M.A.; Owens, R.A. Quantitative proteomic profiling of ochratoxin A repression in Penicillium nordicum by protective cultures. Int. J. Food Microbiol. 2019, 305, 108243. [Google Scholar] [CrossRef]
- Zhou, T.; Wang, X.; Luo, J.; Ye, B.; Zhou, Y.; Zhou, L.; Lai, T. Identification of differentially expressed genes involved in spore germination of Penicillium expansum by comparative transcriptome and proteome approaches. Microbiologyopen 2018, 7, e00562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daviss, B. Growing Pains for Metabolomics. Scientist 2005, 19, 25–28. [Google Scholar]
- Hollywood, K.; Brison, D.R.; Goodacre, R. Metabolomics: Current technologies and future trends. Proteomics 2006, 6, 4716–4723. [Google Scholar] [CrossRef] [PubMed]
- Gika, H.G.; Theodoridis, G.A.; Wingate, J.E.; Wilson, I.D. Within-day reproducibility of an HPLC-MS-based method for metabonomic analysis: Application to human urine. J. Proteome Res. 2007, 6, 3291–3303. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Chu, J.; Zhuang, Y.; Gulik, W.; Noorman, H. A dynamic model-based preparation of uniformly-13C-labeled internal standards facilitates quantitative metabolomics analysis of Penicillium chrysogenum. J. Biotechnol. 2019, 299. [Google Scholar] [CrossRef] [PubMed]
- Mashego, M.R.; Wu, L.; Van Dam, J.C.; Ras, C.; Vinke, J.L.; Van Winden, W.A.; Van Gulik, W.M.; Heijnen, J.J. MIRACLE: Mass isotopomer ratio analysis of U-13C-labeled extracts. A new method for accurate quantification of changes in concentrations of intracellular metabolites. Biotechnol. Bioeng. 2004, 85, 620–628. [Google Scholar] [CrossRef]
- Jang, C.; Chen, L.; Rabinowitz, J.D. Metabolomics and Isotope Tracing. Cell 2018, 173, 822–837. [Google Scholar] [CrossRef]
- Song, P.; Yuan, K.; Qin, T.; Zhang, K.; Ji, X.J.; Ren, L.; Guan, R.; Wen, J.; Huang, H. Metabolomics profiling reveals the mechanism of increased pneumocandin B0 production by comparing mutant and parent strains. J. Ind. Microbiol. Biotechnol. 2018, 45, 767–780. [Google Scholar] [CrossRef]
- Ding, M.-Z.; Cheng, J.-S.; Xiao, W.-H.; Qiao, B.; Yuan, Y.-J. Comparative metabolomic analysis on industrial continuous and batch ethanol fermentation processes by GC-TOF-MS. Metabolomics 2008, 5, 229–238. [Google Scholar] [CrossRef]
- Zheng, X.; Yu, J.; Cairns, T.C.; Zhang, L.; Zhang, Z.; Zhang, Q.; Zheng, P.; Sun, J.; Ma, Y. Comprehensive Improvement of Sample Preparation Methodologies Facilitates Dynamic Metabolomics of Aspergillus niger. Biotechnol. J. 2019, 14, e1800315. [Google Scholar] [CrossRef]
- Wang, G.; Zhao, J.; Wang, X.; Wang, T.; Zhuang, Y.; Chu, J.; Zhang, S.; Noorman, H.J. Quantitative metabolomics and metabolic flux analysis reveal impact of altered trehalose metabolism on metabolic phenotypes of Penicillium chrysogenum in aerobic glucose-limited chemostats. Biochem. Eng. J. 2019, 146, 41–51. [Google Scholar] [CrossRef]
- Martin, J.F.; van den Berg, M.A.; Ver Loren van Themaat, E.; Liras, P. Sensing and transduction of nutritional and chemical signals in filamentous fungi: Impact on cell development and secondary metabolites biosynthesis. Biotechnol. Adv. 2019, 37, 107392. [Google Scholar] [CrossRef] [PubMed]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Estrada, C.; Martín, J.F.; Cueto, L.; Barreiro, C. Omics Approaches Applied to Penicillium chrysogenum and Penicillin Production: Revealing the Secrets of Improved Productivity. Genes 2020, 11, 712. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11060712
García-Estrada C, Martín JF, Cueto L, Barreiro C. Omics Approaches Applied to Penicillium chrysogenum and Penicillin Production: Revealing the Secrets of Improved Productivity. Genes. 2020; 11(6):712. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11060712
Chicago/Turabian StyleGarcía-Estrada, Carlos, Juan F. Martín, Laura Cueto, and Carlos Barreiro. 2020. "Omics Approaches Applied to Penicillium chrysogenum and Penicillin Production: Revealing the Secrets of Improved Productivity" Genes 11, no. 6: 712. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11060712