Shedding the Light on Litopenaeus vannamei Differential Muscle and Hepatopancreas Immune Responses in White Spot Syndrome Virus (WSSV) Exposure

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Material and Methods
2.1. Sampling and WSSV Exposure
2.2. Library Construction, Sequencing, and Trimming
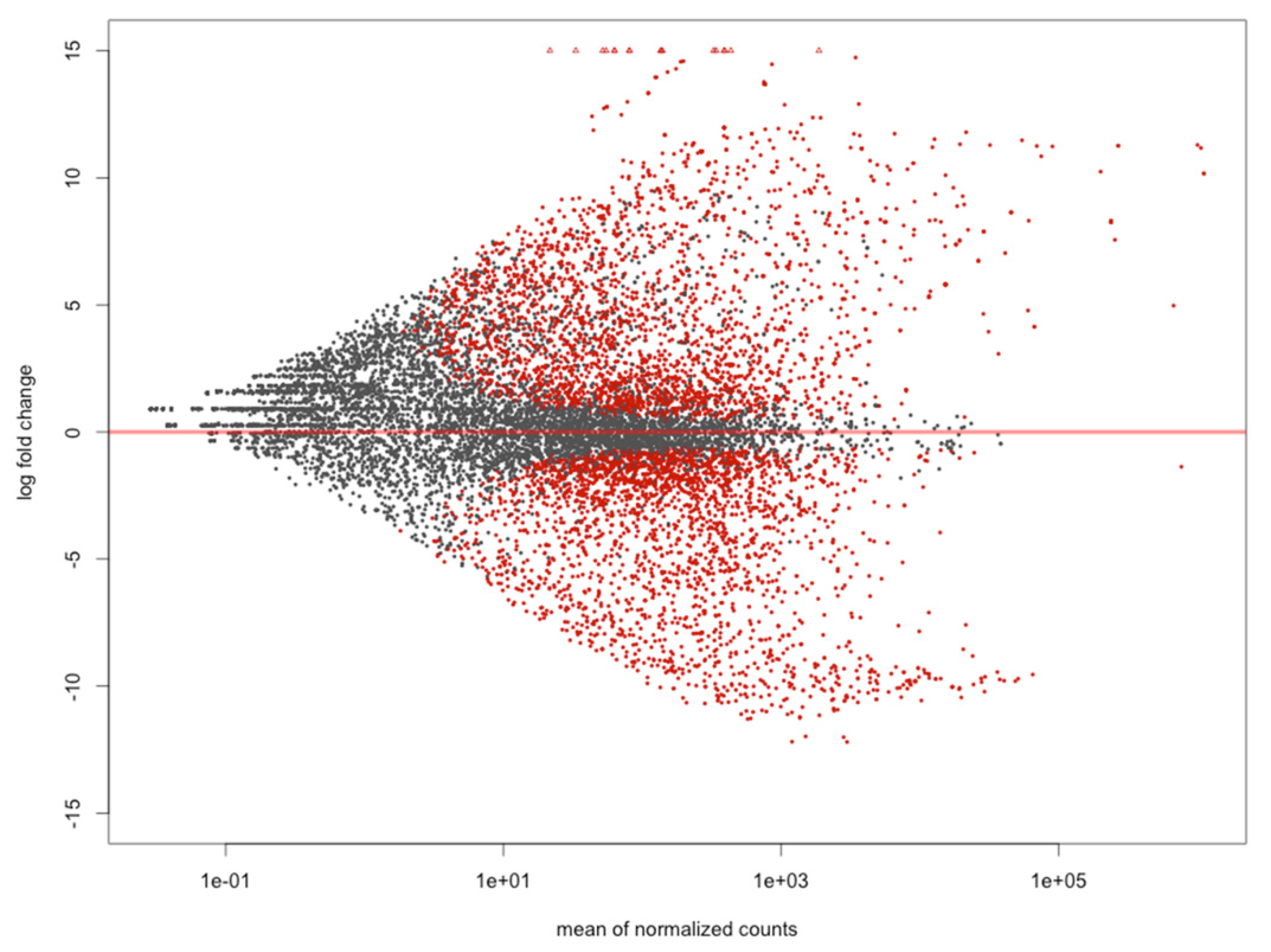
2.3. Mapping and Differentially Expressed Genes Analyses
2.4. Functional Annotation and Enrichment in Gene Ontology (GO)
2.5. DEGs Protein-Protein Interactions (PPI) Networks
3. Results
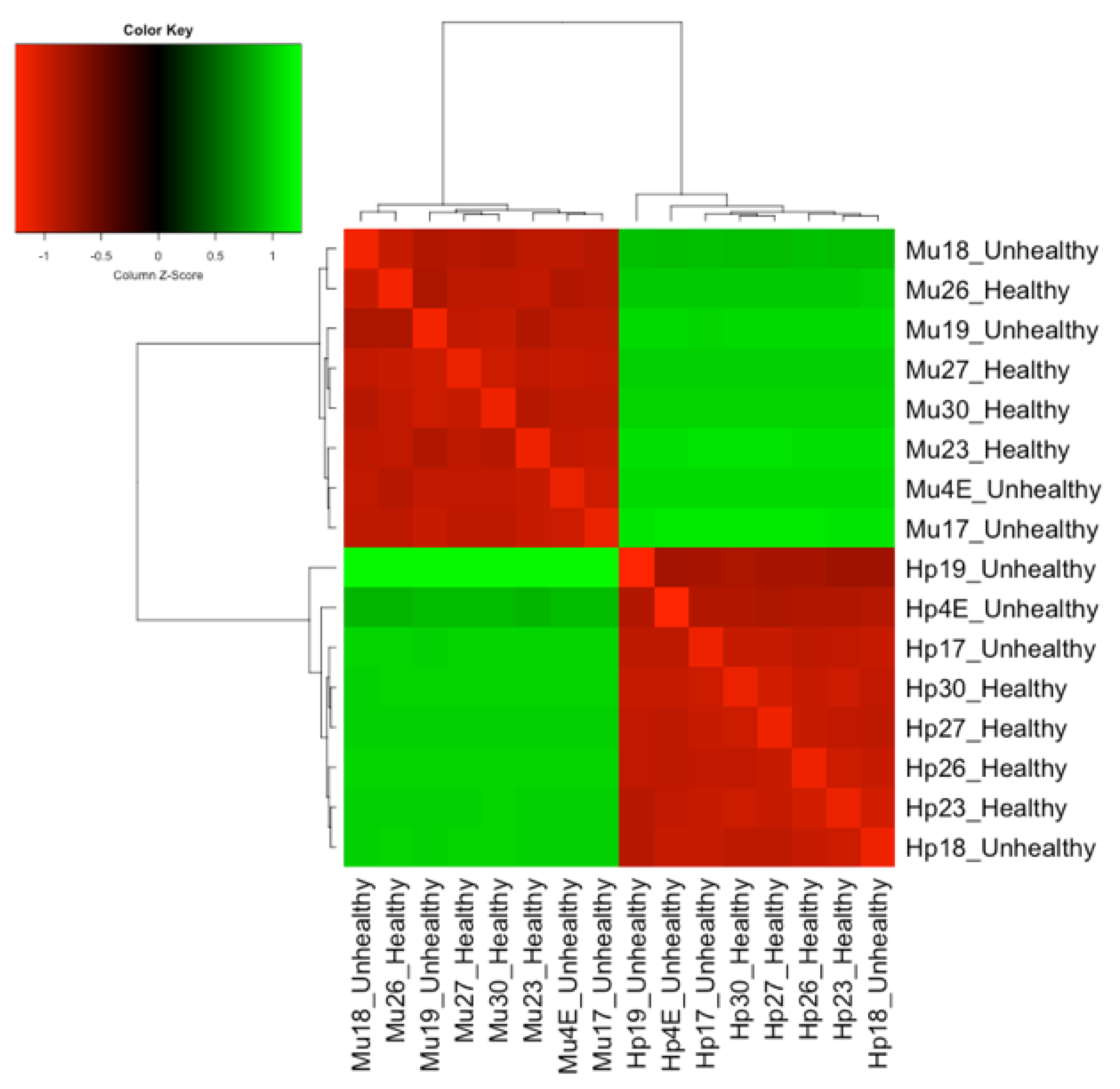
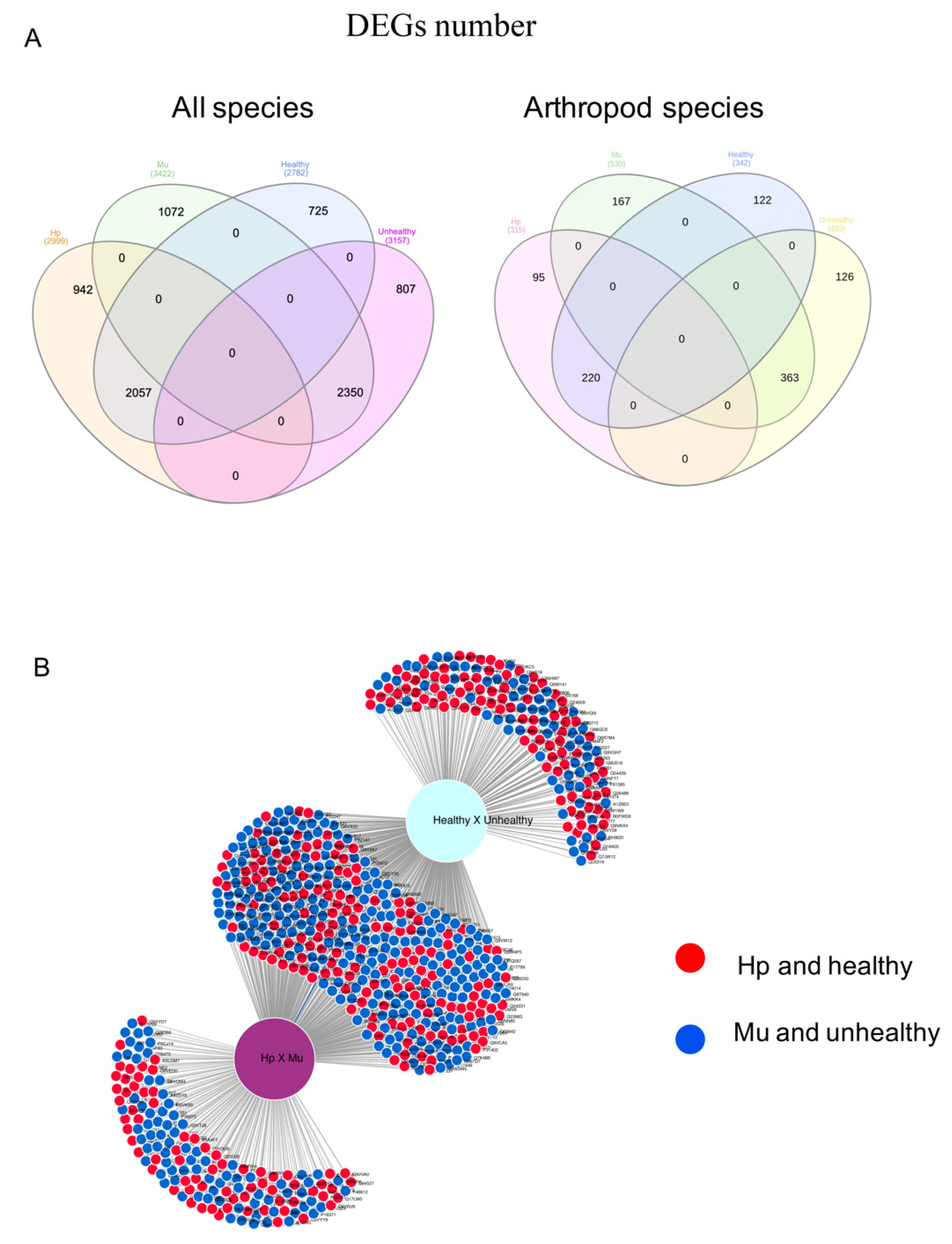
3.1. Differentially Expressed Genes between Hepatopancreas and Muscle (Hp x Mu) Tissues
3.2. Multi-Factor Analysis of Differentially Expressed Genes between Healthy and Unhealthy (He x Un) Conditions
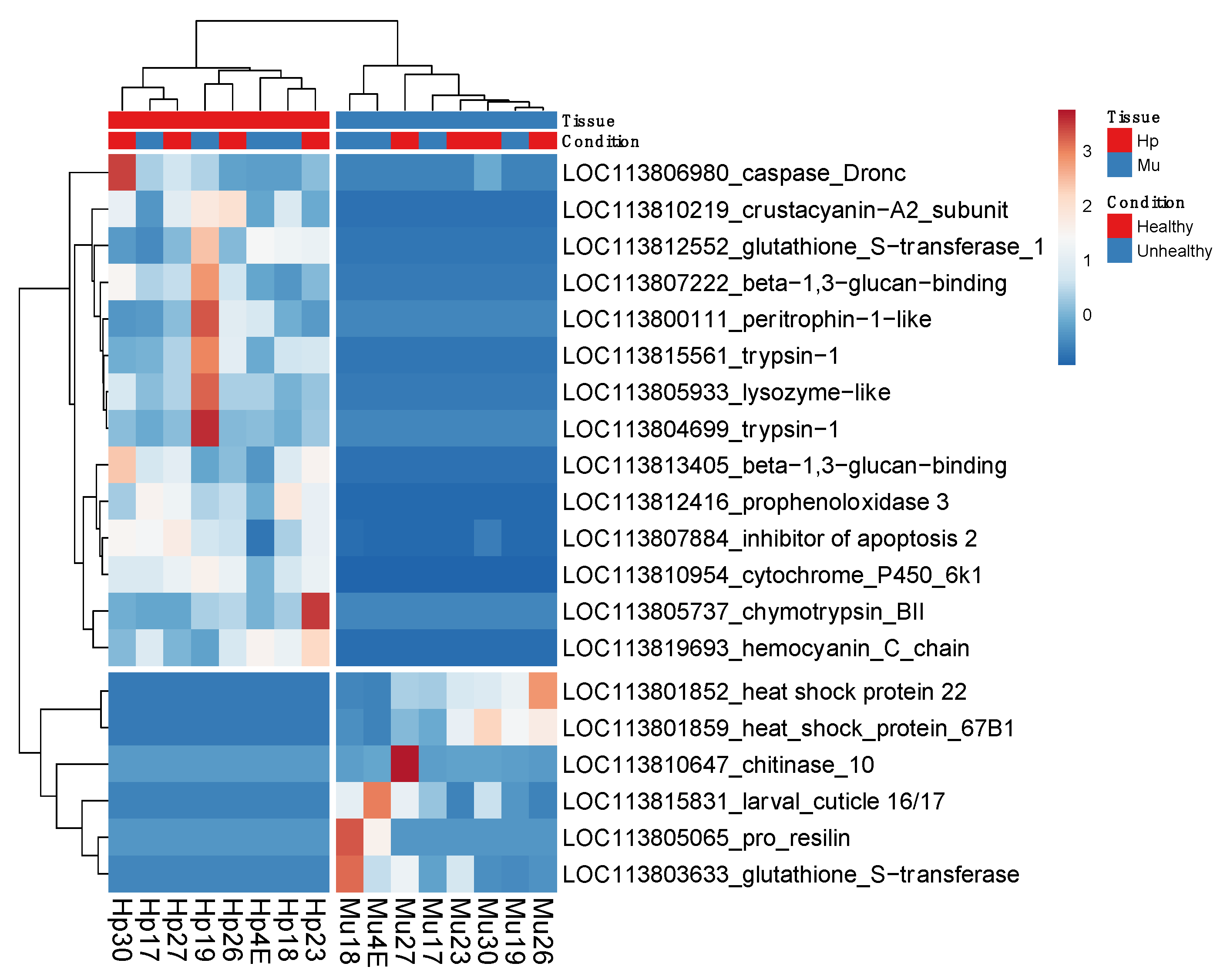
3.3. Common Genes between Tissues and Conditions
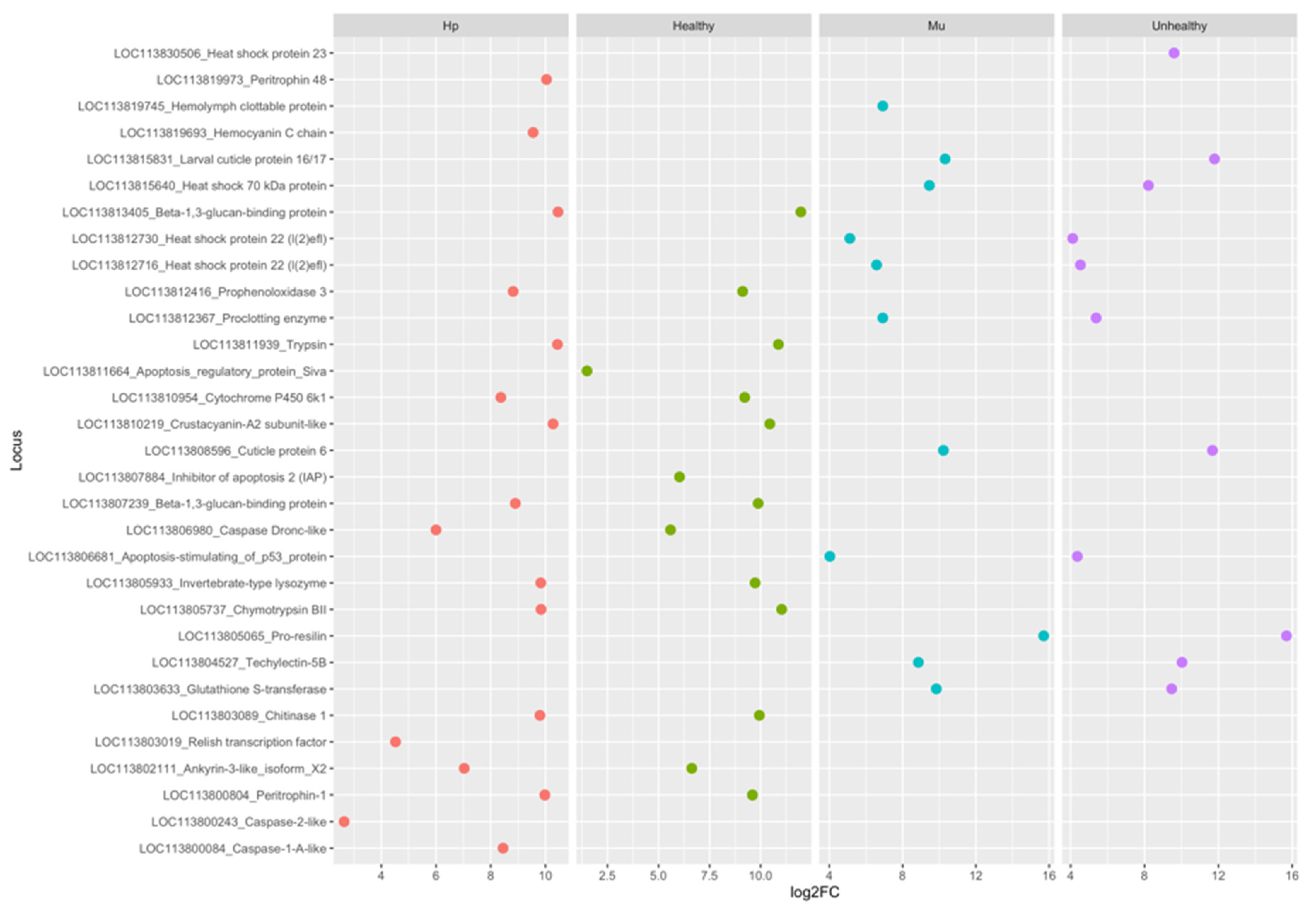
3.4. Protein-Protein Interaction (PPI) Networks for the Most Up-Regulated Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
Ethics Statement
References
- Maciel, J.C.; Francisco, C.J.; Miranda-Filho, K.C. Compensatory growth and feed restriction in marine shrimp production, with emphasis on biofloc technology. Aquacult. Int. 2018, 26, 203–212. [Google Scholar] [CrossRef] [Green Version]
- Fan, L.; Wang, A.; Miao, Y.; Liao, S.; Ye, C.; Lin, Q. Comparative proteomic identification of the hepatopancreas response to cold stress in white shrimp, Litopenaeus vannamei. Aquaculture 2016, 454, 27–34. [Google Scholar] [CrossRef]
- Perez-Enriquez, R.; Hernández-Martínez, F.; Cruz, P. Genetic diversity status of White shrimp Penaeus (Litopenaeus) vannamei broodstock in Mexico. Aquaculture 2009, 297, 44–50. [Google Scholar] [CrossRef]
- Wang, F.; Li, S.; Xiang, J.; Li, F. Transcriptome analysis reveals the activation of neuroendocrine-immune system in shrimp hemocytes at the early stage of WSSV infection. BMC Genom. 2019, 20, 247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peruzza, L.; Shekhar, M.S.; Kumar, K.V.; Swathi, A.; Karthic, K.; Hauton, C.; Vijayan, K.K. Temporal changes in transcriptome profile provide insights of White Spot Syndrome Virus infection in Litopenaeus vannamei. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Sookruksawong, S.; Sun, F.; Liu, Z.; Tassanakajon, A. RNA-Seq analysis reveals genes associated with resistance to Taura syndrome virus (TSV) in the Pacific white shrimp Litopenaeus vannamei. Dev. Comp. Immunol. 2013, 41, 523–533. [Google Scholar] [CrossRef]
- Zeng, D.; Chen, X.; Xie, D.; Zhao, Y.; Yang, C.; Li, Y.; Ma, N.; Peng, M.; Yang, Q.; Liao, Z.; et al. Transcriptome Analysis of Pacific White Shrimp (Litopenaeus vannamei) Hepatopancreas in Response to Taura Syndrome Virus (TSV) Experimental Infection. PLoS ONE 2013, 8, e57515. [Google Scholar] [CrossRef]
- Kanokudom, S.; Prateeprat, T.; Attasart, P.; Roytrakul, S.; Panyim, S.; Smith, D.R.; Assavalapsakul, W. In vitro neutralization of yellow head virus infection in shrimp using recombinant PmYRP65 protein. Aquaculture 2018, 486, 266–270. [Google Scholar] [CrossRef]
- Tassanakajon, A.; Somboonwiwat, K.; Supungul, P.; Tang, S. Discovery of immune molecules and their crucial functions in shrimp immunity. Fish Shellfish Immunol. 2013, 34, 954–967. [Google Scholar] [CrossRef]
- Wang, P.-H.; Gu, Z.-H.; Huang, X.-D.; Liu, B.-D.; Deng, X.; Ai, H.-S.; Wang, J.; Yin, Z.-X.; Weng, S.-P.; Yu, X.-Q.; et al. An immune deficiency homolog from the white shrimp, Litopenaeus vannamei, activates antimicrobial peptide genes. Mol. Immunol. 2009, 46, 1897–1904. [Google Scholar] [CrossRef]
- Qian, Z.; Liu, T.; Liu, Q.; He, S.; Liu, Y.; Hou, F.; Wang, X.; Mi, X.; Cai, C.; Liu, X. p53 is involved in shrimp survival via its regulation roles on MnSOD and GPx in response to acute environmental stresses. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2014, 159, 38–51. [Google Scholar] [CrossRef] [PubMed]
- Rewitz, K.F.; Styrishave, B.; Løbner-Olesen, A.; Andersen, O. Marine invertebrate cytochrome P450: Emerging insights from vertebrate and insect analogies. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2006, 143, 363–381. [Google Scholar] [CrossRef]
- Ai, H.-S.; Huang, Y.-C.; Li, S.-D.; Weng, S.-P.; Yu, X.-Q.; He, J.-G. Characterization of a prophenoloxidase from hemocytes of the shrimp Litopenaeus vannamei that is down-regulated by white spot syndrome virus. Fish Shellfish Immunol. 2008, 25, 28–39. [Google Scholar] [CrossRef] [PubMed]
- Cerenius, L.; Soderhall, K. The prophenoloxidase-activating system in invertebrates. Immunol. Rev. 2004, 198, 116–126. [Google Scholar] [CrossRef] [PubMed]
- Cheng, W.; Tsai, I.-H.; Huang, C.-J.; Chiang, P.-C.; Cheng, C.-H.; Yeh, M.-S. Cloning and characterization of hemolymph clottable proteins of kuruma prawn (Marsupenaeus japonicus) and white shrimp (Litopenaeus vannamei). Dev. Comp. Immunol. 2008, 32, 265–274. [Google Scholar] [CrossRef]
- Zhong, S.; Mao, Y.; Wang, J.; Liu, M.; Zhang, M.; Su, Y. Transcriptome analysis of Kuruma shrimp (Marsupenaeus japonicus) hepatopancreas in response to white spot syndrome virus (WSSV) under experimental infection. Fish Shellfish Immunol. 2017, 70, 710–719. [Google Scholar] [CrossRef] [PubMed]
- Kongton, K.; Phongdara, A.; Tonganunt-Srithaworn, M.; Wanna, W. Molecular cloning and expression analysis of the interferon-γ-inducible lysosomal thiol reductase gene from the shrimp Penaeus monodon. Mol. Biol. Rep. 2011, 38, 3463–3470. [Google Scholar] [CrossRef] [PubMed]
- Nehyba, J.; Hrdličková, R.; Bose, H.R. Dynamic Evolution of Immune System Regulators: The History of the Interferon Regulatory Factor Family. Mol. Biol. Evol. 2009, 26, 2539–2550. [Google Scholar] [CrossRef]
- Huang, T.; Zhang, X. Host defense against DNA virus infection in shrimp is mediated by the siRNA pathway. Eur. J. Immunol. 2013, 43, 137–146. [Google Scholar] [CrossRef]
- Xu, J.; Han, F.; Zhang, X. Silencing shrimp white spot syndrome virus (WSSV) genes by siRNA. Antivir. Res. 2007, 73, 126–131. [Google Scholar] [CrossRef]
- Verbruggen, B.; Bickley, L.K.; Van Aerle, R.; Bateman, K.S.; Stentiford, G.D.; Santos, E.M.; Tyler, C.R. Molecular Mechanisms of White Spot Syndrome Virus Infection and Perspectives on Treatments. Viruses 2016, 8, 23. [Google Scholar] [CrossRef] [Green Version]
- Kulkarni, A.D.; Kiron, V.; Rombout, J.H.W.M.; Brinchmann, M.F.; Fernandes, J.M.O.; Sudheer, N.S.; Singh, B.I.S. Protein profiling in the gut of Penaeus monodon gavaged with oral WSSV-vaccines and live white spot syndrome virus. Proteomics 2014, 14, 1660–1673. [Google Scholar] [CrossRef]
- Ye, T.; Wu, X.; Wu, W.; Dai, C.; Yuan, J. Ferritin protect shrimp Litopenaeus vannamei from WSSV infection by inhibiting virus replication. Fish Shellfish Immunol. 2015, 42, 138–143. [Google Scholar] [CrossRef] [PubMed]
- Santos, C.A.; Andrade, S.C.S.; Teixeira, A.K.; Farias, F.; Kurkjian, K.; Guerrelhas, A.C.; Rocha, J.L.; Galetti, P.M., Jr.; Freitas, P.D. Litopenaeus vannamei Transcriptome Profile of Populations Evaluated for Growth Performance and Exposed to White Spot Syndrome Virus (WSSV). Front. Genet. 2018, 9, 120. [Google Scholar] [CrossRef] [Green Version]
- Rao, R.; Bing Zhu, Y.; Alinejad, T.; Tiruvayipati, S.; Lin Thong, K.; Wang, J.; Bhassu, S. RNA-seq analysis of Macrobrachium rosenbergii hepatopancreas in response to Vibrio parahaemolyticus infection. Gut Pathog. 2015, 7, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, K.-K.; Li, D.-F.; Zhang, M.-C.; Yang, H.-J.; Ruan, L.-W.; Xu, X. Cloning and characterization of three novel WSSV recognizing lectins from shrimp Marsupenaeus japonicus. Fish Shellfish Immunol. 2010, 28, 596–603. [Google Scholar] [CrossRef]
- World Organization for Animal Health (OIE). Infection with White Spot Syndrome Virus; Chapter 2.2.8.; OIE: Paris, France, 2019. [Google Scholar]
- Pfaffl, M.W.; Horgan, G.W.; Dempfle, L. Relative expression software tool (REST©) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Res. 2002, 30, 10. [Google Scholar] [CrossRef] [PubMed]
- Ririe, K.M.; Rasmussen, R.P.; Wittwer, C.T. Product Differentiation by Analysis of DNA Melting Curves during the Polymerase Chain Reaction. Anal. Biochem. 1997, 245, 154–160. [Google Scholar] [CrossRef] [PubMed]
- Serviço Nacional de Aprendizagem Rural (SENAR). Larvicultura e Cultivo de Camarão Marinho: Do Náuplio a Pós-Larva; SENAR: Ceará, Brazil, 2016. [Google Scholar]
- Chomczynski, P.; Mackey, K. Short technical reports. Modification of the TRI reagent procedure for isolation of RNA from polysaccharide-and proteoglycan-rich sources. Biotechniques 1995, 19, 942–945. [Google Scholar] [PubMed]
- Zhang, X.; Yuan, J.; Sun, Y.; Li, S.; Gao, Y.; Yu, Y.; Liu, C.; Wang, Q.; Lv, X.; Zhang, X.; et al. Penaeid shrimp genome provides insights into benthic adaptation and frequent molting. Nat. Commun. 2019, 10, 356. [Google Scholar] [CrossRef] [Green Version]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010, 11, R106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B (Methodol.) 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Restrepo, L.; Reyes, A.; Bajaña, L.; Betancourt, I.; Bayot, B. Draft Genome Sequence of a White Spot Syndrome Virus Isolate Obtained in Ecuador. Genome Announc. 2018, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patro, R.; Duggal, G.; Love, M.I.; Irizarry, R.A.; Kingsford, C. Salmon provides fast and bias-aware quantification of transcript expression. Nat. Methods 2017, 14, 417–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bryant, D.M.; Johnson, K.; DiTommaso, T.; Tickle, T.; Couger, M.B.; Payzin-Dogru, D.; Lee, T.J.; Leigh, N.D.; Kuo, T.-H.; Davis, F.G.; et al. A Tissue-Mapped Axolotl De Novo Transcriptome Enables Identification of Limb Regeneration Factors. Cell Rep. 2017, 18, 762–776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- The UniProt Consortium. UniProt: The universal protein knowledgebase. Nucleic Acids Res. 2017, 45, 158–169. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S.; Sato, Y.; Furumichi, M.; Tanabe, M. KEGG for integration and interpretation of large-scale molecular data sets. Nucleic Acids Res. 2012, 40, D109–D114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene Ontology: Tool for the unification of biology. Nat. Gen. 2000, 25, 25–29. [Google Scholar] [CrossRef] [Green Version]
- Young, M.D.; Wakefield, M.J.; Smyth, G.K.; Oshlack, A. Gene ontology analysis for RNA-seq: Accounting for selection bias. Genome Biol. 2010, 11, R14. [Google Scholar] [CrossRef] [Green Version]
- Wallenius, K.T. Biased Sampling; The Noncentral Hypergeometric Probability Distribution; Technical Report No. LIE ONR 70; University of Stanford: Stanford, CA, USA, 1963; p. 66. [Google Scholar]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Heberle, H.; Meirelles, G.V.; da Silva, F.R.; Telles, G.P.; Minghim, R. InteractiVenn: A web-based tool for the analysis of sets through Venn diagrams. BMC Bioinform. 2015, 16, 169. [Google Scholar] [CrossRef]
- Sun, L.; Dong, S.; Ge, Y.; Fonseca, J.P.; Robinson, Z.T.; Mysore, K.S.; Mehta, P. DiVenn: An Interactive and Integrated Web-Based Visualization Tool for Comparing Gene Lists. Front. Genet. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Chou, H.Y.; Huang, C.Y.; Lo, C.F.; Kou, G.H. Studies on transmission of white spot syndrome associated baculovirus WSBV/Penaeus monodon and P. japonicus via waterborne contact and oral ingestion. Aquaculture 1998, 1–4, 263–276. [Google Scholar] [CrossRef]
- Leonardo, V.A.D.; Bonnichon, V.; Roch, P.; Parrinello, N.; Bonami, J.-R. Comparative WSSV infection routes in the shrimp genera Marsupenaeus and Palaemon. J. Fish Dis. 2005, 28, 565–569. [Google Scholar] [CrossRef]
- Homvises, T.; Tassanakajon, A.; Somboonwiwat, K. Penaeus monodon SERPIN, PmSERPIN6, is implicated in the shrimp innate immunity. Fish Shellfish Immunol. 2010, 29, 890–898. [Google Scholar] [CrossRef]
- Bowornsakulwong, T.; Charoensapsri, W.; Rattanarojpong, T.; Khunrae, P. The expression and purification of WSSV134 from white spot syndrome virus and its inhibitory effect on caspase activity from Penaeus monodon. Protein Expr. Purif. 2017, 130, 123–128. [Google Scholar] [CrossRef]
- Sutthangkul, J.; Amparyup, P.; Charoensapsri, W.; Senapin, S.; Phiwsaiya, K.; Tassanakajon, A. Suppression of Shrimp Melanization during White Spot Syndrome Virus Infection. J. Biol. Chem. 2015. [Google Scholar] [CrossRef] [Green Version]
- Pradeep, B.; Rai, P.; Mohan, S.A.; Shekhar, M.S.; Karunasagar, I. Biology, Host Range, Pathogenesis and Diagnosis of White spot syndrome virus. Indian J. Virol. 2012, 23, 161–174. [Google Scholar] [CrossRef]
- Li, S.; Zhang, X.; Sun, Z.; Li, F.; Xiang, J. Transcriptome Analysis on Chinese Shrimp Fenneropenaeus chinensis during WSSV Acute Infection. PLoS ONE 2013, 8, e58627. [Google Scholar] [CrossRef] [Green Version]
- Buisson, M.; Manet, E.; Trescol-Biemont, M.C.; Gruffat, H.; Durand, B.; Sergeant, A. The Epstein-Barr virus (EBV) early protein EB2 is a posttranscriptional activator expressed under the control of EBV transcription factors EB1 and R. J. Virol. 1989, 63, 5276–5284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holley-Guthrie, E.A.; Quinlivan, E.B.; Mar, E.C.; Kenney, S. The Epstein-Barr virus (EBV) BMRF1 promoter for early antigen (EA-D) is regulated by the EBV transactivators, BRLF1 and BZLF1, in a cell-specific manner. J. Virol. 1990, 64, 3753–3759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, M.-F.; Lo, C.-F.; van Hulten, M.C.W.; Tzeng, H.-F.; Chou, C.-M.; Huang, C.-J.; Wang, C.-H.; Lin, J.-Y.; Vlak, J.M.; Kou, G.-H. Transcriptional Analysis of the Ribonucleotide Reductase Genes of Shrimp White Spot Syndrome Virus. Virology 2000, 277, 92–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamada, Y.; Kimura, H.; Morishima, T.; Daikoku, T.; Maeno, K.; Nishiyama, Y. The Pathogenicity of Ribonucleotide Reductase-Null Mutants of Herpes Simplex Virus Type 1 in Mice. J. Infect. Dis. 1991, 164, 1091–1097. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Li, M.; Ke, W.; Ji, Y.; Bian, X.; Yan, X. Identification of the immediate-early genes of white spot syndrome virus. Virology 2009, 385, 267–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.; Weng, S.; He, J. WSSV—Host interaction: Host response and immune evasion. Fish Shellfish Immunol. 2019, 84, 558–571. [Google Scholar] [CrossRef]
- Tassanakajon, A.; Rimphanitchayakit, V.; Visetnan, S.; Amparyup, P.; Somboonwiwat, K.; Charoensapsri, W.; Tang, S. Shrimp humoral responses against pathogens: Antimicrobial peptides and melanization. Dev. Comp. Immunol. 2018, 80, 81–93. [Google Scholar] [CrossRef]
- Wang, X.-W.; Wang, J.-X. Pattern recognition receptors acting in innate immune system of shrimp against pathogen infections. Fish Shellfish Immunol. 2013, 34, 981–989. [Google Scholar] [CrossRef]
- Pan, D.; He, N.; Yang, Z.; Liu, H.; Xu, X. Differential gene expression profile in hepatopancreas of WSSV-resistant shrimp (Penaeus japonicus) by suppression subtractive hybridization. Dev. Comp. Immunol. 2005, 29, 103–112. [Google Scholar] [CrossRef]
- Jiang, H.; Li, F.; Zhang, J.; Zhang, J.; Huang, B.; Yu, Y.; Xiang, J. Comparison of Protein Expression Profiles of the Hepatopancreas in Fenneropenaeus chinensis Challenged with Heat-inactivated Vibrio anguillarum and White Spot Syndrome Virus. Mar. Biotechnol. 2014, 16, 111–123. [Google Scholar] [CrossRef]
- Zhao, Z.-Y.; Yin, Z.-X.; Weng, S.-P.; Guan, H.-J.; Li, S.-D.; Xing, K.; Chan, S.-M.; He, J.-G. Profiling of differentially expressed genes in hepatopancreas of white spot syndrome virus-resistant shrimp (Litopenaeus vannamei) by suppression subtractive hybridisation. Fish Shellfish Immunol. 2007, 22, 520–534. [Google Scholar] [CrossRef] [PubMed]
- Supungul, P.; Rimphanitchayakit, V.; Aoki, T.; Hirono, I.; Tassanakajon, A. Molecular characterization and expression analysis of a c-type and two novel muramidase-deficient i-type lysozymes from Penaeus monodon. Fish Shellfish Immunol. 2010, 28, 490–498. [Google Scholar] [CrossRef]
- Mai, W.; Wang, W. Protection of blue shrimp (Litopenaeus stylirostris) against the White Spot Syndrome Virus (WSSV) when injected with shrimp lysozyme. Fish Shellfish Immunol. 2010, 28, 727–733. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.T.; Wang, J.; Mao, Y.; Liu, M.; Niu, S.; Qiao, Y.; Su, Y.-Q.; Wang, C.-Z.; Zheng, Z.-P. Identification and expression analysis of a new invertebrate lysozyme in Kuruma shrimp (Marsupenaeus japonicus). Fish Shellfish Immunol. 2016, 49, 336–343. [Google Scholar] [CrossRef] [PubMed]
- Cerenius, L.; Lee, B.L.; Söderhäll, K. The proPO-system: Pros and cons for its role in invertebrate immunity. Trends Immunol. 2008, 29, 263–271. [Google Scholar] [CrossRef]
- Li, F.; Xiang, J. Signaling pathways regulating innate immune responses in shrimp. Fish Shellfish Immunol. 2013, 34, 973–980. [Google Scholar] [CrossRef]
- Ai, H.-S.; Liao, J.-X.; Huang, X.-D.; Yin, Z.-X.; Weng, S.-P.; Zhao, Z.-Y.; Li, S.-D.; Yu, X.-Q.; He, J.-G. A novel prophenoloxidase 2 exists in shrimp hemocytes. Dev. Comp. Immunol. 2009, 33, 59–68. [Google Scholar] [CrossRef]
- Roux, M.M.; Pain, A.; Klimpel, K.R.; Dhar, A.K. The Lipopolysaccharide and β-1,3-Glucan Binding Protein Gene is Upregulated in White Spot Virus-Infected Shrimp (Penaeus stylirostris). J. Virol. 2002, 76, 7140–7149. [Google Scholar] [CrossRef] [Green Version]
- Lai, X.; Kong, J.; Wang, Q.; Wang, W.; Meng, X. Cloning and characterization of a β-1,3-glucan-binding protein from shrimp Fenneropenaeus chinensis. Mol. Biol. Rep. 2011, 38, 4527–4535. [Google Scholar] [CrossRef]
- Phupet, B.; Pitakpornpreecha, T.; Baowubon, N.; Runsaeng, P.; Utarabhand, P. Lipopolysaccharide- and β-1,3-glucan-binding protein from Litopenaeus vannamei: Purification, cloning and contribution in shrimp defense immunity via phenoloxidase activation. Dev. Comp. Immunol. 2018, 81, 167–179. [Google Scholar] [CrossRef]
- Danwattananusorn, T.; Kondo, H.; Aoki, T.; Hirono, I. Molecular cloning, characterization and expression analysis of a chymotrypsin-like serine protease from kuruma shrimp Marsupenaeus japonicus. Fish Sci. 2009, 75, 1231–1238. [Google Scholar] [CrossRef]
- Duan, Y.; Liu, P.; Li, J.; Li, J.; Wang, Y.; Chen, P. The responsive expression of a chitinase gene in the ridgetail white prawn Exopalaemon carinicauda against Vibrio anguillarum and WSSV challenge. Cell Stress Chaperones 2014, 19, 549–558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, T.; Ren, C.; Wang, Y.; Luo, P.; Jiang, X.; Huang, W.; Chen, C.; Hu, C. Molecular cloning, inducible expression and antibacterial analysis of a novel i-type lysozyme (lyz-i2) in Pacific white shrimp, Litopenaeus vannamei. Fish Shellfish Immunol. 2016, 54, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Hou, F.; Wang, X.; Qian, Z.; Liu, Q.; Liu, Y.; He, S.; Mi, X.; Bai, C.; Sun, C.; Liu, X. Identification and functional studies of Akirin, a potential positive nuclear factor of NF-κB signaling pathways in the Pacific white shrimp, Litopenaeus vannamei. Dev. Comp. Immunol. 2013, 41, 703–714. [Google Scholar] [CrossRef]
- Wang, D.; Li, S.; Li, F. Screening of genes regulated by relish in Chinese shrimp Fenneropenaeus chinensis. Dev. Comp. Immunol. 2013, 41, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.-C.; Leu, J.-H.; Kou, G.-H.; Wang, A.H.-J.; Lo, C.-F. Protein expression profiling of the shrimp cellular response to white spot syndrome virus infection. Dev. Comp. Immunol. 2007, 31, 672–686. [Google Scholar] [CrossRef]
- Wongpanya, R.; Aoki, T.; Hirono, I.; Yasuike, M.; Tassanakajon, A. Analysis of Gene Expression in Haemocytes of Shrimp Penaeus monodon Challenged with White Spot Syndrome Virus by cDNA Microarray. Sci. Asia 2007, 33, 165–174. [Google Scholar] [CrossRef]
- Leu, J.-H.; Chang, C.-C.; Wu, J.-L.; Hsu, C.-W.; Hirono, I.; Aoki, T.; Juan, H.-F.; Lo, C.-F.; Kou, G.-H.; Huang, H.-C. Comparative analysis of differentially expressed genes in normal and white spot syndrome virus infected Penaeus monodon. BMC Genom. 2007, 8, 120. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Li, F.; Wang, B.; Xiang, J. A new shrimp peritrophin-like gene from Exopalaemon carinicauda involved in white spot syndrome virus (WSSV) infection. Fish Shellfish Immunol. 2013, 35, 840–846. [Google Scholar] [CrossRef]
- Xie, S.; Li, F.; Zhang, X.; Zhang, J.; Xiang, J. Peritrophin-like protein from Litopenaeus vannamei (LvPT) involved in white spot syndrome virus (WSSV) infection in digestive tract challenged with reverse gavage. Chin. J. Oceanol. Limnol. 2017, 35, 1524–1530. [Google Scholar] [CrossRef]
- Xie, S.; Zhang, X.; Zhang, J.; Li, F.; Xiang, J. Envelope Proteins of White Spot Syndrome Virus (WSSV) Interact with Litopenaeus vannamei Peritrophin-Like Protein (LvPT). PLoS ONE 2015, 10, e0144922. [Google Scholar] [CrossRef] [PubMed]
- Peruzza, L.; Thamizhvanan, S.; Vimal, S.; Vinaya Kumar, K.; Shekhar, M.S.; Smith, V.J.; Hauton, C.; Vijayan, K.K.; Hameed, A.S.S. A comparative synthesis of transcriptomic analyses reveals major differences between WSSV-susceptible Litopenaeus vannamei and WSSV-refractory Macrobrachium rosenbergii. Dev. Comp. Immunol. 2020, 104, 103564. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhou, J.; White, K.P. RNA-seq differential expression studies: More sequence or more replication? Bioinformatics 2014, 30, 301–304. [Google Scholar] [CrossRef]
- Everaert, C.; Luypaert, M.; Maag, J.L.V.; Cheng, Q.X.; Dinger, M.E.; Hellemans, J.; Mestdagh, P. Benchmarking of RNA-sequencing analysis workflows using whole-transcriptome RT-qPCR expression data. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
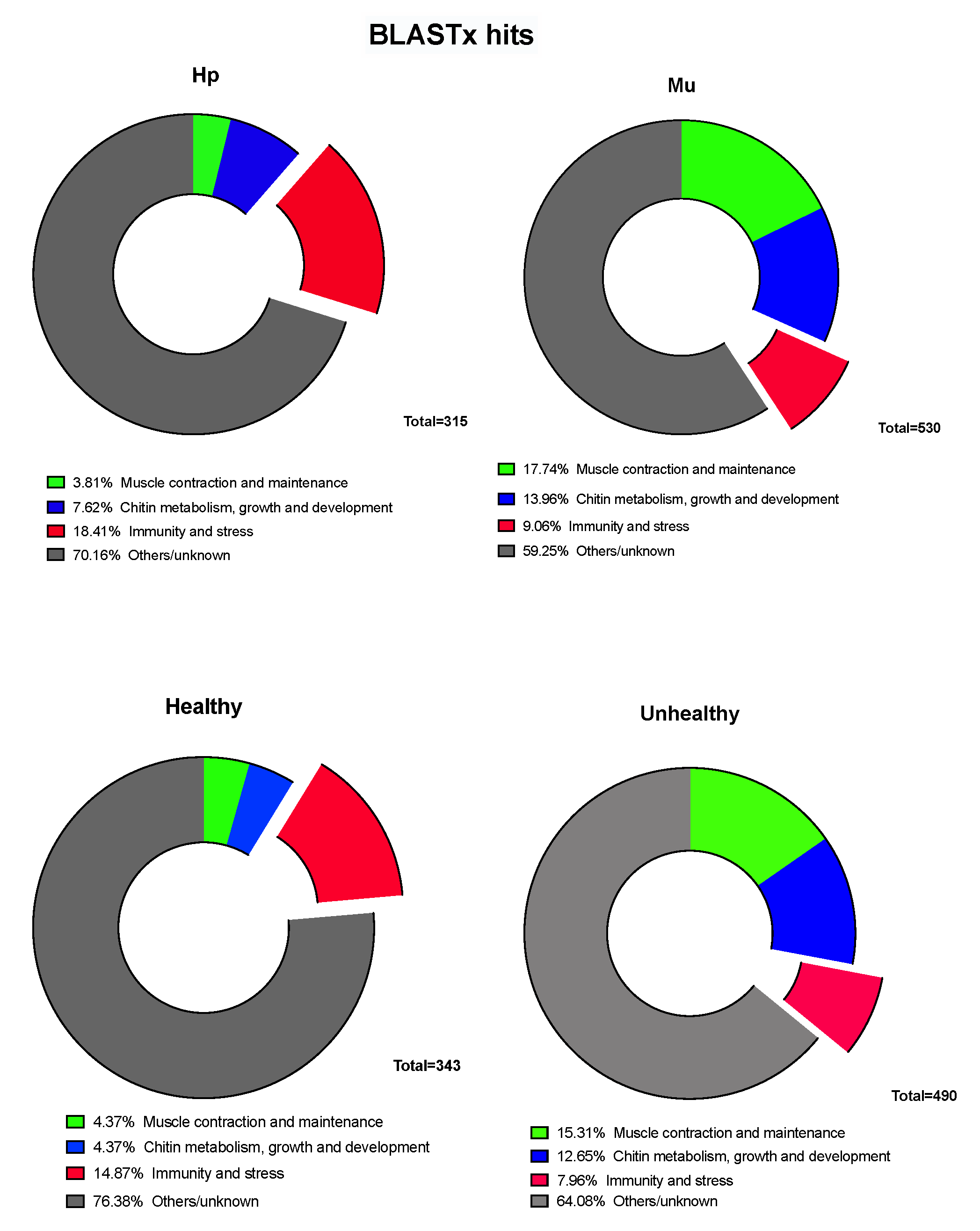{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GO ID | GO Category | Tissue | Ajusted p-Value | No. Genes | GO Term |
|---|---|---|---|---|---|
| GO:0044281 | BP | Healthy | 4.21 × 10−8 | 305 | Small molecule metabolic process |
| GO:0009117 | BP | Healthy | 1.6 × 10−4 | 64 | Nucleotide metabolic process |
| GO:0045454 | BP | Healthy | 2.0 × 10−3 | 18 | Cell redox homeostasis |
| GO:0070062 | CC | Healthy | 3.0 × 10−4 | 183 | Extracellular exosome |
| GO:0072546 | CC | Healthy | 4.0 × 10−3 | 7 | ER membrane protein complex |
| GO:0043492 | MF | Healthy | 1.5 × 10−4 | 27 | ATPase activity, coupled to movement of substances |
| GO:0001872 | MF | Healthy | 7.1 × 10−3 | 4 | (1-3)-β-D-glucan binding |
| GO:1902903 | BP | Unhealthy | 1.0 × 10−4 | 36 | Regulation of supramolecular fiber organization |
| GO:0044042 | BP | Unhealthy | 1.0 × 10−4 | 17 | Glucan metabolic process |
| GO:0035556 | BP | Unhealthy | 1.0 × 10−4 | 118 | Intracellular signal transduction |
| GO:0001558 | BP | Unhealthy | 6.0 × 10−4 | 44 | Regulation of cell growth |
| GO:0030018 | CC | Unhealthy | 2.2 × 10−3 | 26 | Z disc |
| GO:0044430 | CC | Unhealthy | 3.1 × 10−3 | 142 | Cytoskeletal part |
| GO:0042383 | CC | Unhealthy | 3.7 × 10−3 | 23 | Sarcolemma |
| GO:0051015 | MF | Unhealthy | 3.1 × 10−3 | 17 | Actin filament binding |
| GO:0008092 | MF | Unhealthy | 3.6 × 10−3 | 61 | Cytoskeletal protein binding |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santos, C.A.; Andrade, S.C.S.; Fernandes, J.M.O.; Freitas, P.D. Shedding the Light on Litopenaeus vannamei Differential Muscle and Hepatopancreas Immune Responses in White Spot Syndrome Virus (WSSV) Exposure. Genes 2020, 11, 805. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11070805
Santos CA, Andrade SCS, Fernandes JMO, Freitas PD. Shedding the Light on Litopenaeus vannamei Differential Muscle and Hepatopancreas Immune Responses in White Spot Syndrome Virus (WSSV) Exposure. Genes. 2020; 11(7):805. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11070805
Chicago/Turabian StyleSantos, Camilla A., Sónia C. S. Andrade, Jorge M. O. Fernandes, and Patrícia D. Freitas. 2020. "Shedding the Light on Litopenaeus vannamei Differential Muscle and Hepatopancreas Immune Responses in White Spot Syndrome Virus (WSSV) Exposure" Genes 11, no. 7: 805. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11070805