
A Novel QTL and a Candidate Gene Are Associated with the Progressive Motility of Franches-Montagnes Stallion Spermatozoa after Thaw


Abstract
:1. Introduction
2. Materials and Methods
2.1. Phenotypes
2.2. Raw Data Transformation
2.3. Genotypes
2.4. Genetic Analyses
3. Results
3.1. Phenotypes and Heritabilities
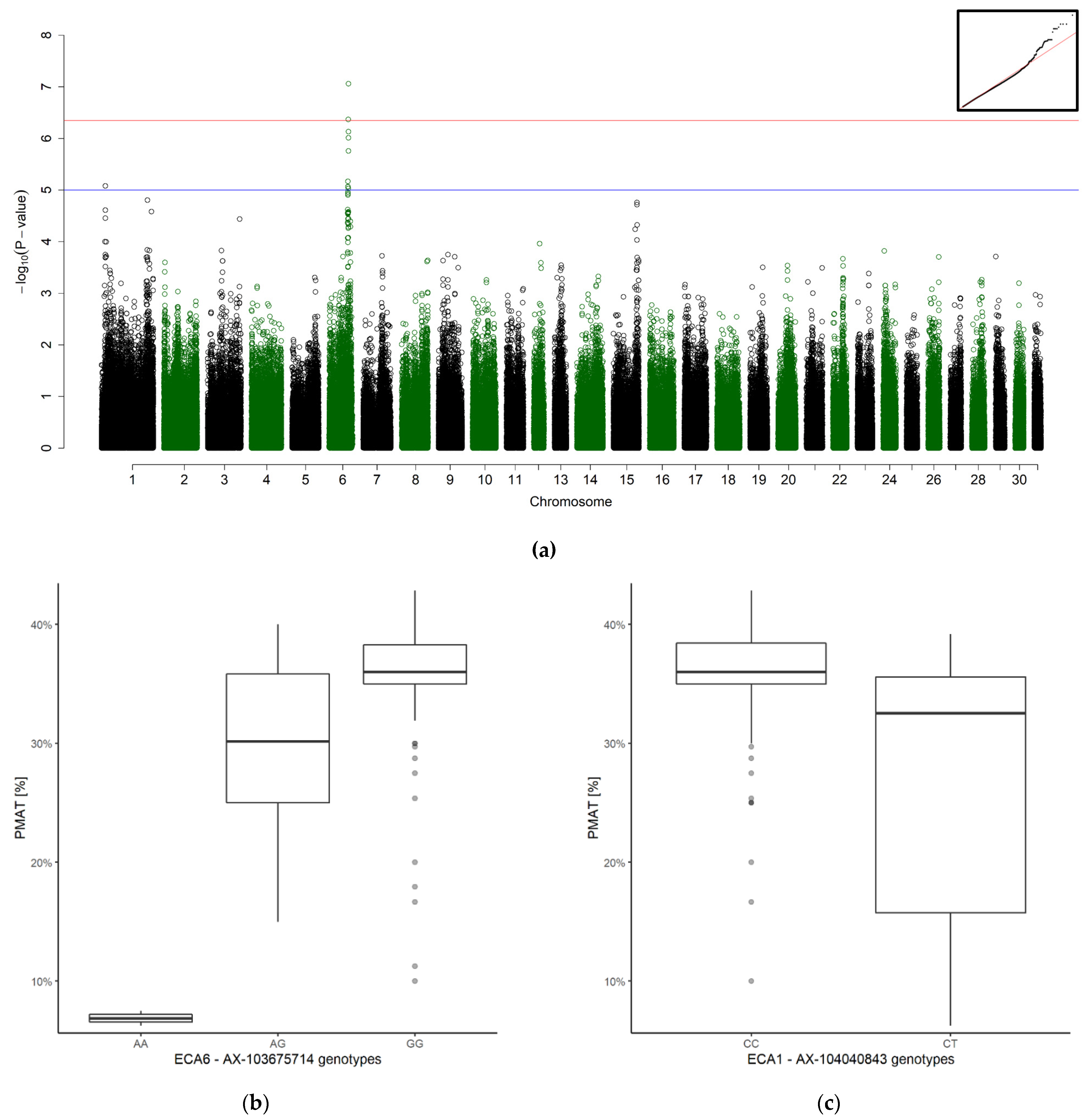
3.2. Genome-Wide Association Study
3.2.1. Covariate Structure
3.2.2. Overall Results of the GWAS
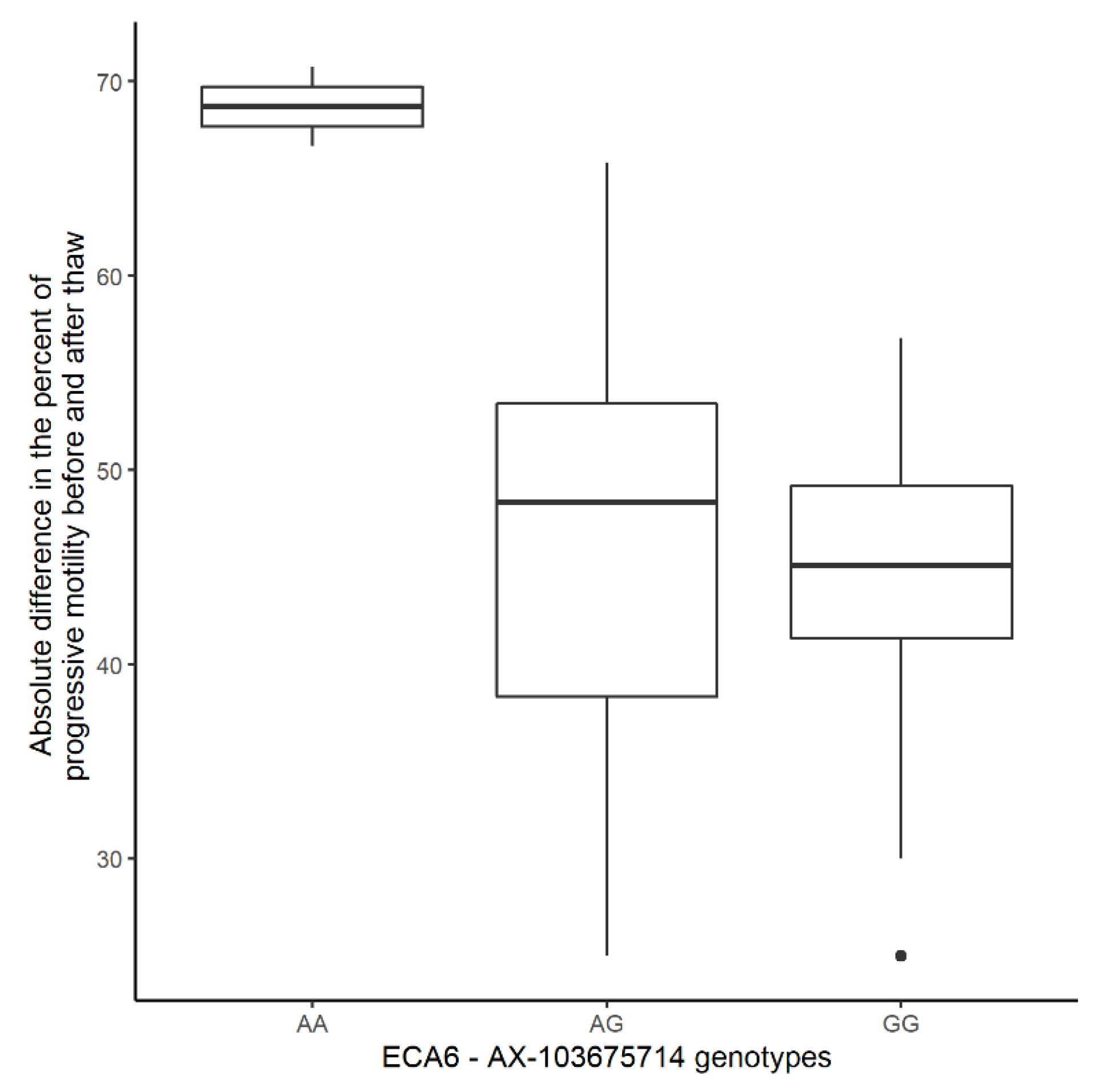
3.2.3. Significant Association for Progressive Motility after Thaw
3.2.4. Suggestive Results for Other Traits
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Animal Testing Permit
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Miller, C. Optimizing the use of frozen–thawed equine semen. Theriogenology 2008, 70, 463–468. [Google Scholar] [CrossRef] [PubMed]
- Gottschalk, M.; Sieme, H.; Martinsson, G.; Distl, O. Relationships among stallion fertility and semen traits using estimated breeding values of German Warmblood stallions. Theriogenology 2017, 89, 68–71. [Google Scholar] [CrossRef] [PubMed]
- Giesecke, K.; Hamann, H.; Stock, K.; Woehlke, A.; Sieme, H.; Distl, O. Evaluation of SPATA1-associated markers for stallion fertility. Anim. Genet. 2009, 40, 359–365. [Google Scholar] [CrossRef] [PubMed]
- Giesecke, K.; Hamann, H.; Stock, K.; Klewitz, J.; Martinsson, G.; Distl, O.; Sieme, H. Evaluation of ACE, SP17, and FSHB as candidates for stallion fertility in Hanoverian warmblood horses. Anim. Reprod. Sci. 2011, 126, 200–206. [Google Scholar] [CrossRef] [PubMed]
- Giesecke, K.; Hamann, H.; Sieme, H.; Distl, O. Evaluation of prolactin receptor (PRLR) as candidate gene for male fertility in Hanoverian warmblood horses. Reprod. Domest. Anim. 2010, 45, e124–e130. [Google Scholar]
- Gottschalk, M.; Metzger, J.; Martinsson, G.; Sieme, H.; Distl, O. Genome-wide association study for semen quality traits in German Warmblood stallions. Anim. Reprod. Sci. 2016, 171, 81–86. [Google Scholar] [CrossRef]
- Schrimpf, R.; Dierks, C.; Martinsson, G.; Sieme, H.; Distl, O. Genome-wide association study identifies phospholipase C zeta 1 (PLCz1) as a stallion fertility locus in Hanoverian warmblood horses. PLoS ONE 2014, 9, e109675. [Google Scholar] [CrossRef]
- Watson, P.F. The causes of reduced fertility with cryopreserved semen. An. Reprod. Sci. 2000, 60, 481–492. [Google Scholar] [CrossRef]
- Holt, W. Basic aspects of frozen storage of semen. Anim. Reprod. Sci. 2000, 62, 3–22. [Google Scholar] [CrossRef]
- Aurich, J.; Kuhl, J.; Tichy, A.; Aurich, C. Efficiency of semen cryopreservation in stallions. Animals 2020, 10, 1033. [Google Scholar] [CrossRef]
- Yeste, M.; Estrada, E.; Rocha, L.G.; Marín, H.; Rodríguez-Gil, J.E.; Miró, J. Cryotolerance of stallion spermatozoa is related to ROS production and mitochondrial membrane potential rather than to the integrity of sperm nucleus. Andrology 2015, 3, 395–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graham, J.K. Cryopreservation of stallion spermatozoa. Vet. Clin. N. Am. Equine Pract. 1996, 12, 131–147. [Google Scholar] [CrossRef]
- Greiser, T.; Sieme, H.; Martinsson, G.; Distl, O. Genetic parameters and estimated breeding values for traits of raw and frozen-thawed semen in German Warmblood stallions. Anim. Reprod. Sci. 2019, 210, 106194. [Google Scholar] [CrossRef] [PubMed]
- Janett, F.; Thun, R.; Bettschen, S.; Burger, D.; Hassig, M. Seasonal changes of semen quality and freezability in Franches–Montagnes stallions. Anim. Reprod. Sci. 2003, 77, 213–221. [Google Scholar] [CrossRef]
- Jeannerat, E.; Marti, E.; Berney, C.; Janett, F.; Bollwein, H.; Sieme, H.; Burger, D.; Wedekind, C. Stallion semen quality depends on major histocompatibility complex matching to teaser mare. Mol. Ecol. 2018, 27, 1025–1035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wach-Gygax, L.; Burger, D.; Malama, E.; Bollwein, H.; Fleisch, A.; Jeannerat, E.; Thomas, S.; Schuler, G.; Janett, F. Seasonal changes of DNA fragmentation and quality of raw and cold-stored stallion spermatozoa. Theriogenology 2017, 99, 98–104. [Google Scholar] [CrossRef] [Green Version]
- Gottschalk, M.; Sieme, H.; Martinsson, G.; Distl, O. Analysis of breed effects on semen traits in light horse, warmblood, and draught horse breeds. Theriogenology 2016, 85, 1375–1381. [Google Scholar] [CrossRef]
- Labitzke, D.; Sieme, H.; Martinsson, G.; Distl, O. Genetic Parameters and Breeding Values for Semen Characteristics in H anoverian Stallions. Reprod. Domest. Anim. 2014, 49, 584–587. [Google Scholar] [CrossRef]
- Greiser, T.; Sieme, H.; Martinsson, G.; Distl, O. Breed and stallion effects on frozen-thawed semen in warmblood, light and quarter horses. Theriogenology 2020, 142, 8–14. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2013. [Google Scholar]
- Frischknecht, M.; Neuditschko, M.; Jagannathan, V.; Drögemüller, C.; Tetens, J.; Thaller, G.; Leeb, T.; Rieder, S. Imputation of sequence level genotypes in the Franches-Montagnes horse breed. Genet. Sel. Evol. 2014, 46, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.; Bender, D.; Maller, J.; Sklar, P.; De Bakker, P.I.; Daly, M.J. PLINK: A tool set for whole-genome association and population-based linkage analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalbfleisch, T.S.; Rice, E.; DePriest, M.S.; Walenz, B.P.; Hestand, M.S.; Vermeesch, J.R.; O’Connell, B.L.; Fiddes, I.T.; Vershinina, A.O.; Petersen, J.L. EquCab3, an Updated Reference Genome for the Domestic Horse. bioRxiv 2018, 306928. [Google Scholar] [CrossRef]
- Aulchenko, Y.S.; Ripke, S.; Isaacs, A.; Van Duijn, C.M. GenABEL: An R library for genome-wide association analysis. Bioinformatics 2007, 23, 1294–1296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gmel, A.I.; Druml, T.; von Niederhäusern, R.; Leeb, T.; Neuditschko, M. Genome-wide association studies based on equine joint angle measurements reveal new QTL affecting the conformation of horses. Genes 2019, 10, 370. [Google Scholar] [CrossRef] [Green Version]
- Kanai, M.; Tanaka, T.; Okada, Y. Empirical estimation of genome-wide significance thresholds based on the 1000 Genomes Project data set. J. Hum. Genet. 2016, 61, 861–866. [Google Scholar] [CrossRef]
- Yang, J.; Lee, S.H.; Goddard, M.E.; Visscher, P.M. GCTA: A tool for genome-wide complex trait analysis. Am. J. Hum. Genet. 2011, 88, 76–82. [Google Scholar] [CrossRef] [Green Version]
- Pinto, F.M.; Ravina, C.G.; Fernández-Sánchez, M.; Gallardo-Castro, M.; Cejudo-Román, A.; Candenas, L. Molecular and functional characterization of voltage-gated sodium channels in human sperm. Reprod. Biol. Endocrinol. 2009, 7, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marques, D.B.; Bastiaansen, J.W.; Broekhuijse, M.L.; Lopes, M.S.; Knol, E.F.; Harlizius, B.; Guimarães, S.E.; Silva, F.F.; Lopes, P.S. Weighted single-step GWAS and gene network analysis reveal new candidate genes for semen traits in pigs. Genet. Sel. Evol. 2018, 50, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahmoun, M.; Lavery, R.; Laurent-Chaballier, S.; Bellora, N.; Philip, G.K.; Rossitto, M.; Symon, A.; Pailhoux, E.; Cammas, F.; Chung, J. In mammalian foetal testes, SOX9 regulates expression of its target genes by binding to genomic regions with conserved signatures. Nucleic Acids Res. 2017, 45, 7191–7211. [Google Scholar] [CrossRef] [Green Version]
- Langenhan, T. Adhesion G protein–coupled receptors—Candidate metabotropic mechanosensors and novel drug targets. Basic Clin. Pharmacolo. Toxicol. 2020, 126, 5–16. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.; Yang, L.; Begum, S.; Xu, L. GPR56 is essential for testis development and male fertility in mice. Dev. Dyn. 2010, 239, 3358–3367. [Google Scholar] [CrossRef] [Green Version]
- Davies, B.; Baumann, C.; Kirchhoff, C.; Ivell, R.; Nubbemeyer, R.; Habenicht, U.-F.; Theuring, F.; Gottwald, U. Targeted deletion of the epididymal receptor HE6 results in fluid dysregulation and male infertility. Mol. Cell. Biol. 2004, 24, 8642–8648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bryant, J.M.; Donahue, G.; Wang, X.; Meyer-Ficca, M.; Luense, L.J.; Weller, A.H.; Bartolomei, M.S.; Blobel, G.A.; Meyer, R.G.; Garcia, B.A. Characterization of BRD4 during mammalian postmeiotic sperm development. Mol. Cell. Biol. 2015, 35, 1433–1448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, B.C.; Gatti, M.; Goldberg, M.L. Bipolar spindle attachments affect redistributions of ZW10, a Drosophila centromere/kinetochore component required for accurate chromosome segregation. J. Cell Biol. 1996, 134, 1127–1140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swegen, A.; Aitken, R. Characterisation of the stallion sperm proteome. J. Equine Vet. Sci. 2014, 34, 35–37. [Google Scholar] [CrossRef]
- Janett, F.; Thun, R.; Ryhiner, A.; Burger, D.; Hassig, M.; Hertzberg, H. Influence of Eqvalan®(ivermectin) on quality and freezability of stallion semen. Theriogenology 2001, 55, 785–792. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Trait | Mean | SD | Min | Max | h2 | SE |
|---|---|---|---|---|---|---|
| VOL | 24.48 | 12.91 | 4.00 | 70.50 | 0.63 | 0.26 |
| CON | 284.00 | 91.99 | 122.80 | 580.00 | 0.48 | 0.27 |
| TSC | 6.12 | 2.94 | 1.81 | 17.34 | 0.02 | 0.24 |
| PM | 80.11 | 8.51 | 50.00 | 90.00 | 0.00 | 0.23 |
| PMAT | 34.51 | 7.32 | 6.25 | 45.33 | 0.03 | 0.21 |
| Trait | ECA | Position EquCab 3.0 | #SNP | SNP with the Lowest p-Value | p-Value | Number of Genotypes | Genotypes of Stallions | Reference Allele | Alternate Allele | ||
|---|---|---|---|---|---|---|---|---|---|---|---|
| HOMR | HET | HOMA | |||||||||
| PMAT | 6 | 67,290,346–69,885,065 | 11 | AX-103675714 | 1.69 × 10−7 | 108 | 95 | 11 | 2 | G | A |
| PMAT | 1 | 166,470,601 | 1 | AX-104040843 | 7.45 × 10−6 | 109 | 95 | 14 | 0 | C | T |
| CON | 12 | 1,050,320 | 1 | AX-103675085 | 2.09 × 10−6 | 105 | 92 | 12 | 1 | C | T |
| TSC | 22 | 33,024,116 | 1 | AX-103616662 | 6.71 × 10−6 | 109 | 31 | 52 | 26 | C | T |
| VOL | 1 | 46,782,612 | 1 | AX-104100234 | 8.86 × 10−6 | 109 | 91 | 18 | 0 | G | A |
| VOL | 20 | 42,868,232 | 1 | AX-104083513 | 9.82 × 10−6 | 108 | 1 | 13 | 94 | A | G |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gmel, A.I.; Burger, D.; Neuditschko, M. A Novel QTL and a Candidate Gene Are Associated with the Progressive Motility of Franches-Montagnes Stallion Spermatozoa after Thaw. Genes 2021, 12, 1501. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12101501
Gmel AI, Burger D, Neuditschko M. A Novel QTL and a Candidate Gene Are Associated with the Progressive Motility of Franches-Montagnes Stallion Spermatozoa after Thaw. Genes. 2021; 12(10):1501. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12101501
Chicago/Turabian StyleGmel, Annik Imogen, Dominik Burger, and Markus Neuditschko. 2021. "A Novel QTL and a Candidate Gene Are Associated with the Progressive Motility of Franches-Montagnes Stallion Spermatozoa after Thaw" Genes 12, no. 10: 1501. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12101501