
Associations between ALDH Genetic Variants, Alcohol Consumption, and the Risk of Nasopharyngeal Carcinoma in an East Asian Population

 , and
, and
Abstract
:1. Introduction
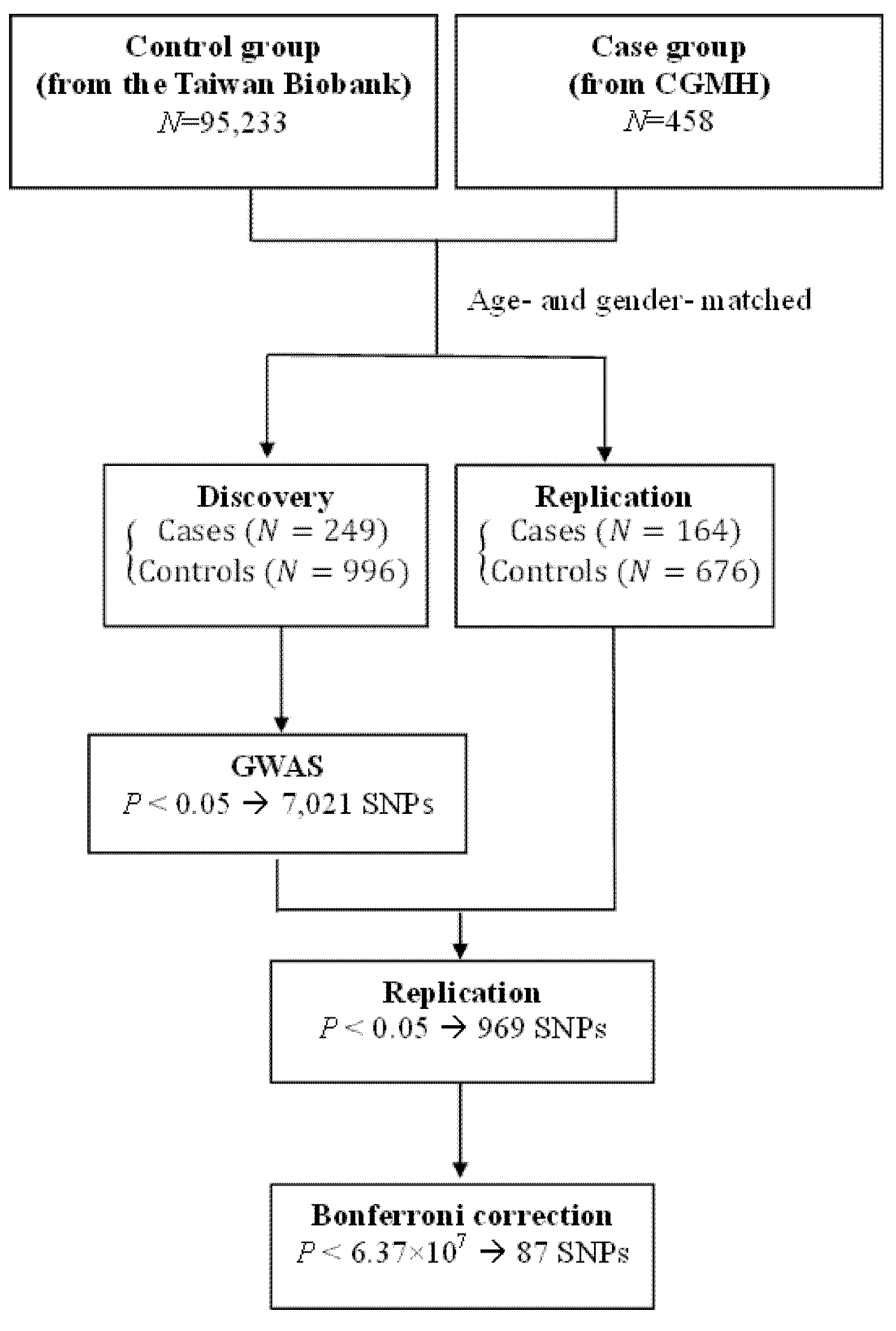
2. Materials and Methods
2.1. Study Population
2.2. Data Collection
2.3. Genotyping and Imputation
2.4. Statistical Analysis
3. Results
3.1. Characteristics of the Study Participants
3.2. ALDH Isoforms and Candidate SNPs Confer Susceptibility for NPC
3.3. Cumulative Risk Effect of 13 SNPs on NPC Susceptibility
3.4. ALDH Genes Confer Susceptibility for NPC after Stratification for Alcohol Use
3.5. The Effects of Interaction between Alcohol Consumption and SNPs on the Risk of NPC
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Chan, A.T.C.; Le, Q.-T.; Blanchard, P.; Sun, Y.; Ma, J. Nasopharyngeal carcinoma. Lancet 2019, 394, 64–80. [Google Scholar] [CrossRef]
- Yu, G.; Hsu, W.-L.; Coghill, A.E.; Yu, K.J.; Wang, C.-P.; Lou, P.-J.; Liu, Z.; Jones, K.; Vogt, A.; Wang, M.; et al. Whole-Exome Sequencing of Nasopharyngeal Carcinoma Families Reveals Novel Variants Potentially Involved in Nasopharyngeal Carcinoma. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef]
- Tse, K.-P.; Su, W.-H.; Chang, K.-P.; Tsang, N.-M.; Yu, C.-J.; Tang, P.; See, L.-C.; Hsueh, C.; Yang, M.-L.; Hao, S.-P.; et al. Genome-wide Association Study Reveals Multiple Nasopharyngeal Carcinoma-Associated Loci within the HLA Region at Chromosome 6p21.3. Am. J. Hum. Genet. 2009, 85, 194–203. [Google Scholar] [CrossRef] [Green Version]
- Bei, J.-X.; Li, Y.; Jia, W.-H.; Feng, B.-J.; Zhou, G.; Chen, L.-Z.; Feng, Q.-S.; Low, H.-Q.; Zhang, H.; He, F.; et al. A genome-wide association study of nasopharyngeal carcinoma identifies three new susceptibility loci. Nat. Genet. 2010, 42, 599–603. [Google Scholar] [CrossRef]
- Bei, J.-X.; Su, W.-H.; Yu, K.; Chin, Y.-M.; Lou, P.-J.; Hsu, W.-L.; McKay, J.D.; Chen, C.-J.; Chang, Y.-S.; Ching-Ching International Nasopharyngeal Carcinoma (NPC) Genetics Working Group. A GWAS Meta-analysis and Replication Study Identifies a Novel Locus within CLPTM1L/TERT Associated with Nasopharyngeal Carcinoma in Individuals of Chinese Ancestry. Cancer Epidemiol. Biomark. Prev. 2016, 25, 188–192. [Google Scholar] [CrossRef] [Green Version]
- Dai, W.; Zheng, H.; Cheung, A.K.L.; Tang, C.; Ko, J.; Wong, B.W.Y.; Leong, M.M.L.; Sham, P.C.; Cheung, F.; Kwong, D.L.-W.; et al. Whole-exome sequencing identifies MST1R as a genetic susceptibility gene in nasopharyngeal carcinoma. Proc. Natl. Acad. Sci. USA 2016, 113, 3317–3322. [Google Scholar] [CrossRef] [Green Version]
- Okekpa, S.; Mydin, R.B.S.M.N.; Mangantig, E.; Azmi, N.S.A.; Zahari, S.N.S.; Kaur, G.; Musa, Y. Nasopharyngeal Carcinoma (NPC) Risk Factors: A Systematic Review and Meta-Analysis of the Association with Lifestyle, Diets, Socioeconomic and Sociodemographic in Asian Region. Asian Pac. J. Cancer Prev. 2019, 20, 3505–3514. [Google Scholar] [CrossRef]
- Polesel, J.; Franceschi, S.; Talamini, R.; Negri, E.; Barzan, L.; Montella, M.; Libra, M.; Vaccher, E.; Franchin, G.; La Vecchia, C.; et al. Tobacco smoking, alcohol drinking, and the risk of different histological types of nasopharyngeal cancer in a low-risk population. Oral Oncol. 2011, 47, 541–545. [Google Scholar] [CrossRef]
- Ji, X.; Zhang, W.; Xie, C.; Wang, B.; Zhang, G.; Zhou, F. Nasopharyngeal carcinoma risk by histologic type in central China: Impact of smoking, alcohol and family history. Int. J. Cancer 2010, 129, 724–732. [Google Scholar] [CrossRef]
- Du, T.; Chen, K.; Zheng, S.; Bao, M.; Huang, Y.; Wu, K. Association Between Alcohol Consumption and Risk of Nasopharyngeal Carcinoma: A Comprehensive Meta-Analysis of Epidemiological Studies. Alcohol. Clin. Exp. Res. 2019, 43, 2262–2273. [Google Scholar] [CrossRef]
- Chen, L.; Gallicchio, L.; Boyd-Lindsley, K.; Tao, X.; Robinson, K.A.; Lam, T.K.; Herman, J.G.; Caulfield, L.E.; Guallar, E.; Alberg, A.J. Alcohol Consumption and the Risk of Nasopharyngeal Carcinoma: A Systematic Review. Nutr. Cancer 2008, 61, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Gross, E.R.; Zambelli, V.O.; Small, B.A.; Ferreira, J.C.B.; Chen, C.-H.; Mochly-Rosen, D. A Personalized Medicine Approach for Asian Americans with the Aldehyde Dehydrogenase 2*2 Variant. Annu. Rev. Pharmacol. Toxicol. 2015, 55, 107–127. [Google Scholar] [CrossRef] [Green Version]
- Crous-Bou, M.; Rennert, G.; Cuadras, D.; Salazar, R.; Cordero, D.; Rennert, H.S.; Lejbkowicz, F.; Kopelovich, L.; Lipkin, S.M.; Gruber, S.B.; et al. Polymorphisms in Alcohol Metabolism Genes ADH1B and ALDH2, Alcohol Consumption and Colorectal Cancer. PLoS ONE 2013, 8, e80158. [Google Scholar] [CrossRef] [Green Version]
- Seitz, H.K.; Stickel, F. Acetaldehyde as an underestimated risk factor for cancer development: Role of genetics in ethanol metabolism. Genes Nutr. 2010, 5, 121–128. [Google Scholar] [CrossRef] [Green Version]
- Perez-Miller, S.J.; Hurley, T.D. Coenzyme Isomerization is Integral to Catalysis in Aldehyde Dehydrogenase†,‡. Biochemistry 2003, 42, 7100–7109. [Google Scholar] [CrossRef]
- Tsai, S.-T.; Wong, T.-Y.; Ou, C.-Y.; Fang, S.-Y.; Chen, K.-C.; Hsiao, J.-R.; Huang, C.-C.; Lee, W.-T.; Lo, H.-I.; Huang, J.-S.; et al. The interplay between alcohol consumption, oral hygiene, ALDH2 and ADH1B in the risk of head and neck cancer. Int. J. Cancer 2014, 135, 2424–2436. [Google Scholar] [CrossRef]
- Brooks, P.J.; Enoch, M.-A.; Goldman, D.; Li, T.-K.; Yokoyama, A. The Alcohol Flushing Response: An Unrecognized Risk Factor for Esophageal Cancer from Alcohol Consumption. PLoS Med. 2009, 6, e1000050. [Google Scholar] [CrossRef]
- Chen, M.-H.; Weng, J.-J.; Cheng, C.-T.; Wu, R.-C.; Huang, S.-C.; Wu, C.-E.; Chung, Y.-H.; Liu, C.-Y.; Chang, M.-H.; Chiang, K.-C.; et al. ALDH1A3, the Major Aldehyde Dehydrogenase Isoform in Human Cholangiocarcinoma Cells, Affects Prognosis and Gemcitabine Resistance in Cholangiocarcinoma Patients. Clin. Cancer Res. 2016, 22, 4225–4235. [Google Scholar] [CrossRef] [Green Version]
- Luo, W.-R.; Yao, K.-T. Cancer stem cell characteristics, ALDH1 expression in the invasive front of nasopharyngeal carcinoma. Virchows Arch. 2014, 464, 35–43. [Google Scholar] [CrossRef]
- Chang, P.M.-H.; Chen, C.-H.; Yeh, C.-C.; Lu, H.-J.; Liu, T.-T.; Chen, M.-H.; Liu, C.-Y.; Wu, A.T.H.; Yang, M.-H.; Tai, S.-K.; et al. Transcriptome analysis and prognosis of ALDH isoforms in human cancer. Sci. Rep. 2018, 8, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Vasiliou, V.; Nebert, D.W. Analysis and update of the human aldehyde dehydrogenase (ALDH) gene family. Hum. Genom. 2005, 2, 138–143. [Google Scholar] [CrossRef] [Green Version]
- Su, W.-H.; Shugart, Y.Y.; Chang, K.-P.; Tsang, N.-M.; Tse, K.-P.; Chang, Y.-S. How Genome-Wide SNP-SNP Interactions Relate to Nasopharyngeal Carcinoma Susceptibility. PLoS ONE 2013, 8, e83034. [Google Scholar] [CrossRef]
- Wei, C.-Y.; Yang, J.-H.; Yeh, E.-C.; Tsai, M.-F.; Kao, H.-J.; Lo, C.-Z.; Chang, L.-P.; Lin, W.-J.; Hsieh, F.-J.; Belsare, S.; et al. Genetic profiles of 103,106 individuals in the Taiwan Biobank provide insights into the health and history of Han Chinese. NPJ Genom. Med. 2021, 6, 1–10. [Google Scholar] [CrossRef]
- Howie, B.N.; Donnelly, P.; Marchini, J. A Flexible and Accurate Genotype Imputation Method for the Next Generation of Genome-Wide Association Studies. PLoS Genet. 2009, 5, e1000529. [Google Scholar] [CrossRef] [Green Version]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.R.; Bender, D.; Maller, J.; Sklar, P.; de Bakker, P.I.W.D.; Daly, M.J.; et al. PLINK: A Tool Set for Whole-Genome Association and Population-Based Linkage Analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sham, P.C.; Purcell, S.M. Statistical power and significance testing in large-scale genetic studies. Nat. Rev. Genet. 2014, 15, 335–346. [Google Scholar] [CrossRef] [PubMed]
- Moreb, J.; Schweder, M.; Suresh, A.; Zucali, J.R. Overexpression of the human aldehyde dehydrogenase class I results in in-creased resistance to 4-hydroperoxycyclophosphamide. Cancer Gene Ther. 1996, 3, 24–30. [Google Scholar]
- Yang, C.; Wang, X.; Liao, X.; Han, C.; Yu, T.; Qin, W.; Zhu, G.; Su, H.; Yu, L.; Liu, X.; et al. Aldehyde dehydrogenase 1 (ALDH1) isoform expression and potential clinical implications in hepatocellular carcinoma. PLoS ONE 2017, 12, e0182208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishikawa, S.; Konno, M.; Hamabe, A.; Hasegawa, S.; Kano, Y.; Ohta, K.; Fukusumi, T.; Sakai, D.; Kudo, T.; Haraguchi, N.; et al. Aldehyde dehydrogenasehigh gastric cancer stem cells are resistant to chemotherapy. Int. J. Oncol. 2013, 42, 1437–1442. [Google Scholar] [CrossRef] [Green Version]
- Tomita, H.; Tanaka, K.; Tanaka, T.; Hara, A. Aldehyde dehydrogenase 1A1 in stem cells and cancer. Oncotarget 2016, 7, 11018–11032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuo, K.; Hiraki, A.; Hirose, K.; Ito, H.; Suzuki, T.; Wakai, K.; Tajima, K. Impact of theAlcohol-Dehydrogenase(ADH)1CandADH1Bpolymorphisms on drinking behavior in nonalcoholic Japanese. Hum. Mutat. 2007, 28, 506–510. [Google Scholar] [CrossRef]
- Wang, W.; Wang, C.; Xu, H.; Gao, Y. Aldehyde Dehydrogenase, Liver Disease and Cancer. Int. J. Biol. Sci. 2020, 16, 921–934. [Google Scholar] [CrossRef] [PubMed]
- Tan, B.Y.; Ning, N. Association of ADH1B Arg47His polymorphism with the risk of cancer: A meta-analysis. Biosci. Rep. 2019, 39, BSR20181915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, S.-J.; Yokoyama, A.; Yokoyama, T.; Huang, Y.-C.; Wu, S.-Y.; Shao, Y.; Niu, J.; Wang, J.; Liu, Y.; Zhou, X.-Q.; et al. Relationship between genetic polymorphisms of ALDH2 and ADH1B and esophageal cancer risk: A meta-analysis. World J. Gastroenterol. 2010, 16, 4210–4220. [Google Scholar] [CrossRef]
- Park, B.; Kim, J.-H.; Lee, E.-G.; Jung, S.-Y.; Lee, S.Y.; Kang, H.-S.; Han, J.H. Role of aldehyde dehydrogenases, alcohol dehydrogenase 1B genotype, alcohol consumption, and their combination in breast cancer in East-Asian women. Sci. Rep. 2020, 10, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Krupenko, S.A.; Krupenko, N.I. ALDH1L1 and ALDH1L2 Folate Regulatory Enzymes in Cancer. Adv. Exp. Med. Biol. 2018, 1032, 127–143. [Google Scholar] [CrossRef]
- Krupenko, S.A.; Horita, D.A. The Role of Single-Nucleotide Polymorphisms in the Function of Candidate Tumor Suppressor ALDH1L1. Front. Genet. 2019, 10, 1013. [Google Scholar] [CrossRef]
- Lourembam, D.S.; Singh, A.R.; Sharma, T.D.; Singh, T.S.; Singh, T.R.; Singh, L.S. Evaluation of Risk Factors for Nasopharyngeal Carcinoma in a High-risk Area of India, the Northeastern Region. Asian Pac. J. Cancer Prev. 2015, 16, 4927–4935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghosh, S.K.; Singh, A.S.; Mondal, R.; Kapfo, W.; Khamo, V.; Singh, Y.I. Dysfunction of mitochondria due to environmental carcinogens in nasopharyngeal carcinoma in the ethnic group of Northeast Indian population. Tumor Biol. 2014, 35, 6715–6724. [Google Scholar] [CrossRef] [PubMed]
- Lakhanpal, M.; Singh, L.C.; Rahman, T.; Sharma, J.; Singh, M.M.; Kataki, A.C.; Verma, S.; Chauhan, P.S.; Singh, Y.M.; Wajid, S.; et al. Contribution of susceptibility locus at HLA class I region and environmental factors to occurrence of nasopharyngeal cancer in Northeast India. Tumor Biol. 2014, 36, 3061–3073. [Google Scholar] [CrossRef]
- Yong, S.K.; Ha, T.C.; Yeo, M.C.R.; Gaborieau, V.; McKay, J.D.; Wee, J. Associations of lifestyle and diet with the risk of nasopharyngeal carcinoma in Singapore: A case–control study. Chin. J. Cancer 2017, 36, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, C.W.; Chang, W.S.; Gong, C.L.; Shih, L.C.; Chen, L.Y.; Lin, E.Y.; Li, H.T.; Yen, S.T.; Wu, C.N.; Bau, D.T. Contribution of Matrix Metallopeptidase-1 Genotypes, Smoking, Alcohol Drinking and Areca Chewing to Nasopharyngeal Carcinoma Sus-ceptibility. Anticancer Res. 2016, 36, 3335–3340. [Google Scholar] [PubMed]
- Chang, E.T.; Ye, W.; Zeng, Y.-X.; Adami, H.-O. The Evolving Epidemiology of Nasopharyngeal Carcinoma. Cancer Epidemiol. Biomark. Prev. 2021, 30, 1035–1047. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Diehl, S.; Pfeiffer, R.; Chen, C.-J.; Hsu, W.-L.; Dosemeci, M.; Cheng, Y.-J.; Sun, B.; Goldstein, A.M.; Hildesheim, A. Evaluation of Risk Factors for Nasopharyngeal Carcinoma in High-Risk Nasopharyngeal Carcinoma Families in Taiwan. Cancer Epidemiol. Biomark. Prev. 2005, 14, 900–905. [Google Scholar] [CrossRef] [Green Version]
- Chang, E.T.; Liu, Z.; Hildesheim, A.; Liu, Q.; Cai, Y.; Zhang, Z.; Chen, G.; Xie, S.-H.; Cao, S.-M.; Shao, J.-Y.; et al. Active and Passive Smoking and Risk of Nasopharyngeal Carcinoma: A Population-Based Case-Control Study in Southern China. Am. J. Epidemiol. 2017, 185, 1272–1280. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Database 1-Discovery (N = 1245) | Database 2-Replication (N = 840) | Database 3-Total (N = 2085) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Controls (N = 996) | Cases (N = 249) | p Value | Controls (N = 676) | Cases (N = 164) | p Value | Controls (N = 1672) | Cases (N = 413) | p Value | |
| Age | 48.16 (9.92) | 48.01 (9.43) | 0.827 | 47.84 (11.00) | 47.92 (10.91) | 0.934 | 48.03 (10.37) | 47.98 (10.03) | 0.919 |
| Gender | 1.000 | 0.924 | 0.939 | ||||||
| Female | 228 (22.9) | 57 (22.9) | 188 (27.8) | 45 (27.4) | 416 (24.9) | 102 (24.7) | |||
| Male | 768 (77.1) | 192 (77.1) | 488 (72.2) | 119 (72.6) | 1256 (75.1) | 311 (75.3) | |||
| Alcohol users | <0.001 ** | <0.001 ** | <0.001 ** | ||||||
| No | 830 (83.4) | 156 (62.7) | 581 (85.9) | 105 (64.0) | 1411 (84.4) | 261 (63.2) | |||
| Yes | 165 (16.6) | 93 (37.3) | 95 (14.1) | 59 (36.0) | 260 (15.6) | 152 (36.8) | |||
| Betel quid chewers | <0.001 ** | <0.001 ** | <0.001 ** | ||||||
| No | 937 (94.3) | 189 (75.9) | 631 (93.6) | 129 (78.7) | 1568 (94.0) | 318 (77.0) | |||
| Yes | 57 (5.7) | 60 (24.1) | 43 (6.4) | 35 (21.3) | 100 (6.0) | 95 (23.0) | |||
| Cigarette smokers | 0.005 * | <0.001 ** | <0.001 ** | ||||||
| No | 620 (62.3) | 131 (52.6) | 464 (68.6) | 80 (48.8) | 1084 (64.9) | 211 (51.1) | |||
| Yes | 375 (37.7) | 118 (47.4) | 212 (31.4) | 84 (51.2) | 587 (35.1) | 202 (48.9) | |||
| rsID | Genes | Chr. | pb38 | Risk Allele | Discovery | Replication | Total | |||
|---|---|---|---|---|---|---|---|---|---|---|
| OR (95% CI) | OR (95% CI) | Genotype Frequency | OR (95% CI) | p Value | ||||||
| Cases | Controls | |||||||||
| rs7534676 | ALDH4A1;RP13-279N23.2 | 1 | 18,893,311 | C | 0.35 (0.2–0.62) | 0.34 (0.18–0.65) | 3/20/320 | 16/294/1336 | 0.35 (0.23–0.53) | 8.92 × 107 * |
| rs7554974 | ALDH4A1;RP13-279N23.2 | 1 | 18,897,992 | T | 0.41 (0.25–0.69) | 0.34 (0.18–0.64) | 3/23/323 | 17/299/1340 | 0.38 (0.26–0.57) | 2.13 × 106 * |
| rs7518631 | ALDH4A1;RP13-279N23.2 | 1 | 18,899,249 | A | 0.43 (0.26–0.72) | 0.34 (0.18–0.64) | 3/24/322 | 18/299/1339 | 0.40 (0.27–0.58) | 3.24 × 106 * |
| rs72936453 | ALDH4A1;RP13-279N23.2 | 1 | 18,899,852 | T | 0.43 (0.26–0.72) | 0.34 (0.18–0.63) | 3/24/323 | 18/299/1339 | 0.39 (0.27–0.58) | 3.01 × 106 * |
| rs1229984 | ADH1B | 4 | 99,318,162 | T | 0.95 (0.70–1.30) | 1.06 (0.76–1.50) | 16/86/132 | 104/656/912 | 1.00 (0.79–1.25) | 9.72 × 101 |
| rs17111068 | ALDH18A1 | 10 | 95,638,799 | T | 0.12 (0.04–0.38) | 0.12 (0.03–0.49) | 0/5/304 | 6/204/1462 | 0.12 (0.05–0.30) | 3.91 × 106 * |
| rs76655136 | ALDH3B2;RP11-119D9.1 | 11 | 67,695,970 | C | 0.14 (0.05–0.38) | 0.37 (0.17–0.83) | 0/11/333 | 11/191/1470 | 0.23 (0.12–0.42) | 3.02 × 106 * |
| rs10778364 | ALDH1L2;C12orf45 | 12 | 105,047,085 | T | 0.68 (0.52–0.89) | 0.66 (0.47–0.94) | 8/115/246 | 96/598/960 | 0.67 (0.54–0.83) | 1.93 × 104 |
| rs671 | ALDH2 | 12 | 11,180,3962 | A | 1.32 (1.05–1.67) | 1.10 (0.82–1.47) | 39/163/191 | 146/684/842 | 1.23 (1.03–1.48) | 2.25 × 102 |
| rs34200934 | RP11-344A16.2;ALDH1A2 | 15 | 57,767,226 | T | 0.70 (0.50–0.97) | 0.66 (0.44–1.01) | 8/66/307 | 30/414/1185 | 0.69 (0.53–0.89) | 4.76 × 103 |
| rs11636446 | RP11-344A16.2;ALDH1A2 | 15 | 57,780,522 | C | 0.67 (0.46–0.98) | 0.65 (0.42–1.01) | 3/60/315 | 21/364/1282 | 0.67 (0.50–0.89) | 5.41 × 103 |
| rs79071218 | RP11-344A16.2;ALDH1A2 | 15 | 57,885,374 | C | 0.19 (0.08–0.48) | 0.41 (0.18–0.93) | 0/12/356 | 4/189/1479 | 0.28 (0.15–0.51) | 3.50 × 105 |
| rs1975431 | RP11-344A16.2;ALDH1A2 | 15 | 57,887,256 | C | 0.19 (0.08–0.48) | 0.36 (0.15–0.87) | 0/11/356 | 4/188/1480 | 0.26 (0.14–0.48) | 2.33 × 105 |
| rs28829404 | ALDH1A2;LIPC | 15 | 58,379,641 | T | 0.40 (0.22–0.73) | 0.26 (0.09–0.71) | 1/14/346 | 6/196/1470 | 0.34 (0.20–0.58) | 5.32 × 105 |
| OR (95% CI) | p Value | |
|---|---|---|
| Alcohol consumption | 2.61 (1.60–4.26) | <0.001 ** |
| Betel quid chewing | 2.63 (1.40–4.94) | 0.003 * |
| Cigarette smoking | 0.79 (0.49–1.27) | 0.334 |
| Cumulative risk allele | ||
| <median | Ref. | Ref. |
| ≥median | 3.98 (2.45–6.46) | <0.001 ** |
| Alcohol Use = No | Alcohol Use = Yes | |||||||
|---|---|---|---|---|---|---|---|---|
| Controls (N = 1411) | Cases (N = 261) | OR (95% CI) | p Value | Controls (N = 260) | Cases (N = 152) | OR (95% CI) | p Value | |
| rs1229984 | ||||||||
| CC/CT | 647 (45.9) | 68 (43.3) | Ref. | Ref. | 112 (43.1) | 34 (44.2) | Ref. | Ref. |
| TT | 764 (54.1) | 89 (56.7) | 1.11 (0.80–1.55) | 0.544 | 148 (56.9) | 43 (55.8) | 0.96 (0.57–1.60) | 0.867 |
| rs671 | ||||||||
| GG/AG | 1270 (90.0) | 211 (84.5) | Ref. | Ref. | 255 (98.1) | 142 (100.0) | Ref. | Ref. |
| AA | 141 (10.0) | 39 (15.5) | 1.66 (1.13–2.43) | 0.010 * | 5 (1.9) | 0 (0.0) | 0.00 (0.00–) | 0.999 |
| Recessive (AA/AG) | 755 (53.5) | 149 (59.4) | 1.27 (0.97–1.67) | 0.087 | 75 (28.8) | 53 (37.3) | 1.47 (0.95–2.27) | 0.082 |
| rs7534676 | ||||||||
| TT/TC | 253 (18.2) | 16 (7.3) | Ref. | Ref. | 56 (21.8) | 7 (5.6) | Ref. | Ref. |
| CC | 1135 (81.8) | 203 (92.7) | 2.83 (1.67–4.79) | <0.001 * | 201 (78.2) | 117 (94.4) | 4.66 (2.06–10.55) | <0.001 * |
| rs7554974 | ||||||||
| CC/CT | 259 (18.5) | 19 (8.5) | Ref. | Ref. | 56 (21.8) | 7 (5.6) | Ref. | Ref. |
| TT | 1139 (81.5) | 205 (91.5) | 2.45 (1.51–4.00) | <0.001 * | 201 (78.2) | 118 (94.4) | 4.70 (2.07–10.64) | <0.001 * |
| rs7518631 | ||||||||
| GG/GA | 260 (18.6) | 19 (8.5) | Ref. | Ref. | 56 (21.8) | 8 (6.3) | Ref. | Ref. |
| AA | 1138 (81.4) | 204 (91.5) | 2.45 (1.51–4.00) | <0.001 * | 201 (78.2) | 118 (93.7) | 4.11 (1.89–8.92) | <0.001 * |
| rs72936453 | ||||||||
| AA/AT | 260 (18.6) | 19 (8.5) | Ref. | Ref. | 56 (21.8) | 8 (6.3) | Ref. | Ref. |
| TT | 1138 (81.4) | 204 (91.5) | 2.45 (1.51–4.00) | <0.001 * | 201 (78.2) | 119 (93.7) | 4.14 (1.91–8.99) | <0.001 * |
| rs17111068 | ||||||||
| GG/GT | 178 (12.6) | 4 (2.0) | Ref. | Ref. | 32 (12.3) | 1 (0.9) | Ref. | Ref. |
| TT | 1233 (87.4) | 195 (98.0) | 7.04 (2.58–19.18) | <0.001 * | 228 (87.7) | 109 (99.1) | 15.30 (2.06–113.42) | 0.008 * |
| rs76655136 | ||||||||
| TT/TC | 167 (11.8) | 6 (2.7) | Ref. | Ref. | 35 (13.5) | 5 (4.1) | Ref. | Ref. |
| CC | 1244 (88.2) | 215 (97.3) | 4.81 (2.10–11.00) | <0.001 * | 225 (86.5) | 118 (95.9) | 3.67 (1.40–9.62) | 0.008 * |
| rs10778364 | ||||||||
| CC/CT | 586 (42.0) | 76 (32.8) | Ref. | Ref. | 107 (41.3) | 47 (34.3) | Ref. | Ref. |
| TT | 808 (58.0) | 156 (67.2) | 1.49 (1.10–2.00) | 0.008 * | 152 (58.7) | 90 (65.7) | 1.35 (0.88–2.07) | 0.174 |
| rs34200934 | ||||||||
| GG/GT | 365 (26.6) | 42 (17.1) | Ref. | Ref. | 79 (30.9) | 32 (23.7) | Ref. | Ref. |
| TT | 1007 (73.4) | 204 (82.9) | 1.76 (1.24–2.51) | 0.002 * | 177 (69.1) | 103 (76.3) | 1.44 (0.89–2.32) | 0.137 |
| rs11636446 | ||||||||
| AA/AC | 321 (22.8) | 36 (15.2) | Ref. | Ref. | 64 (24.7) | 27 (19.1) | Ref. | Ref. |
| CC | 1086 (77.2) | 201 (84.8) | 1.65 (1.13–2.40) | 0.009 * | 195 (75.3) | 114 (80.9) | 1.39 (0.84–2.30) | 0.206 |
| rs79071218 | ||||||||
| TT/TC | 170 (12.0) | 7 (3.1) | Ref. | Ref. | 23 (8.8) | 5 (3.5) | Ref. | Ref. |
| CC | 1241 (88.0) | 219 (96.9) | 4.29 (1.99–9.25) | <0.001 * | 237 (91.2) | 137 (96.5) | 2.66 (0.99–7.15) | 0.053 |
| rs1975431 | ||||||||
| AA/AC | 169 (12.0) | 7 (3.1) | Ref. | Ref. | 23 (8.8) | 4 (2.8) | Ref. | Ref. |
| CC | 1242 (88.0) | 219 (96.9) | 4.26 (1.97–9.19) | <0.001 * | 237 (91.2) | 137 (97.2) | 3.32 (113–9.81) | 0.030 * |
| rs28829404 | ||||||||
| CC/CT | 163 (11.6) | 13 (5.7) | Ref. | Ref. | 39 (15.0) | 2 (1.5) | Ref. | Ref. |
| TT | 1248 (88.4) | 216 (94.3) | 2.17 (1.21–3.89) | 0.009 * | 221 (85.0) | 130 (98.5) | 11.47 (2.73–48.29) | 0.001 * |
| Cumulative risk alleles | ||||||||
| <Median | 615 (46.3) | 14 (19.4) | Ref. | Ref. | 142 (56.6) | 8 (21.1) | Ref. | Ref. |
| ≥Median | 713 (53.7) | 58 (80.6) | 3.57 (1.97–6.47) | <0.001 * | 109 (43.4) | 30 (78.9) | 4.89 (2.15–11.08) | <0.001 * |
| Controls (N = 1672) | Cases (N = 413) | p Value a | OR (95% CI) | p Value b | |
|---|---|---|---|---|---|
| rs1229984*Alcohol | <0.001 | ||||
| CC/CT*Non-Alcohol | 647 (38.7) | 68 (29.1) | Ref. | Ref. | |
| TT*Non-Alcohol or CC/CT*Alcohol | 876 (52.4) | 123 (52.6) | 1.34 (0.98–1.83) | 0.070 | |
| TT*Alcohol | 148 (8.9) | 43 (18.4) | 2.76 (1.81–4.21) | <0.001 * | |
| rs671*Alcohol | - | ||||
| GG/AG*Non-Alcohol | 1270 (76.0) | 212 (53.9) | Ref. | Ref. | |
| AA*Non-Alcohol or GG/AG*Alcohol | 396 (23.7) | 181 (46.1) | 2.74 (2.18–3.44) | <0.001 * | |
| AA*Alcohol | 5 (0.3) | 0 (0.0) | - (–) | 0.999 | |
| (Recessive model) | <0.001 | ||||
| GG*Non-Alcohol | 656 (39.3) | 102 (26.0) | Ref. | Ref. | |
| AA/AG*Non-Alcohol or GG*Alcohol | 940 (56.3) | 238 (60.6) | 1.63 (1.27–2.10) | <0.001 * | |
| AA/AG*Alcohol | 75 (4.5) | 53 (13.5) | 4.55 (3.02–6.84) | <0.001 * | |
| rs7534676*Alcohol | <0.001 | ||||
| TT/TC*Non-Alcohol | 253 (15.4) | 16 (4.7) | Ref. | Ref. | |
| CC*Non-Alcohol or TT/TC*Alcohol | 1191 (72.4) | 210 (61.2) | 2.79 (1.65–4.72) | <0.001 * | |
| CC*Alcohol | 201 (12.2) | 117 (34.1) | 9.20 (5.29–16.02) | <0.001 * | |
| rs7554974*Alcohol | <0.001 | ||||
| CC/CT*Non-Alcohol | 259 (15.6) | 19 (5.4) | Ref. | Ref. | |
| TT*Non-Alcohol or CC/CT*Alcohol | 1195 (72.2) | 212 (60.7) | 2.42 (1.48–3.94) | <0.001 * | |
| TT*Alcohol | 201 (12.1) | 118 (33.8) | 8.00 (4.77–13.44) | <0.001 * | |
| rs7518631*Alcohol | <0.001 | ||||
| GG/GA*Non-Alcohol | 260 (15.7) | 19 (5.4) | Ref. | Ref. | |
| AA*Non-Alcohol or GG/GA*Alcohol | 1194 (72.1) | 212 (60.7) | 2.43 (1.49–3.96) | <0.001* | |
| AA*Alcohol | 201 (12.1) | 118 (33.8) | 8.03 (4.78–13.49) | <0.001 * | |
| rs72936453*Alcohol | <0.001 | ||||
| AA/AT*Non-Alcohol | 260 (15.7) | 19 (5.4) | Ref. | Ref. | |
| TT*Non-Alcohol or AA/AT*Alcohol | 1194 (72.1) | 212 (60.6) | 2.43 (1.49–3.96) | <0.001 * | |
| TT*Alcohol | 201 (12.1) | 119 (34.0) | 8.10 (4.83–13.60) | <0.001 * | |
| rs17111068*Alcohol | <0.001 | ||||
| GG/GT*Non-Alcohol | 178 (10.7) | 4 (1.3) | Ref. | Ref. | |
| TT*Non-Alcohol or GG/GT*Alcohol | 1265 (75.7) | 196 (63.4) | 6.90 (2.53–18.79) | <0.001 * | |
| TT*Alcohol | 228 (13.6) | 109 (35.3) | 21.27 (7.70–58.81) | <0.001 * | |
| rs76655136*Alcohol | <0.001 | ||||
| TT/TC*Non-Alcohol | 167 (10.0) | 6 (1.7) | Ref. | Ref. | |
| CC*Non-Alcohol or TT/TC*Alcohol | 1279 (76.5) | 220 (64.0) | 4.79 (2.09–10.95) | <0.001 * | |
| CC*Alcohol | 225 (13.5) | 118 (34.3) | 14.60 (6.28–33.96) | <0.001 * | |
| rs10778364*Alcohol | <0.001 | ||||
| CC/CT*Non-Alcohol | 586 (35.5) | 76 (20.6) | Ref. | Ref. | |
| TT Non-Alcohol or CC/CT*Alcohol | 915 (55.4) | 203 (55.0) | 1.71 (1.29–2.27) | <0.001 * | |
| TT*Alcohol | 152 (9.2) | 90 (24.4) | 4.57 (3.21–6.50) | <0.001 * | |
| rs34200934*Alcohol | <0.001 | ||||
| GG/GT*Non-Alcohol | 365 (22.4) | 42 (11.0) | Ref. | Ref. | |
| TT*Non-Alcohol or GG/GT*Alcohol | 1086 (66.7) | 236 (61.9) | 1.89 (1.33–2.68) | <0.001 * | |
| TT*Alcohol | 177 (10.9) | 103 (27.0) | 5.06 (3.39–7.55) | <0.001 * | |
| rs11636446*Alcohol | <0.001 | ||||
| AA/AC*Non-Alcohol | 321 (19.3) | 36 (9.5) | Ref. | Ref. | |
| CC*Non-Alcohol or AA/AC*Alcohol | 1150 (69.0) | 228 (60.3) | 1.77 (1.22–2.57) | 0.003 * | |
| CC*Alcohol | 195 (11.7) | 114 (30.2) | 5.22 (3.44–7.89) | <0.001 * | |
| rs79071218*Alcohol | <0.001 | ||||
| TT/TC*Non-Alcohol | 170 (10.2) | 7 (1.9) | Ref. | Ref. | |
| CC*Non-Alcohol or TT/TC*Alcohol | 1264 (75.6) | 224 (60.9) | 4.30 (1.99–9.29) | <0.001 * | |
| CC*Alcohol | 237 (14.2) | 137 (37.2) | 14.04 (6.41–30.77) | <0.001 * | |
| rs1975431*Alcohol | <0.001 | ||||
| AA/AC*Non-Alcohol | 169 (10.1) | 7 (1.9) | Ref. | Ref. | |
| CC*Non-Alcohol or AA/AC*Alcohol | 1265 (75.7) | 223 (60.8) | 4.26 (1.97–9.19) | <0.001 * | |
| CC*Alcohol | 237 (14.2) | 137 (37.3) | 13.96 (6.37–30.59) | <0.001 * | |
| rs28829404*Alcohol | <0.001 | ||||
| CC/CT*Non-Alcohol | 163 (9.8) | 13 (3.6) | Ref. | Ref. | |
| TT*Non-Alcohol or CC/CT*Alcohol | 1287 (77.0) | 218 (60.4) | 2.12 (1.19–3.80) | 0.011 * | |
| TT*Alcohol | 221 (13.2) | 130 (36.0) | 7.38 (4.03–13.51) | <0.001 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liao, W.-L.; Chan, F.-C.; Chang, K.-P.; Chang, Y.-W.; Chen, C.-H.; Su, W.-H.; Chang, H.-H. Associations between ALDH Genetic Variants, Alcohol Consumption, and the Risk of Nasopharyngeal Carcinoma in an East Asian Population. Genes 2021, 12, 1547. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12101547
Liao W-L, Chan F-C, Chang K-P, Chang Y-W, Chen C-H, Su W-H, Chang H-H. Associations between ALDH Genetic Variants, Alcohol Consumption, and the Risk of Nasopharyngeal Carcinoma in an East Asian Population. Genes. 2021; 12(10):1547. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12101547
Chicago/Turabian StyleLiao, Wen-Ling, Fu-Chun Chan, Kai-Ping Chang, Ya-Wen Chang, Che-Hong Chen, Wen-Hui Su, and Hen-Hong Chang. 2021. "Associations between ALDH Genetic Variants, Alcohol Consumption, and the Risk of Nasopharyngeal Carcinoma in an East Asian Population" Genes 12, no. 10: 1547. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12101547