
Candidate Genes of Regulation of Skeletal Muscle Energy Metabolism in Athletes

 ,
,  ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
3. Results
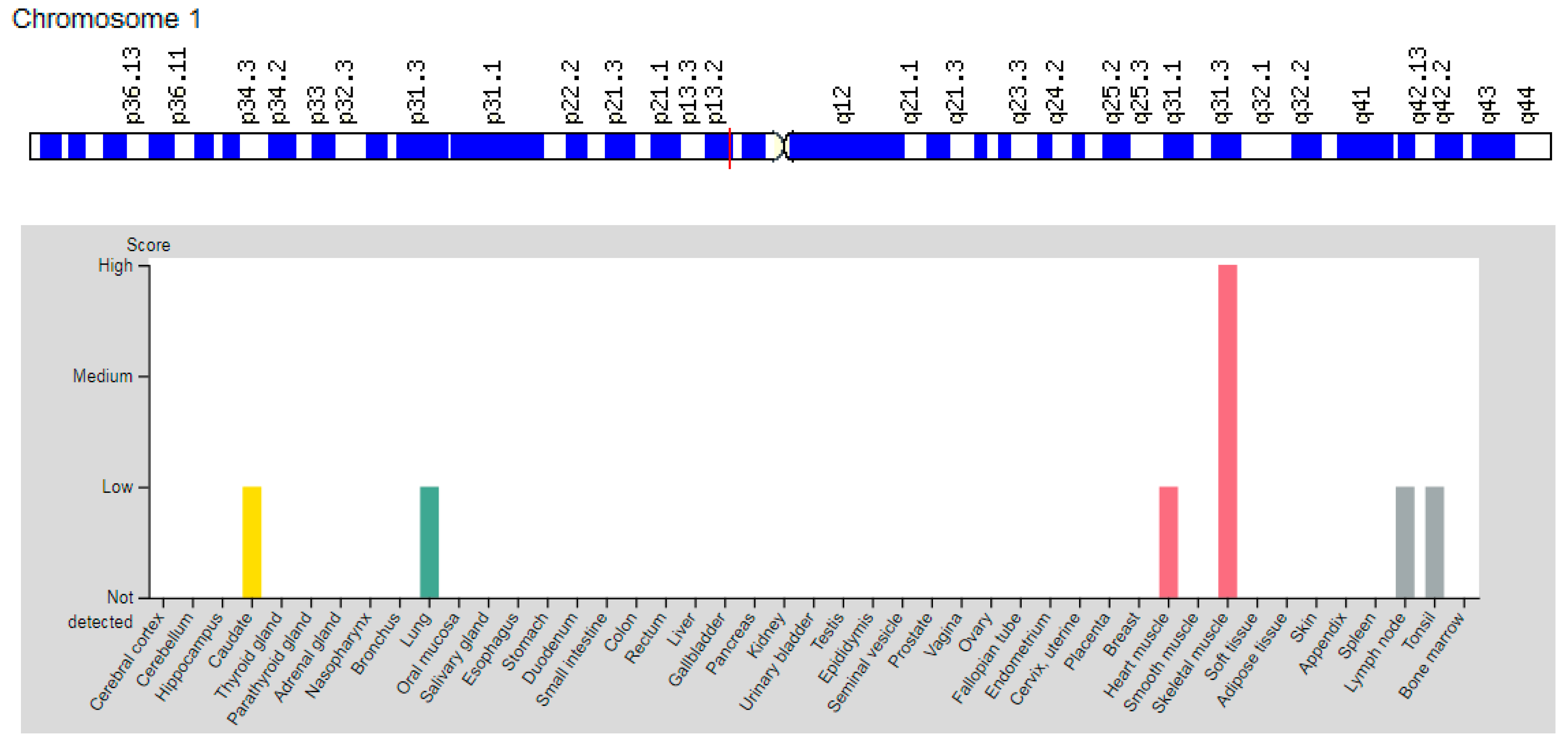
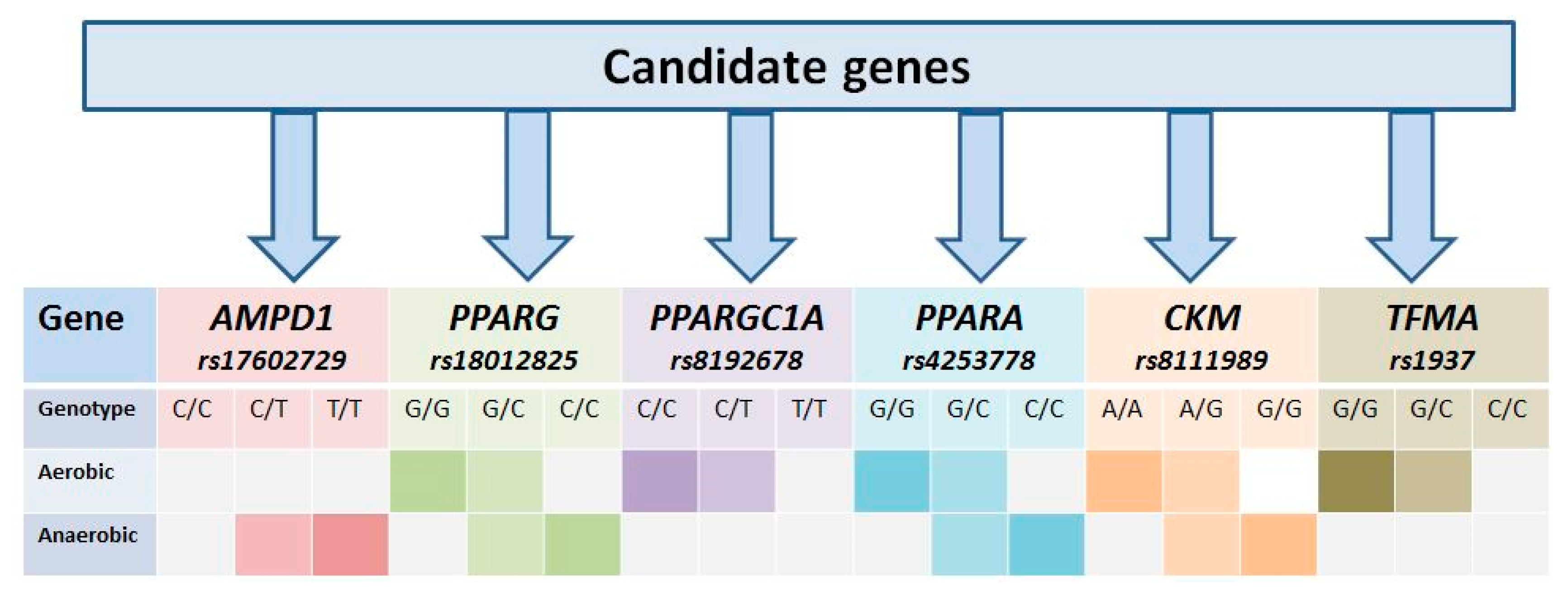
3.1. Gene AMPD1
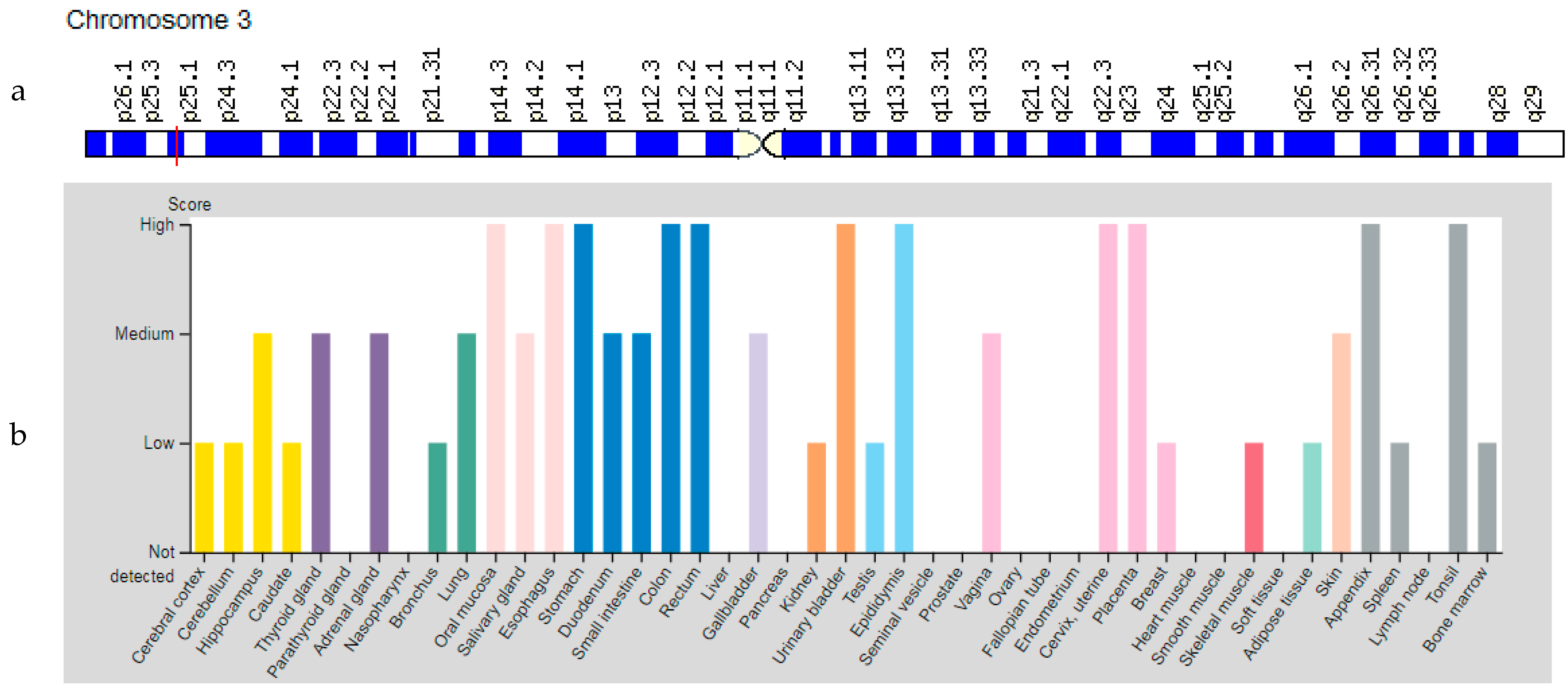
3.2. PPARG Gene
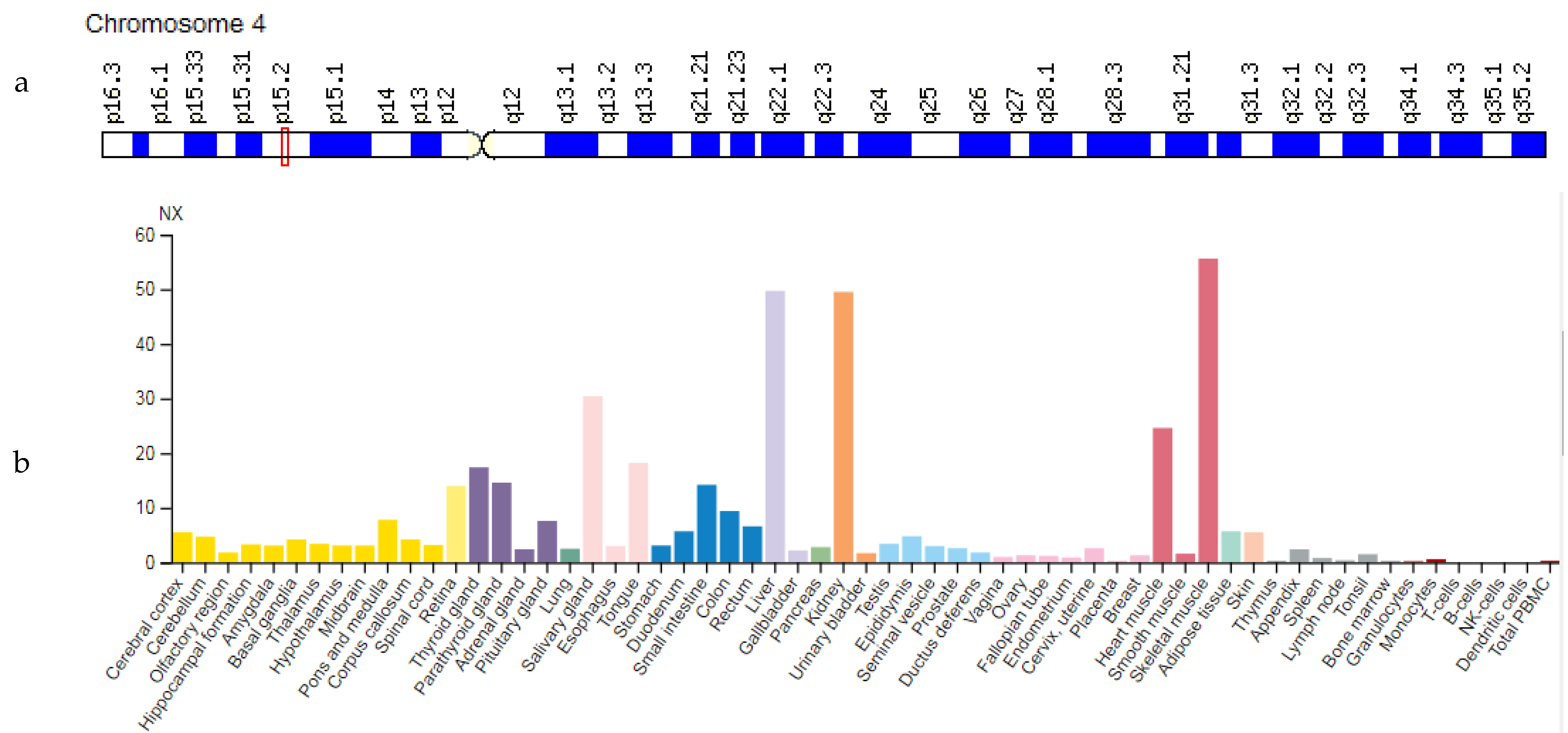
3.3. PPARGC1A Gene
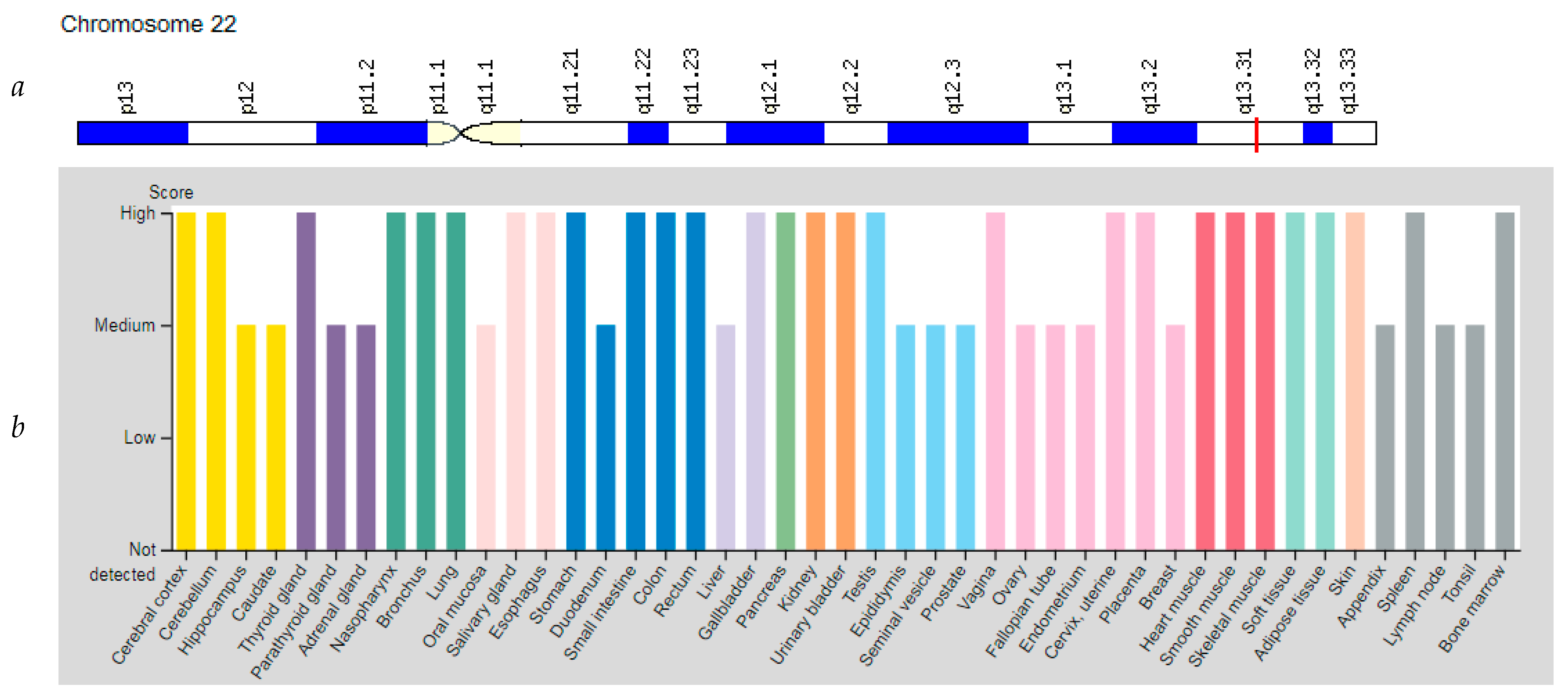
3.4. Gene PPARA
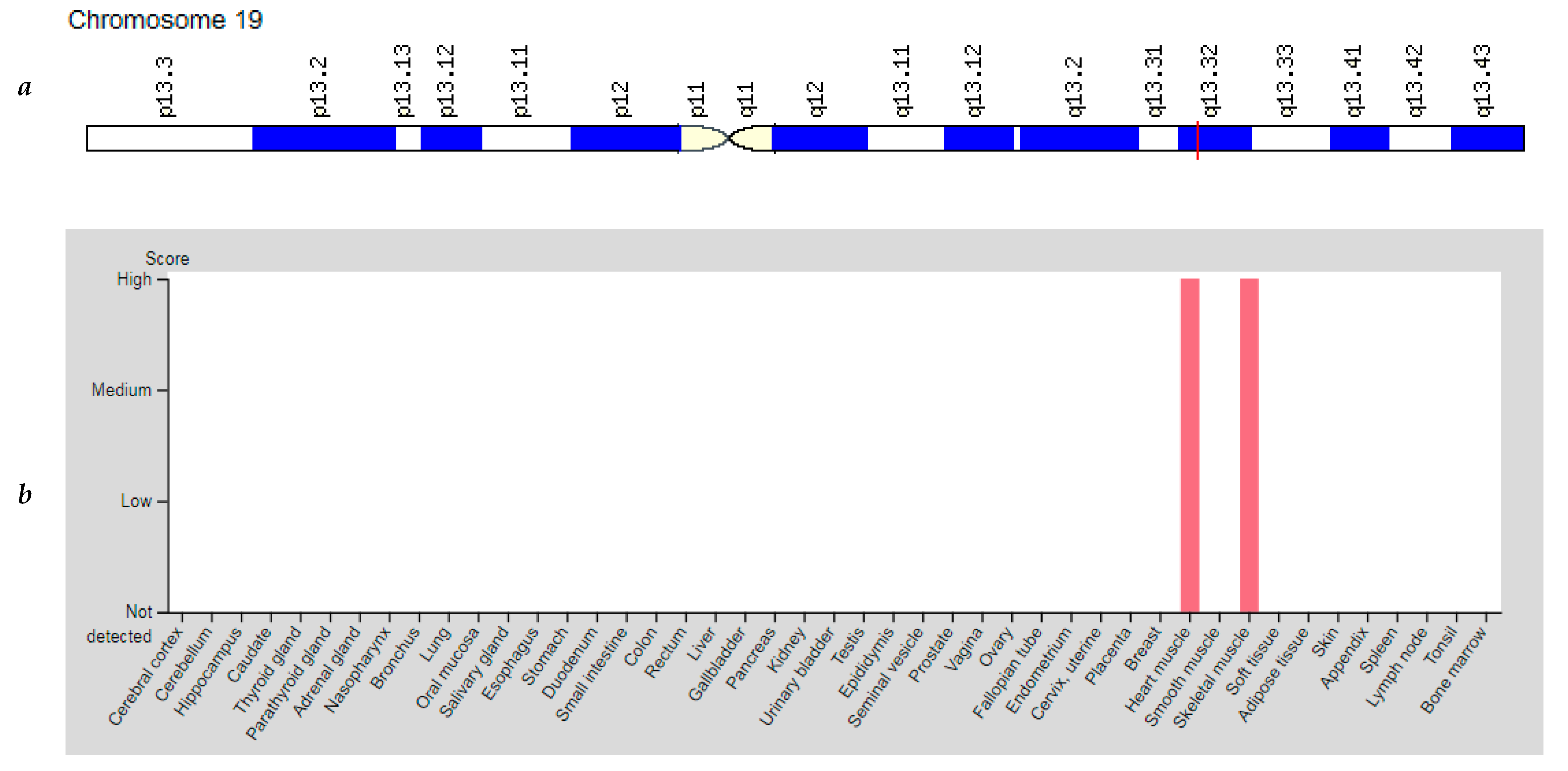
3.5. CKM Gene
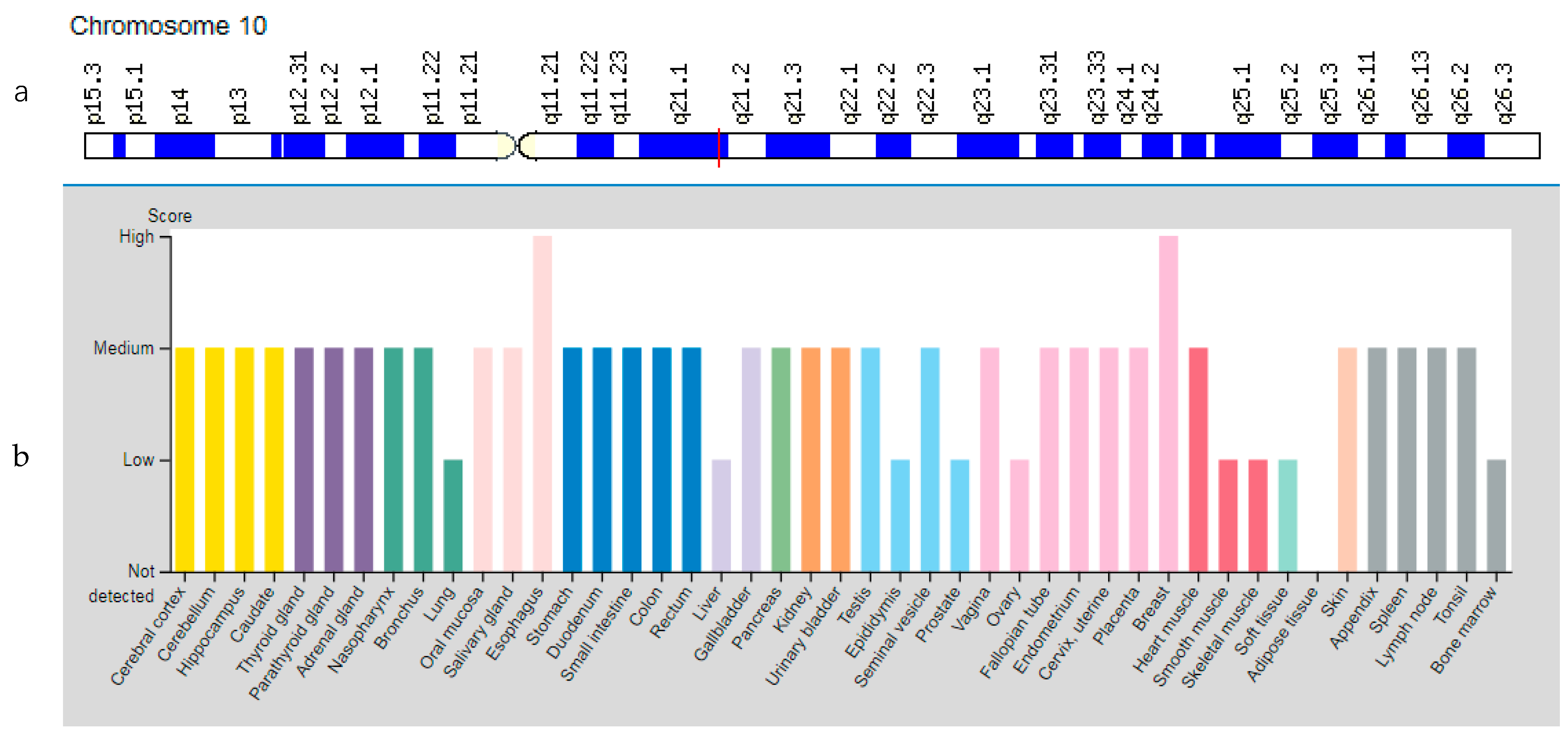
3.6. TFAM Gene
4. Discussion
5. Limitations
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Balberova, O.V.; Sidorkina, E.G.; Koshkina, K.S.; Plachy, J.K.; Bykov, E.V. Model characteristics of competition performance in terms of atletes, functional fitness. Sci. Educ. Today 2021, 3, 161–176. [Google Scholar] [CrossRef]
- Kryazhev, V.D.; Kryazheva, S.V.; Alenurov, E.A.; Bokova, L.V. Competitive and training areas in cyclical locomotion at top-qualified athletes. Sci. Lett. P.F. Lesgaft Univ. 2020, 10, 205–213. [Google Scholar] [CrossRef]
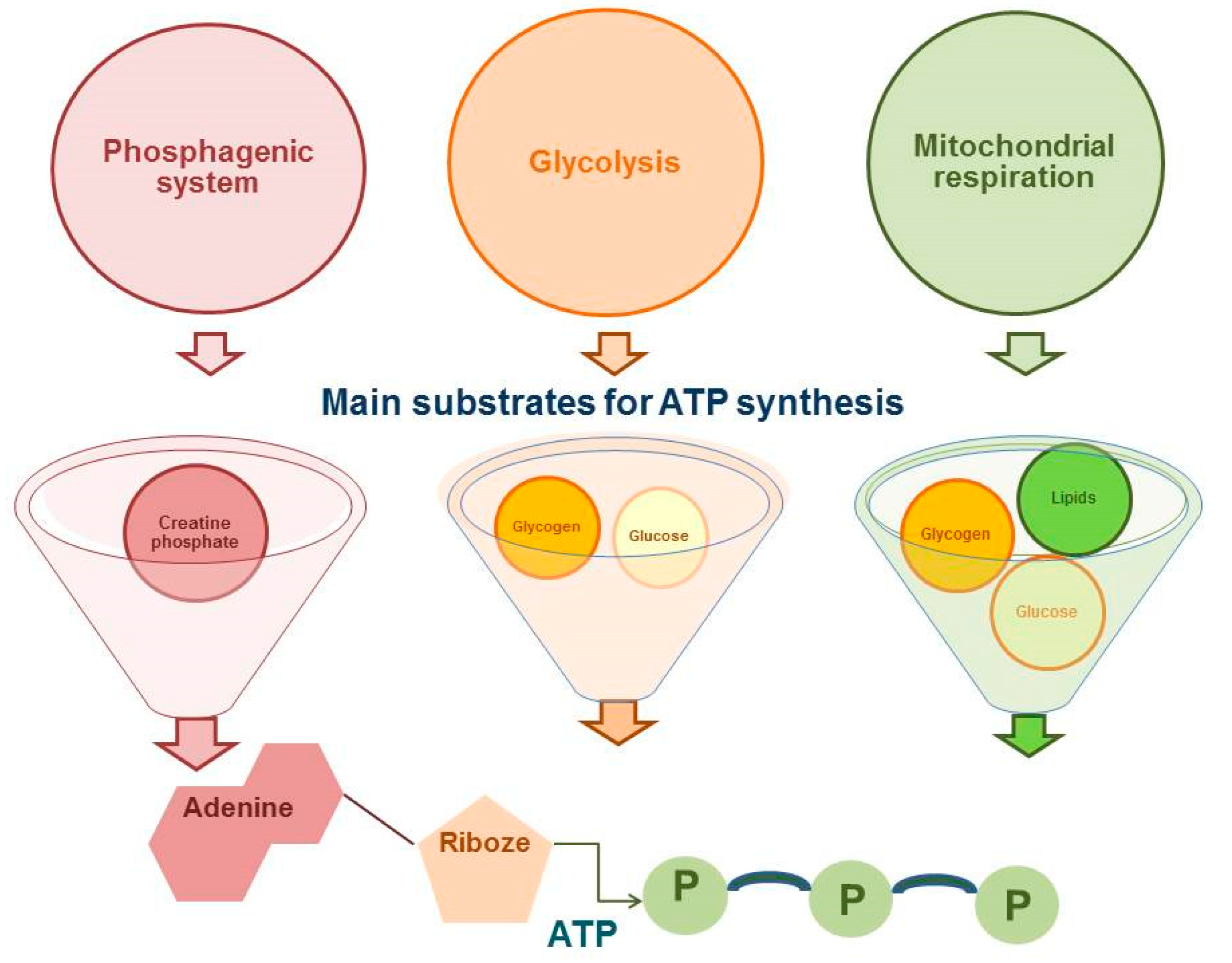
- Hargreaves, M.; Spriet, L.L. Skeletal muscle energy metabolism during exercise. Nat. Metab. 2020, 2, 817–828. [Google Scholar] [CrossRef]
- Baker, S.; Mc Cormick, M.C.; Robergs, R.A. Interaction among Skeletal Muscle Metabolic Energy Systems during Intense Exercise. J. Nutr. Metab. 2010, 2010, 905612. [Google Scholar] [CrossRef] [Green Version]
- Balberova, O.V. Candidate genes and single-nucleotide gene variants associated with muscle and tendon injuries in cyclic sports athletes. Pers. Psychiatry Neurol. 2021, 1, 64–72. [Google Scholar] [CrossRef]
- Fedotovskaya, O.N.; Danilova, A.A.; Akhmetov, I.I. Effect of AMPD1 gene polymorphism on muscle activity in humans. Bull. Exp. Biol. Med. 2013, 154, 489–491. [Google Scholar] [CrossRef]
- Tharabenjasin, P.; Pabalan, N.; Jarjanazi, H. Association of PPARGC1A Gly428Ser (rs8192678) polymorphism with potential for athletic ability and sports performance: A meta-analysis. PLoS ONE 2019, 14, e0200967. [Google Scholar] [CrossRef] [PubMed]
- Meijer, J.P.; Jaspers, R.T.; Rittweger, J.; Seynnes, O.R.; Kamandulis, S.; Brazaitis, M.; Skurvydas, A.; Pišot, R.; Šimunič, B.; Narici, M.V.; et al. Single muscle fibre contractile properties differ between body-builders, power athletes and control subjects. Exp. Physiol. 2015, 100, 1331–1341. [Google Scholar] [CrossRef] [PubMed]
- McKusick, V.A. OMIM: Online Mendelian Inheritance in Man. Available online: https://omim.org/entry/123310 (accessed on 17 July 2021).
- GeneCards: The Human Gene Database. Available online: https://www.genecards.org/ (accessed on 20 August 2021).
- GTEx Portal. Available online: https://www.gtexportal.org/ (accessed on 28 August 2021).
- Rubio, J.C.; Martín, M.A.; Rabadán, M.; Gómez-Gallego, F.; San Juan, A.F.; Alonso, J.M.; Chicharro, J.L.; Pérez, M.; Arenas, J.; Lucia, A. Frequency of the C34T mutation of the AMPD1 gene in world-class endurance athletes: Does this mutation impair performance? J. Appl. Physiol. 2005, 98, 2108–2112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmetov, I.I.; Egorova, E.S.; Gabdrakhmanova, L.J.; Fedotovskaya, O.N. Genes and athletic performance: An Update. Med. Sport Sci. 2016, 61, 41–54. [Google Scholar] [CrossRef] [PubMed]
- Norman, B.; Mahnke-Zizelman, D.K.; Vallis, A.; Sabina, R.L. Genetic and other determinants of AMP deaminase activity in healthy adult skeletal muscle. J. Appl. Physiol. 1998, 85, 1273–1278. [Google Scholar] [CrossRef]
- Ginevičienė, V.; Jakaitienė, A.; Pranculis, A.; Milašius, K.; Tubelis, L.; Utkus, A. AMPD1 rs17602729 is associated with physical performance of sprint and power in elite Lithuanian athletes. BMC Genet. 2014, 15, 58. [Google Scholar] [CrossRef] [Green Version]
- Ahmetov, I.I.; Mozhayskaya, I.A.; Lyubaeva, E.V.; Vinogradova, O.L.; Rogozkin, V.A. PPARG Gene polymorphism and locomotor activity in humans. Bull. Exp. Biol. Med. 2008, 146, 630–632. [Google Scholar] [CrossRef] [PubMed]
- Meirhaeghe, A.; Crowley, V.; Lenaghan, C.; Lelliott, C.; Green, K.; Stewart, A.; Hart, K.; Schinner, S.; Sethi, J.K.; Yeo, G.; et al. Characterization of the human, mouse and rat PGC1 beta (peroxisome-proliferator-activated receptor-gamma co-activator 1 beta) gene in vitro and in vivo. Biochem. J. 2003, 373 Pt 1, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Bray, M.S.; Hagberg, J.M.; Pérusse, L.; Rankinen, T.; Roth, S.M.; Wolfarth, B.; Bouchard, C. The human gene map for performance and health-related fitness phenotypes: The 2006–2007 update. Med. Sci. Sports Exerc. 2009, 41, 35–73. [Google Scholar] [CrossRef] [PubMed]
- Semple, R.K.; Chatterjee, V.K.; O’Rahilly, S. PPAR gamma and human metabolic disease. J. Clin. Investig. 2006, 116, 581–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stumvoll, M.; Häring, H. The peroxisome proliferator-activated receptor-gamma2 Pro12Ala polymorphism. Diabetes 2002, 51, 2341–2347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaccaro, O.; Lapice, E.; Monticelli, A.; Giacchetti, M.; Castaldo, I.; Galasso, R.; Pinelli, M.; Donnarumma, G.; Rivellese, A.A.; Cocozza, S.; et al. Pro12Ala polymorphism of the PPARgamma2 locus modulates the relationship between energy intake and body weight in type 2 diabetic patients. Diabetes Care 2007, 30, 1156–1161. [Google Scholar] [CrossRef] [Green Version]
- Kahara, T.; Takamura, T.; Hayakawa, T.; Nagai, Y.; Yamaguchi, H.; Katsuki, T.; Katsuki, K.; Katsuki, M.; Kobayashi, K. PPARgamma gene polymorphism is associated with exercise-mediated changes of insulin resistance in healthy men. Metabolism 2003, 52, 209–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ilyutik, A.; Gilep, I. The relationship of gene polymorphisms with the development of physical qualities in athletes (based on the material of speed skating). Sci. Olymp. Sports 2017, 3, 51–57. (In Russian) [Google Scholar]
- Mansoori, A.; Amini, M.; Kolahdooz, F.; Seyedrezazadeh, E. Obesity and Pro12Ala Polymorphism of Peroxisome Proliferator-Activated Receptor-gamma Gene in Healthy Adults: A Systematic Review and Meta-Analysis. Ann. Nutr. Metab. 2015, 67, 104–118. [Google Scholar] [CrossRef] [PubMed]
- Akhmetov, I.I.; Popov, D.V.; Mozhaĭskaia, I.A.; Missina, S.S.; Astratenkova, I.V.; Vinogradova, O.L.; Rogozkin, V.A. Association of regulatory genes polymorphisms with aerobic and anaerobic performance of athletes. Ross. Fiziol. Zhurnal Im. IM Sechenova 2007, 93, 837–843. [Google Scholar]
- Petr, M.; Maciejewska-Skrendo, A.; Zajac, A.; Chycki, J.; Stastny, P. Association of Elite Sports Status with Gene Variants of Peroxisome Proliferator Activated Receptors and Their Transcriptional Coactivator. Int. J. Mol. Sci. 2019, 21, 162. [Google Scholar] [CrossRef] [Green Version]
- Maciejewska-Karlowska, A.; Sawczuk, M.; Cieszczyk, P.; Zarebska, A.; Sawczyn, S. Association between the Pro12Ala polymorphism of the peroxisome proliferator-activated receptor gamma gene and strength athlete status. PLoS ONE 2013, 8, e67172. [Google Scholar] [CrossRef] [PubMed]
- Handschin, C.; Spiegelman, B.M. The role of exercise and PGC1alpha in inflammation and chronic disease. Nature 2008, 454, 463–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmetov, I.I.; Mozhayskaya, I.A.; Flavell, D.M.; Astratenkova, I.V.; Komkova, A.I.; Lyubaeva, E.V.; Tarakin, P.P.; Shenkman, B.S.; Vdovina, A.B.; Netreba, A.I.; et al. PPARalpha gene variation and physical performance in Russian athletes. Eur. J. Appl. Physiol. 2006, 97, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Mathai, A.S.; Bonen, A.; Benton, C.R.; Robinson, D.L.; Graham, T.E. Rapid exercise-induced changes in PGC-1alpha mRNA and protein in human skeletal muscle. J. Appl. Physiol. 2008, 105, 1098–1105. [Google Scholar] [CrossRef] [Green Version]
- Sharma, R.; Matharoo, K.; Kapoor, R.; Bhanwer, A.J.S. Association of PGC-1α gene with type 2 diabetes in three unrelated endogamous groups of North-West India (Punjab): A case-control and meta-analysis study. Mol. Genet. Genom. 2018, 293, 317–329. [Google Scholar] [CrossRef]
- Yang, Y.; Mo, X.; Chen, S.; Lu, X.; Gu, D. Association of peroxisome proliferator-activated receptor gamma coactivator 1 Alpha (PPARGC1A) gene polymorphisms and type 2 diabetes mellitus: A meta-analysis. Diabetes Metab. Res. Rev. 2011, 27, 177–184. [Google Scholar] [CrossRef]
- Caló, M.C.; Vona, G. Gene polymorphisms and elite athletic performance. J. Anthropol. Sci. 2008, 86, 113–131. [Google Scholar]
- Maciejewska, A.; Sawczuk, M.; Cieszczyk, P.; Mozhayskaya, I.A.; Ahmetov, I.I. The PPARGC1A gene Gly482Ser in Polish and Russian athletes. J. Sports Sci. 2012, 30, 101–113. [Google Scholar] [CrossRef] [PubMed]
- Olesen, J.; Kiilerich, K.; Pilegaard, H. PGC-1alpha-mediated adaptations in skeletal muscle. Pflüg. Arch. 2010, 460, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Ahmetov, I.I.; Williams, A.G.; Popov, D.V.; Lyubaeva, E.V.; Hakimullina, A.M.; Fedotovskaya, O.N.; Mozhayskaya, I.A.; Vinogradova, O.L.; Astratenkova, I.V.; Montgomery, H.E.; et al. The combined impact of metabolic gene polymorphisms on elite endurance athlete status and related phenotypes. Hum. Genet. 2009, 126, 751–761. [Google Scholar] [CrossRef]
- Cieszczyk, P.; Sawczuk, M.; Maciejewska, A.; Ficek, K.; Eider, J. Variation in peroxisome proliferator activated receptor α gene in elite combat athletes. Eur. J. Sport Sci. 2011, 11, 119–123. [Google Scholar] [CrossRef]
- Tural, E.; Kara, N.; Agaoglu, S.A.; Elbistan, M.; Tasmektepligil, M.Y.; Imamoglu, O. PPAR-α and PPARGC1A gene variants have strong effects on aerobic performance of Turkish elite endurance athletes. Mol. Biol. Rep. 2014, 41, 5799–5804. [Google Scholar] [CrossRef] [PubMed]
- Eynon, N.; Ruiz, J.R.; Oliveira, J.; Duarte, J.A.; Birk, R.; Lucia, A. Genes and elite athletes: A roadmap for future research. J. Physiol. 2011, 589 Pt 13, 3063–3070. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Leon, S.; Tuvblad, C.; Forero, D.A. Sports genetics: The PPARA gene and athletes’ high ability in endurance sports. A systematic review and meta-analysis. Biol. Sport 2016, 33, 3–6. [Google Scholar] [CrossRef]
- Drozdovska, S.B.; Dosenko, V.E.; Ahmetov, I.I.; Ilyin, V.N. The association of gene polymorphisms with athlete status in ukrainians. Biol. Sport 2013, 30, 163–167. [Google Scholar] [CrossRef] [Green Version]
- Maciejewska, A.; Sawczuk, M.; Cięszczyk, P. Variation in the PPARα gene in Polish rowers. J. Sci. Med. Sport 2011, 14, 58–64. [Google Scholar] [CrossRef]
- Gronek, P.; Holdys, J.; Kryściak, J.; Stanisławski, D. CKM Gene G (Ncoi-) Allele Has a Positive Effect on Maximal Oxygen Uptake in Caucasian Women Practicing Sports Requiring Aerobic and Anaerobic Exercise Metabolism. J. Hum. Kinet. 2013, 39, 137–145. [Google Scholar] [CrossRef]
- Zhou, D.Q.; Hu, Y.; Liu, G.; Gong, L.; Xi, Y.; Wen, L. Muscle-specific creatine kinase gene polymorphism and running economy responses to an 18-week 5000-m training programme. Br. J. Sports Med. 2006, 40, 988–991. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Sun, Y.; Liang, H.; Yu, D.; Hu, S. A meta-analysis of the association of CKM gene rs8111989 polymorphism with sport performance. Biol. Sport 2017, 34, 323–330. [Google Scholar] [CrossRef] [PubMed]
- Baird, M.F.; Graham, S.M.; Baker, J.S.; Bickerstaff, G.F. Creatine-kinase- and exercise-related muscle damage implications for muscle performance and recovery. J. Nutr. Metab. 2012, 20, 960363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, E.P.; Li, Y.H.; Qian, J.D.; Yan, H.W. Association of CKMM gene A/G polymorphism and athletic performance of uyghurnationality. Zhongguo Ying Yong Sheng Li Xue Za Zhi 2016, 32, 82–86. (In Chinese) [Google Scholar] [PubMed]
- Rivera, M.A.; Dionne, F.T.; Wolfarth, B.; Chagnon, M.; Simoneau, J.A.; Pérusse, L.; Boulay, M.R.; Gagnon, J.; Song, T.M.; Keul, J.; et al. Muscle-specific creatine kinase gene polymorphisms in elite endurance athletes and sedentary controls. Med. Sci. Sports Exerc. 1997, 29, 1444–1447. [Google Scholar] [CrossRef] [PubMed]
- Heled, Y.; Bloom, M.S.; Wu, T.J.; Stephens, Q.; Deuster, P.A. CK-MM and ACE genotypes and physiological prediction of the creatine kinase response to exercise. J. Appl. Physiol. 2007, 103, 504–510. [Google Scholar] [CrossRef]
- Fedotovskaia, O.N.; Popov, D.V.; Vinogradova, O.L.; Akhmetov, I.I. Association of the muscle-specific creatine kinase (CKMM) gene polymorphism with physical performance of athletes. Fiziol Cheloveka 2012, 38, 105–109. [Google Scholar] [CrossRef]
- Akhmetov, I.I.; Popov, D.V.; Missina, S.S.; Vinogradova, O.L.; Rogozkin, V.A. Association of the mitochondrial transcription factor (TFAM) gene polymorphism with physical performance of athletes. Fiziol Cheloveka 2010, 36, 121–125. [Google Scholar]
- Theilen, N.T.; Kunkel, G.H.; Tyagi, S.C. The Role of Exercise and TFAM in Preventing Skeletal Muscle Atrophy. J. Cell. Physiol. 2017, 232, 2348–2358. [Google Scholar] [CrossRef]
- Garatachea, N.; Pareja-Galeano, H.; Sanchis-Gomar, F.; Santos-Lozano, A.; Fiuza-Luces, C.; Morán, M.; Emanuele, E.; Joyner, M.J.; Lucia, A. Exercise attenuates the major hallmarks of aging. Rejuvenation Res. 2015, 18, 57–89. [Google Scholar] [CrossRef] [Green Version]
- Yan, Z.; Lira, V.A.; Greene, N.P. Exercise training-induced regulation of mitochondrial quality. Exerc. Sport Sci. Rev. 2012, 40, 159–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gordon, J.W.; Rungi, A.A.; Inagaki, H.; Hood, D.A. Effects of contractile activity on mitochondrial transcription factor A expression in skeletal muscle. J. Appl. Physiol. 2001, 90, 389–396. [Google Scholar] [CrossRef] [PubMed]
- Ekstrand, M.I.; Falkenberg, M.; Rantanen, A.; Park, C.B.; Gaspari, M.; Hultenby, K.; Rustin, P.; Gustafsson, C.M.; Larsson, N.G. Mitochondrial transcription factor A regulates mtDNA copy number in mammals. Hum. Mol. Genet. 2004, 13, 935–944. [Google Scholar] [CrossRef] [PubMed]
- Larsson, N.G.; Wang, J.; Wilhelmsson, H.; Oldfors, A.; Rustin, P.; Lewandoski, M.; Barsh, G.S.; Clayton, D.A. Mitochondrial transcription factor A is necessary for mtDNA maintenance and embryogenesis in mice. Nat. Genet. 1998, 18, 231–236. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Hu, Y.; Feng, L.; Bao, D.; Xi, Y.; Wen, L.; Lucia, A. Relationship between TFAM gene polymorphisms and endurance capacity in response to training. Int. J. Sports Med. 2007, 28, 1059–1064. [Google Scholar] [CrossRef] [PubMed]
- Mavlyanov, I.; Parpiev, S.; Sadikov, A.; Kurganov, S.; Makhmudov, D. Relative features of the PPARA (rs4253778), PPARGC1A(rs8192678) and PPARG2(rs1801282) polimorfism genes in athletes engaged in cyclic types of sports. Eur. J. Mol. Clin. Med. 2020, 7, 1860–1869. Available online: https://ejmcm.com/article_2395.html (accessed on 12 June 2021).









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene: MIM (Protein) | Location | Clinical Manifestations of Mutation: MIM | Inheritance |
|---|---|---|---|
| AMPD1: 102770 (Adenosine monophosphate deaminase 1 type) | 1p13.2 (16 exons) | Myopathy Due to Myoadenylate Deaminase Deficiency: 615511 | AD |
| CKM: 123310 (Creatine Kinase, Muscle) | 19q13.32 (8 exons) | Myotonic Dystrophy: 160900 | AD |
| PPARA: 170998 (Peroxisome Proliferator Activated Receptor Alpha) | 22q13.31 (14 exons) | Fatty Liver Disease: 613282 | Mu |
| PPARG: 601487 (Peroxisome proliferator activated receptor gamma) | 3p25.2 (14 exons) | Lipodystrophy, Obesity, Diabetes Mellitus Type 2 Familial Partial Lipodystrophy Type 3: 604367 | Mu AD |
| PPARGC1A: 604517 (Peroxisome Proliferator-Activated Receptor Gamma Coactivator 1 Alpha) | 4p15.2 (24 exons) | Amyotrophic Lateral Sclerosis Type 1: 105400 Huntington Disease: 604802 | AD/AR AR |
| TFAM: 600438 (Transcription Factor A, Mitochondrial) | 10q21.1 (8 exons) | Mitochondrial DNA Depletion Syndrome Type 15: 617156 | AR |
| Gene | Localization, Chromosome | Protein/ Enzyme | Effects on Energy Metabolism of Skeletal Muscle |
|---|---|---|---|
| AMPD1 | 1p13.2 | Adenosine monophosphate deaminase 1 type (AMPD1) | AMPD1 catalyzes the deamination of adenosine monophosphate and inosine monophosphate in skeletal muscle and plays an important role in the purine nucleotide cycle. |
| PPARG | 3p25.2 | Peroxisome proliferator activated receptor gamma (PPARG) | PPARG controls the peroxisome beta-oxidation pathway of fatty acids and is a key regulator of adipocyte differentiation and glucose homeostasis. |
| PPARGC1A | 4p15.2 | Peroxisome Proliferator-Activated Receptor Gamma Coactivator 1 Alpha (PPARGC1A) | PPARGC1A regulates genes involved in energy metabolism, provides a direct link between external physiological stimuli and regulation of mitochondrial biogenesis, and is the main factor regulating the determination of muscle fiber type. |
| PPARA | 22q13.31 | Peroxisome Proliferator Activated Receptor Alpha (PPARA) | PPARA is involved in the regulation of energy metabolism, regulates the expression of genes encoding several key muscle enzymes involved in fatty acid oxidation. |
| CKM | 19q13.32 | Creatine Kinase, Muscle (CKM) | CKM catalyzes the transfer of phosphate between ATP and various phosphogenic groups such as creatine phosphate; CKM isozymes play a central role in energy transduction in tissues with high energy requirements such as skeletal muscle, heart, brain. |
| TFAM | 10q21.1 | Transcription Factor A, Mitochondrial (TFAM) | TFAM is responsible for regulating mitochondrial DNA replication and transcription and also protects cells from oxidative stress. |
| Gene | Expression Level in Skeletal Muscles (RPKM) | Expression Level in Myocardium (RPKM) | Expression Level in Lung (RPKM) |
|---|---|---|---|
| AMPD1 | 225.7 | 0.023 | 0.363 |
| PPARG | 2.097 | 4.716 | 19.45 |
| PPARGC1A | 11.02 | 9.406 | 3.292 |
| PPARA | 12.34 | 7.889 | 5.624 |
| CKM | 25890.0 | 2987 | 4.667 |
| TFAM | 6.0 | 4.754 | 9.471 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Balberova, O.V.; Bykov, E.V.; Medvedev, G.V.; Zhogina, M.A.; Petrov, K.V.; Petrova, M.M.; Al-Zamil, M.; Trefilova, V.V.; Goncharova, P.S.; Shnayder, N.A. Candidate Genes of Regulation of Skeletal Muscle Energy Metabolism in Athletes. Genes 2021, 12, 1682. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12111682
Balberova OV, Bykov EV, Medvedev GV, Zhogina MA, Petrov KV, Petrova MM, Al-Zamil M, Trefilova VV, Goncharova PS, Shnayder NA. Candidate Genes of Regulation of Skeletal Muscle Energy Metabolism in Athletes. Genes. 2021; 12(11):1682. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12111682
Chicago/Turabian StyleBalberova, Olga V., Evgeny V. Bykov, German V. Medvedev, Margarita A. Zhogina, Kirill V. Petrov, Marina M. Petrova, Mustafa Al-Zamil, Vera V. Trefilova, Polina S. Goncharova, and Natalia A. Shnayder. 2021. "Candidate Genes of Regulation of Skeletal Muscle Energy Metabolism in Athletes" Genes 12, no. 11: 1682. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12111682