
First DNA Sequencing in Beninese Indigenous Cattle Breeds Captures New Milk Protein Variants

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Sampling, DNA Extraction and Sequencing
2.2. Processing of Sequence Data
2.3. Functional Annotation of DNA Sequence Variants, Frequencies of SNP Alleles and Casein Haplotypes
3. Results
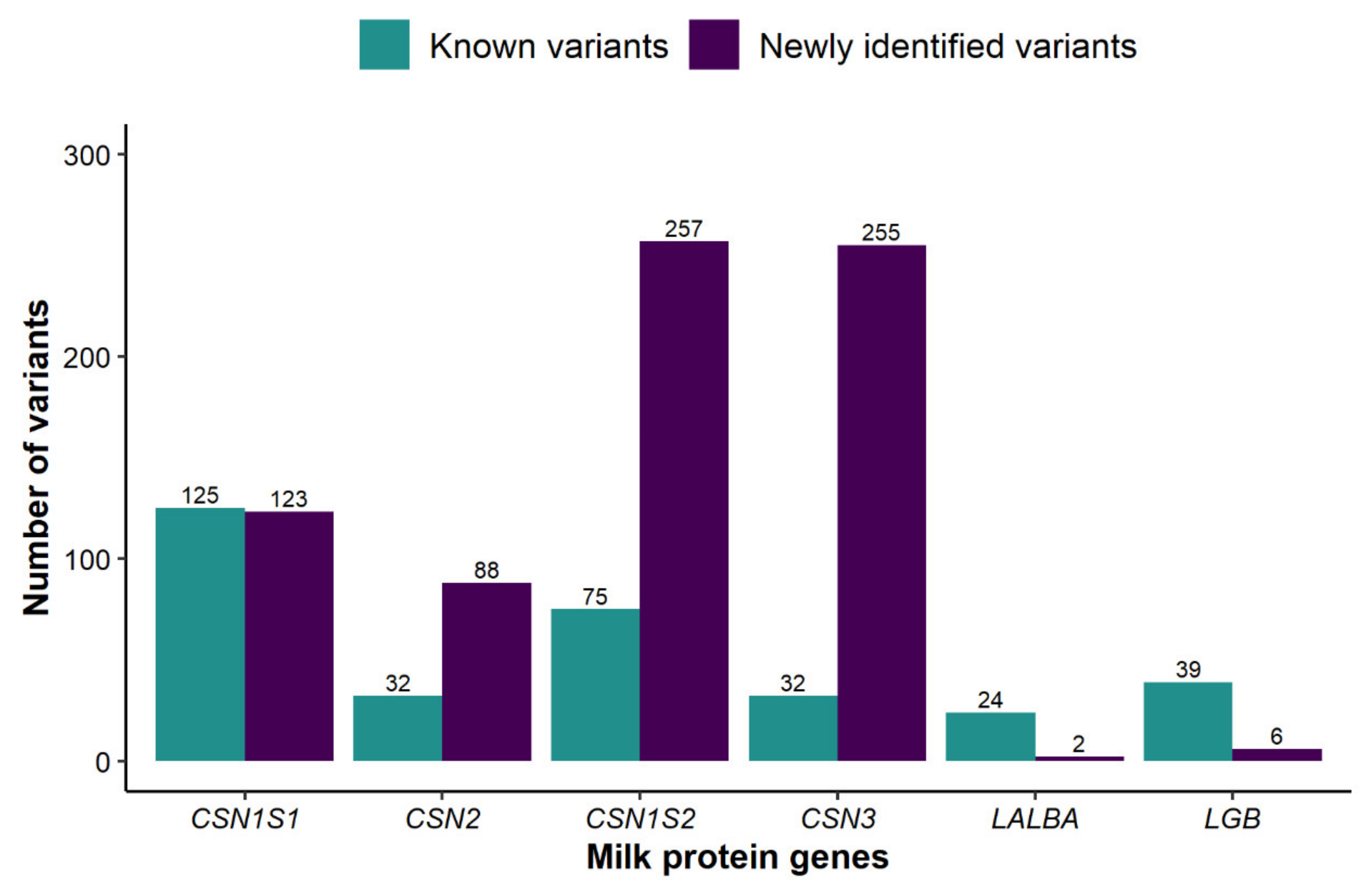
3.1. Distribution of DNA Sequence Variants
3.2. Causal Genetic Polymorphisms of Milk Protein Variants
3.2.1. CSN1S1
3.2.2. CSN2
3.2.3. CSN1S2
3.2.4. CSN3
3.2.5. LALBA
3.2.6. LGB
3.3. Casein Haplotypes
3.4. Functional Effects of Variants in the Splice Sites, 5′- and 3′-Untranslated Regions of Milk Protein Genes
4. Discussion
4.1. Sequence Polymorphisms and Milk Proteins in Beninese Cattle
4.1.1. CSN1S1
4.1.2. CSN2
4.1.3. CSN1S2
4.1.4. CSN3
4.1.5. LALBA
4.1.6. LGB
4.2. Casein Haplotypes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alassane, Y.; Ahounou, S.; Toleba, S.; Adjakpa, A.; Dotche, I.; Houaga, I.; Moula, N.; AntoineMoussiaux, N.; Hornick, J.; Youssao, A. Zootechnical performance of Girolando cattle at Kpinnou Breeding Farm, South-West of Benin Republic. J. Adv. Veter-Anim. Res. 2018, 5, 123. [Google Scholar] [CrossRef]
- Alk, I.T.; Yari, H.M.; Gbangboche, A.B.; Lokossou, R. Reproductive Performance and Milk Production of Girolando Cows in the Ranch of Kpinnou, South-West of Benin Republic. J. Anim. Veter-Adv. 2011, 10, 2588–2592. [Google Scholar] [CrossRef]
- Kévin, K.S.; Charles, D.G.-K.; Valentine, Y.-G.; Souleymane, S.; Maurice, K.; Issaka, Y.A.K. Genetic Diversity of Benin Cattle Populations Using Microsatellite Markers. Int. J. Anim. Sci. Technol. 2019, 3, 7. [Google Scholar] [CrossRef]
- Vanvanhossou, S.F.U.; Koura, I.B.; Dossa, L.H. The implications of herd entrustment practice for the sustainable use of cattle genetic resources in the (agro)-pastoral systems of West Africa: A case study from Benin. Pastoralism 2021, 11, 1–14. [Google Scholar] [CrossRef]
- Houessou, S.O.; Dossa, L.H.; Assogba, C.A.; Diogo, R.V.C.; Vanvanhossou, S.F.U.S.; Schlecht, E. The role of cross-border transhumance in influencing resident herders’ cattle husbandry practices and use of genetic resources. Animal 2020, 14, 2378–2386. [Google Scholar] [CrossRef] [PubMed]
- Vanvanhossou, S.F.U.; Dossa, L.H.; König, S. Sustainable Management of Animal Genetic Resources to Improve Low-Input Livestock Production: Insights into Local Beninese Cattle Populations. Sustainability 2021, 13, 9874. [Google Scholar] [CrossRef]
- Dossa, L.H.; Vanvanhossou, S.F.U.S. The indigenous Somba cattle of the hilly Atacora region in North-West Benin: Threats and opportunities for its sustainable use. Trop. Anim. Health Prod. 2015, 48, 349–359. [Google Scholar] [CrossRef] [PubMed]
- Hiernaux, P.; Adamou, K.; Zezza, A.; Ayantunde, A.A.; Federighi, G. Milk offtake of cows in smallholder farms of semiarid Sahel: Low yields with high value! Rev. Elev. Med. Vet. Pays. Trop. 2016, 69, 143–153. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, A.S.; Rahmatalla, S.; Bortfeldt, R.; Arends, D.; Reissmann, M.; Brockmann, G.A. Milk protein polymorphisms and casein haplotypes in Butana cattle. J. Appl. Genet. 2016, 58, 261–271. [Google Scholar] [CrossRef] [PubMed]
- Lemay, D.; Lynn, D.J.; Martin, W.F.; Neville, M.C.; Casey, T.M.; Rincon, G.; Kriventseva, E.V.; Barris, W.C.; Hinrichs, A.S.; Molenaar, A.J.; et al. The bovine lactation genome: Insights into the evolution of mammalian milk. Genome Biol. 2009, 10, R43. [Google Scholar] [CrossRef] [PubMed]
- Sebastiani, C.; Arcangeli, C.; Ciullo, M.; Torricelli, M.; Cinti, G.; Fisichella, S.; Biagetti, M. Frequencies Evaluation of β-Casein Gene Polymorphisms in Dairy Cows Reared in Central Italy. Animals 2020, 10, 252. [Google Scholar] [CrossRef] [Green Version]
- Ferretti, L.; Leone, P.; Sgaramella, V. Long range restriction analysis of the bovine casein genes. Nucleic Acids Res. 1990, 18, 6829–6833. [Google Scholar] [CrossRef] [Green Version]
- Farrell, H.M.; Jimenez-Flores, R.; Bleck, G.T.; Brown, E.M.; Butler, J.E.; Creamer, L.K.; Hicks, C.L.; Hollar, C.M.; Ng-Kwai-Hang, K.F.; Swaisgood, H.E. Nomenclature of the Proteins of Cows’ Milk—Sixth Revision. J. Dairy Sci. 2004, 87, 1641–1674. [Google Scholar] [CrossRef] [Green Version]
- Caroli, A.M.; Chessa, S.; Erhardt, G.J. Invited review: Milk protein polymorphisms in cattle: Effect on animal breeding and human nutrition. J. Dairy Sci. 2009, 92, 5335–5352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallinat, J.L.; Qanbari, S.; Drögemüller, C.; Pimentel, E.C.G.; Thaller, G.; Tetens, J. DNA-based identification of novel bovine casein gene variants. J. Dairy Sci. 2013, 96, 699–709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Visker, M.H.P.W.; Heck, J.M.L.; Van Valenberg, H.J.F.; Van Arendonk, J.A.M.; Bovenhuis, H. Short communication: A new bovine milk-protein variant: α-Lactalbumin variant D. J. Dairy Sci. 2012, 95, 2165–2169. [Google Scholar] [CrossRef]
- Mahé, M.-F.; Miranda, G.; Queval, R.; Bado, A.; Zafindrajaona, P.S.; Grosclaude, F. Genetic polymorphism of milk proteins in African Bos taurus and Bos indicus populations. Characterization of variants αs1-Cn H and κ-Cn. J. Genet. Sel. Evol. 1999, 31, 239–253. [Google Scholar] [CrossRef]
- Goudarzi, K.M.; Belemsaga, D.M.; Ceriotti, G.; Laloë, D.; Fagbohoum, F.; Kouagou, N.T.; Sidibé, I.; Codjia, V.; Crimella, M.C.; Grosclaude, F.; et al. Caractérisation de la race bovine Somba à l’aide de marqueurs moléculaires. Rev. Elev. Med. Vet. Pays Trop. 2001, 54, 129. [Google Scholar] [CrossRef] [Green Version]
- Ceriotti, G.; Marletta, D.; Caroli, A.; Erhardt, G. Milk protein loci polymorphism in taurine (Bos taurus) and zebu (Bos indicus) populations bred in hot climate. J. Anim. Breed. Genet. 2004, 121, 404–415. [Google Scholar] [CrossRef]
- Boettcher, P.J.; Caroli, A.; Stella, A.; Chessa, S.; Budelli, E.; Canavesi, F.; Ghiroldi, S.; Pagnacco, G. Effects of Casein Haplotypes on Milk Production Traits in Italian Holstein and Brown Swiss Cattle. J. Dairy Sci. 2004, 87, 4311–4317. [Google Scholar] [CrossRef]
- Bonfatti, V.; Di Martino, G.; Cecchinato, A.; Vicario, D.; Carnier, P. Effects of β-κ-casein (CSN2-CSN3) haplotypes and β-lactoglobulin (BLG) genotypes on milk production traits and detailed protein composition of individual milk of Simmental cows. J. Dairy Sci. 2010, 93, 3797–3808. [Google Scholar] [CrossRef]
- Nilsen, H.; Olsen, H.G.; Hayes, B.; Sehested, E.; Svendsen, M.; Nome, T.; Meuwissen, T.; Lien, S. Casein haplotypes and their association with milk production traits in Norwegian Red cattle. Genet. Sel. Evol. 2009, 41, 24. [Google Scholar] [CrossRef] [Green Version]
- Hohmann, L.G.; Weimann, C.; Scheper, C.; Erhardt, G.; König, S. Associations between maternal milk protein genotypes with preweaning calf growth traits in beef cattle. J. Anim. Sci. 2020, 98, skaa280. [Google Scholar] [CrossRef] [PubMed]
- Deth, R.; Clarke, A.; Ni, J.; Trivedi, M. Clinical evaluation of glutathione concentrations after consumption of milk containing different subtypes of β-casein: Results from a randomized, cross-over clinical trial. Nutr. J. 2015, 15, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Ionel, N.R.; Elena, I.D.; Toma, C.L. Effects of Sires Genotyping for κ-Casein on B Allele Frequency in Romanian Brown Cattle. Res. J. Biotechnol. 2017, 12, 12. [Google Scholar]
- Losi, G.; Capella, P.; Castagnetti, G.B.; Grazia, L.; Zambonelli, C.; Mariani, P.; Russo, V. Influenza delle varianti genetiche della caseina k sulla formazione e sulle caratteristiche della cagliata. Sci. Tecnol. Aliment. 1973, 3, 373. [Google Scholar]
- Martin, P.; Szymanowska, M.; Zwierzchowski, L.; Leroux, C. The impact of genetic polymorphisms on the protein composition of ruminant milks. Reprod. Nutr. Dev. 2002, 42, 433–459. [Google Scholar] [CrossRef] [PubMed]
- Comin, A.; Cassandro, M.; Chessa, S.; Ojala, M.; Zotto, R.D.; De Marchi, M.; Carnier, P.; Gallo, L.; Pagnacco, G.; Bittante, G. Effects of Composite β- and κ-Casein Genotypes on Milk Coagulation, Quality, and Yield Traits in Italian Holstein Cows. J. Dairy Sci. 2008, 91, 4022–4027. [Google Scholar] [CrossRef] [PubMed]
- Heck, J.M.L.; Schennink, A.; van Valenberg, H.; Bovenhuis, H.; Visker, M.H.P.W.; van Arendonk, J.; Van Hooijdonk, A.C.M. Effects of milk protein variants on the protein composition of bovine milk. J. Dairy Sci. 2009, 92, 1192–1202. [Google Scholar] [CrossRef]
- Braunschweig, M.; Hagger, C.; Stranzinger, G.; Puhan, Z. Associations Between Casein Haplotypes and Milk Production Traits of Swiss Brown Cattle. J. Dairy Sci. 2000, 83, 1387–1395. [Google Scholar] [CrossRef]
- Szymanowska, M.; Malewski, T.; Zwierzchowski, L. Transcription factor binding to variable nucleotide sequences in 5′-flanking regions of bovine casein genes. Int. Dairy J. 2004, 14, 103–115. [Google Scholar] [CrossRef]
- Chan, T.-M.; Leung, K.-S.; Lee, K.-H. TFBS identification based on genetic algorithm with combined representations and adaptive post-processing. Bioinformatics 2007, 24, 341–349. [Google Scholar] [CrossRef] [Green Version]
- Boeva, V. Analysis of Genomic Sequence Motifs for Deciphering Transcription Factor Binding and Transcriptional Regulation in Eukaryotic Cells. Front. Genet. 2016, 7, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ha, M.; Kim, V.N. Regulation of microRNA biogenesis. Nat. Rev. Mol. Cell Biol. 2014, 15, 509–524. [Google Scholar] [CrossRef] [PubMed]
- Broughton, J.; Lovci, M.T.; Huang, J.L.; Yeo, E.; Pasquinelli, A.E. Pairing beyond the Seed Supports MicroRNA Targeting Specificity. Mol. Cell 2016, 64, 320–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibeagha-Awemu, E.M.; Kgwatalala, P.; Zhao, X. A critical analysis of production-associated DNA polymorphisms in the genes of cattle, goat, sheep, and pig. Mamm. Genome 2008, 19, 591–617. [Google Scholar] [CrossRef] [PubMed]
- Szymanowska, M.; Siadkowska, E.; Lukaszewicz, M.; Zwierzchowski, L. Association of nucleotide-sequence polymorphism in the 5’-flanking regions of bovine casein genes with casein content in cow’s milk. Le Lait 2004, 84, 579–590. [Google Scholar] [CrossRef] [Green Version]
- Kishore, A.; Sodhi, M.; Mukesh, M.; Mishra, B.P.; Sobti, R.C. Sequence analysis and identification of new variations in the 5′-flanking region of αS2-casein gene in Indian zebu cattle. Mol. Biol. Rep. 2013, 40, 4473–4481. [Google Scholar] [CrossRef]
- Ganai, N.; Bovenhuis, H.; Van Arendonk, J.A.M.; Visker, M.H.P.W. Novel polymorphisms in the bovineβ-lactoglobulingene and their effects onβ-lactoglobulin protein concentration in milk. Anim. Genet. 2009, 40, 127–133. [Google Scholar] [CrossRef]
- Lum, L.S.; Dovč, P.; Medrano, J.F. Polymorphisms of Bovine β-Lactoglobulin Promoter and Differences in the Binding Affinity of Activator Protein-2 Transcription Factor. J. Dairy Sci. 1997, 80, 1389–1397. [Google Scholar] [CrossRef]
- Ibeagha-Awemu, E.; Prinzenberg, E.-M.; Erhardt, G. High variability of milk protein genes in Bos indicus cattle breeds of Cameroon and Nigeria and characterization of a new αs1-casein promoter allele. J. Dairy Res. 2005, 72, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Prinzenberg, E.-M.; Weimann, C.; Brandt, H.; Bennewitz, J.; Kalm, E.; Schwerin, M.; Erhardt, G. Polymorphism of the Bovine CSN1S1 Promoter: Linkage Mapping, Intragenic Haplotypes, and Effects on Milk Production Traits. J. Dairy Sci. 2003, 86, 2696–2705. [Google Scholar] [CrossRef] [Green Version]
- Jann, O.C.; Ibeagha-Awemu, E.; Özbeyaz, C.; Zaragoza, P.; Williams, J.; Marsan, P.A.; Lenstra, J.A.; Moazami-Goudarzi, K.; Erhardt, G. Geographic distribution of haplotype diversity at the bovine casein locus. Genet. Sel. Evol. 2004, 36, 243–257. [Google Scholar] [CrossRef] [Green Version]
- Scheper, C.; Bohlouli, M.; Brügemann, K.; Weimann, C.; Vanvanhossou, S.F.U.; König, S.; Dossa, L.H. The role of agro-ecological factors and transboundary transhumance in shaping the genetic diversity in four indigenous cattle populations of Benin. J. Anim. Breed. Genet. 2020, 137, 622–640. [Google Scholar] [CrossRef]
- Vanvanhossou, S.F.U.; Scheper, C.; Dossa, L.H.; Yin, T.; Brügemann, K.; König, S. A multi-breed GWAS for morphometric traits in four Beninese indigenous cattle breeds reveals loci associated with conformation, carcass and adaptive traits. BMC Genom. 2020, 21, 783. [Google Scholar] [CrossRef] [PubMed]
- Lühken, G.; Caroli, A.; Ibeagha-Awemu, E.M.; Erhardt, G. Characterization and genetic analysis of bovine αs1-caseinIvariant. Anim. Genet. 2009, 40, 479–485. [Google Scholar] [CrossRef]
- Illumina. bcl2fastq2 Conversion Software: Version 2.20.0. 2017. Available online: https://support.illumina.com/sequencing/sequencing_software/bcl2fastq-conversion-software.html (accessed on 24 June 2021).
- Li, H. Aligning sequence reads, clone sequences and assembly contigs with BWA-MEM. arXiv 2013, arXiv:1303.3997. [Google Scholar]
- McLaren, W.; Gil, L.; Hunt, S.E.; Riat, H.S.; Ritchie, G.R.S.; Thormann, A.; Flicek, P.; Cunningham, F. The Ensembl Variant Effect Predictor. Genome Biol. 2016, 17, 122. [Google Scholar] [CrossRef] [Green Version]
- European Variation Archive. Milk Protein Gene Polymorphisms in the Beninese Indigenous Cattle Breeds. Available online: https://wwwdev.ebi.ac.uk/eva/?eva-study=PRJEB47999 (accessed on 13 October 2021).
- Tetens, J.L. Genetic Variability of Bovine and Ovine Milk Protein Genes. Ph.D. Thesis, Christian-Albrechts-Universität zu Kiel, Kiel, Germany, 2014. [Google Scholar]
- Danecek, P.; Auton, A.; Abecasis, G.; Albers, C.A.; Banks, E.; DePristo, M.A.; Handsaker, R.; Lunter, G.; Marth, G.T.; Sherry, S.T.; et al. The variant call format and VCFtools. Bioinformatics 2011, 27, 2156–2158. [Google Scholar] [CrossRef]
- Sinnwell, J.P.; Schaid, D.J. Statistical Analysis of Haplotypes with Traits and Covariates when Linkage Phase is Ambiguous, R package version 1.8.7; Mayo Clinic Division of Health Sciences Research: Rochester, MN, USA, 2021. [Google Scholar]
- Reese, M.G.; Eeckman, F.H.; Kulp, D.; Haussler, D. Improved Splice Site Detection in Genie. J. Comput. Biol. 1997, 4, 311–323. [Google Scholar] [CrossRef] [PubMed]
- Yevshin, I.; Sharipov, R.; Kolmykov, S.; Kondrakhin, Y.; Kolpakov, F. GTRD: A database on gene transcription regulation—2019 update. Nucleic Acids Res. 2018, 47, D100–D105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolmykov, S.; Yevshin, I.; Kulyashov, M.; Sharipov, R.; Kondrakhin, Y.; Makeev, V.J.; Kulakovskiy, I.V.; Kel, A.; Kolpakov, F. GTRD: An integrated view of transcription regulation. Nucleic Acids Res. 2020, 49, D104–D111. [Google Scholar] [CrossRef]
- Agarwal, V.; Bell, G.W.; Nam, J.-W.; Bartel, D.P. Predicting effective microRNA target sites in mammalian mRNAs. eLife 2015, 4, e05005. [Google Scholar] [CrossRef]
- Lewis, B.P.; Burge, C.B.; Bartel, D.P. Conserved Seed Pairing, Often Flanked by Adenosines, Indicates that Thousands of Human Genes are MicroRNA Targets. Cell 2005, 120, 15–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meier, S.; Korkuć, P.; Arends, D.; Brockmann, G.A. DNA Sequence Variants and Protein Haplotypes of Casein Genes in German Black Pied Cattle (DSN). Front. Genet. 2019, 10, 1129. [Google Scholar] [CrossRef] [PubMed]
- Hanotte, O.; Dessie, T.; Kemp, S. Time to Tap Africa’s Livestock Genomes. Science 2010, 328, 1640–1641. [Google Scholar] [CrossRef] [PubMed]
- Cartegni, L.; Wang, J.; Zhu, Z.; Zhang, M.Q.; Krainer, A.R. ESEfinder: A web resource to identify exonic splicing enhancers. Nucleic Acids Res. 2003, 31, 3568–3571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, M.; Xia, W.; Zhang, X.; Liu, Y.; Jiang, M. Identification and verification of differentially expressed genes in yak mammary tissue during the lactation cycle. J. Dairy Res. 2020, 87, 158–165. [Google Scholar] [CrossRef]
- Sumner-Thomson, J.M.; Vierck, J.L.; McNamara, J.P. Differential expression of genes in adipose tissue of first-lactation dairy cattle. J. Dairy Sci. 2011, 94, 361–369. [Google Scholar] [CrossRef] [PubMed]
- Hindson, C.M.; Chevillet, J.; Briggs, H.A.; Gallichotte, E.N.; Ruf, I.K.; Hindson, B.J.; Vessella, R.L.; Tewari, M. Absolute quantification by droplet digital PCR versus analog real-time PCR. Nat. Methods 2013, 10, 1003–1005. [Google Scholar] [CrossRef] [PubMed]
- Costa-Silva, J.; Domingues, D.; Lopes, F.M. RNA-Seq differential expression analysis: An extended review and a software tool. PLoS ONE 2017, 12, e0190152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lisson, M.; Lochnit, G.; Erhardt, G. In vitro gastrointestinal digestion of bovine αS1- and αS2-casein variants gives rise to different IgE-binding epitopes. Int. Dairy J. 2014, 34, 47–55. [Google Scholar] [CrossRef]
- Giambra, I.J.; Jäger, S.; Erhardt, G. Isoelectric focusing reveals additional casein variants in German sheep breeds. Small Rumin. Res. 2010, 90, 11–17. [Google Scholar] [CrossRef]
- Najafi, M.; Mianji, G.R.; Pirsaraie, Z.A. Cloning and comparative analysis of gene structure in promoter site of alpha-s1 casein gene in Naeinian goat and sheep. Meta Gene 2014, 2, 854–861. [Google Scholar] [CrossRef] [PubMed]
- Cosenza, G.; Pauciullo, A.; Colimoro, L.; Mancusi, A.; Rando, A.; Di Berardino, D.; Ramunno, L. An SNP in the goat CSN2 promoter region is associated with the absence of β-casein in milk. Anim. Genet. 2007, 38, 655–658. [Google Scholar] [CrossRef] [PubMed]
- Ibeagha-Awemu, E.; Prinzenberg, E.M.; Jann, O.C.; Lühken, G.; Ibeagha, A.E.; Zhao, X.; Erhardt, G. Molecular Characterization of Bovine CSN1S2B and Extensive Distribution of Zebu-Specific Milk Protein Alleles in European Cattle. J. Dairy Sci. 2007, 90, 3522–3529. [Google Scholar] [CrossRef]
- Shen, B.; Zhang, L.; Lian, C.; Lu, C.; Zhang, Y.; Pan, Q.; Yang, R.; Zhao, Z. Deep Sequencing and Screening of Differentially Expressed MicroRNAs Related to Milk Fat Metabolism in Bovine Primary Mammary Epithelial Cells. Int. J. Mol. Sci. 2016, 17, 200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Sousa, M.A.P.; de Athayde, F.R.F.; Maldonado, M.B.C.; de Lima, A.O.; Fortes, M.R.S.; Lopes, F.L. Single nucleotide polymorphisms affect miRNA target prediction in bovine. PLoS ONE 2021, 16, e0249406. [Google Scholar] [CrossRef] [PubMed]
- Damiani, G.; Florio, S.; Budelli, E.; Bolla, P.; Caroli, A. Single nucleotide polymorphisms (SNPs) within Bov-A2 SINE in the second intron of bovine and buffalo k-casein (CSN3 ) gene. Anim. Genet. 2000, 31, 277–279. [Google Scholar] [CrossRef]
- Tetens, J.L.; Qanbari, S.; Drögemüller, C.; Pimentel, E.C.G.; Bennewitz, J.; Thaller, G. Bos indicus introgression into (peri-)alpine cattle breeds—evidence from the analysis of bovine whey protein variants. Anim. Genet. 2014, 45, 585–588. [Google Scholar] [CrossRef]
- Bell, K.; Hopper, K.E.; McKenzie, H.A. Bovine a-Lactalbumin C and aS1-, ß- and ?-Caseins of Bali (Banteng) Cattle, Bos (Bihos) Javanicus. Aust. J. Biol. Sci. 1981, 34, 149. [Google Scholar] [CrossRef] [Green Version]
- Pausch, H.; MacLeod, I.M.; Fries, R.; Emmerling, R.; Bowman, P.J.; Daetwyler, H.D.; Goddard, M.E. Evaluation of the accuracy of imputed sequence variant genotypes and their utility for causal variant detection in cattle. Genet. Sel. Evol. 2017, 49, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Medrano, J.F.; Aguilar-Cordova, E. Polymerase chain reaction amplification of bovine β-lactoglobulin genomic sequences and identification of genetic variants by RFLP analysis. Anim. Biotechnol. 1990, 1, 73–77. [Google Scholar] [CrossRef]
- Martin, P.; Ollivier-Bousquet, M.; Grosclaude, F. Genetic polymorphism of caseins: A tool to investigate casein micelle organization. Int. Dairy J. 1999, 9, 163–171. [Google Scholar] [CrossRef]
- Formaggioni, P.; Summer, A.; Malacarne, M.; Mariani, P. Milk protein polymorphism: Detection and diffusion of the genetic variants in Bos genus. Ann. Fac. Med. Vet. Univ. Parma 1999, 19, 127–165. [Google Scholar]
- Mohr, U.; Koczan, D.; Linder, D.; Hobom, G.; Erhardt, G. A single point mutation results in A allele-specific exon skipping in the bovine αs1-casein mRNA. Gene 1994, 143, 187–192. [Google Scholar] [CrossRef]
- Rahmatalla, S.A.; Arends, D.; Ahmed, A.S.; Hassan, L.M.A.; Krebs, S.; Reissmann, M.; Brockmann, G.A. Capture Sequencing to Explore and Map Rare Casein Variants in Goats. Front. Genet. 2021, 12, 620253. [Google Scholar] [CrossRef]
- Rando, A.; Ramunno, L.; Masina, P. Mutations in casein genes. Zootec. Nutr. Anim. 2000, 26, 105–114. [Google Scholar]
- Mariani, P.; Pecorari, M. Fattori genetici, attitudine alla caseificazione e resa del latte in formaggio. Sci. Tecn. Latt.-Cas 1987, 38, 286–326. [Google Scholar]
- Merin, U.; Bernstein, S.; Bloch-Damti, A.; Yagil, R.; van Creveld, C.; Lindner, P.; Gollop, N. A comparative study of milk serum proteins in camel (Camelus dromedarius) and bovine colostrum. Livest. Prod. Sci. 2001, 67, 297–301. [Google Scholar] [CrossRef]
- Shabo, Y.; Barzel, R.; Margoulis, M.; Yagil, R. Camel milk for food allergies in children. Isr. Med Assoc. J. 2005, 7, 796–798. [Google Scholar] [PubMed]
- El-Agamy, E.-S.I. Camel Milk. In Handbook of Milk of Non-Bovine Mammals; Park, Y.W., Haenlein, G.F.W., Wendorff, W.L., Eds.; John Wiley & Sons, Ltd.: Oxford, UK, 2017; pp. 409–480. ISBN 9781119110316. [Google Scholar]
- Rando, A.; Di Gregorio, P.; Ramunno, L.; Mariani, P.; Fiorella, A.; Senese, C.; Marletta, D.; Masina, P. Characterization of the CSN1AG Allele of the Bovine αs1-Casein Locus by the Insertion of a Relict of a Long Interspersed Element. J. Dairy Sci. 1998, 81, 1735–1742. [Google Scholar] [CrossRef]
- Cartegni, L.; Chew, S.L.; Krainer, A. Listening to silence and understanding nonsense: Exonic mutations that affect splicing. Nat. Rev. Genet. 2002, 3, 285–298. [Google Scholar] [CrossRef] [PubMed]
- Braunschweig, M.H.; Leeb, T. Aberrant Low Expression Level of Bovine β-Lactoglobulin Is Associated with a C to A Transversion in the BLG Promoter Region. J. Dairy Sci. 2006, 89, 4414–4419. [Google Scholar] [CrossRef]
- Brooke-Taylor, S.; Dwyer, K.; Woodford, K.; Kost, N. Systematic Review of the Gastrointestinal Effects of A1 Compared with A2 β-Casein. Adv. Nutr. 2017, 8, 739–748. [Google Scholar] [CrossRef]
- Gautier, P.; Escobar Stuardo, L. Rapport de la Mission de suivi de L’évaluation PVS des Services Vétérinaires du Bénin, OIE—World Organisation for Animal Health, Paris, France. 2013. Available online: https://rr-africa.oie.int/wp-content/uploads/2019/11/pvs_fu_rapport-benin.pdf (accessed on 20 June 2021).
- FAO. Phenotypic Characterization of Animal Genetic Resources; Food and Agriculture Organization of the United Nations: Rome, Italy, 2012. [Google Scholar]
- Vapnek, J.; Chapman, M. Legislative and Regulatory Options for Animal Welfare; Food and Agriculture Organization of the United Nations: Rome, Italy, 2011; ISBN 9251067112. [Google Scholar]



{kind=link}
{kind=link}
| Gene Name | Gene Sequence | |||
|---|---|---|---|---|
| Sequence ID | Location | Strand 1 | Exons (n) | |
| CSN1S1 | ENSBTAG00000007695 | 6:85410518:85429868 | + | 19 |
| CSN2 | ENSBTAG00000002632 | 6:85448564:85458344 | − | 9 |
| CSN1S2 | ENSBTAG00000005005 | 6:85529305:85549156 | + | 18 |
| CSN3 | ENSBTAG00000039787 | 6:85645254:85659526 | + | 5 |
| LALBA | ENSBTAG00000005859 | 5:31182832:31213745 | + | 4 |
| LGB | ENSBTAG00000014678 | 11:103255224:103264876 | + | 7 |
| CSN1S1 | CSN2 | CSN1S2 | CSN3 | LALBA | LGB | |
|---|---|---|---|---|---|---|
| n (%) | ||||||
| SNP | 20 (8.06) | 8 (5.83) | 26 (7.53) | 15 (5.23) | 3 (11.54) | 1 (2.22) |
| InDel 1 | 228 (91.94) | 114 (94.17) | 313 (92.47) | 275 (94.77) | 23 (88.46) | 44 (97.78) |
| Total | 248 | 120 | 332 | 287 | 26 | 45 |
| Genes | SNP | Gene Transcript | Variant Frequency | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| ID 1 | BTA:bp 2 | Allele | Exon | Protein 3 | Amino Acid 4 | Variant 5 | Crossbreed (n = 27) | Lagune (n = 20) | Somba (n = 20) | Total (n = 20) | |
| CSN1S1 | rs43703010 | 6:85427427 | A | 17 | 192 (207) | Glu | B | 0.62 | 0.97 | 0.93 | 0.81 |
| G | Gly | C | 0.38 | 0.03 | 0.08 | 0.19 | |||||
| CSN2 | rs715383373 | 6:85450908 | T | 7 | 197 (247) | Val | A1 | 0.97 | 1.00 | 1.00 | 0.99 |
| C | Ala | L | 0.03 | 0.00 | 0.00 | 0.01 | |||||
| rs43703011 | 6:85451298 | A | 7 | 67 (117) | His | A1 | 0.39 | 0.80 | 0.63 | 0.57 | |
| C | Pro | A2 | 0.61 | 0.20 | 0.37 | 0.43 | |||||
| CSN1S2 | rs441966828 | 6:85533780 | C | 3 | 8 (23) | Ser | A | 0.98 | 1.00 | 1.00 | 0.99 |
| T | Phe | B | 0.02 | 0.00 | 0.00 | 0.01 | |||||
| CSN3 | ss7626433428 | 6:85656526 | C | 4 | 66 (87) | Ala | B | 1.00 | 0.75 | 1.00 | 0.97 |
| T | Val | K | 0.00 | 0.25 | 0.00 | 0.03 | |||||
| rs43703015 | 6:85656736 | T | 4 | 136 (157) | Ile | B | 0.50 | 0.75 | 0.80 | 0.72 | |
| C | Thr | A | 0.50 | 0.25 | 0.20 | 0.28 | |||||
| rs43703016 | 6:85656772 | C | 4 | 148 (169) | Ala | B | 0.50 | 0.50 | 0.80 | 0.69 | |
| A | Asp | A | 0.50 | 0.50 | 0.20 | 0.31 | |||||
| LALBA | rs714688595 | 5:31184282 | C | 1 | 10 (58) | Arg | B | 0.96 | 1.00 | 0.88 | 0.94 |
| T | Trp | F | 0.04 | 0.00 | 0.13 | 0.06 | |||||
| rs722550244 | 5:31184283 | G | 1 | 10 (58) | Arg | B | 0.88 | 1.00 | 1.00 | 0.95 | |
| A | Gln | A | 0.12 | 0.00 | 0.00 | 0.05 | |||||
| rs465119286 | 5:31184696 | A | 2 | 41(89) | Ile | B | 0.88 | 1.00 | 0.95 | 0.93 | |
| G | Val | E | 0.12 | 0.00 | 0.05 | 0.07 | |||||
| LGB | rs109625649 | 11:103259232 | C | 4 | 118 (134) | Ala | B | 0.89 | 0.50 | 0.81 | 0.80 |
| T | Val | B1 | 0.11 | 0.50 | 0.19 | 0.20 | |||||
| ss7626433430 | 11:103257980 | - | 3 | Thr92 | - | B | 0.96 | 1.00 | 1.00 | 0.98 | |
| A | Thr92Asnfs*13 | - | K | 0.04 | 0.00 | 0.00 | 0.02 | ||||
| Haplotypes 1 | Crossbreed (n = 27) | Lagune (n = 20) | Somba (n = 20) | Total (n = 20) |
|---|---|---|---|---|
| B-A1-A-B | 0.01 | 0.50 | 0.47 | 0.39 |
| B-A2-A-B | 0.20 | 0.18 | 0.25 | 0.22 |
| B-A1-A-A | 0.39 | 0.29 | 0.20 | 0.20 |
| C-A2-A-A | <0.01 | <0.01 | 0.02 | 0.19 |
| C-A2-A-B | 0.38 | <0.01 | 0.02 | <0.01 |
| Gene | Location (BTA:bp 1) | Allele (ref/alt) 2 | ID 3 | Position | miRNA | TF | ||
|---|---|---|---|---|---|---|---|---|
| Loss 4 | Gain 4 | Loss 4 | Gain 4 | |||||
| CSN1S1 | 6:85411136 | C/T | rs379920406 | 5′-UTR | - | bta-miR-2291 | - | - |
| 6:85411147 | G/A | rs517257790 | 5′-UTR | - | - | - | - | |
| 6:85411192 | C/T | rs520777414 | 5′-UTR | - | - | - | - | |
| 6:85411221 | A/G | rs133040184 | 5′-UTR | bta-miR-133c | bta-miR-9-5p | - | - | |
| 6:85411236 | A/G | rs137119956 | 5′-UTR | bta-miR-2446 | - | E2F-6 | - | |
| 6:85411427 | A/C | rs135615686 | 5′-UTR | - | - | RAR | - | |
| 6:85411578 | A/G | rs109817504 | 5′-UTR | - | - | ATF-3; B-ATF; c-Jun; SMARCC1; SMARCC2 | - | |
| 6:85411618 | C/G | rs109757609 | 5′-UTR | bta-miR-2284e | bta-miR-7864 | - | Sp1; IRF-4 | |
| 6:85411677 | G/A | rs134534951 | 5′ UTR | - | - | STAT5A | - | |
| 6:85411780 | T/C | rs110163054 | 5′-UTR | bta-miR-15a; bta-miR-562 | - | - | - | |
| 6:85413195 | C/T | rs110899610 | 5′-UTR | bta-miR-2325b; bta-miR-2420; bta-miR-453 | bta-miR-376a | - | - | |
| 6:85428962 | G/A | rs466704456 | 3′-UTR | bta-miR-874 | bta-miR-219-5p; bta-miR-2355-3p | - | - | |
| 6:85429024 | C/T | ss7626432754 | 3′-UTR | - | - | - | - | |
| 6:85429077 | A/G | rs435231328 | 3′-UTR | bta-miR-338 | - | - | - | |
| 6:85429241 | A/G | rs211141048 | 3′-UTR | bta-miR-380-3p | - | SMAD4 | - | |
| 6:85429252 | C/A | rs716970086 | 3′-UTR | bta-miR-371 | - | SMAD4 | - | |
| CSN2 | 6:85449252 | C/A | rs440770944 | 3′-UTR | bta-miR-2464-3p | - | - | p63 |
| CSN1S2 | 6:85531897 | T/C | rs109900747 | 5′-UTR | - | bta-miR-452 | Sp1; HNF-4alpha; HNF-4gamma; FOXO1 | c-Ets-1 |
| 6:85548443 | A/G | rs211156498 | 3′-UTR | bta-miR-2318; bta-miR-2479; bta-miR-2480 | bta-miR-2463; bta-miR-452 | CP2-L1 | - | |
| 6:85548520 | A/G | rs109274107 | 3′-UTR | - | bta-miR-1388-3p | - | - | |
| CSN3 | 6:85656841 | T/A | rs109787476 | 3′-UTR | bta-miR-496 | bta-miR-2284w | ZNF274 | |
| 6:85658779 | C/A | ss7626433382 | 3′-UTR | bta-miR-193a-3p | - | - | - | |
| 6:85658873 | G/A | rs134516686 | 3′-UTR | - | - | - | - | |
| LALBA | 5:31183709 | G/C | rs444727593 | 5′-UTR | - | bta-miR-2475 | - | - |
| 5:31183736 | G/A | rs458466372 | 5′-UTR | bta-miR-2284r; bta-miR-2284s | - | - | - | |
| 5:31183766 | T/C | rs471361585 | 5′-UTR | - | bta-miR-2370-5p | - | - | |
| 5:31183789 | A/G | rs440012037 | 5′-UTR | bta-miR-7864 | - | - | ||
| 5:31183806 | C/T | rs717249686 | 5′-UTR | - | - | - | KLF4 | |
| 5:31183848 | C/A | rs460157851 | 5′-UTR | - | bta-miR-2472; bta-miR-6517 | - | AP-2alpha; IRF-9, STAT2;STAT1 | |
| 5:31183921 | A/G | rs448925171 | 5′-UTR | bta-miR-2325c; bta-miR-6120-3p | bta-miR-218; bta-miR-2452 | - | - | |
| 5:31183924 | T/C | rs719631407 | 5′-UTR | bta-miR-6120-3p | bta-miR-218 | - | - | |
| 5:31184082 | T/C | rs462561324 | 5′-UTR | - | - | - | - | |
| 5:31185981 | C/T | rs718675014 | 3′-UTR | - | bta-miR-2425-5p; bta-miR-2467-3p | N-Myc; N-Myc | c-Myc | |
| 5:31186027 | A/G | rs110359174 | 3′-UTR | bta-miR-143 | - | - | - | |
| LGB | 11:103255847 | G/A | rs516356159 | 5′-UTR | - | - | - | RCoR2 |
| 11:103255918 | C/T | rs41255682 | 5′-UTR | bta-miR-1777a; bta-miR-2454-5p | - | SIX5 | - | |
| 11:103255930 | G/C | rs41255683 | 5′-UTR | - | - | - | STAT1; STAT3 | |
| 11:103260824 | G/A | rs1116405113 | 3′-UTR | bta-miR-7860 | - | - | - | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vanvanhossou, S.F.U.; Giambra, I.J.; Yin, T.; Brügemann, K.; Dossa, L.H.; König, S. First DNA Sequencing in Beninese Indigenous Cattle Breeds Captures New Milk Protein Variants. Genes 2021, 12, 1702. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12111702
Vanvanhossou SFU, Giambra IJ, Yin T, Brügemann K, Dossa LH, König S. First DNA Sequencing in Beninese Indigenous Cattle Breeds Captures New Milk Protein Variants. Genes. 2021; 12(11):1702. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12111702
Chicago/Turabian StyleVanvanhossou, Sèyi Fridaïus Ulrich, Isabella Jasmin Giambra, Tong Yin, Kerstin Brügemann, Luc Hippolyte Dossa, and Sven König. 2021. "First DNA Sequencing in Beninese Indigenous Cattle Breeds Captures New Milk Protein Variants" Genes 12, no. 11: 1702. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12111702