
Expression Pattern of Nitric Oxide Synthase during Development of the Marine Gastropod Mollusc, Crepidula fornicata


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Care and Embryos Fixation
2.2. Sequence Mining and Analysis
2.3. Cloning and Sequencing
2.4. Gene Expression Analysis
2.5. Microscopy
3. Results and Discussion
3.1. One Ortholog for Nos with Neural Signatures Is Present in C. fornicata
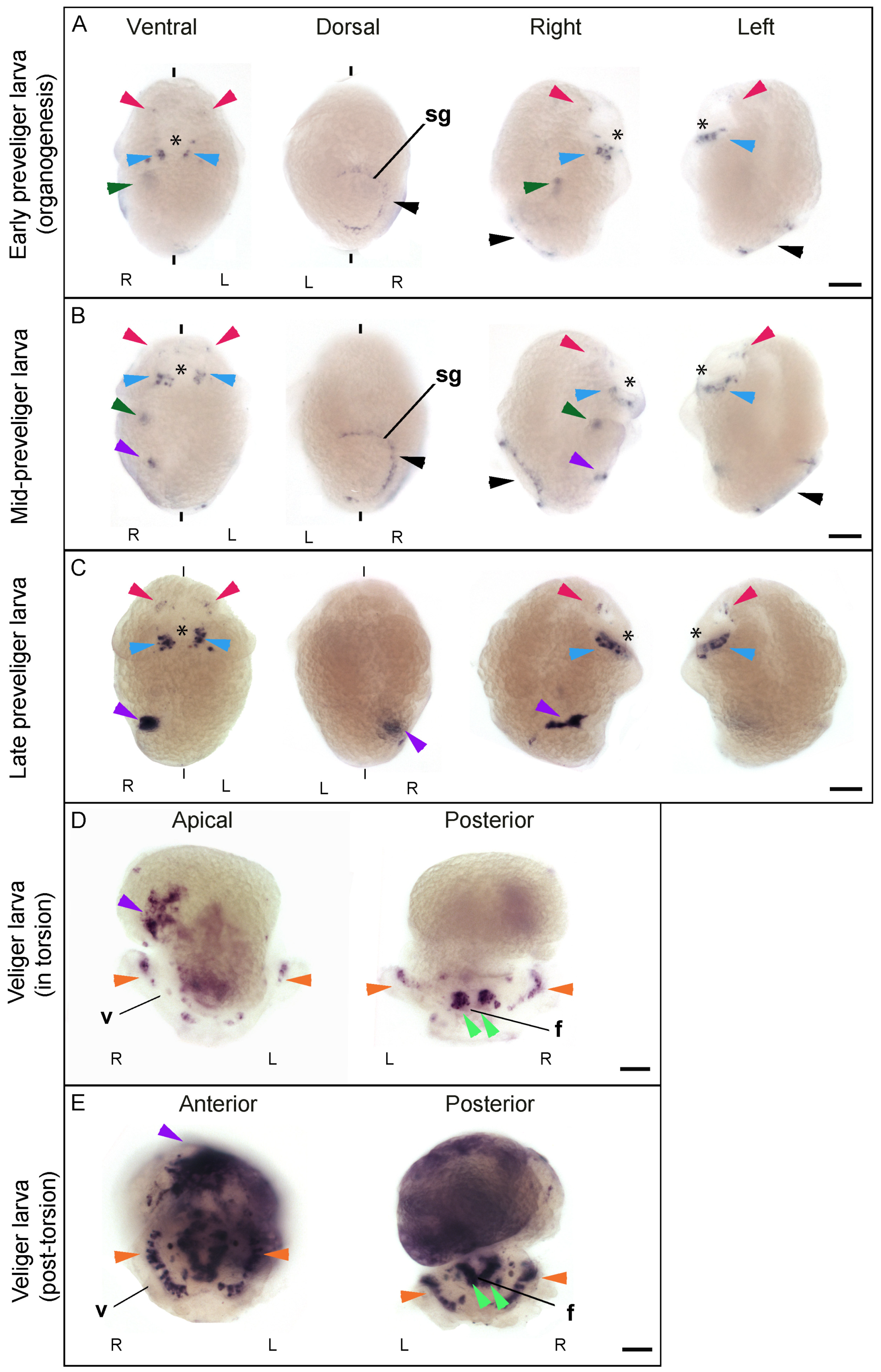
3.2. Cfo-Nos Expression during Development Suggests an Involvement in Shell Formation and Neural Specification
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Biggers, W.J.; Pires, A.; Pechenik, J.A.; Johns, E.; Patel, P.; Polson, T.; Polson, J. Inhibitors of nitric oxide synthase induce larval settlement and metamorphosis of the polychaete annelid Capitella teleta. Invertebr. Reprod. Dev. 2012, 56, 1–13. [Google Scholar] [CrossRef]
- Bishop, C.D.; Brandhorst, B.P. Development of Nitric Oxide Synthase-Defined Neurons in the Sea Urchin Larval Ciliary Band and Evidence for a Chemosensory Function During Metamorphosis. Dev. Dyn. 2007, 236, 1535–1546. [Google Scholar] [CrossRef]
- Comes, S.; Locascio, A.; Silvestre, F.; d’Ischia, M.; Russo, G.L.; Tosti, E.; Branno, M.; Palumbo, A. Regulatory roles of nitric oxide during larval development and metamorphosis in Ciona intestinalis. Dev. Biol. 2007, 306, 772–784. [Google Scholar] [CrossRef]
- Bishop, C.D.; Bates, W.R.; Brandhorst, B.P. Regulation of metamorphosis in ascidians involves NO/cGMP signaling and HSP90. J. Exp. Zool. 2001, 289, 374–384. [Google Scholar] [CrossRef]
- Ueda, N.; Richards, G.S.; Degnan, B.M.; Kranz, A.; Adamska, M.; Croll, R.P.; Degnan, S.M. An ancient role for nitric oxide in regulating the animal pelagobenthic life cycle: Evidence from a marine sponge. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef] [Green Version]
- Ueda, N.; Degnan, S.M. Nitric oxide acts as a positive regulator to induce metamorphosis of the ascidian Herdmania momus. PLoS ONE 2013, 8, e72797. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.T.; Zhang, Y.; Liu, Y.Z.; Li, Y.F.; Yoshida, A.; Osatomi, K.; Yang, J.L.; Liang, X. Nitric Oxide Negatively Regulates Larval Metamorphosis in Hard-Shelled Mussel (Mytilus coruscus). Front. Mar. Sci. 2020, 7. [Google Scholar] [CrossRef]
- Pechenik, J.; Cochrane, D.E.; Li, W.; West, E.T.; Pires, A.; Leppo, M. Nitric oxide inhibits metamorphosis in larvae of Crepidula fornicata, the slippershell snail. Biol. Bull. 2007, 213, 160–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Froggett, S.J.; Leise, E.M. Metamorphosis in the marine snail Ilyanassa obsoleta, yes or NO? Biol. Bull. 1999, 196, 57–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romero, M.R.; Phuong, M.A.; Bishop, C.D.; Krug, P.J. Nitric oxide signaling differentially affects habitat choice by two larval morphs of the sea slug Alderia willowi: Mechanistic insight into evolutionary transitions in dispersal strategies. J. Exp. Biol. 2012, 216, 1114–1125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogeler, S.; Carboni, S.; Li, X.; Nevejan, N.; Monaghan, S.J.; Ireland, J.H.; Joisse, A. Bivalves are NO Different: Nitric Oxide as Negative Regulator of Metamorphosis in the Pacific Oyster, Crassostrea gigas. BMC Dev. Biol. 2020, 20, 23. [Google Scholar] [CrossRef] [PubMed]
- Mattiello, T.; Fiore, G.; Brown, E.R.; d’Ischia, M.; Palumbo, A. Nitric oxide mediates the glutamate-dependent pathway for neurotransmission in Sepia officinalis chromatophore organs. J. Biol. Chem. 2010, 285, 24154–24163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elphick, M.R.; Kemenes, G.; Staras, K.; O’Shea, M. Behavioral role for nitric oxide in chemosensory activation of feeding in a mollusc. J. Neurosci. 1995, 15, 7653–7664. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Zhou, Z.; Wang, L.; Shi, X.; Wang, J.; Yue, F.; Yi, Q.; Yang, C.; Song, L. The immunomodulation of inducible nitric oxide in scallop Chlamys farreri. Fish. Shellfish Immunol. 2013, 34, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Zhou, Z.; Wang, L.; Wang, L.; Yue, F.; Wang, J.; Song, L. A scallop nitric oxide synthase (NOS) with structure similar to neuronal NOS and its involvement in the immune defense. PLoS ONE 2013, 8, e69158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gajbhiye, D.S.; Khandeparker, L. Involvement of inducible nitric oxide synthase (iNOS) in immune-functioning of Paphia malabarica (Chemnitz, 1782). Fish. Shellfish Immunol. 2019, 84, 384–389. [Google Scholar] [CrossRef]
- Miller, N.; Saada, R.; Fishman, S.; Hurwitz, I.; Susswein, A.J. Neurons controlling Aplysia feeding inhibit themselves by continuous NO production. PLoS ONE 2011, 6, e17779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakura, M.; Kabetani, M.; Watanabe, S.; Kirino, Y. Impairment of olfactory discrimination by blockade of nitric oxide activity in the terrestrial slug Limax valentianus. Neurosci. Lett. 2004, 370, 257–261. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, R.; Misawa, K.; Ito, E. Genomic structure of nitric oxide synthase in the terrestrial slug is highly conserved. Gene 2008, 415, 74–81. [Google Scholar] [CrossRef]
- Serfőző, Z.; Nacsa, K.; Veréb, Z.; Battonyai, I.; Hegedűs, C.; Balogh, C.; Elekes, K. Nitric oxide-coupled signaling in odor elicited molecular events in the olfactory center of the terrestrial snail, Helix pomatia. Cell Signal. 2017, 30, 67–81. [Google Scholar] [CrossRef] [Green Version]
- Toni, M.; De Angelis, F.; di Patti, M.C.; Cioni, C. Nitric Oxide Synthase in the Central Nervous System and Peripheral Organs of Stramonita haemastoma: Protein Distribution and Gene Expression in Response to Thermal Stress. Mar. Drugs 2015, 13, 6636–6664. [Google Scholar] [CrossRef] [Green Version]
- Andreakis, N.; D’Aniello, S.; Albalat, R.; Patti, F.P.; Garcia-Fernàndez, J.; Procaccini, G.; Sordino, P.; Palumbo, A. Evolution of the nitric oxide synthase family in metazoans. Mol. Biol. Evol. 2011, 28, 163–179. [Google Scholar] [CrossRef] [Green Version]
- Annona, G.; Caccavale, F.; Pascual-Anaya, J.; Kuratani, S.; De Luca, P.; Palumbo, A.; D’Aniello, S. Nitric Oxide regulates mouth development in amphioxus. Sci. Rep. 2017, 7, 8432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hadfield, M.G. Why and how marine invertebrate larvae metamorphose so fast. Semin. Cell Dev. Biol. 2000, 11, 437–443. [Google Scholar] [CrossRef]
- Henry, J.Q.; Perry, K.J.; Martindale, M.Q. Cell specification and the role of the polar lobe in the gastropod mollusc Crepidula fornicata. Dev. Biol. 2006, 297, 295–307. [Google Scholar] [CrossRef] [Green Version]
- Perry, K.J.; Lyons, D.C.; Truchado-Garcia, M.; Fischer, A.H.; Helfrich, L.W.; Johansson, K.B.; Diamond, J.C.; Grande, C.; Henry, J.Q. Deployment of regulatory genes during gastrulation and germ layer specification in a model spiralian mollusc Crepidula. Dev. Dyn. 2015, 244, 1215–1248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kenny, N.J.; Truchado-García, M.; Grande, C. Deep, multi-stage transcriptome of the schistosomiasis vector Biomphalaria glabrata provides platform for understanding molluscan disease-related pathways. BMC Infect. Dis. 2016, 16, 618. [Google Scholar] [CrossRef] [Green Version]
- Kearse, K.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Olson, S.A. MacVector: An integrated sequence analysis program for the Macintosh. Methods Mol. Biol. 1994, 25, 195–201. [Google Scholar] [CrossRef]
- Lee, H.J.; Zheng, J.J. PDZ domains and their binding partners: Structure, specificity, and modification. Cell Commun. Signal. 2010, 8, 8. [Google Scholar] [CrossRef] [Green Version]
- Kone, B.C.; Kuncewicz, T.; Zhang, W.; Yu, Z.Y. Protein interactions with nitric oxide synthases: Controlling the right time, the right place, and the right amount of nitric oxide. Am. J. Physiol. Renal Physiol. 2003, 285, F178–F190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishida, C.R.; Ortiz De Montellano, P.R. Autoinhibition of endothelial nitric-oxide synthase. Identification of an electron transfer control element. J. Biol. Chem. 1999, 274, 14692–14698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dickinson, J.G.; Nason, J.; Croll, R.P. Histochemical localization of FMRFamide, serotonin and catecholamines in embryonic Crepidula fornicata (Gastropoda, Prosobranchia). Zoomorphology 1999, 119, 49–62. [Google Scholar] [CrossRef]
- Nielsen, C. Larval nervous systems: True larval and precocious adult. J. Exp. Biol. 2015, 218, 629–636. [Google Scholar] [CrossRef] [Green Version]
- Grande, C.; Patel, N.H. Nodal signalling is involved in left-right asymmetry in snails. Nature 2009, 457, 1007–1011. [Google Scholar] [CrossRef] [Green Version]
- Kurita, Y.; Wada, H. Evidence that gastropod torsion is driven by asymmetric cell proliferation activated by TGF-β signalling. Biol. Lett. 2011, 7, 759–762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashimoto, N.; Kurita, Y.; Wada, H. Developmental role of dpp in the gastropod shell plate and co-option of the dpp signaling pathway in the evolution of the operculum. Dev. Biol. 2012, 366, 367–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henry, J.J.; Collin, R.; Perry, K.J. The slipper snail, Crepidula: An emerging lophotrochozoan model system. Biol. Bull. 2010, 218, 211–229. [Google Scholar] [CrossRef] [Green Version]
- Sinigaglia, C.; Busengdal, H.; Lerner, A.; Oliveri, P.; Rentzsch, F. Molecular characterization of the apical organ of the anthozoan Nematostella vectensis. Dev. Biol. 2015, 398, 120–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Ilyanassa obsoleta | Metamorphosis (negative regulator) | [9] |
| Crepidula fornicata | Metamorphosis (negative regulator) | [8] |
| Alderia willowi | Metamorphosis (negative regulator) | [10] |
| Mytilus coruscus | Metamorphosis (negative regulator) | [7] |
| Crassostera gigas | Metamorphosis (negative regulator), Immune Response | [11] |
| Chlamys farreri | Immune Response | [14,15] |
| Paphia malabarica | Immune Response | [16] |
| Sepia oficinalis | Development, Neurotransmission | [12] |
| Lymnaea stagnalis | Neurotransmission, Chemosensory activation | [13] |
| Aplysia californica | Feeding, Chemosensory activation | [17] |
| Lehmannia valentiana | Olfaction | [18] |
| Limax valentianus | Olfaction | [19] |
| Helix pomatia | Olfaction | [20] |
| Stramonita haemastoma | Sensory Perception, Stress Response | [21] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Truchado-Garcia, M.; Caccavale, F.; Grande, C.; D’Aniello, S. Expression Pattern of Nitric Oxide Synthase during Development of the Marine Gastropod Mollusc, Crepidula fornicata. Genes 2021, 12, 314. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12020314
Truchado-Garcia M, Caccavale F, Grande C, D’Aniello S. Expression Pattern of Nitric Oxide Synthase during Development of the Marine Gastropod Mollusc, Crepidula fornicata. Genes. 2021; 12(2):314. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12020314
Chicago/Turabian StyleTruchado-Garcia, Marta, Filomena Caccavale, Cristina Grande, and Salvatore D’Aniello. 2021. "Expression Pattern of Nitric Oxide Synthase during Development of the Marine Gastropod Mollusc, Crepidula fornicata" Genes 12, no. 2: 314. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12020314