
Molecular Profiling and Gene Banking of Rabbit EPCs Derived from Two Biological Sources

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Experimental Design
2.3. Collection of Biological Material and Isolation of Rabbit Endothelial Progenitor Cells
2.4. Culture of Human Endothelial Cell Line
2.5. Flow Cytometry
2.6. Polymerase Chain Reaction (PCR) Analyses
2.6.1. Real-Time PCR
2.6.2. Droplet Digital PCR
2.7. Cell Differentiation into the Neurogenic Lineage
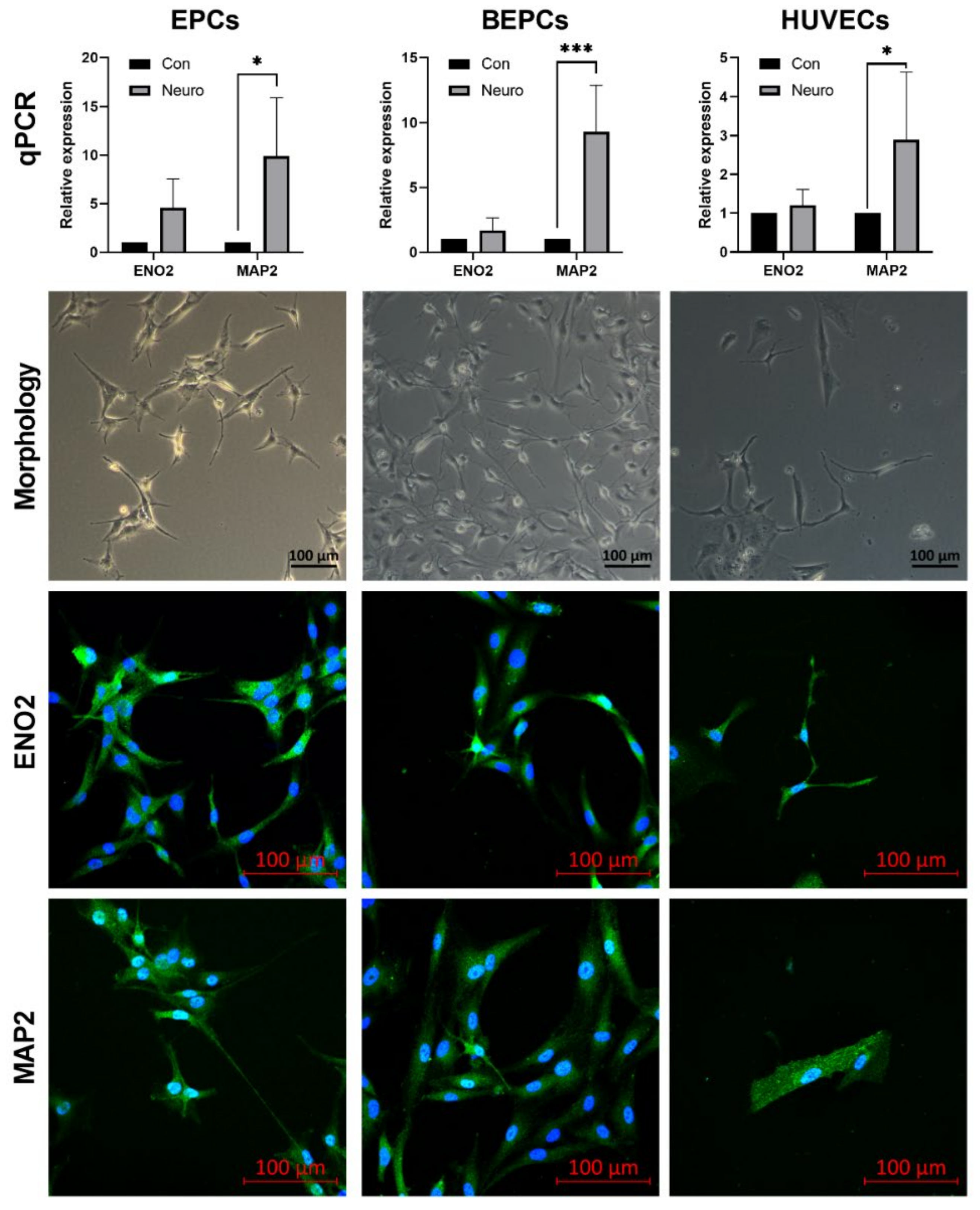
2.7.1. Quantitative Real–Time PCR (qPCR) Analysis of Neuronal Markers
2.7.2. Expression of the Neuronal Markers Assessed by Confocal Microscopy
2.8. Cryopreservation of the Rabbit Endothelial Progenitor Cells (EPCs)
2.9. Statistical Analysis
3. Results
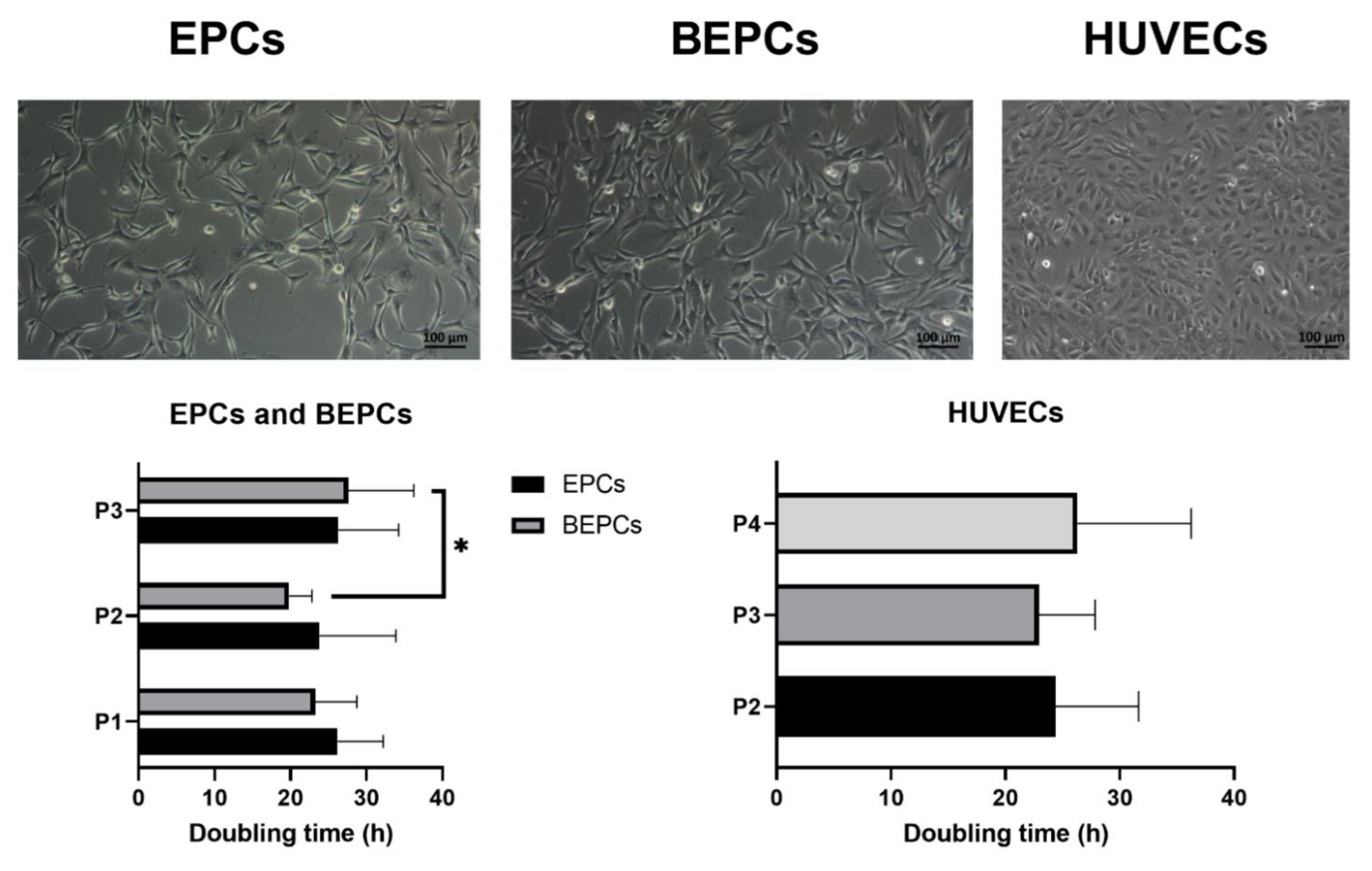
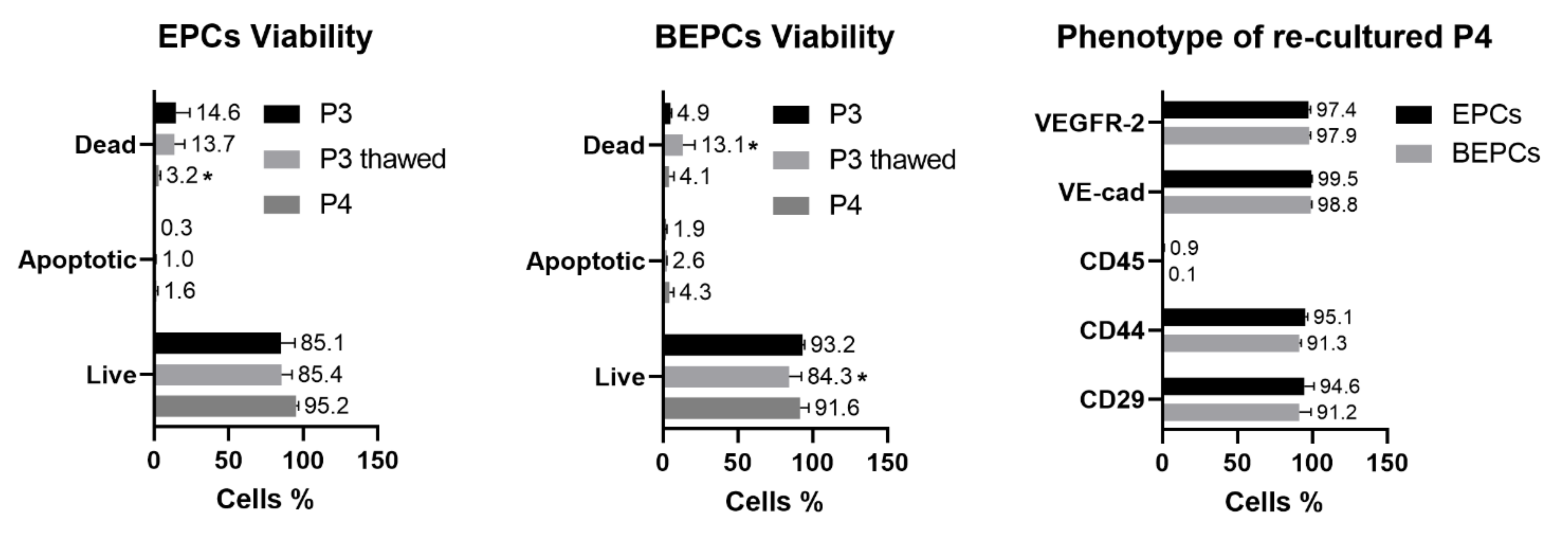
3.1. Cell Morphology, Proliferation and Viability
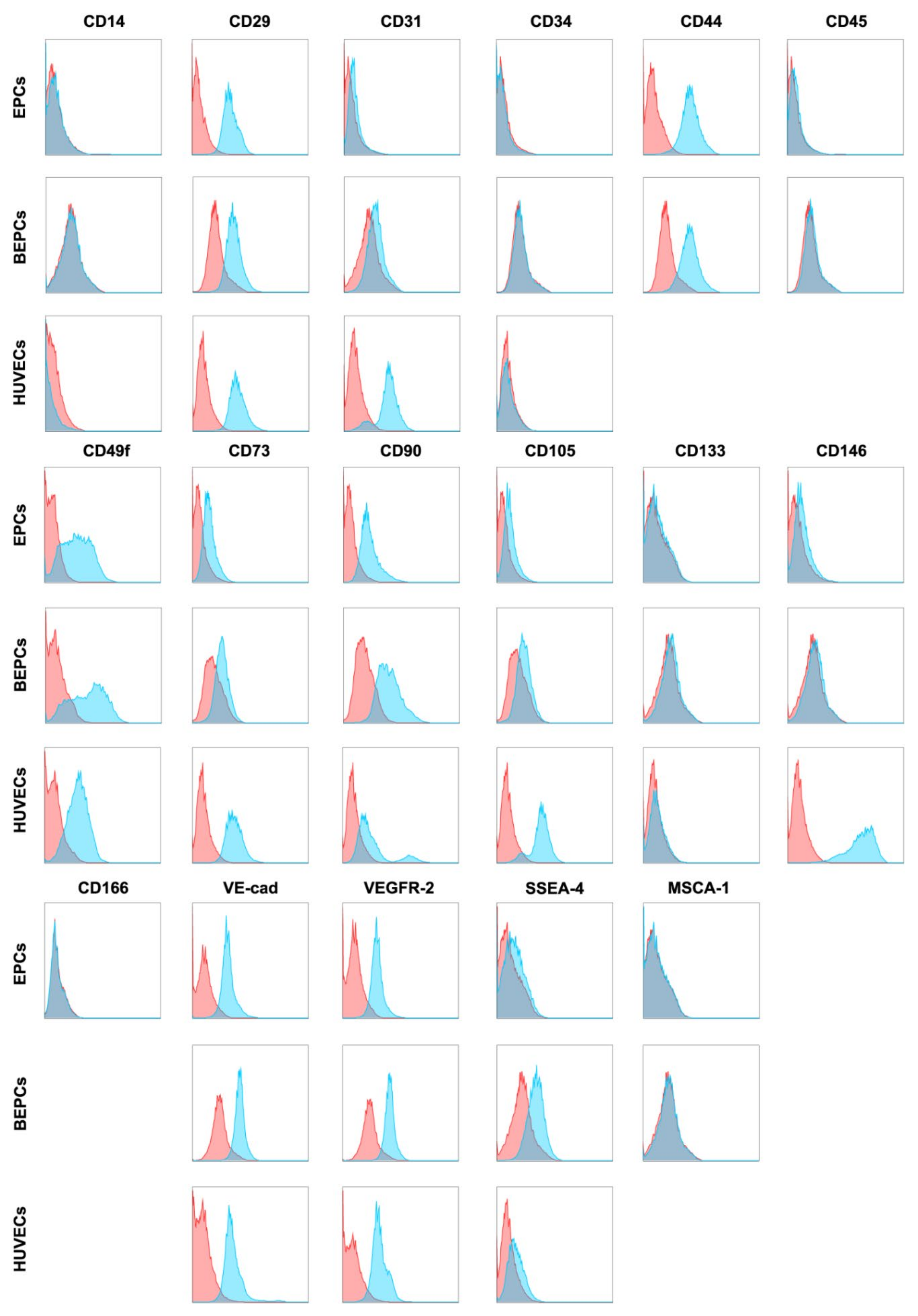
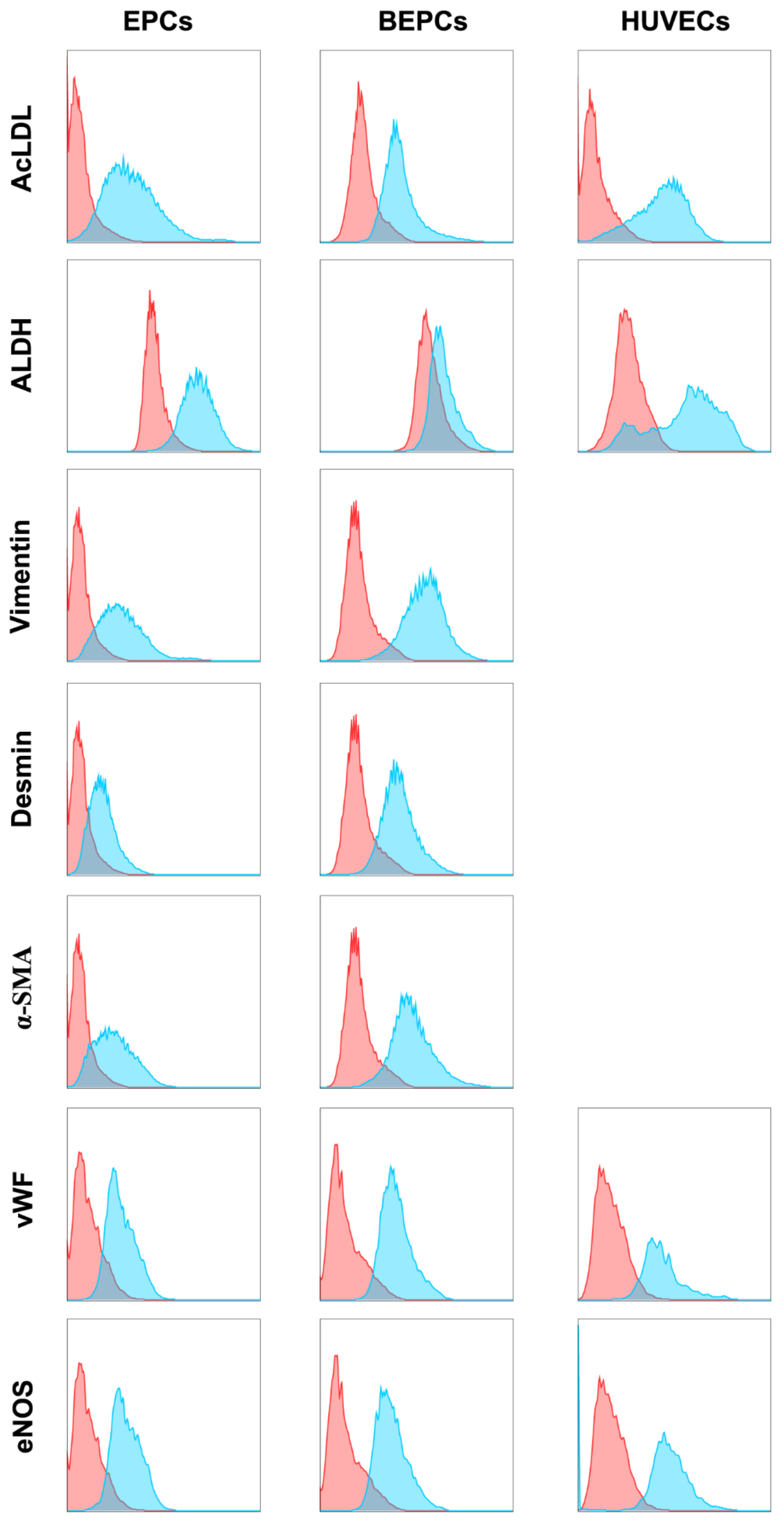
3.2. Phenotypic Analyses of the Primary Endothelial Cells
3.2.1. Immunophenotyping
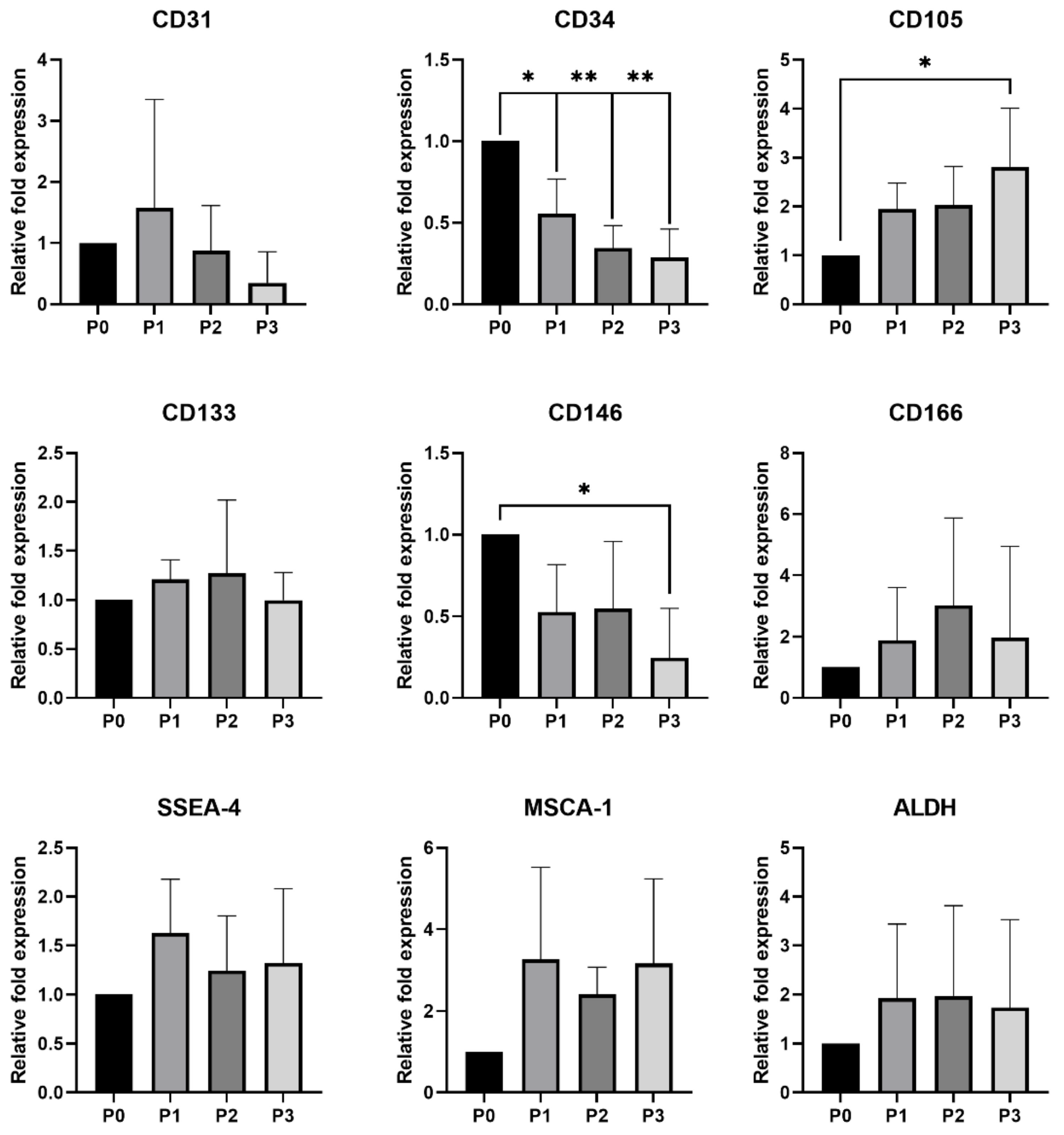
3.2.2. qPCR Analysis
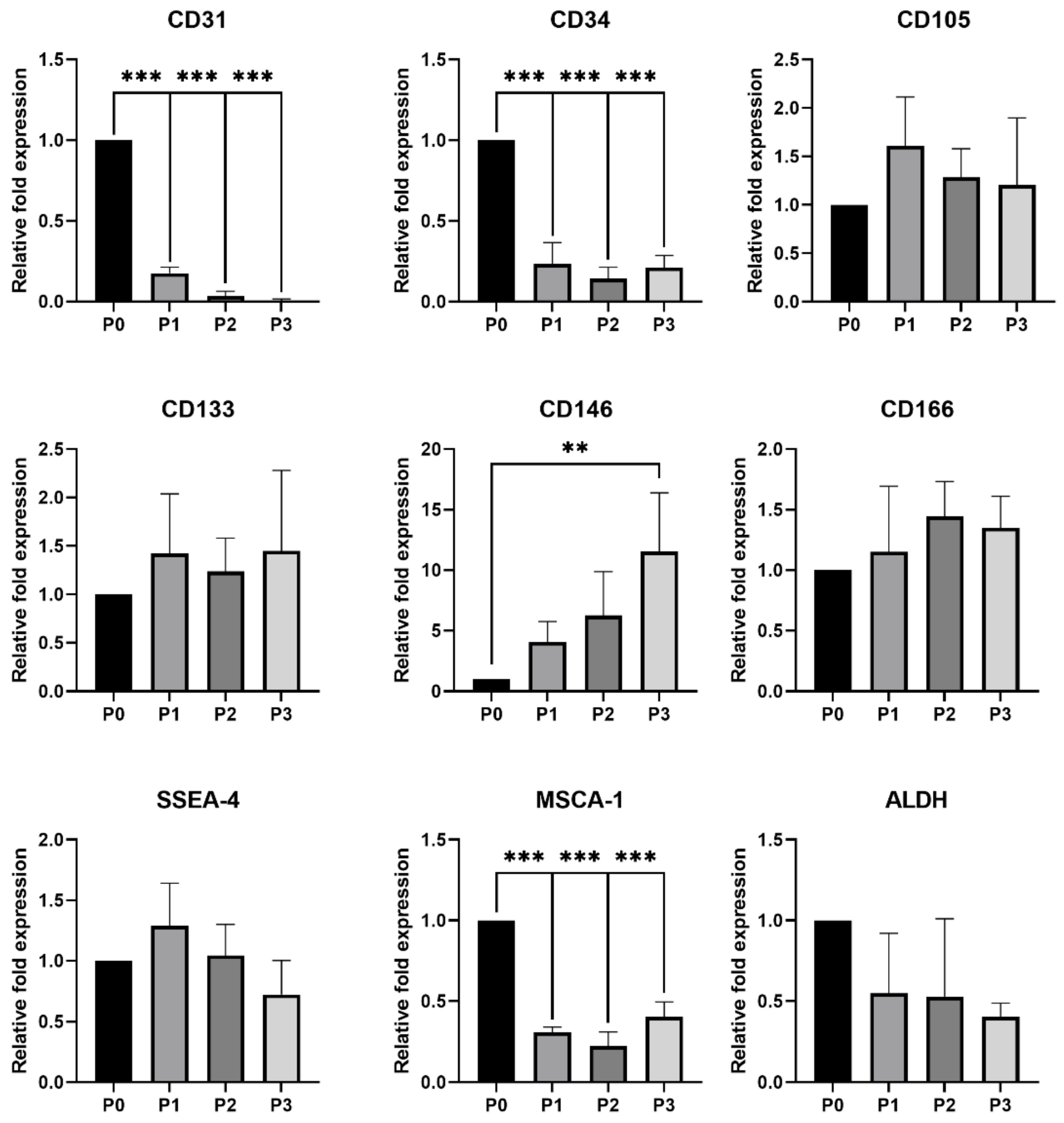
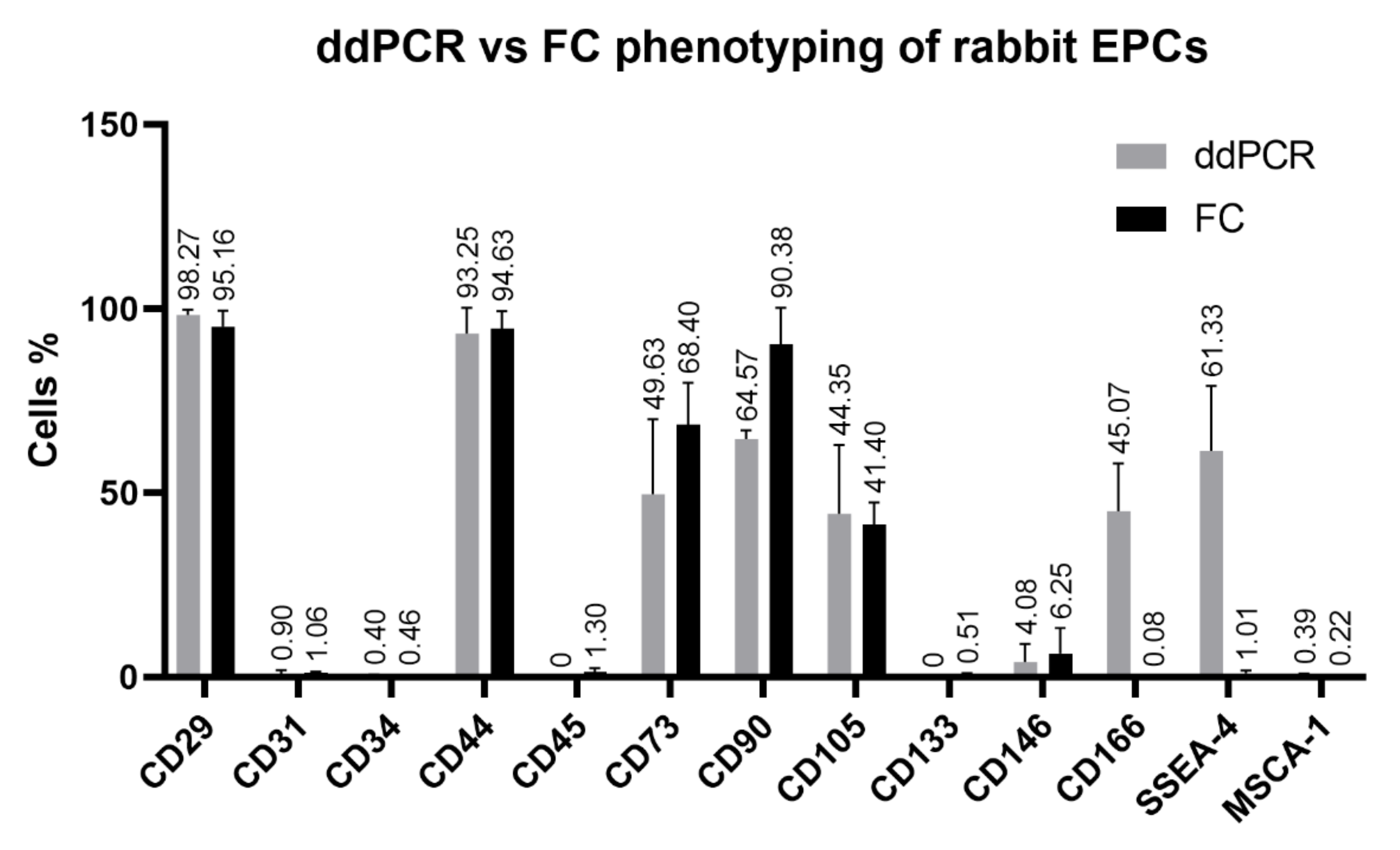
3.2.3. Digital Droplet PCR (ddPCR) Analysis
3.3. Neurogenic Differentiation of the Endothelial Progenitor Cells
3.4. Cryopreservation of the Rabbit EPCs and BEPCs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A


Appendix B

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Marker | Species | Homology (%) | NCBI Reference |
|---|---|---|---|
| CD14 | Rabbit | 73.07 | NP_001075664.1 |
| Human | 100 | NP_000582.1 | |
| CD29 (ITB1) | Rabbit | 100 | AAB34684.1 |
| Human | 100 | AAA79835.1 | |
| CD31 (PECAM1) | Rabbit | 73.99 | XP_008269937.1 |
| Human | 100 | NP_000433.4 | |
| CD34 | Rabbit | 71.76 | XP_008266693.1 |
| Human | 100 | NP_001020280.1 | |
| CD49f (ITGA6) | Rabbit | 92.74 | XP_017198420.1 |
| Human | 100 | NP_001073286.1 | |
| CD73 (NT5E) | Rabbit | 89.39 | XP_002714603.1 |
| Human | 100 | NP_002517.1 | |
| CD90 (THY1) | Rabbit | 82.23 | XP_002722764.1 |
| Human | 100 | NP_006279.2 | |
| CD105 (ENG) | Rabbit | 72.84 | XP_008249251.1 |
| Human | 100 | NP_001108225.1 | |
| CD133 (PROM1) | Rabbit | 68.31 | XP_017205932.1 |
| Human | 100 | NP_006008.1 | |
| CD146 (MCAM) | Rabbit | 82.44 | XP_008248732.2 |
| Human | 100 | NP_006491.2 | |
| CD166 (ALCAM) | Rabbit | 95.20 | XP_002716771.1 |
| Human | 100 | NP_001618.2 | |
| VE-cadherin (CDH5) | Rabbit | 83.72 | XP_008255604.2 |
| Human | 100 | NP_001786.2 | |
| VEGFR-2 (KDR) | Rabbit | 87.54 | NP_001182599.1 |
| Human | 100 | NP_002244.1 | |
| SSEA-4 (ST3GAL2) | Rabbit | 98.29 | XP_002711761.1 |
| Human | 100 | NP_008858.1 | |
| MSCA-1 (ALPL) | Rabbit | 91.93 | XP_017201978.1 |
| Human | 100 | NP_000469.3 | |
| Vimentin | Rabbit | 96.78 | XP_002717466.1 |
| Human | 100 | NP_003371.2 | |
| Desmin | Rabbit | 98.51 | NP_001164952.1 |
| Human | 100 | NP_001918.3 | |
| α-SMA (ACTA2) | Rabbit | 100 | NP_001095152.1 |
| Human | 100 | NP_001604.1 | |
| vWF | Rabbit | 83.02 | NP_001316017.1 |
| Human | 100 | NP_000543.3 | |
| eNOS (NOS3) | Rabbit | 99.15 | AAG24287.1 |
| Human | 100 | NP_000594.2 |
References
- Harris, D.T. Stem Cell Banking for Regenerative and Personalized Medicine. Biomedicines 2014, 2, 50–79. [Google Scholar] [CrossRef] [Green Version]
- Vašíček, J.; Kováč, M.; Baláži, A.; Kulíková, B.; Tomková, M.; Olexíková, L.; Čurlej, J.; Bauer, M.; Schnabl, S.; Hilgarth, M.; et al. Combined approach for characterization and quality assessment of rabbit bone marrow-derived mesenchymal stem cells intended for gene banking. New Biotechnol. 2020, 54, 1–12. [Google Scholar] [CrossRef]
- Chopra, H.; Hung, M.K.; Kwong, D.L.; Zhang, C.F.; Pow, E.H.N. Insights into endothelial progenitor cells: Origin, classification, potentials, and prospects. Stem Cells Int. 2018, 2018, 7015. [Google Scholar] [CrossRef] [PubMed]
- Yoder, M.C. Endothelial stem and progenitor cells (stem cells): (2017 Grover Conference Series). Pulm. Circ. 2017, 8, 9. [Google Scholar] [CrossRef] [Green Version]
- Asahara, T.; Murohara, T.; Sullivan, A.; Silver, M.; Van Der Zee, R.; Li, T.; Witzenbichler, B.; Schatteman, G.; Isner, J.M. Isolation of putative progenitor endothelial cells for angiogenesis. Science 1997, 275, 964–966. [Google Scholar] [CrossRef]
- Hur, J.; Yoon, C.-H.; Kim, H.-S.; Choi, J.-H.; Kang, H.-J.; Hwang, K.-K.; Oh, B.-H.; Lee, M.-M.; Park, Y.-B. Characterization of Two Types of Endothelial Progenitor Cells and Their Different Contributions to Neovasculogenesis. Arter. Thromb. Vasc. Biol. 2004, 24, 288–293. [Google Scholar] [CrossRef] [PubMed]
- Rehman, J.; Li, J.; Orschell, C.M.; March, K.L. Peripheral Blood “Endothelial Progenitor Cells” Are Derived From Monocyte/Macrophages and Secrete Angiogenic Growth Factors. Circ. 2003, 107, 1164–1169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.; Weisdorf, D.J.; Solovey, A.; Hebbel, R.P. Origins of circulating endothelial cells and endothelial outgrowth from blood. J. Clin. Investig. 2000, 105, 71–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ingram, D.A.; Mead, L.E.; Tanaka, H.; Meade, V.; Fenoglio, A.; Mortell, K.; Pollok, K.; Ferkowicz, M.J.; Gilley, D.; Yoder, M.C. Identification of a novel hierarchy of endothelial progenitor cells using human peripheral and umbilical cord blood. Blood 2004, 104, 2752–2760. [Google Scholar] [CrossRef]
- Masek, L.C.; Sweetenham, J.W. Isolation and culture of endothelial cells from human bone marrow. Br. J. Haematol. 1994, 88, 855–865. [Google Scholar] [CrossRef] [PubMed]
- Rafii, S.; Shapiro, F.; Rimarachin, J.; Nachman, R.L.; Ferris, B.; Weksler, B.; Moore, M.A.; Asch, A.S. Isolation and characterization of human bone marrow microvascular endothelial cells: Hematopoietic progenitor cell adhesion. Blood 1994, 84, 10–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Timmermans, F.; Van Hauwermeiren, F.; De Smedt, M.; Raedt, R.; Plasschaert, F.; De Buyzere, M.L.; Gillebert, T.C.; Plum, J.; Vandekerckhove, B. Endothelial outgrowth cells are not derived from CD133+ cells or CD45+ hematopoietic precursors. Arter. Thromb. Vasc. Biol. 2007, 27, 1572–1579. [Google Scholar] [CrossRef] [Green Version]
- Peichev, M.; Naiyer, A.J.; Pereira, D.; Zhu, Z.; Lane, W.J.; Williams, M.; Oz, M.C.; Hicklin, D.J.; Witte, L.; Moore, M.A.S.; et al. Expression of VEGFR-2 and AC133 by circulating human CD34+ cells identifies a population of functional endothelial precursors. Blood 2000, 95, 952–958. [Google Scholar] [CrossRef]
- Medina, R.J.; O’Neill, C.L.; Sweeney, M.; Guduric-Fuchs, J.; Gardiner, T.A.; Simpson, D.A.; Stitt, A.W. Molecular analysis of endothelial progenitor cell (EPC) subtypes reveals two distinct cell populations with different identities. BMC Med Genom. 2010, 3, 18. [Google Scholar] [CrossRef] [Green Version]
- Gulati, R.; Jevremovic, D.; Peterson, T.E.; Chatterjee, S.; Shah, V.; Vile, R.G.; Simari, R.D. Diverse Origin and Function of Cells With Endothelial Phenotype Obtained From Adult Human Blood. Circ. Res. 2003, 93, 1023–1025. [Google Scholar] [CrossRef] [PubMed]
- Mukai, N.; Akahori, T.; Komaki, M.; Li, Q.; Kanayasu-Toyoda, T.; Ishii-Watabe, A.; Kobayashi, A.; Yamaguchi, T.; Abe, M.; Amagasa, T.; et al. A comparison of the tube forming potentials of early and late endothelial progenitor cells. Exp. Cell Res. 2008, 314, 430–440. [Google Scholar] [CrossRef]
- Ito, H.; Rovira, I.I.; Bloom, M.L.; Takeda, K.; Ferrans, V.J.; Quyyumi, A.A.; Finkel, T. Endothelial progenitor cells as putative targets for angiostatin. Cancer Res. 1999, 59, 5875–5877. [Google Scholar] [PubMed]
- Hill, J.M.; Zalos, G.; Halcox, J.P.J.; Schenke, W.H.; Waclawiw, M.A.; Quyyumi, A.A.; Finkel, T. Circulating endothelial progenitor cells, vascular function, and cardiovascular risk. N. Engl. J. Med. 2003, 348, 593–600. [Google Scholar] [CrossRef]
- Richardson, M.R.; Yoder, M.C. Endothelial progenitor cells: Quo Vadis? J. Mol. Cell. Cardiol. 2011, 50, 266–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, Q.; Rafii, S.; Wu, M.H.-D.; Wijelath, E.S.; Yu, C.; Ishida, A.; Fujita, Y.; Kothari, S.; Mohle, R.; Sauvage, L.R.; et al. Evidence for Circulating Bone Marrow-Derived Endothelial Cells. Blood 1998, 92, 362–367. [Google Scholar] [CrossRef] [PubMed]
- Gehling, U.M.; Ergun, S.; Schumacher, U.; Wagener, C.; Pantel, K.; Otte, M.; Schuch, G.; Schafhausen, P.; Mende, T.; Kilic, N.; et al. In vitro differentiation of endothelial cells from AC133-positive progenitor cells. Blood 2000, 95, 3106–3112. [Google Scholar] [CrossRef] [PubMed]
- Kaushal, S.; Amiel, G.E.; Guleserian, K.J.; Shapira, O.M.; Perry, T.E.; Sutherland, F.W.; Rabkin, E.; Moran, A.M.; Schoen, F.J.; Atala, A.; et al. Functional small-diameter neovessels created using endothelial progenitor cells expanded ex vivo. Nat. Med. 2001, 7, 1035–1040. [Google Scholar] [CrossRef]
- Kanayasu-Toyoda, T.; Yamaguchi, T.; Oshizawa, T.; Hayakawa, T. CD31 (PECAM-1)-bright cells derived from AC133-positive cells in human peripheral blood as endothelial-precursor cells. J. Cell. Physiol. 2003, 195, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Wijelath, E.S.; Rahman, S.; Murray, J.; Patel, Y.; Savidge, G.; Sobel, M. Fibronectin promotes VEGF-induced CD34+ cell differentiation into endothelial cells. J. Vasc. Surg. 2004, 39, 655–660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peters, E.B. Endothelial Progenitor Cells for the Vascularization of Engineered Tissues. Tissue Eng. Part B Rev. 2018, 24, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Kutikhin, A.G.; Tupikin, A.E.; Matveeva, V.G.; Shishkova, D.K.; Antonova, L.V.; Kabilov, M.R.; Velikanova, E.A. Human peripheral blood-derived endothelial colony-forming cells are highly similar to mature vascular endothelial cells yet demonstrate a transitional transcriptomic signature. Cells 2020, 9, 876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, N.; Du, C.-Q.; Wang, S.-S.; Xie, K.; Zhang, S.-L.; Miao, J.-Y. D609 induces vascular endothelial cells and marrow stromal cells differentiation into neuron-like cells. Acta Pharmacol. Sin. 2004, 25, 442–446. [Google Scholar] [PubMed]
- Su, L.; Zhao, J.; Zhao, B.X.; Miao, J.Y.; Yin, D.L.; Zhang, S.L. Safrole oxide induced human umbilical vein vascular endothelial cell differentiation into neuron-like cells by depressing the reactive oxygen species level at the low concentration. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2006, 1763, 247–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, L.; Zhao, J.; Zhao, B.X.; Zhang, S.L.; Miao, J.Y. Safrole oxide induces human umbilical vein endothelial cell transdifferentiation to 5-hydroxytryptaminergic neuron-like cells through tropomyosin receptor kinase A/cyclooxygenase 2/nuclear factor-kappa B/interleukin 8 signaling. Int. J. Biochem. Cell Biol. 2011, 43, 1512–1522. [Google Scholar] [CrossRef]
- Wang, X.; Zhao, Z.; Zhang, H.; Hou, J.; Feng, W.; Zhang, M.; Guo, J.; Xia, J.; Ge, Q.; Chen, X.; et al. Simultaneous isolation of mesenchymal stem cells and endothelial progenitor cells derived from murine bone marrow. Exp. Ther. Med. 2018, 16, 5171–5177. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Cui, X.; Cheng, L.; Guan, X.; Li, H.; Li, X.; Cheng, M. Actin Stabilization by Jasplakinolide Affects the Function of Bone Marrow-Derived Late Endothelial Progenitor Cells. PLoS ONE 2012, 7, e50899. [Google Scholar] [CrossRef] [Green Version]
- Qiu, J.; Li, W.; Feng, S.; Wang, M.; He, Z. Transplantation of bone marrow-derived endothelial progenitor cells attenuates cerebral ischemia and reperfusion injury by inhibiting neuronal apoptosis, oxidative stress and nuclear factor-κB expression. Int. J. Mol. Med. 2013, 31, 91–98. [Google Scholar] [CrossRef] [Green Version]
- Guan, X.M.; Cheng, M.; Li, H.; Cui, X.D.; Li, X.; Wang, Y.L.; Sun, J.L.; Zhang, X.Y. Biological properties of bone marrow-derived early and late endothelial progenitor cells in different culture media. Mol. Med. Rep. 2013, 8, 1722–1728. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Gong, Y.; Lian, J.; Wang, L.; Kretlow, J.D.; Zhou, G.; Cao, Y.; Liu, W.; Zhang, W.J. Expansion of Endothelial Progenitor Cells in High Density Dot Culture of Rat Bone Marrow Cells. PLoS ONE 2014, 9, e107127. [Google Scholar] [CrossRef]
- Gong, X.; Li, B.; Yang, Y.; Huang, Y.; Sun, Y.; Liu, M.; Jia, X.; Fan, Y. Bone marrow derived endothelial progenitor cells retain their phenotype and functions after a limited number of culture passages and cryopreservation. Cytotechnology 2019, 71, 1–14. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Shirota, T.; Yasui, H.; Matsuda, T. Canine endothelial progenitor cell-lined hybrid vascular graft with nonthrombogenic potential. J. Thorac. Cardiovasc. Surg. 2003, 126, 455–464. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Riha, G.M.; Yang, H.; Li, M.; Yao, Q.; Chen, C. Differentiation and Proliferation of Endothelial Progenitor Cells from Canine Peripheral Blood Mononuclear Cells. J. Surg. Res. 2005, 126, 193–198. [Google Scholar] [CrossRef] [PubMed]
- Fedorovich, N.E.; Haverslag, R.T.; Dhert, W.J.; Alblas, J. The Role of Endothelial Progenitor Cells in Prevascularized Bone Tissue Engineering: Development of Heterogeneous Constructs. Tissue Eng. Part A 2010, 16, 2355–2367. [Google Scholar] [CrossRef] [PubMed]
- Bai, C.; Hou, L.; Zhang, M.; Pu, Y.; Guan, W.; Ma, Y. Characterization of vascular endothelial progenitor cells from chicken bone marrow. BMC Vet.-Res. 2012, 8, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, T.; Kalka, C.; Masuda, H.; Chen, D.; Silver, M.; Kearney, M.; Magner, M.; Isner, J.M.; Asahara, T. Ischemia- and cytokine-induced mobilization of bone marrow-derived endothelial progenitor cells for neovascularization. Nat. Med. 1999, 5, 434–438. [Google Scholar] [CrossRef]
- Gao, X.; Chen, W.; Liang, Z.; Chen, L. Autotransplantation of circulating endothelial progenitor cells protects against lipopolysaccharide-induced acute lung injury in rabbit. Int. Immunopharmacol. 2011, 11, 1584–1590. [Google Scholar] [CrossRef] [PubMed]
- Amini, A.R.; Laurencin, C.T.; Nukavarapu, S.P. Differential analysis of peripheral blood- and bone marrow-derived endothelial progenitor cells for enhanced vascularization in bone tissue engineering. J. Orthop. Res. 2012, 30, 1507–1515. [Google Scholar] [CrossRef]
- He, T.; Smith, L.A.; Harrington, S.; Nath, K.A.; Caplice, N.M.; Katusic, Z.S. Transplantation of Circulating Endothelial Progenitor Cells Restores Endothelial Function of Denuded Rabbit Carotid Arteries. Stroke 2004, 35, 2378–2384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Z.-L.; Mai, X.-L.; Sun, J.-H.; Ju, S.-H.; Yang, X.; Ni, Y.; Teng, G.-J. Inhibited atherosclerotic plaque formation by local administration of magnetically labeled endothelial progenitor cells (EPCs) in a rabbit model. Atherosclerosis 2009, 205, 80–86. [Google Scholar] [CrossRef]
- Kovac, M.; Vasicek, J.; Kulikova, B.; Bauer, M.; Curlej, J.; Balazi, A.; Chrenek, P. Different RNA and protein expression of surface markers in rabbit amniotic fluid-derived mesenchymal stem cells. Biotechnol. Prog. 2017, 33, 1601–1613. [Google Scholar] [CrossRef] [PubMed]
- Vašíček, J.; Shehata, M.; Schnabl, S.; Hilgarth, M.; Hubmann, R.; Jäger, U.; Bauer, M.; Chrenek, P. Critical assessment of the efficiency of CD34 and CD133 antibodies for enrichment of rabbit hematopoietic stem cells. Biotechnol. Prog. 2018, 34, 1278–1289. [Google Scholar] [CrossRef]
- Kulikova, B.; Kovac, M.; Bauer, M.; Tomkova, M.; Olexikova, L.; Vasicek, J.; Balazi, A.; Makarevich, A.V.; Chrenek, P. Survivability of rabbit amniotic fluid-derived mesenchymal stem cells post slow-freezing or vitrification. Acta Histochem. 2019, 121, 491–499. [Google Scholar] [CrossRef] [PubMed]
- Svoradová, A.; Vašíček, J.; Ostró, A.; Chrenek, P. Aldehyde dehydrogenase in fresh primordial germ cells as a marker of cell ‘stemness’. Zygote 2019, 27, 46–48. [Google Scholar] [CrossRef]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T.L. Primer-BLAST: A tool to design target-specific primers for polymerase chain reaction. BMC Bioinform. 2012, 13, 134. [Google Scholar] [CrossRef] [Green Version]
- Vašíček, J.; Baláži, A.; Parkányi, V.; Bauer, M. Different MACS sorting strategies for the enrichment of Lin– (CD34+ CD45–) hematopoietic progenitor cells: Preliminary study. Ann. Univ. Paedagog. Crac. Studia Nat. 2018, 3, 83–89. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Esfandiari, B.; Soliemani, M.; Kaviani, S.; Parivar, K. Rapid Neural Differentiation of Human Adipose Tissuederived Stem Cells Using NGF, Forskolin and bFGF. Biomed. Pharmacol. J. 2016, 9, 39–48. [Google Scholar] [CrossRef]
- Rafii, S.; Lyden, D. Therapeutic stem and progenitor cell transplantation for organ vascularization and regeneration. Nat. Med. 2003, 9, 702–712. [Google Scholar] [CrossRef]
- Medina, R.J.; O’Neill, C.L.; Humphreys, M.W.; Gardiner, T.A.; Stitt, A.W. Outgrowth endothelial cells: Characterization and their potential for reversing ischemic retinopathy. Investig. Ophthalmol. Vis. Sci. 2010, 51, 5906–5913. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Tao, Y.; Ren, S.; Liu, H.; Zhou, H.; Hu, J.; Tang, Y.; Zhang, B.; Chen, H. Simultaneous harvesting of endothelial progenitor cells and mesenchymal stem cells from the human umbilical cord. Exp. Ther. Med. 2017, 15, 806–812. [Google Scholar] [CrossRef] [Green Version]
- Fina, L.; Molgaard, H.V.; Robertson, D.; Bradley, N.J.; Monaghan, P.; Delia, D.; Sutherland, D.R.; Baker, M.A.; Greaves, M.F. Expression of the CD34 gene in vascular endothelial cells. Blood 1990, 75, 2417–2426. [Google Scholar] [CrossRef] [Green Version]
- Aplin, A.E.; Howe, A.; Alahari, S.K.; Juliano, R.L. Signal transduction and signal modulation by cell adhesion receptors: The role of integrins, cadherins, immunoglobulin-cell adhesion molecules, and selectins. Pharmacol. Rev. 1998, 50, 197–263. [Google Scholar]
- Bourguignon, L.Y.; Lokeshwar, V.B.; He, J.; Chen, X.; Bourguignon, G.J. A CD44-like endothelial cell transmembrane glycoprotein (GP116) interacts with extracellular matrix and ankyrin. Mol. Cell. Biol. 1992, 12, 4464–4471. [Google Scholar] [CrossRef] [Green Version]
- Liesveld, J.L.; Frediani, K.E.; Harbol, A.W.; DiPersio, J.F.; Abboud, C.N. Characterization of the adherence of normal and leukemic CD34+ cells to endothelial monolayers. Leukemia 1994, 8, 2111–2117. [Google Scholar]
- Xu, Y.; Swerlick, R.A.; Sepp, N.; Bosse, D.; Ades, E.W.; Lawley, T.J. Characterization of Expression and Modulation of Cell Adhesion Molecules on an Immortalized Human Dermal Microvascular Endothelial Cell Line (HMEC-1). J. Investig. Dermatol. 1994, 102, 833–837. [Google Scholar] [CrossRef] [Green Version]
- Trochon, V.; Mabilat, C.; Bertrand, P.; Legrand, Y.; Smadja-Joffe, F.; Soria, C.; Delpech, B.; Lu, H. Evidence of involvement of CD44 in endothelial cell proliferation, migration and angiogenesisin vitro. Int. J. Cancer 1996, 66, 664–668. [Google Scholar] [CrossRef]
- Krebsbach, P.H.; Villa-Diaz, L.G. The Role of Integrin α6 (CD49f) in Stem Cells: More than a Conserved Biomarker. Stem Cells Dev. 2017, 26, 1090–1099. [Google Scholar] [CrossRef]
- Grünewald, J.K.G.; Ridley, A.J. CD73 represses pro-inflammatory responses in human endothelial cells. J. Inflamm. 2010, 7, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kisselbach, L.; Merges, M.; Bossie, A.; Boyd, A. CD90 Expression on human primary cells and elimination of contaminating fibroblasts from cell cultures. Cytotechnology 2009, 59, 31–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masedunskas, A.; King, J.A.; Tan, F.; Cochran, R.; Stevens, T.; Sviridov, D.; Ofori-Acquah, S.F. Activated leukocyte cell adhesion molecule is a component of the endothelial junction involved in transendothelial monocyte migration. FEBS Lett. 2006, 580, 2637–2645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohneda, O.; Ohneda, K.; Arai, F.; Lee, J.; Miyamoto, T.; Fukushima, Y.; Dowbenko, D.; Lasky, L.A.; Suda, T. ALCAM (CD166): Its role in hematopoietic and endothelial development. Blood 2001, 98, 2134–2142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Green, L.; Ofstein, R.H.; Rapp, B.; Saadatzadeh, M.R.; Bhavsar, J.R.; Fajardo, A.; Dalsing, M.C.; Ingram, D.A.; Murphy, M.P. Adult venous endothelium is a niche for highly proliferative and vasculogenic endothelial colony-forming cells. J. Vasc. Surg. 2017, 66, 1854–1863. [Google Scholar] [CrossRef] [Green Version]
- Sobiesiak, M.; Sivasubramaniyan, K.; Hermann, C.; Tan, C.; Örgel, M.; Treml, S.; Cerabona, F.; De Zwart, P.; Ochs, U.; Müller, C.A.; et al. The Mesenchymal Stem Cell Antigen MSCA-1 is Identical to Tissue Non-specific Alkaline Phosphatase. Stem Cells Dev. 2010, 19, 669–677. [Google Scholar] [CrossRef]
- Fei, R.G.; Penn, P.E.; Wolf, N.S. A method to establish pure fibroblast and endothelial cell colony cultures from murine bone marrow. Exp. Hematol. 1990, 18, 953–957. [Google Scholar]
- Dave, J.M.; Bayless, K.J. Vimentin as an Integral Regulator of Cell Adhesion and Endothelial Sprouting. Microcirculation 2014, 21, 333–344. [Google Scholar] [CrossRef] [PubMed]
- Costa, M.L.; Escaleira, R.; Cataldo, A.; Oliveira, F.; Mermelstein, C.S. Desmin: Molecular interactions and putative functions of the muscle intermediate filament protein. Braz. J. Med Biol. Res. 2004, 37, 1819–1830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, X.; Dunn, J.; Dickinson, A.M.; Gillespie, J.I.; Baudouin, S.V. Smooth Muscle α-Actin Expression in Endothelial Cells Derived from CD34+ Human Cord Blood Cells. Stem Cells Dev. 2004, 13, 521–527. [Google Scholar] [CrossRef]
- Povsic, T.J.; Zavodni, K.L.; Vainorius, E.; Kherani, J.F.; Goldschmidt-Clermont, P.J.; Peterson, E.D. Common endothelial progenitor cell assays identify discrete endothelial progenitor cell populations. Am. Heart J. 2009, 157, 335–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohmura-Kakutani, H.; Akiyama, K.; Maishi, N.; Ohga, N.; Hida, Y.; Kawamoto, T.; Iida, J.; Shindoh, M.; Tsuchiya, K.; Shinohara, N.; et al. Identification of Tumor Endothelial Cells with High Aldehyde Dehydrogenase Activity and a Highly Angiogenic Phenotype. PLoS ONE 2014, 9, e113910. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Yuan, X.; Ou, Y.; Cai, Y.; Wang, S.; Sun, Q.; Zhang, W. Autophagy and apoptosis during adult adipose-derived stromal cells differentiation into neuron-like cells in vitro. Neural Regen. Res. 2012, 7, 1205–1212. [Google Scholar] [CrossRef] [PubMed]
- Mu, T.; Qin, Y.; Liu, B.; He, X.; Liao, Y.; Sun, J.; Qiu, J.; Li, X.; Zhong, Y.; Cai, J. In VitroNeural Differentiation of Bone Marrow Mesenchymal Stem Cells Carrying the FTH1 Reporter Gene and Detection with MRI. BioMed Res. Int. 2018, 2018, 8602. [Google Scholar] [CrossRef] [PubMed] [Green Version]







| Marker | Host/Isotype | Clone | Conjugate | Company |
|---|---|---|---|---|
| CD14 | mouse IgG2a | TÜK4 | PE | Dako Cytomation |
| CD29 | mouse IgG1 | P4G11 | FITC | Merck |
| CD31 | mouse IgG1 | C31.7 | PE | Novus Biologicals |
| CD34 | mouse IgG1 | QBEnd-10 | FITC | Thermo Fisher Scientific |
| CD44 | mouse IgG1 | W4/86 | purified 1 | Bio-Rad |
| CD45 | mouse IgG1 | L12/201 | purified 1 | Bio-Rad |
| CD49f | rat IgG2a | GoH3 | AF647 | BioLegend |
| CD73 | mouse IgG1 | AD2 | FITC | BD Biosciences |
| CD90 | mouse IgG1 | 5E10 | FITC | BD Biosciences |
| CD105 | mouse IgG1 | 266 | FITC | BD Biosciences |
| CD133 | mouse IgG1 | AC133 | PE | Miltenyi Biotec |
| CD146 | mouse IgG1 | P1H12 | PE | eBioscience |
| CD166 | rabbit IgG | polyclonal | purified 1 | Bioss |
| VE-cadherin | mouse IgG1 | F-8 | AF647 | Santa Cruz Biotechnology |
| Flk-1 (VEGFR-2) | mouse IgG1 | D-8 | AF647 | Santa Cruz Biotechnology |
| SSEA-4 | mouse IgG3 | MC-813-70 | PE | eBioscience |
| MSCA-1 | mouse IgG1 | W8B2 | PE | Miltenyi Biotec |
| Vimentin | mouse IgG2a | Vim 3B4 | purified 1 | Dako Cytomation |
| Desmin | mouse IgG1 | D33 | purified 1 | Dako Cytomation |
| α-SMA | mouse IgG2a | 1A4 | purified 1 | Dako Cytomation |
| vWF | mouse IgG1 | C-12 | FITC | Santa Cruz Biotechnology |
| NOS3 (eNOS) | mouse IgG2a | A-9 | FITC | Santa Cruz Biotechnology |
| Gene | Product Size (bp) | Forward Primer | Reverse Primer | Reference |
|---|---|---|---|---|
| CD29 | 287 | 5´-AGAATGTCACCAACCGTAGCA-3´ | 5´-CACAAAGGAGCCAAACCCA-3´ | [2] |
| CD31 | 138 | 5´-GTGATAATTGCCGCCTTGAT-3´ | 5´-GTTGGGATCTGACACGGTCT-3´ | XM_008271715.2 1 |
| CD34 | 155 | 5´-CTGAGGTTAGGGCTCAGTGC-3´ | 5´-GGAGTAGCTCTGGTGGCTTG-3´ | [50] |
| CD44 | 112 | 5´-TCATCCTGGCATCCCTCTTG-3´ | 5´-CCGTTGCCATTGTTGATCAC-3´ | [2] |
| CD45 | 262 | 5´-TACTCTGCCTCCCGTTG-3´ | 5´-GCTGAGTGTCTGCGTGTC-3´ | [2] |
| CD73 | 170 | 5´-CTCCTTTCCTCTCAAATCCAG-3´ | 5´-GTCCACGCCCTTCACTTTC-3´ | [2] |
| CD90 | 293 | 5´-CTGCTGCTGCTCTCACTGTC-3´ | 5´-ACAGAAGCAGCTTTGGGAAA-3´ | [2] |
| CD105 | 109 | 5´-TGACATACAGCACCAGCCAG-3´ | 5´-AGCTCTGACACCTCGTTTGG-3´ | [2] |
| CD133 | 124 | 5´-TCATTCCGATGGAACAGTCA-3´ | 5´-ATGAAGTTCTGGGCGTCATC-3´ | XM_017350443.1 1 |
| CD146 | 116 | 5´-GACAATGGCATCCTGGTCTT-3´ | 5´-AGTAGCTCGTGGCGTTCACT-3´ | [2] |
| CD166 | 345 | 5´-GCTCCCCAGTATTTATTGCCTTC-3´ | 5´-GTAGCACCTTTCCATTCCTGTA-3´ | [2] |
| ST3GAL2 (SSEA-4) | 126 | 5´-CTGGGAGAATAACCGGTACG-3´ | 5´-GCTCAGTTGCCTCGGTAGAC-3´ | [2] |
| ALPL (MSCA-1) | 137 | 5´-CCCTCATGTGATGGCTTACG-3´ | 5´-CTCAGAACAGGACGCTCAGG-3´ | [2] |
| ALDH | 135 | 5´-CTGGGAAAAGCAACCTGAAG-3´ | 5´-AACACTGGCCCTGATGGTAG-3´ | AB176450.1 1 |
| B2M | 118 | 5´-ATTCACGCCCAATGATAAGG-3´ | 5´-ATCCTCAGACCTCCATGCTG-3´ | [2] |
| Cell Status /Passage | Cell Type | P0 | P1 | P2 | P3 |
|---|---|---|---|---|---|
| Live | EPCs | 83.8 ± 4.8 | 84.7 ± 1.3 | 91.5 ± 2.9 | 85.1 ± 9.3 |
| BEPCs | 93.0 ± 3.3 | 89.2 ± 6.7 | 92.5 ± 2.8 | 93.2 ± 1.3 | |
| Apoptotic | EPCs | 0.9 ± 0.6 | 1.0 ± 0.8 | 0.6 ± 0.1 | 0.3 ± 0.1 |
| BEPCs | 1.0 ± 0.4 | 1.3 ± 0.9 | 2.1 ± 0.9 | 1.9 ± 0.8 | |
| Dead | EPCs | 15.4 ± 4.6 | 14.2 ± 1.9 | 7.9 ± 3.0 | 14.6 ± 9.4 |
| BEPCs | 6.0 ± 3.1 | 9.5 ± 5.9 | 5.5 ± 2.1 | 4.9 ± 0.9 |
| Marker/Passage | Cell Type | P0 | P1 | P2 | P3 | HUVECs (P3) |
|---|---|---|---|---|---|---|
| CD14 | EPCs | 8.0 ± 7.9 | 2.0 ± 2.3 * | 0.9 ± 0.7 ** | 1.2 ± 0.9 ** | 0.1 ± 0.0 |
| BEPCs | 7.5 ± 6.4 | 1.1 ± 1.0 * | 0.1 ± 0.1 ** | 0.1 ± 0. 0 ** | ||
| CD29 | EPCs | 84.3 ± 9.1 | 90.7 ± 6.4 | 94.3 ± 6.3 * | 95.2 ± 4.3 ** | 91.2 ± 0.3 |
| BEPCs | 81.1 ± 4.0 | 93.5 ± 7.6 | 89.4 ± 11.4 | 91.8 ± 4.3 | ||
| CD31 | EPCs | 11.6 ± 9.0 | 4.3 ± 3.9 * | 2.0 ± 1.2 ** | 1.1 ± 0.4 ** | 81.5 ± 1.2 |
| BEPCs | 19.8 ± 8.9 | 7.7 ± 1.2 ** | 4.7 ± 2.5 *** | 1.8 ± 1.0 *** | ||
| CD34 | EPCs | 0.8 ± 0.3 | 0.6 ± 0.3 | 0.4 ± 0.3 | 0.5 ± 0.2 | 0.6 ± 0.6 |
| BEPCs | 0.7 ± 0.5 | 0.4 ±0.2 | 0.1 ± 0.1 | 0.1 ± 0.1 | ||
| CD44 | EPCs | 83.6 ± 12.2 | 85.9 ± 13.5 | 96.5 ± 2.2 | 94.6 ± 4.7 | n/a |
| BEPCs | 96.7 ± 1.9 | 96.6 ± 4.3 | 96.9 ± 1.0 | 94.2 ± 3.9 | ||
| CD45 | EPCs | 10.1 ± 9.1 | 3.0 ± 3.4 ** | 1.8 ± 2.4 ** | 1.3 ± 1.1 *** | n/a |
| BEPCs | 15.7 ± 10.3 | 1.8 ± 1.1 ** | 0.3 ± 0.1 *** | 0.2 ± 0.1 *** | ||
| CD49f | EPCs | n/a | n/a | n/a | 85.5 ± 6.8 | 94.1 ± 8.4 |
| BEPCs | n/a | n/a | n/a | 85.8 ± 11.0 | ||
| CD73 | EPCs | n/a | n/a | n/a | 68.4 ± 11.5 | 91.0 ± 1.4 |
| BEPCs | n/a | n/a | n/a | 71.3 ± 6.9 | ||
| CD90 | EPCs | n/a | n/a | n/a | 90.4 ± 9.82 | 12.6 ± 0.5 |
| BEPCs | n/a | n/a | n/a | 88.1 ± 11.9 | ||
| CD105 | EPCs | n/a | n/a | n/a | 41.4 ± 6.0 | 83.7 ± 1.2 |
| BEPCs | n/a | n/a | n/a | 33.0 ± 21.2 | ||
| CD133 | EPCs | 0.7 ± 0.3 | 0.4 ± 0.3 | 0.3 ± 0.2 | 0.5 ± 0.5 | 0.5 ± 0.2 |
| BEPCs | 1.2 ± 1.2 | 0.4 ± 0.2 | 0.2 ± 0.1 | 0.3 ± 0.4 | ||
| CD146 | EPCs | 2.3 ± 1.8 | 2.0 ± 2.3 | 2.6 ± 3.8 | 6.3 ± 7.1 | 83.7 ± 18.5 |
| BEPCs | 1.5 ± 1.1 | 3.9 ± 1.9 * | 0.9 ± 0.8 | 0.3 ± 0.4 | ||
| CD166 | EPCs | 0.2 ± 0.2 | 0.1 ± 0.0 | 0.1 ± 0.0 | 0.1 ± 0.0 | n/a |
| BEPCs | n/a | n/a | n/a | n/a | ||
| VE-cadherin | EPCs | 94.8 ± 4.3 | 98.1 ± 1.2 * | 98.8 ± 1.0 ** | 98.4 ± 1.7 * | 84.3 ± 12.7 |
| BEPCs | 99.1 ± 0.4 | 99.8 ± 0.1 | 99.8 ± 0.2 | 96.0 ± 6.7 | ||
| VEGFR-2 | EPCs | 90.2 ± 8.2 | 94.5 ± 8.6 | 97.1 ± 1.8 | 97.4 ± 2.3 | 94.8 ± 5.3 |
| BEPCs | 97.8 ± 2.0 | 99.4 ± 0.3 | 99.2 ± 0.5 | 94.1 ± 9.0 | ||
| SSEA-4 | EPCs | 6.2 ± 5.2 | 5.7 ± 10.1 | 1.4 ± 1.1 | 1.0 ± 0.7 | 1.0 ± 0.1 |
| BEPCs | 59.6 ± 29.7 | 19.5 ± 11.8 * | 14.3 ± 9.7 ** | 25.4 ± 18.0 * | ||
| MSCA-1 | EPCs | 0.5 ± 0.4 | 0.4 ± 0.3 | 0.1 ± 0.2 | 0.2 ± 0.1 | n/a |
| BEPCs | 1.0 ± 0.5 | 0.3 ± 0.2 | 0.0 ± 0.0 | 0.1 ± 0.0 |
| Marker/Passage | Cell Type | P0 | P1 | P2 | P3 | HUVECs (P3) |
|---|---|---|---|---|---|---|
| AcLDL | EPCs | 80.8 ± 10.8 | 92.8 ± 7.8 | 88.7 ± 13.9 | 92.2 ± 5.4 | 86.2 ± 1.0 |
| BEPCs | 99.3 ± 0.5 | 99.9 ± 0.2 | 98.8 ± 1.0 | 97.6 ± 0.9 ** | ||
| ALDH | EPCs | 84.9 ± 5.6 | 78.9 ± 10.3 | 80.4 ± 15.4 | 94.5 ± 5.0 | 80.0 ± 1.8 |
| BEPCs | 24.7 ± 4.9 | 46.1 ± 25.2 | 48.6 ± 40.4 | 25.3 ± 10.5 | ||
| Vimentin | EPCs | 81.3 ± 5.9 | 87.3 ± 7.7 | 89.6 ± 7.3 | 86.3 ± 9.0 | n/a |
| BEPCs | 99.3 ± 0.3 | 98.0 ± 1.5 | 99.4 ± 0.5 | 99.0 ± 0.7 | ||
| Desmin | EPCs | 55.1 ± 18.2 | 59.0 ± 17.9 | 68.3 ± 20.2 | 64.2 ± 14.0 | n/a |
| BEPCs | 89.5 ± 5.8 | 92.3 ± 6.0 | 94.8 ± 3.4 | 92.4 ± 5.4 | ||
| α-SMA | EPCs | 74.0 ± 5.9 | 85.6 ± 11.1 | 88.5 ± 4.9 | 82.5 ± 9.9 | n/a |
| BEPCs | 99.0 ± 0.3 | 96.9 ± 2.0 | 98.1 ± 1.1 | 92.7 ± 10.4 | ||
| vWF | EPCs | 98.0 ± 0.1 | 98.3 ± 0.8 | 99.4 ± 0.3 * | 99.8 ± 0.1 ** | 98.8 ± 1.5 |
| BEPCs | 97.4 ± 1.4 | 99.5 ± 0.3 ** | 99.7 ± 0.2 *** | 99.8 ± 0.1 *** | ||
| eNOS | EPCs | 94.8 ± 6.0 | 99.0 ± 0.3 | 99.4 ± 0.2 | 99.7 ± 0.1 | 98.9 ± 1.0 |
| BEPCs | 99.8 ± 0.1 | 99.9 ± 0.1 | 99.9 ± 0.1 | 99.9 ± 0.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vašíček, J.; Baláži, A.; Bauer, M.; Svoradová, A.; Tirpáková, M.; Tomka, M.; Chrenek, P. Molecular Profiling and Gene Banking of Rabbit EPCs Derived from Two Biological Sources. Genes 2021, 12, 366. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12030366
Vašíček J, Baláži A, Bauer M, Svoradová A, Tirpáková M, Tomka M, Chrenek P. Molecular Profiling and Gene Banking of Rabbit EPCs Derived from Two Biological Sources. Genes. 2021; 12(3):366. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12030366
Chicago/Turabian StyleVašíček, Jaromír, Andrej Baláži, Miroslav Bauer, Andrea Svoradová, Mária Tirpáková, Marián Tomka, and Peter Chrenek. 2021. "Molecular Profiling and Gene Banking of Rabbit EPCs Derived from Two Biological Sources" Genes 12, no. 3: 366. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12030366