
Potential Genetic Contributions of the Central Nervous System to a Predisposition to Elite Athletic Traits: State-of-the-Art and Future Perspectives


Abstract
:1. Introduction
2. Brain Areas and Functions Related to a Predisposition to High EC: Findings of Rodent-Based Studies
2.1. Emerging Roles of Pain Analgesia System
2.2. Differential Roles of the Dopaminergic and Serotonergic Systems
2.3. Emerging Roles of Neuronal Circuits Related to Neuroplasticity
2.4. Potential Roles of Thermoregulation-Related Factors
2.5. A Summary of Findings of Rodent-Based Studies
3. CNS-Related Genes That Potentially Contribute to a Predisposition to Elite Athletic Traits: Findings of Human Studies
3.1. Genes Related to Motor Skills, Motor Learning, Motor Performance, and Sleep
3.2. Genes Related to the Serotonergic System
3.3. Genes Related to the Dopaminergic System
3.4. Transcranial Direct Current Stimulation (tDCS) and Genes Potentially Related to Responsiveness to tDCS
3.5. A Summary of Findings of Human Studies
4. Ethical Issues
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| 5-HIAA | 5-hydroxyindoleacetic acid |
| 5-HT | 5-hydroxytryptamine |
| CNS | Central nervous system |
| CPu | Caudate putamen |
| DA | Dopamine |
| DOPAC | 3,4-dihydroxyphenylacetic acid |
| EC | Exercise capacity |
| GABA | γ-aminobutyric acid |
| NAc | Nucleus accumbens |
| PFC | Prefrontal cortex |
| SNPs | Single nucleotide polymorphisms |
| tDCS | Transcranial direct current stimulation |
References
- De Moor, M.H.; Spector, T.D.; Cherkas, L.F.; Falchi, M.; Hottenga, J.J.; Boomsma, D.I.; De Geus, E.J. Genome-wide linkage scan for athlete status in 700 British female DZ twin pairs. Twin Res. Hum. Genet. 2007, 10, 812–820. [Google Scholar] [CrossRef] [Green Version]
- Maciejewska-Skrendo, A.; Cieszczyk, P.; Chycki, J.; Sawczuk, M.; Smolka, W. Genetic Markers Associated with Power Athlete Status. J. Hum. Kinet. 2019, 68, 17–36. [Google Scholar] [CrossRef] [Green Version]
- Guest, N.S.; Horne, J.; Vanderhout, S.M.; El-Sohemy, A. Sport Nutrigenomics: Personalized Nutrition for Athletic Performance. Front. Nutr. 2019, 6, 8. [Google Scholar] [CrossRef]
- Pickering, C.; Kiely, J.; Grgic, J.; Lucia, A.; Del Coso, J. Can Genetic Testing Identify Talent for Sport? Genes 2019, 10, 972. [Google Scholar] [CrossRef] [Green Version]
- Jacob, Y.; Spiteri, T.; Hart, N.H.; Anderton, R.S. The Potential Role of Genetic Markers in Talent Identification and Athlete Assessment in Elite Sport. Sports 2018, 6, 88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foley, T.E.; Brooks, L.R.; Gilligan, L.J.; Burghardt, P.R.; Koch, L.G.; Britton, S.L.; Fleshner, M. Brain activation patterns at exhaustion in rats that differ in inherent exercise capacity. PLoS ONE 2012, 7, e45415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Z.; Davis, H.I.; Wolff, A.; Northoff, G. Thalamo-Sensorimotor Functional Connectivity Correlates with World Ranking of Olympic, Elite, and High Performance Athletes. Neural Plast. 2017, 2017, 1473783. [Google Scholar] [CrossRef]
- Roberts, M.D.; Ruegsegger, G.N.; Brown, J.D.; Booth, F.W. Mechanisms Associated With Physical Activity Behavior: Insights From Rodent Experiments. Exerc. Sport Sci. Rev. 2017, 45, 217–222. [Google Scholar] [CrossRef]
- Arvanitakis, Z.; Shah, R.C.; Bennett, D.A. Diagnosis and Management of Dementia: Review. JAMA 2019, 322, 1589–1599. [Google Scholar] [CrossRef] [PubMed]
- Erickson, K.I.; Voss, M.W.; Prakash, R.S.; Basak, C.; Szabo, A.; Chaddock, L.; Kim, J.S.; Heo, S.; Alves, H.; White, S.M.; et al. Exercise training increases size of hippocampus and improves memory. Proc. Natl. Acad. Sci. USA 2011, 108, 3017–3022. [Google Scholar] [CrossRef] [Green Version]
- Law, C.K.; Lam, F.M.; Chung, R.C.; Pang, M.Y. Physical exercise attenuates cognitive decline and reduces behavioural problems in people with mild cognitive impairment and dementia: A systematic review. J. Physiother. 2020, 66, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Hillman, C.H.; Erickson, K.I.; Kramer, A.F. Be smart, exercise your heart: Exercise effects on brain and cognition. Nat. Rev. Neurosci. 2008, 9, 58–65. [Google Scholar] [CrossRef]
- Ma, C.L.; Ma, X.T.; Wang, J.J.; Liu, H.; Chen, Y.F.; Yang, Y. Physical exercise induces hippocampal neurogenesis and prevents cognitive decline. Behav. Brain Res. 2017, 317, 332–339. [Google Scholar] [CrossRef] [PubMed]
- Di Liegro, C.M.; Schiera, G.; Proia, P.; Di Liegro, I. Physical Activity and Brain Health. Genes 2019, 10, 720. [Google Scholar] [CrossRef] [Green Version]
- Roberts, M.D.; Gilpin, L.; Parker, K.E.; Childs, T.E.; Will, M.J.; Booth, F.W. Dopamine D1 receptor modulation in nucleus accumbens lowers voluntary wheel running in rats bred to run high distances. Physiol. Behav. 2012, 105, 661–668. [Google Scholar] [CrossRef]
- Caetano-Anolles, K.; Rhodes, J.S.; Garland, T., Jr.; Perez, S.D.; Hernandez, A.G.; Southey, B.R.; Rodriguez-Zas, S.L. Cerebellum Transcriptome of Mice Bred for High Voluntary Activity Offers Insights into Locomotor Control and Reward-Dependent Behaviors. PLoS ONE 2016, 11, e0167095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koch, L.G.; Britton, S.L. Artificial selection for intrinsic aerobic endurance running capacity in rats. Physiol. Genom. 2001, 5, 45–52. [Google Scholar] [CrossRef] [Green Version]
- Rabelo, P.C.; Almeida, T.F.; Guimaraes, J.B.; Barcellos, L.A.; Cordeiro, L.M.; Moraes, M.M.; Coimbra, C.C.; Szawka, R.E.; Soares, D.D. Intrinsic exercise capacity is related to differential monoaminergic activity in the rat forebrain. Brain Res. Bull. 2015, 112, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Rabelo, P.C.R.; Horta, N.A.C.; Cordeiro, L.M.S.; Poletini, M.O.; Coimbra, C.C.; Szawka, R.E.; Soares, D.D. Intrinsic exercise capacity in rats influences dopamine neuroplasticity induced by physical training. J. Appl. Physiol. 2017, 123, 1721–1729. [Google Scholar] [CrossRef]
- Rabelo, P.C.R.; Cordeiro, L.M.S.; Aquino, N.S.S.; Fonseca, B.B.B.; Coimbra, C.C.; Wanner, S.P.; Szawka, R.E.; Soares, D.D. Rats with higher intrinsic exercise capacities exhibit greater preoptic dopamine levels and greater mechanical and thermoregulatory efficiencies while running. J. Appl. Physiol. 2019, 126, 393–402. [Google Scholar] [CrossRef] [Green Version]
- Hosp, J.A.; Luft, A.R. Dopaminergic meso-cortical projections to m1: Role in motor learning and motor cortex plasticity. Front. Neurol. 2013, 4, 145. [Google Scholar] [CrossRef] [Green Version]
- Ong, W.Y.; Stohler, C.S.; Herr, D.R. Role of the Prefrontal Cortex in Pain Processing. Mol. Neurobiol. 2019, 56, 1137–1166. [Google Scholar] [CrossRef] [Green Version]
- Lv, Q.; Wu, F.; Gan, X.; Yang, X.; Zhou, L.; Chen, J.; He, Y.; Zhang, R.; Zhu, B.; Liu, L. The Involvement of Descending Pain Inhibitory System in Electroacupuncture-Induced Analgesia. Front. Integr. Neurosci. 2019, 13, 38. [Google Scholar] [CrossRef]
- Cordeiro, L.M.S.; Rabelo, P.C.R.; Moraes, M.M.; Teixeira-Coelho, F.; Coimbra, C.C.; Wanner, S.P.; Soares, D.D. Physical exercise-induced fatigue: The role of serotonergic and dopaminergic systems. Braz. J. Med. Biol. Res. 2017, 50, e6432. [Google Scholar] [CrossRef]
- Kupnicka, P.; Listos, J.; Tarnowski, M.; Kolasa-Wolosiuk, A.; Wasik, A.; Lukomska, A.; Barczak, K.; Gutowska, I.; Chlubek, D.; Baranowska-Bosiacka, I. Fluoride Affects Dopamine Metabolism and Causes Changes in the Expression of Dopamine Receptors (D1R and D2R) in Chosen Brain Structures of Morphine-Dependent Rats. Int. J. Mol. Sci. 2020, 21, 2361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, T.W.; Tsai, S.F.; Kuo, Y.M. Physical Exercise Enhances Neuroplasticity and Delays Alzheimer’s Disease. Brain Plast. 2018, 4, 95–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keloglan, S.; Sahin, L.; Cevik, O.S. Long-term treadmill exercise upregulated hippocampal learning-related genes without improving cognitive behavior in social isolated rats. Folia Morphol. Warsz. 2019, 4, 668–675. [Google Scholar] [CrossRef] [PubMed]
- Halliday, M.R.; Abeydeera, D.; Lundquist, A.J.; Petzinger, G.M.; Jakowec, M.W. Intensive treadmill exercise increases expression of hypoxia-inducible factor 1alpha and its downstream transcript targets: A potential role in neuroplasticity. Neuroreport 2019, 30, 619–627. [Google Scholar] [CrossRef]
- Li, X.; Wang, L.; Zhang, S.; Hu, X.; Yang, H.; Xi, L. Timing-Dependent Protection of Swimming Exercise against d-Galactose-Induced Aging-Like Impairments in Spatial Learning/Memory in Rats. Brain Sci. 2019, 9, 236. [Google Scholar] [CrossRef] [Green Version]
- Ewald, R.C.; Cline, H.T. NMDA receptors and brain development. In Biology of the NMDA Receptor; Van Dongen, A.M., Ed.; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2009. [Google Scholar]
- Wanner, S.P.; Primola-Gomes, T.N.; Pires, W.; Guimaraes, J.B.; Hudson, A.S.; Kunstetter, A.C.; Fonseca, C.G.; Drummond, L.R.; Damasceno, W.C.; Teixeira-Coelho, F. Thermoregulatory responses in exercising rats: Methodological aspects and relevance to human physiology. Temp. Austin 2015, 2, 457–475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morrison, S.F.; Nakamura, K. Central Mechanisms for Thermoregulation. Annu. Rev. Physiol. 2019, 81, 285–308. [Google Scholar] [CrossRef]
- Ahmetov, I.I.; Egorova, E.S.; Gabdrakhmanova, L.J.; Fedotovskaya, O.N. Genes and Athletic Performance: An Update. Med. Sport Sci. 2016, 61, 41–54. [Google Scholar] [CrossRef]
- Peplonska, B.; Safranow, K.; Adamczyk, J.; Boguszewski, D.; Szymanski, K.; Soltyszewski, I.; Barczak, A.; Siewierski, M.; Ploski, R.; Sozanski, H.; et al. Association of serotoninergic pathway gene variants with elite athletic status in the Polish population. J. Sports Sci. 2019, 37, 1655–1662. [Google Scholar] [CrossRef]
- Jacob, Y.; Chivers, P.; Anderton, R.S. Genetic predictors of match performance in sub-elite Australian football players: A pilot study. J. Exerc. Sci. Fit. 2019, 17, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Jacob, Y.; Cripps, A.; Evans, T.; Chivers, P.T.; Joyce, C.; Anderton, R.S. Identification of genetic markers for skill and athleticism in sub-elite Australian football players: A pilot study. J. Sports Med. Phys. Fit. 2018, 58, 241–248. [Google Scholar] [CrossRef]
- Morin-Moncet, O.; Beaumont, V.; de Beaumont, L.; Lepage, J.F.; Theoret, H. BDNF Val66Met polymorphism is associated with abnormal interhemispheric transfer of a newly acquired motor skill. J. Neurophysiol. 2014, 111, 2094–2102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noohi, F.; Boyden, N.B.; Kwak, Y.; Humfleet, J.; Burke, D.T.; Muller, M.L.; Bohnen, N.I.; Seidler, R.D. Association of COMT val158met and DRD2 G>T genetic polymorphisms with individual differences in motor learning and performance in female young adults. J. Neurophysiol. 2014, 111, 628–640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, G.; Xing, L.; Wu, D.; Bhattacharyya, B.J.; Jones, C.R.; McMahon, T.; Chong, S.Y.C.; Chen, J.A.; Coppola, G.; Geschwind, D.; et al. A Rare Mutation of beta1-Adrenergic Receptor Affects Sleep/Wake Behaviors. Neuron 2019, 103, 1044–1055 e1047. [Google Scholar] [CrossRef] [Green Version]
- Charest, J.; Grandner, M.A. Sleep and Athletic Performance: Impacts on Physical Performance, Mental Performance, Injury Risk and Recovery, and Mental Health. Sleep Med. Clin. 2020, 15, 41–57. [Google Scholar] [CrossRef]
- Xing, L.; Shi, G.; Mostovoy, Y.; Gentry, N.W.; Fan, Z.; McMahon, T.B.; Kwok, P.Y.; Jones, C.R.; Ptacek, L.J.; Fu, Y.H. Mutant neuropeptide S receptor reduces sleep duration with preserved memory consolidation. Sci. Transl. Med. 2019, 11. [Google Scholar] [CrossRef] [Green Version]
- Petr, M.; Maciejewska-Skrendo, A.; Zajac, A.; Chycki, J.; Stastny, P. Association of Elite Sports Status with Gene Variants of Peroxisome Proliferator Activated Receptors and Their Transcriptional Coactivator. Int. J. Mol. Sci. 2019, 21, 162. [Google Scholar] [CrossRef] [Green Version]
- Bethea, C.L.; Lima, F.B.; Centeno, M.L.; Weissheimer, K.V.; Senashova, O.; Reddy, A.P.; Cameron, J.L. Effects of citalopram on serotonin and CRF systems in the midbrain of primates with differences in stress sensitivity. J. Chem. Neuroanat. 2011, 41, 200–218. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Maejima, T.; Wyler, S.C.; Casadesus, G.; Herlitze, S.; Deneris, E.S. Pet-1 is required across different stages of life to regulate serotonergic function. Nat. Neurosci. 2010, 13, 1190–1198. [Google Scholar] [CrossRef]
- Currier, D.; Mann, J.J. Stress, genes and the biology of suicidal behavior. Psychiatr. Clin. N. Am. 2008, 31, 247–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belmer, A.; Quentin, E.; Diaz, S.L.; Guiard, B.P.; Fernandez, S.P.; Doly, S.; Banas, S.M.; Pitychoutis, P.M.; Moutkine, I.; Muzerelle, A.; et al. Positive regulation of raphe serotonin neurons by serotonin 2B receptors. Neuropsychopharmacology 2018, 43, 1623–1632. [Google Scholar] [CrossRef] [Green Version]
- Gee, C.J. How does sport psychology actually improve athletic performance? A framework to facilitate athletes’ and coaches’ understanding. Behav. Modif. 2010, 34, 386–402. [Google Scholar] [CrossRef] [PubMed]
- Mang, C.S.; McEwen, L.M.; MacIsaac, J.L.; Snow, N.J.; Campbell, K.L.; Kobor, M.S.; Ross, C.J.D.; Boyd, L.A. Exploring genetic influences underlying acute aerobic exercise effects on motor learning. Sci. Rep. 2017, 7, 12123. [Google Scholar] [CrossRef] [PubMed]
- Levin, H.; Troyanskaya, M.; Petrie, J.; Wilde, E.A.; Hunter, J.V.; Abildskov, T.J.; Scheibel, R.S. Methylphenidate Treatment of Cognitive Dysfunction in Adults After Mild to Moderate Traumatic Brain Injury: Rationale, Efficacy, and Neural Mechanisms. Front. Neurol. 2019, 10, 925. [Google Scholar] [CrossRef]
- Tartar, J.L.; Cabrera, D.; Knafo, S.; Thomas, J.D.; Antonio, J.; Peacock, C.A. The “Warrior” COMT Val/Met Genotype Occurs in Greater Frequencies in Mixed Martial Arts Fighters Relative to Controls. J. Sports Sci. Med. 2020, 19, 38–42. [Google Scholar] [PubMed]
- Nagano-Saito, A.; Dagher, A.; Booij, L.; Gravel, P.; Welfeld, K.; Casey, K.F.; Leyton, M.; Benkelfat, C. Stress-induced dopamine release in human medial prefrontal cortex--18F-fallypride/PET study in healthy volunteers. Synapse 2013, 67, 821–830. [Google Scholar] [CrossRef]
- Reardon, S. ‘Brain doping’ may improve athletes’ performance. Nature 2016, 531, 283–284. [Google Scholar] [CrossRef]
- Holgado, D.; Vadillo, M.A.; Sanabria, D. “Brain-Doping”, Is It a Real Threat? Front. Physiol. 2019, 10, 483. [Google Scholar] [CrossRef]
- Zhu, Z.; Zhou, J.; Manor, B.; Wang, X.; Fu, W.; Liu, Y. Commentary: “Brain-Doping”, Is It a Real Threat? Front. Physiol. 2019, 10, 1489. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Stephenson, M.C.; Morris, P.G.; Jackson, S.R. tDCS-induced alterations in GABA concentration within primary motor cortex predict motor learning and motor memory: A 7 T magnetic resonance spectroscopy study. Neuroimage 2014, 99, 237–243. [Google Scholar] [CrossRef] [Green Version]
- Kolasinski, J.; Hinson, E.L.; Divanbeighi Zand, A.P.; Rizov, A.; Emir, U.E.; Stagg, C.J. The dynamics of cortical GABA in human motor learning. J. Physiol. 2019, 597, 271–282. [Google Scholar] [CrossRef] [Green Version]
- World Anti-Doping Agency. World Anti-Doping Code 2021. Available online: https://www.wada-ama.org/sites/default/files/resources/files/2021_wada_code.pdf (accessed on 11 January 2021).
- Davis, J.M.; Alderson, N.L.; Welsh, R.S. Serotonin and central nervous system fatigue: Nutritional considerations. Am. J. Clin. Nutr. 2000, 72, 573–578. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Gonzalez, L.M.; Sanchez-Oliver, A.J.; Mata, F.; Jodra, P.; Antonio, J.; Dominguez, R. Acute caffeine supplementation in combat sports: A systematic review. J. Int. Soc. Sports Nutr. 2018, 15, 60. [Google Scholar] [CrossRef] [Green Version]
- De Azevedo, A.P.; Guerra, M.A., Jr.; Caldas, L.C.; Guimaraes-Ferreira, L. Acute Caffeine Ingestion did not Enhance Punch Performance in Professional Mixed-Martial Arts Athletes. Nutrients 2019, 11, 1422. [Google Scholar] [CrossRef] [Green Version]
- Foster, P.P.; Rosenblatt, K.P.; Kuljis, R.O. Exercise-induced cognitive plasticity, implications for mild cognitive impairment and Alzheimer’s disease. Front. Neurol. 2011, 2, 28. [Google Scholar] [CrossRef] [Green Version]
- Nascimento, C.M.; Pereira, J.R.; Pires de Andrade, L.; Garuffi, M.; Ayan, C.; Kerr, D.S.; Talib, L.L.; Cominetti, M.R.; Stella, F. Physical exercise improves peripheral BDNF levels and cognitive functions in mild cognitive impairment elderly with different bdnf Val66Met genotypes. J. Alzheimers Dis. 2015, 43, 81–91. [Google Scholar] [CrossRef]
- Karpowicz, K.; Krych, K.; Karpowicz, M.; Nowak, W.; Gronek, P. The relationship between CA repeat polymorphism of the IGF-1 gene and the structure of motor skills in young athletes. Acta Biochim. Pol. 2018, 65, 43–50. [Google Scholar] [CrossRef] [Green Version]
- Munoz, A.; Lopez-Lopez, A.; Labandeira, C.M.; Labandeira-Garcia, J.L. Interactions Between the Serotonergic and Other Neurotransmitter Systems in the Basal Ganglia: Role in Parkinson’s Disease and Adverse Effects of L-DOPA. Front. Neuroanat. 2020, 14, 26. [Google Scholar] [CrossRef] [PubMed]


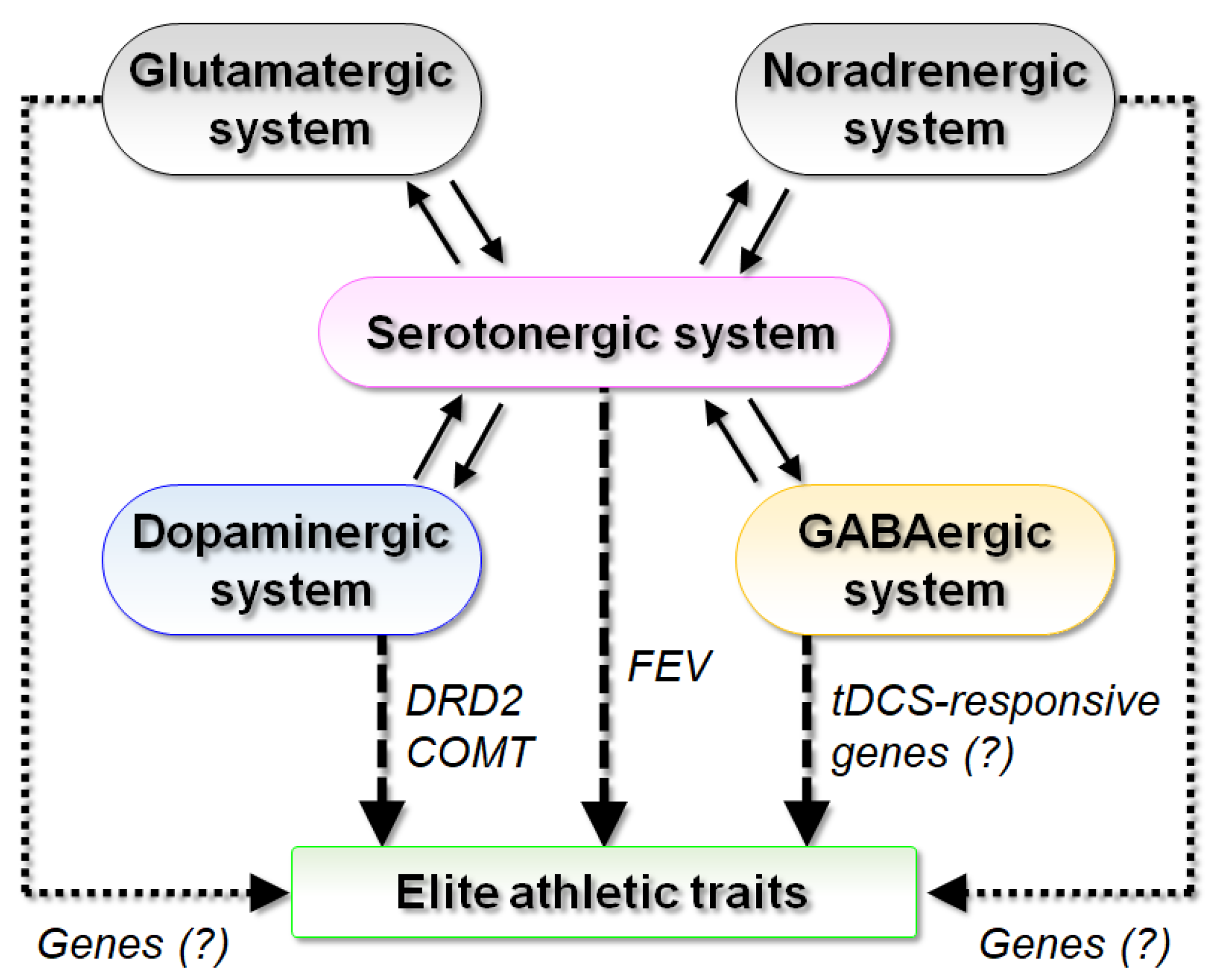{kind=link}
| CNS Functions Potentially Related to EC | Genes Involved/Analyzed | Animals and Related EC | Reference |
|---|---|---|---|
| Pain analgesia system | Not identified | Rats with inherent high EC | [6] |
| Serotonergic system | Not identified | Rats with inherent high EC | [18] |
| Dopaminergic system | Not identified | Neuroplasticity in rats with inherent high EC | [19] |
| Thermoregulatory system | Not identified | Rats with inherent high EC | [20] |
| Genes/Genotypes | Potential CNS Functions Related to Athletic Traits | References |
|---|---|---|
| Brain-derived neurotropic factor (BDNF) A/G (rs6265) | Modulation of interhemispheric transfer of a procedural motor skill | [35,37] |
| Dopamine D2 receptor (DRD2) A/A (rs1076560) | Motor learning and performance | [35,38] |
| β 1 adrenergic receptor (ADRB1) C/T (rs776439595) | Sleep regulation | [39] |
| Fifth Ewing variant (FEV) G/G (rs860573) | Elevated serotonergic activity for optimal performance | [34] |
| Catechol-O-methyltransferase (COMT) G/G (rs4680) | Attainment of optimal dopamine dose under athletic competition | [50] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kitazawa, H.; Hasegawa, K.; Aruga, D.; Tanaka, M. Potential Genetic Contributions of the Central Nervous System to a Predisposition to Elite Athletic Traits: State-of-the-Art and Future Perspectives. Genes 2021, 12, 371. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12030371
Kitazawa H, Hasegawa K, Aruga D, Tanaka M. Potential Genetic Contributions of the Central Nervous System to a Predisposition to Elite Athletic Traits: State-of-the-Art and Future Perspectives. Genes. 2021; 12(3):371. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12030371
Chicago/Turabian StyleKitazawa, Hiroya, Kazuya Hasegawa, Daichi Aruga, and Masashi Tanaka. 2021. "Potential Genetic Contributions of the Central Nervous System to a Predisposition to Elite Athletic Traits: State-of-the-Art and Future Perspectives" Genes 12, no. 3: 371. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12030371