
Cytological Observation and Transcriptome Comparative Analysis of Self-Pollination and Cross-Pollination in Dendrobium Officinale

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. Histological Observe of Pollen Tube Growth
2.3. Total RNA Extraction, Library Preparation and Transcriptome Sequencing
2.4. Reads Mapping to the Reference Genome
2.5. Identification of Differentially Expressed Genes (DEGs)
2.6. Function Annotation and Enrichment Analysis
2.7. qRT-PCR Validate Analysis
3. Results
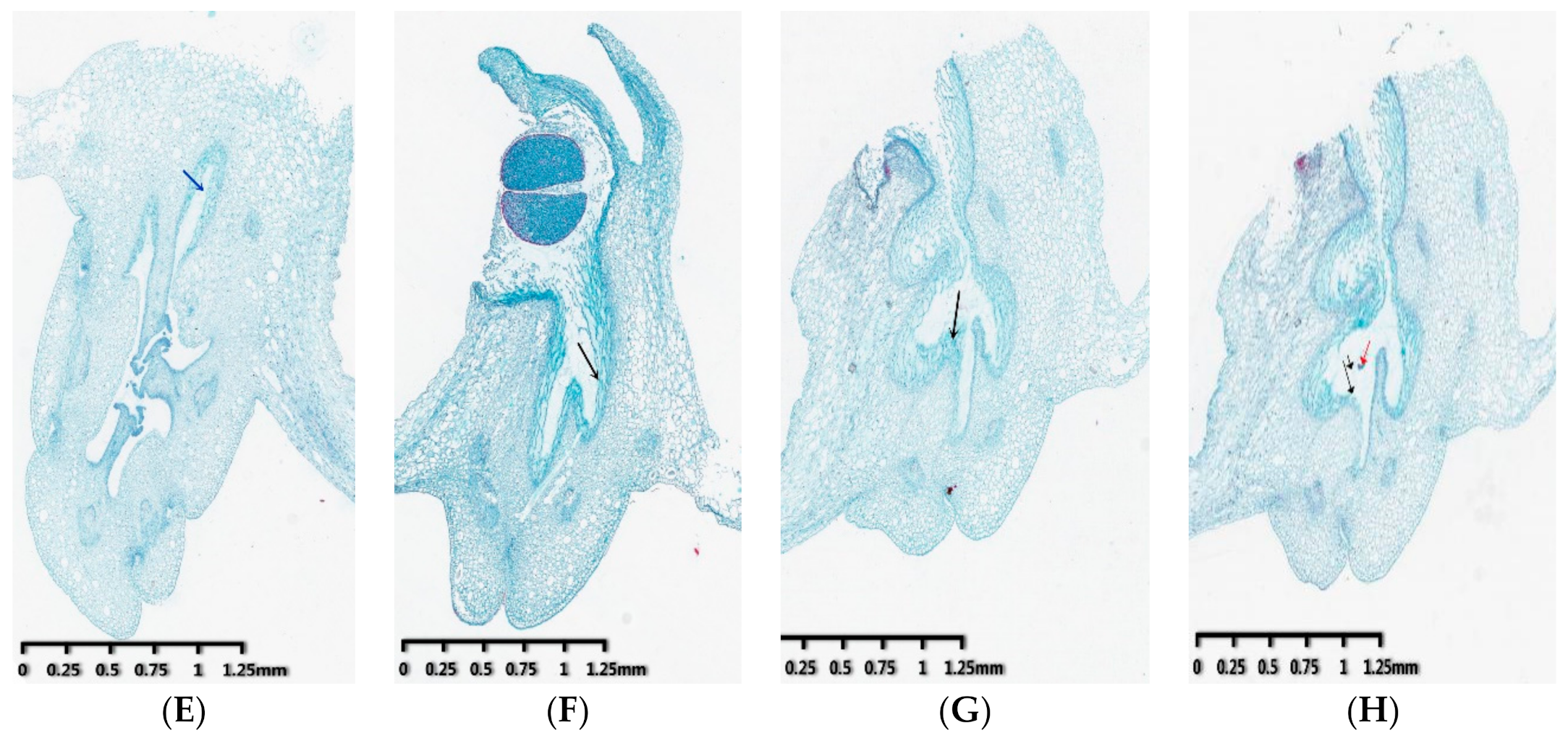
3.1. Observation of Pollen Tube Growth
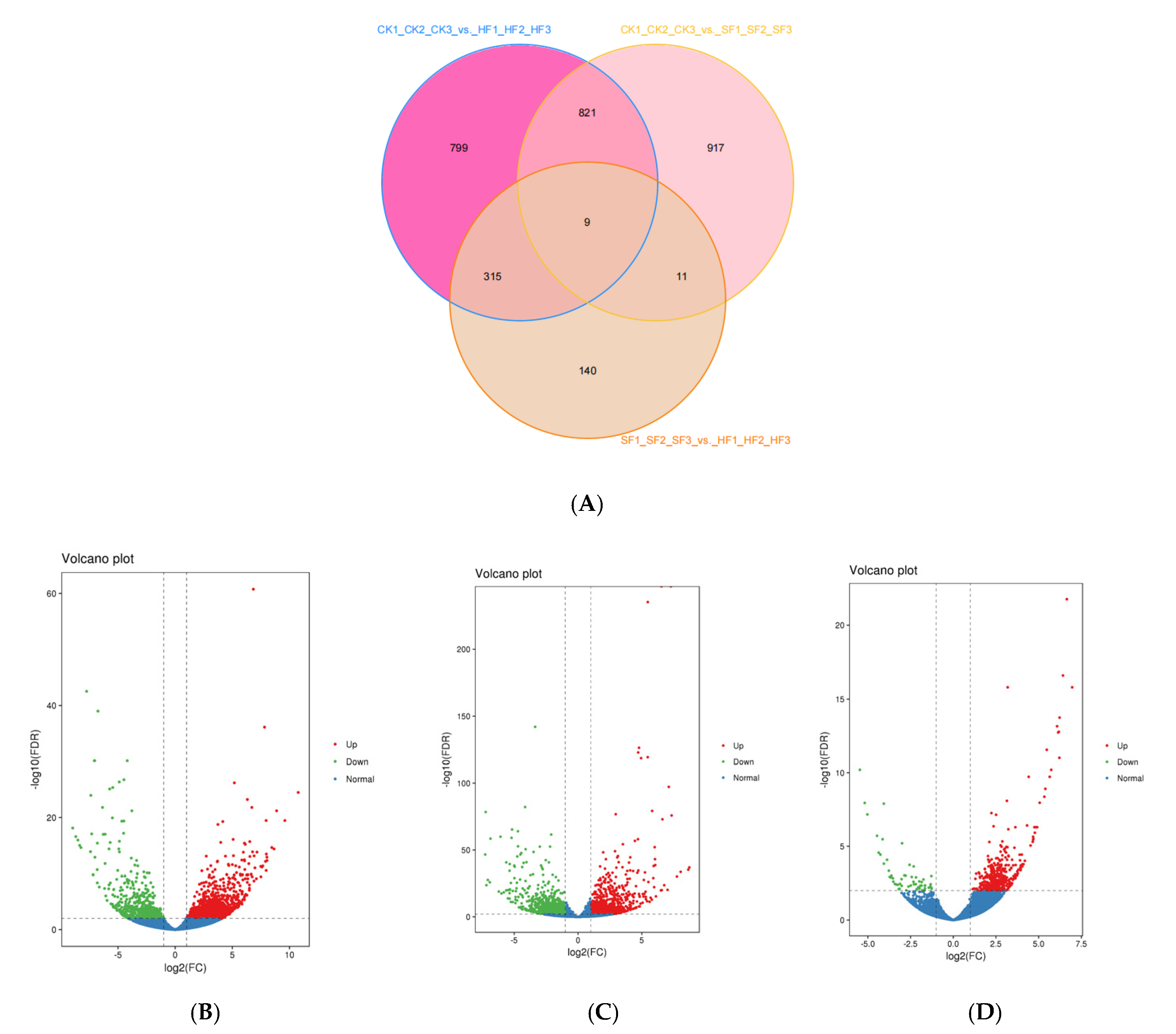
3.2. Transcriptome Sequencing Analysis
3.3. Identification of Differentially Expressed Genes (DEGs)
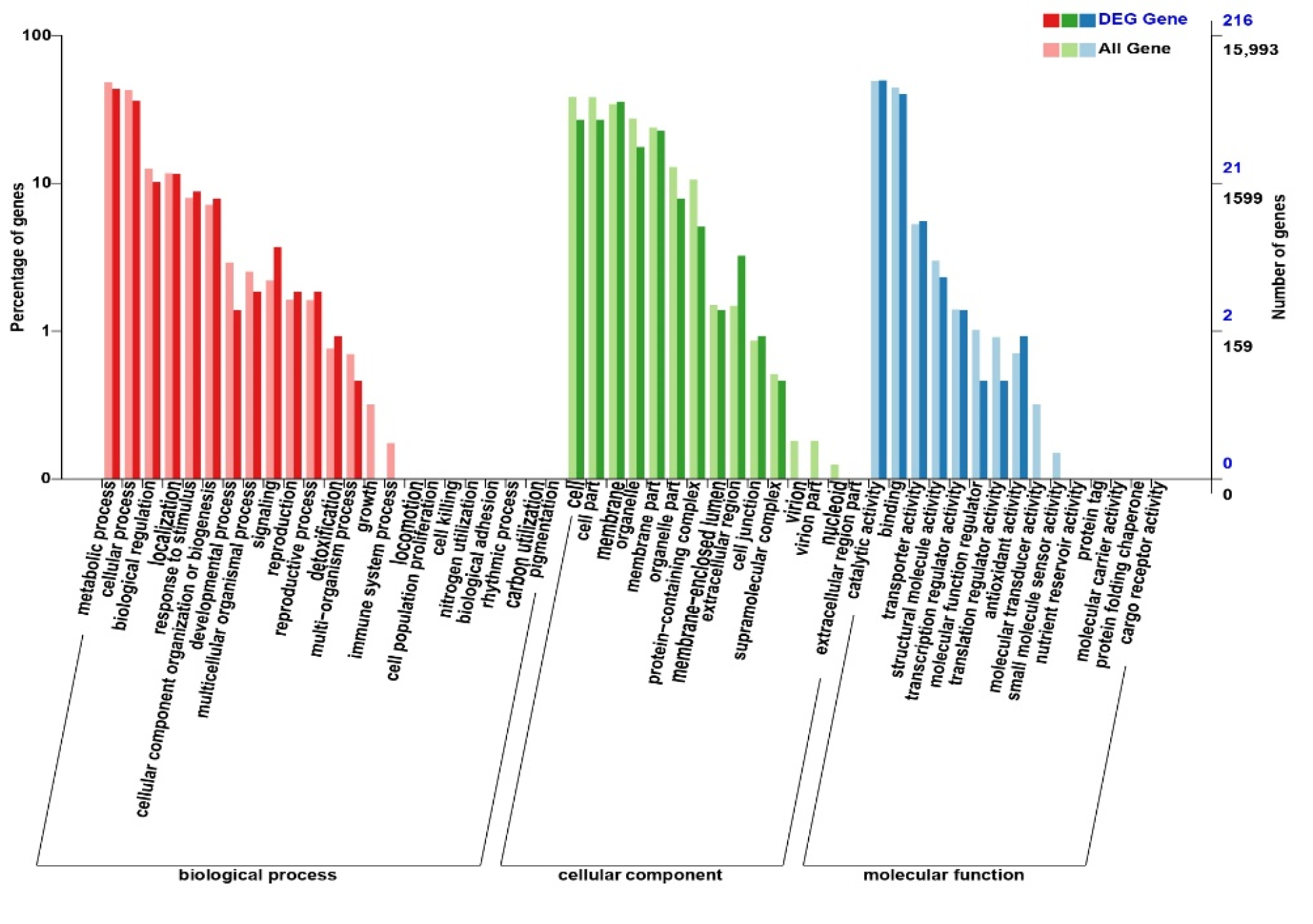
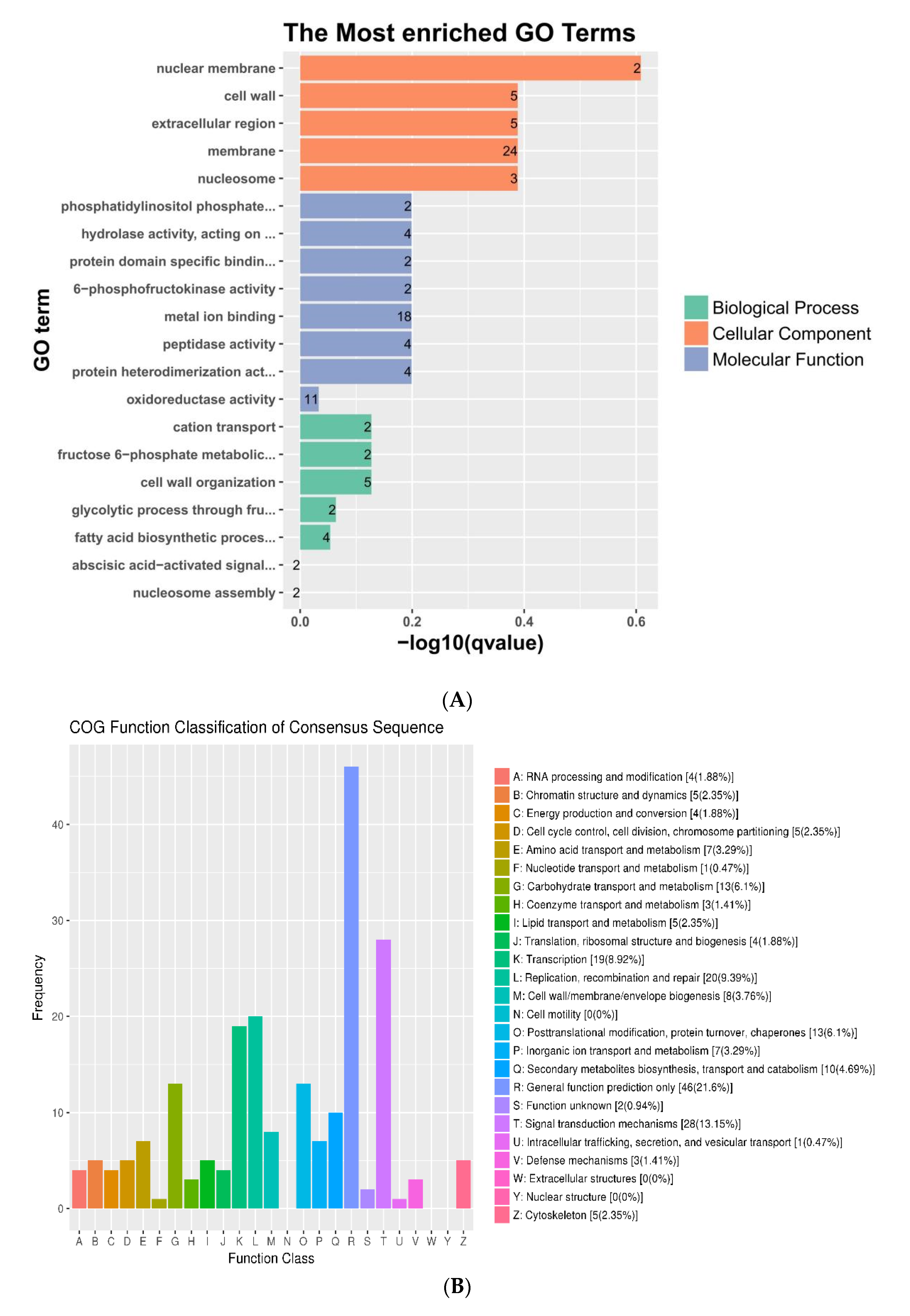
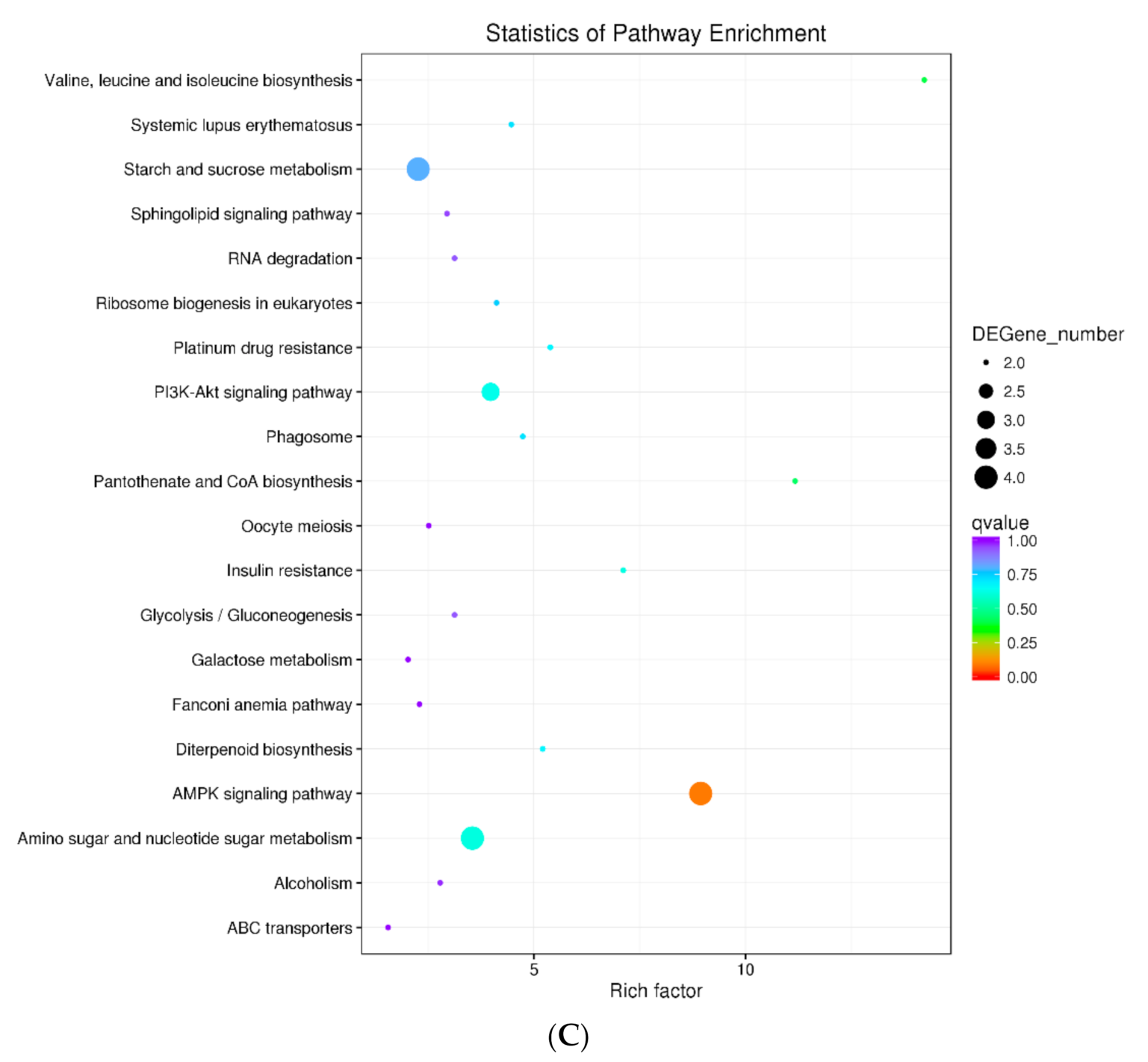
3.4. Functional Annotation and Enrichment of DEGS
3.5. Identification of Candidate Genes Associated with Self-Incompatibility
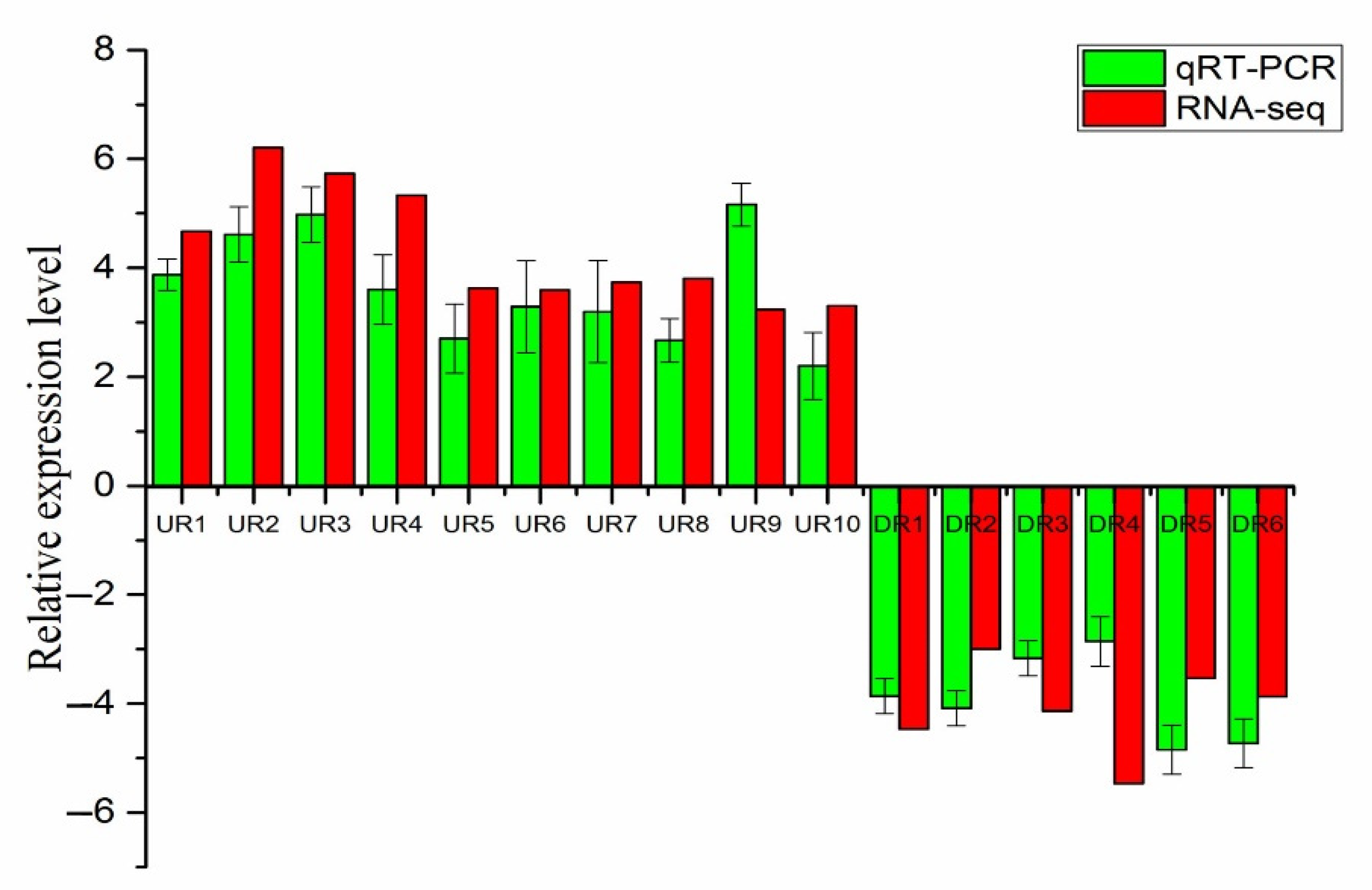
3.6. qRT-PCR Validate Analysis of DEGs
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Meng, L.Z.; Lv, G.P.; Hu, D.J.; Cheong, K.L.; Xie, J.; Zhao, J. Effects of polysaccharides from different species of Dendrobium (Shihu) on macrophage function. Molecules 2013, 18, 5779–5791. [Google Scholar] [CrossRef]
- Yan, L.; Wang, X.; Liu, H.; Tian, Y.; Lian, J.M.; Yang, R.J.; Hao, S.M.; Wang, X.J.; Yang, S.C.; Li, Q.Y.; et al. The genome of Dendrobium officinale illuminates the biology of the important traditional Chinese orchid herb. Mol. Plant 2015, 8, 922–934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, K.; Li, Y.; Tao, S.; Wei, G.; Huang, Y.; Chen, D.; Wu, C. Purification, Characterization and Biological Activity of Polysaccharides from Dendrobium officinale. Molecules 2016, 21, 701. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, Y.; Lyu, P.; Chen, L.; Shen, C.; Sun, C. Comparative transcriptomic analysis reveal the regulation mechanism underlying MeJA-induced accumulation of alkaloids in Dendrobium officinale. J. Plant Res 2019, 132, 419–429. [Google Scholar] [CrossRef]
- Huang, S.; Chen, F.; Cheng, H.; Huang, G. Modification and application of polysaccharide from traditional Chinese medicine such as Dendrobium officinale. Int. J. Biol. Macromol 2020, 157, 385–393. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Chen, S.; Hu, Y.; Yang, Y.; Yuan, J.; Wu, Y. Protective roles and mechanisms of Dendrobium officinal polysaccharides on secondary liver injury in acute colitis. Int. J. Biol. Macromol. 2018, 107, 2201–2210. [Google Scholar] [CrossRef] [PubMed]
- Yue, H.; Liu, Y.; Qu, H.; Ding, K. Structure analysis of a novel heteroxylan from the stem of Dendrobium officinale and anti-angiogenesis activities of its sulfated derivative. Int. J. Biol. Macromol. 2017, 103, 533–542. [Google Scholar] [CrossRef]
- Liu, J.; Zhu, T.; Niu, Q.; Yang, X.; Suo, H.; Zhang, H. Dendrobium nobile alkaloids protects against H2O2-induced neuronal injury by suppressing JAK-STATs pathway activation in N2A cells. Biol. Pharm. Bull. 2020, 43, 716–724. [Google Scholar] [CrossRef] [Green Version]
- Devadas, R.; Pattanayak, S.L.; Singh, D.R. Studies on cross compatibility in Dendrobium species and hybrids. Indian J. Genet. 2016, 76, 344–355. [Google Scholar]
- Zhang, G.Q.; Xu, Q.; Bian, C.; Tsai, W.C.; Yeh, C.M.; Liu, K.W.; Yoshida, K.; Zhang, L.S.; Chang, S.B.; Chen, F.; et al. The Dendrobium catenatum Lindl. genome sequence provides insights into polysaccharide synthase, floral development and adaptive evolution. Sci Rep. 2016, 6, 19029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, B.; Yuan, H.; Yu, Q.X.; Si, J.P. Pollen vigor and development of germplasm of Dendrobium officinale. Chin. J. Chin. Mater. Med. 2011, 36, 755–757. [Google Scholar]
- Bhattacharyya, P.; Kumaria, S. Molecular characterization of Dendrobium nobile Lindl., an endangered medicinal orchid, based on randomly amplified polymorphic DNA. Plant Syst. Evol. 2015, 301, 201–210. [Google Scholar] [CrossRef]
- Bhattacharyya, P.; Ghosh, S.; Mandi, S.S.; Kumaria, S.; Tandon, P. Genetic variability and association of AFLP markers with some important biochemical traits in Dendrobium thyrsiforum, a threatened medicinal orchid. S. Afr. J. Bot. 2017, 109, 214–222. [Google Scholar] [CrossRef]
- Yan, S.; Zhao, T.M.; Li, Y.F.; Hu, Y.D.; Chun, Z. Agronomic and quality traits of diferent Dendrobium nobile in Hejiang. Chin. J. Exp. Tradit Med. Formulae 2018, 24, 73–77. [Google Scholar]
- Igic, B.; Kohn, J.R. Evolutionary relationships among self-incompatibility RNases. Proc. Natl. Acad. Sci. USA 2001, 98, 13167–13171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doucet, J.; Lee, H.K.; Goring, D.R. Pollen acceptance or rejection: A tale of two pathways. Trends Plant Sci. 2016, 21, 1058–1067. [Google Scholar] [CrossRef]
- Mcclure, B.A.; Franklin-Tong, V. Gametophytic self-incompatibility: Understanding the cellular mechanisms involved in “self” pollen tube inhibition. Planta 2006, 224, 233–245. [Google Scholar] [CrossRef] [PubMed]
- Fujii, S.; Kubo, K.; Takayama, S. Non-self- and self-recognition models in plant self-incompatibility. Nat. Plants 2016, 2, 16130. [Google Scholar] [CrossRef] [PubMed]
- McClure, B.A.; Gray, J.E.; Anderson, M.A.; Clarke, A.E. Self-incompatibilityin Nicotiana alata involves degradation of pollen rRNA. Nature 1990, 347, 757–760. [Google Scholar] [CrossRef]
- de Graaf, B.H.J.; Rudd, J.J.; Wheeler, M.J.; Perry, R.M.; Bell, E.M.; Osman, K.; Franklin, F.C.H.; Franklin-Tong, V.E. Self-incompatibility in Papaver targets soluble inorganic pyrophosphatases in pollen. Nature 2006, 444, 490–493. [Google Scholar] [CrossRef] [PubMed]
- Samuel, M.A.; Chaal, B.K.; Lampard, G.; Green, B.R.; Ellis, B. Surviving the passage: Non-canonical stromal targeting of an Arabidopsis mitogen-activated protein kinase kinase. Plant Signal Behav. 2008, 3, 6–12. [Google Scholar] [CrossRef] [Green Version]
- Samuel, M.A.; Mudgil, Y.; Salt, J.N.; Delmas, F.; Ramachandran, S.; Chilelli, A.; Goring, D.R. Interactions between the S-Domain receptor kinases and AtPUB-ARM E3 ubiquitin ligases suggest a conserved signaling pathway in Arabidopsis. Plant Physiol. 2008, 147, 2084–2095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haasen, K.E.; Goring, D.R. The recognition and rejection of self-incompatible pollen in the Brassicaceae. Bot. Stud. 2010, 51, 1–6. [Google Scholar]
- Bower, M.S.; Matias, D.D.; Fernandes-Carvalho, E.; Mazzurco, M.; Gu, T.; Rothstein, S.J.; Goring, D.R. Two members of the thioredoxin-h family interact with the E. kinase domain of a Brassica S locus receptor kinase. Plant Cell 1996, 8, 1641–1650. [Google Scholar]
- Samuel, M.A.; Chong, Y.T.; Haasen, K.E.; Aldea-Brydges, M.G.; Stone, S.L.; Goring, D.R. Cellular pathways regulating responses to compatible and self-incompatible pollen in Brassica and Arabidopsis stigmas intersect at Exo70A1, a putative component of the exocyst complex. Plant Cell 2009, 21, 2655–2671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lao, X.; Suwabe, K.; Niikura, S.; Kakita, M.; Iwano, M.; Takayama, S. Physiological and genetic analysis of CO2-induced breakdown of self-incompatibility in Brassica rapa. J Exp Bot. 2014, 65, 939–951. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Yu, H.; Li, T.; Li, L.; Zhang, G.; Liu, Z.; Huang, T.; Zhang, Y. Comparative proteomics analyses of pollination response in endangered orchid species Dendrobium Chrysanthum. Int. J. Mol. Sci. 2017, 18, 2496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, J.k.; Zuo, J.H.; Huang, Y.H.; Li, C.S.; Chen, Y.L. The origin and germplasm collection for cultivated Dendrobium officinale K. Kimura & Migo individuals revealed by EST-SSR markers. Genet. Resour. Crop Evol. 2020, 67, 1209–1219. [Google Scholar]
- Tian, C.X.; Zhang, Y.M.; Ma, H.L. Primary study on preparation procedure of paraffin section with embryo structure of Poa pratensis. Pratacultural Sci. 2013, 30, 1980–1986. [Google Scholar]
- Trapnell, C.; Williams, B.; Pertea, G.; Mortazavi, A.; Kwan, G.; Baren, M.; Salzberg, S.; Wold, B.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef] [Green Version]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome biol. 2010, 11, R106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Cui, H.M.; Huang, T.H.; Liu, T.K.; Hou, X.L.; Li, Y. Identification and validation of reference genes for RT-qPCR analysis in non-heading Chinese cabbage flowers. Front. Plant Sci. 2016, 7, 811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Hansen, K.D.; Wu, Z.J.; Irizarry, R.A.; Leek, J.T. Sequencing technology does not eliminate biological variability. Nat Biotechnol. 2011, 29, 572–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulze, S.K.; Kanwar, R.; Golzenleuchter, M.; Therneau, T.M.; Beutler, A.S. SERE: Single-parameter quality control and sample comparison for RNA-Seq. BMC Genomics 2012, 13, 524. [Google Scholar] [CrossRef] [Green Version]
- Nie, S.S.; Zhang, M.J.; Zhang, L.G. Genome-wide identification and expression analysis of calmodulin-like (CML) genes in Chinese cabbage (Brassica rapa L. ssp. pekinensis). BMC Genomics 2017, 18, 482. [Google Scholar] [CrossRef]
- Wu, P.; Wang, W.; Duan, W.; Li, Y.; Hou, X. Comprehensive analysis of the CDPK-SnRK superfamily genes in Chinese cabbage and its evolutionary implications in plants. Front. Plant Sci. 2017, 8, 162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohanta, T.K.; Kumar, P.; Bae, H.H. Genomics and evolutionary aspect of calcium signaling event in calmodulin and calmodulin-like proteins in plants. BMC Plant Biol. 2017, 17, 38. [Google Scholar] [CrossRef] [Green Version]
- Rubinstein, A.L.; Broadwater, A.H.; Lowrey, K.B.; Bedinger, P.A. Pex1, a pollen-specific gene with an extensin-like domain. Proc. Natl. Acad. Sci. USA 1995, 92, 3086–3090. [Google Scholar] [CrossRef] [Green Version]
- Wan, J.; Patel, A.; Mathieu, M.; Kim, S.; Xu, D.; Stacey, G. A lectin receptor-like kinase is required for pollen development in Arabidopsis. Plant Mol. Biol. 2008, 67, 469–482. [Google Scholar] [CrossRef]
- Wheeler, M.J.; Graaf, B.H.J.; Hadjiosif, N.; Perry, R.M.; Poulter, N.S.; Osman, K.; Vatovec, S.; Harper, A.; Franklin, F.C.H.; Franklin-Tong, V.E. Identification of the pollen self-incompatibility determinant in Papaver rhoeas. Nature 2009, 459, 992–995. [Google Scholar] [CrossRef]
- Millner, H.J.; McCrea, A.R.; Baldwin, T.C. An investigation of self-incompatibility within the genus Restrepia. Am. J Bot. 2015, 102, 487–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Y.; Lin, R.; Yuan, C.; Ma, L.; Wang, J.; Zhu, Y.; Gao, Y. Cytological study on self and hybridization affinity in Dendrobium officinale. J. Zhejiang A F Univ. 2018, 35, 618–623. [Google Scholar]
- Miao, H.X.; Qin, Y.H.; Silva, J.A.T.; Ye, Z.X.; Hu, G.B. Identification of differentially expressed genes in pistils from self-incompatible Citrus reticulata by suppression subtractive hybridization. Mol. Biol. Rep. 2013, 40, 159–169. [Google Scholar] [CrossRef]
- Miao, H.X.; Qin, Y.H.; Ye, Z.X.; Hu, G.B. Molecular characterization and expression analysis of ubiquitin-activating enzyme E1 gene in Citrus reticulata. Genes 2013, 53, 249–259. [Google Scholar] [CrossRef]
- Zhang, W.; Wei, X.; Meng, H.L.; Ma, C.H.; Jiang, N.H.; Zhang, G.H.; Yang, S.C. Transcriptomic comparison of the self-pollinated and cross-pollinated flowers of Erigeron breviscapus to analyze candidate self-incompatibility-associated genes. BMC Plant Biol. 2015, 15, 248. [Google Scholar] [CrossRef] [Green Version]
- Jung, H.J.; Kayum, M.A.; Thamilarasan, S.K.; Nath, U.K.; Park, J.I.; Chung, M.Y.; Hur, Y.; Nou, I.S. Molecular characterisation and expression profiling of calcineurin B-like (CBL) genes in Chinese cabbage under abiotic stresses. Funct. Plant Biol. 2017, 44, 739. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, W.Z.; Song, L.F.; Zou, J.J.; Su, Z.; Wu, W.H. Transcriptome analyses show changes in gene expression to accompany pollen germination and tube growth in Arabidopsis. Plant Physiol. 2008, 148, 1201–1211. [Google Scholar] [CrossRef] [Green Version]
- Hua, Z.; Kao, T.H. Identification and characterization of components of a putative petunia S-locus F-box-containing E3 ligase complex involved in S-RNase-based self-incompatibility. Plant Cell 2006, 18, 2531–2553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vierstra, R.D. The ubiquitin-26S proteasome system at the nexus of plant biology. Nat. Rev. Mol. Cell Biol. 2009, 10, 385–397. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Fan, J.; Li, J.; Song, Y.; Li, Q.; Zhang, Y.; Xue, Y. SCF (SLF)-mediated cytosolic degradation of S-RNase is required for cross-pollen compatibility in S-RNase-based self-incompatibility in Petunia hybrida. Front. Genet. 2014, 5, 228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, H. Plant reproduction: GABA gradient, guidance and growth. Curr. Biol. 2003, 13, R834–R836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canovas, F.M.; Avila, C.; Canton, F.R.; Canas, R.A.; Torre, F. Ammonium assimilation and amino acid metabolism in conifers. J. Exp. Bot. 2007, 58, 2307–2318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palanivelu, R.; Brass, L.; Edlund, A.F.; Preuss, D. Pollen tube growth and guidance is regulated by POP2, an Arabidopsis gene that controls GABA levels. Cell 2003, 114, 47–59. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| #Sample ID | ReadSum | BaseSum | GC(%) | Q20(%) | Q30(%) |
|---|---|---|---|---|---|
| CK1 | 22,450,889 | 6,735,266,700 | 47.49% | 97.83% | 93.76% |
| CK2 | 21,276,069 | 6,382,820,700 | 47.25% | 97.14% | 92.14% |
| CK3 | 24,726,730 | 7,418,019,000 | 47.09% | 97.46% | 92.97% |
| CK average | 22,817,896 | 6,845,368,800 | 47.28% | 97.48% | 92.96% |
| HF1 | 28,969,506 | 8,690,851,800 | 52.62% | 97.29% | 92.85% |
| HF2 | 20,949,503 | 6,284,850,900 | 47.06% | 97.64% | 93.40% |
| HF3 | 21,063,503 | 6,319,050,900 | 46.80% | 97.74% | 93.63% |
| HF average | 23,660,837 | 7,098,251,200 | 48.83% | 97.56% | 93.29% |
| SF1 | 20,991,442 | 6,297,432,600 | 46.87% | 97.71% | 93.55% |
| SF2 | 27,714,690 | 8,314,407,000 | 46.88% | 97.60% | 93.33% |
| SF3 | 23,222,705 | 6,966,811,500 | 46.65% | 97.65% | 93.49% |
| SF average | 23,976,279 | 7,192,883,700 | 46.80% | 97.65% | 93.46% |
| Total average | 23,485,004 | 7,045,501,233 | 47.63% | 97.56% | 93.24% |
| Total | 211,365,037 | 63,409,511,100 |
| Sample ID | Total Read | Reads Mapped |
|---|---|---|
| CK1 | 44,901,778 | 33,273,053 (74.10%) |
| CK2 | 42,552,138 | 31,039,066 (72.94%) |
| CK3 | 49,453,460 | 34,715,952 (70.20%) |
| HF1 | 57,939,012 | 39,336,622 (67.89%) |
| HF2 | 41,899,006 | 29,965,533 (71.52%) |
| HF3 | 42,127,006 | 30,322,911 (71.98%) |
| SF1 | 41,982,884 | 30,649,680 (73.01%) |
| SF2 | 55,429,380 | 40,043,108 (72.24%) |
| SF3 | 46,445,410 | 33,240,722 (71.57%) |
| Total | 422,730,074 | 302,586,647 (71.58%) |
| DEG Set | All DEG | Up-Regulated | Down-Regulated |
|---|---|---|---|
| CK1_CK2_CK3_ vs._HF1_HF2_HF3 | 1944 | 1358 | 586 |
| CK1_CK2_CK3_ vs._SF1_SF2_SF3 | 1758 | 932 | 826 |
| SF1_SF2_SF3_ vs._HF1_HF2_HF3 | 475 | 417 | 58 |
| GO Classify | All Gene | DEG Gene | Ratio |
|---|---|---|---|
| extracellular region | 238 | 7 | 0.0294 |
| signaling | 352 | 8 | 0.0227 |
| antioxidant activity | 113 | 2 | 0.0177 |
| detoxification | 122 | 2 | 0.0164 |
| reproductive process | 259 | 4 | 0.0154 |
| reproduction | 261 | 4 | 0.0153 |
| response to stimulus | 1275 | 19 | 0.0149 |
| cellular component organization or biogenesis | 1146 | 17 | 0.0148 |
| cell junction | 138 | 2 | 0.0145 |
| transporter activity | 845 | 12 | 0.0142 |
| membrane | 5497 | 77 | 0.0140 |
| catalytic activity | 7876 | 107 | 0.0136 |
| Total gene | 15,993 | 216 | 0.0135 |
| transcription regulator activity | 224 | 3 | 0.0134 |
| localization | 1869 | 25 | 0.0134 |
| membrane part | 3812 | 49 | 0.0129 |
| membrane-enclosed lumen | 240 | 3 | 0.0125 |
| binding | 7089 | 87 | 0.0123 |
| supramolecular complex | 82 | 1 | 0.0122 |
| metabolic process | 7732 | 94 | 0.0122 |
| cellular process | 6833 | 78 | 0.0114 |
| biological regulation | 2010 | 22 | 0.0109 |
| structural molecule activity | 478 | 5 | 0.0105 |
| multicellular organismal process | 404 | 4 | 0.0099 |
| cell part | 6115 | 58 | 0.0095 |
| cell | 6127 | 58 | 0.0095 |
| multi-organism process | 112 | 1 | 0.0089 |
| organelle | 4386 | 38 | 0.0087 |
| organelle part | 2062 | 17 | 0.0082 |
| translation regulator activity | 146 | 1 | 0.0068 |
| protein-containing complex | 1693 | 11 | 0.0065 |
| developmental process | 466 | 3 | 0.0064 |
| molecular function regulator | 163 | 1 | 0.0061 |
| nucleoid | 20 | 0 | 0 |
| virion | 29 | 0 | 0 |
| extracellular region part | 11 | 0 | 0 |
| virion part | 29 | 0 | 0 |
| protein tag | 3 | 0 | 0 |
| cargo receptor activity | 1 | 0 | 0 |
| protein folding chaperone | 2 | 0 | 0 |
| nutrient reservoir activity | 9 | 0 | 0 |
| molecular transducer activity | 51 | 0 | 0 |
| molecular carrier activity | 3 | 0 | 0 |
| small molecule sensor activity | 24 | 0 | 0 |
| cell killing | 4 | 0 | 0 |
| immune system process | 28 | 0 | 0 |
| cell population proliferation | 5 | 0 | 0 |
| carbon utilization | 2 | 0 | 0 |
| nitrogen utilization | 4 | 0 | 0 |
| biological adhesion | 3 | 0 | 0 |
| growth | 51 | 0 | 0 |
| locomotion | 10 | 0 | 0 |
| pigmentation | 2 | 0 | 0 |
| rhythmic process | 3 | 0 | 0 |
| Gene Name | ID | Description | Regulated (log2FC) | References |
|---|---|---|---|---|
| DoCML11 | Dendrobium_GLEAN_10039171 | Calmodulin-like protein 11 | Up (3.59) | [36] |
| DoCML14 | Dendrobium_GLEAN_10057311 | Putative calcium-binding protein CML14 | Up (2.77) | [36] |
| DoCML10 | Dendrobium_GLEAN_10061210 | Probable calcium-binding protein CML10 | Up (2.61) | [36] |
| DoCML42 | Dendrobium_GLEAN_10078224 | Calcium-binding protein CML42 | Up (2.55) | [36] |
| DoCPK20 | Dendrobium_GLEAN_10069252 | Calcium-dependent protein kinase 20 | Up (2.17) | [37] |
| DoCBL7 | Dendrobium_GLEAN_10021981 | Calcineurin B-like protein 7 | Down (−1.67) | [38] |
| DoLRX4 | Dendrobium_GLEAN_10140109 | Pollen-specific leucine-rich repeat extensin-like protein 4 | Up (2.81) | [39] |
| DoL-RLKs1 | Dendrobium_GLEAN_10110992 | L-type lectin-domain containing receptor kinase IX.1 | Up (2.29) | [40] |
| DoARC12 | Dendrobium_GLEAN_10051179 | U-box domain-containing protein 12 | Up (2.35) | [23] |
| DoARC52 | Dendrobium_GLEAN_10116227 | U-box domain-containing protein 52 | Up (2.25) | [23] |
| DoARC15 | Dendrobium_GLEAN_10032857 | U-box domain-containing protein 15 | Up (2.18) | [23] |
| DoARC3 | Dendrobium_GLEAN_10017335 | U-box domain-containing protein 3 | Up (2.25) | [23] |
| DoSRK1 | Dendrobium_GLEAN_10067876 | Putative inactive leucine-rich repeat receptor-like protein kinase | Up (2.68) | [21,22] |
| DoSRK2 | Dendrobium_GLEAN_10005387 | CBL-interacting serine/threonine-protein kinase 12 | Up (2.43) | [21,22] |
| DoSRK3 | Dendrobium_GLEAN_10113311 | serine/threonine-protein kinase D6PKL2 | Up (2.78) | [21,22] |
| DoSRK4 | Dendrobium_GLEAN_10042125 | CBL-interacting serine/threonine-protein kinase 12 | Up (2.62) | [21,22] |
| DoSRK5 | Dendrobium_GLEAN_10128039 | serine/threonine-protein kinase BLUS1 | Up (2.13) | [21,22] |
| DoSRK6 | Dendrobium_GLEAN_10027995 | Putative serine/threonine-protein kinase | Up (2.03) | [21,22] |
| DoSRK7 | Dendrobium_GLEAN_10098521 | Serine/threonine-protein kinase PBS1 | Up (1.86) | [21,22] |
| DoSRK8 | Dendrobium_GLEAN_10000384 | CBL-interacting serine/threonine-protein kinase 12 | Up (2.43) | [21,22] |
| DoSRK9 | Dendrobium_GLEAN_10010756 | Leucine-rich repeat receptor-like serine/threonine-protein kinase | Up (2.06) | [21,22] |
| DoSRK10 | Dendrobium_GLEAN_10047407 | Receptor-like serine/threonine-protein kinase ALE2 | Down (−1.32) | [21,22] |
| DoSRK11 | Dendrobium_GLEAN_10044962 | Putative LRR receptor-like serine/threonine-protein kinase RLK | Up (2.41) | [21,22] |
| DoEXO1 | Dendrobium_GLEAN_10047257 | Exocyst complex component EXO70B1 | Up (2.68) | [24,25] |
| DoEXO2 | Dendrobium_GLEAN_10108122 | Exocyst complex component EXO70B1 | Up (2.83) | [24,25] |
| DoEBI1 | Dendrobium_GLEAN_10013195 | E3 ubiquitin-protein ligase WAV3 | Up (2.24) | [27] |
| DoEBI2 | Dendrobium_GLEAN_10030613 | E3 ubiquitin-protein ligase RGLG4 | Up (2.13) | [27] |
| DoEBI3 | Dendrobium_GLEAN_10106149 | E3 ubiquitin-protein ligase SPL2 | Up (1.62) | [27] |
| DoEBI4 | Dendrobium_GLEAN_10029117 | E3 ubiquitin-protein ligase KEG | Up (2.49) | [27] |
| DoEBI5 | Dendrobium_GLEAN_10062294 | Ubiquitin-like protein-NEDD8-like protein RUB3 | Up (3.09) | [27] |
| DoEBI6 | Dendrobium_GLEAN_10032857 | Ring-type E3 ubiquitin transferase | Up (2.18) | [27] |
| DoEBI7 | Dendrobium_GLEAN_10126475 | E3 ubiquitin-protein ligase XBAT31 | Up (2.07) | [27] |
| DoEBI8 | Dendrobium_GLEAN_10076237 | Probable BOI-related E3 ubiquitin-protein ligase 3 | Up (2.47) | [27] |
| DoEBI9 | Dendrobium_GLEAN_10069672 | BOI-related E3 ubiquitin-protein ligase 1 | Up (2.43) | [27] |
| DoFAT1 | Dendrobium_GLEAN_10055554 | omega-6 fatty acid desaturase | Up (2.34) | [27] |
| DoAMI1 | Dendrobium_GLEAN_10138041 | Serine carboxypeptidase-like 35 | Up (2.31) | [27] |
| DoAMI2 | Dendrobium_GLEAN_10118990 | Amino-acid permease BAT1 like | Down (−2.86) | [27] |
| DoAMI3 | Dendrobium_GLEAN_10033197 | Aspartokinase 1 | Up (1.57) | [27] |
| DoAMI4 | Dendrobium_GLEAN_10141386 | Arogenate dehydratase/prephenate dehydratase 2 | Up (2.21) | [27] |
| DoAMI5 | Dendrobium_GLEAN_10014762 | Probable polyamine transporter At3g19553 | Up (3.17) | [27] |
| DoAMI6 | Dendrobium_GLEAN_10056253 | amino acid transporter | Up (1.53) | [27] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.; Hu, B.; Zhang, F.; Luo, X.; Xie, J. Cytological Observation and Transcriptome Comparative Analysis of Self-Pollination and Cross-Pollination in Dendrobium Officinale. Genes 2021, 12, 432. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12030432
Chen Y, Hu B, Zhang F, Luo X, Xie J. Cytological Observation and Transcriptome Comparative Analysis of Self-Pollination and Cross-Pollination in Dendrobium Officinale. Genes. 2021; 12(3):432. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12030432
Chicago/Turabian StyleChen, Yaling, Benchang Hu, Fantao Zhang, Xiangdong Luo, and Jiankun Xie. 2021. "Cytological Observation and Transcriptome Comparative Analysis of Self-Pollination and Cross-Pollination in Dendrobium Officinale" Genes 12, no. 3: 432. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12030432