
Characterization of Two Ethephon-Induced IDA-Like Genes from Mango, and Elucidation of Their Involvement in Regulating Organ Abscission

 and
and 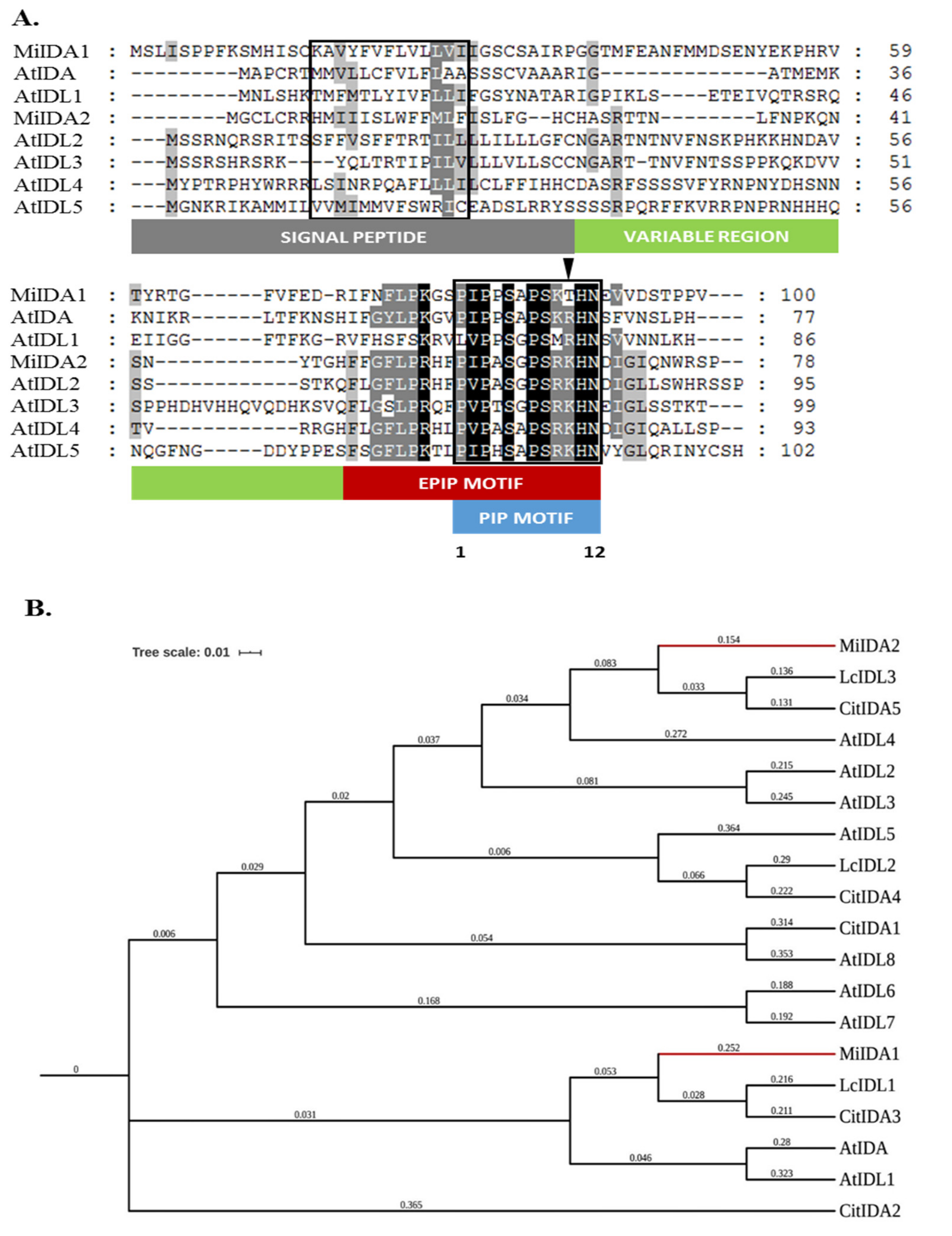{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
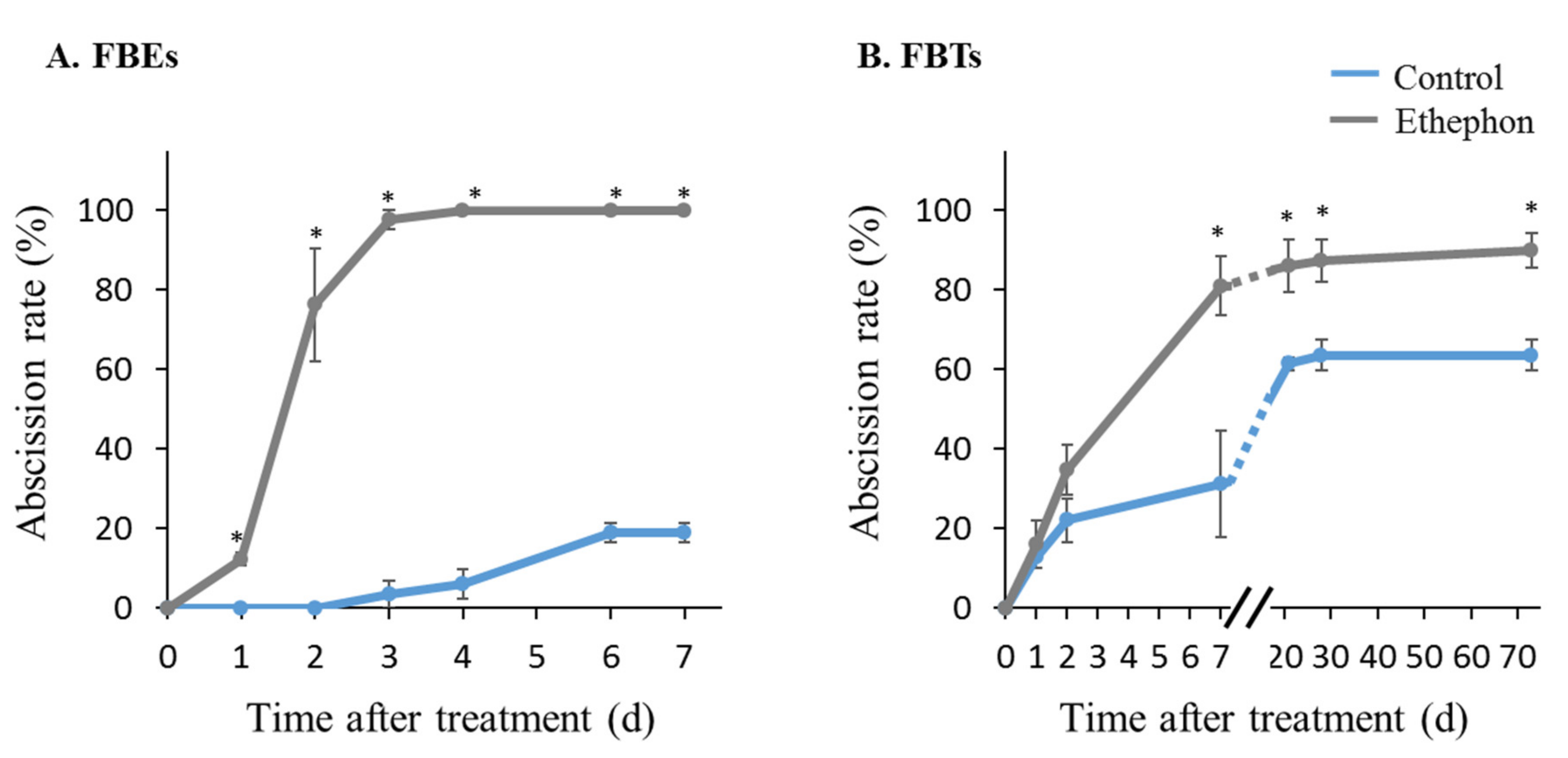
2.1. Plant Material, Induction of Fruitlet Abscission, and Fruitlet AZ Tissue Sampling
2.2. RNA Isolation and cDNA Synthesis
2.3. Cloning of MiIDA1 and MiIDA2 Genes
2.4. Selection of Mango Genes for Expression Analysis
2.5. Real-Time Quantitative (q) PCR Analysis
2.6. Arabidopsis Transformation and Phenotypic Analysis
2.7. BCECF Fluorescence Analysis
2.8. Scanning Electron Microscopy (SEM)
2.9. Statistical Analysis
3. Results
3.1. Identification and Isolation of MiIDA1 and MiIDA2 Transcripts from the Mango ‘Kent’ Cultivar
3.2. An Atypical PIPT-Type Version of ‘Kent’ MiIDA1 Protein Is Present in Various Mango Cultivars
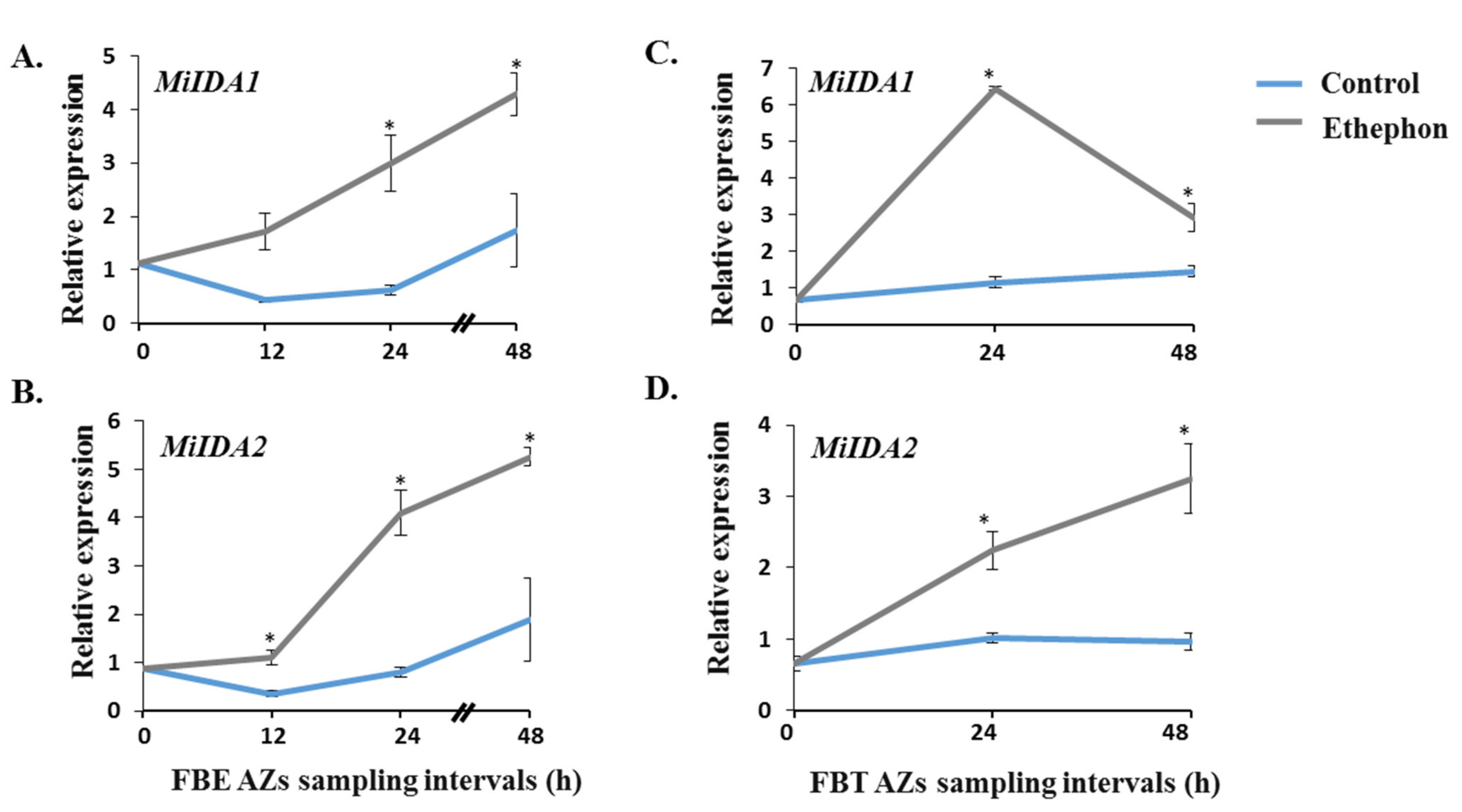
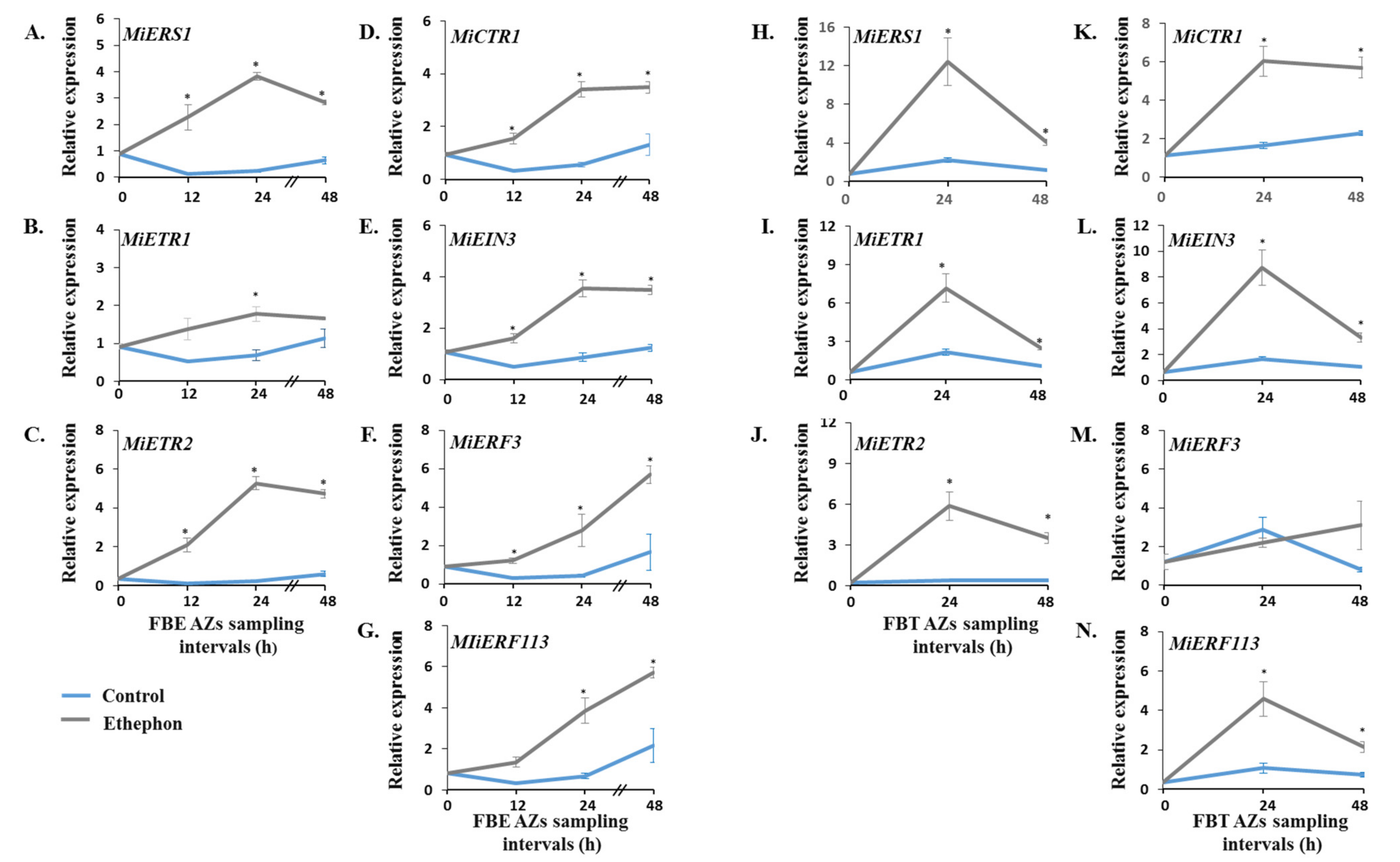
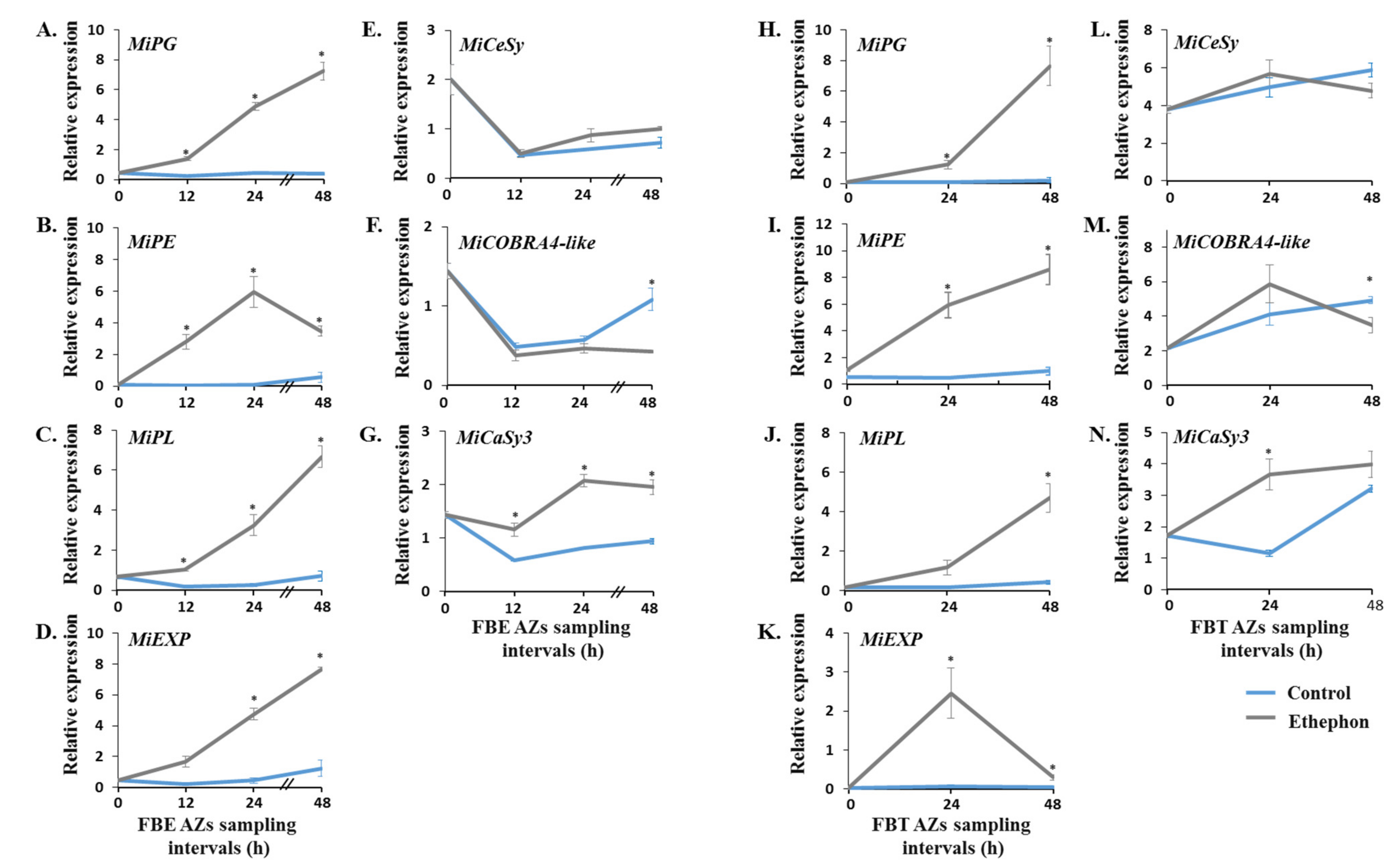
3.3. Monitoring the Effect of Ethephon Treatment on the Expression Patterns of MiIDA1 and MiIDA2, and Distinct Ethylene Signaling-Related Genes
3.4. Monitoring the Effect of Ethephon Treatment on the Expression Profiles of Genes Encoding for Distinct Cell Wall-Modifying Proteins
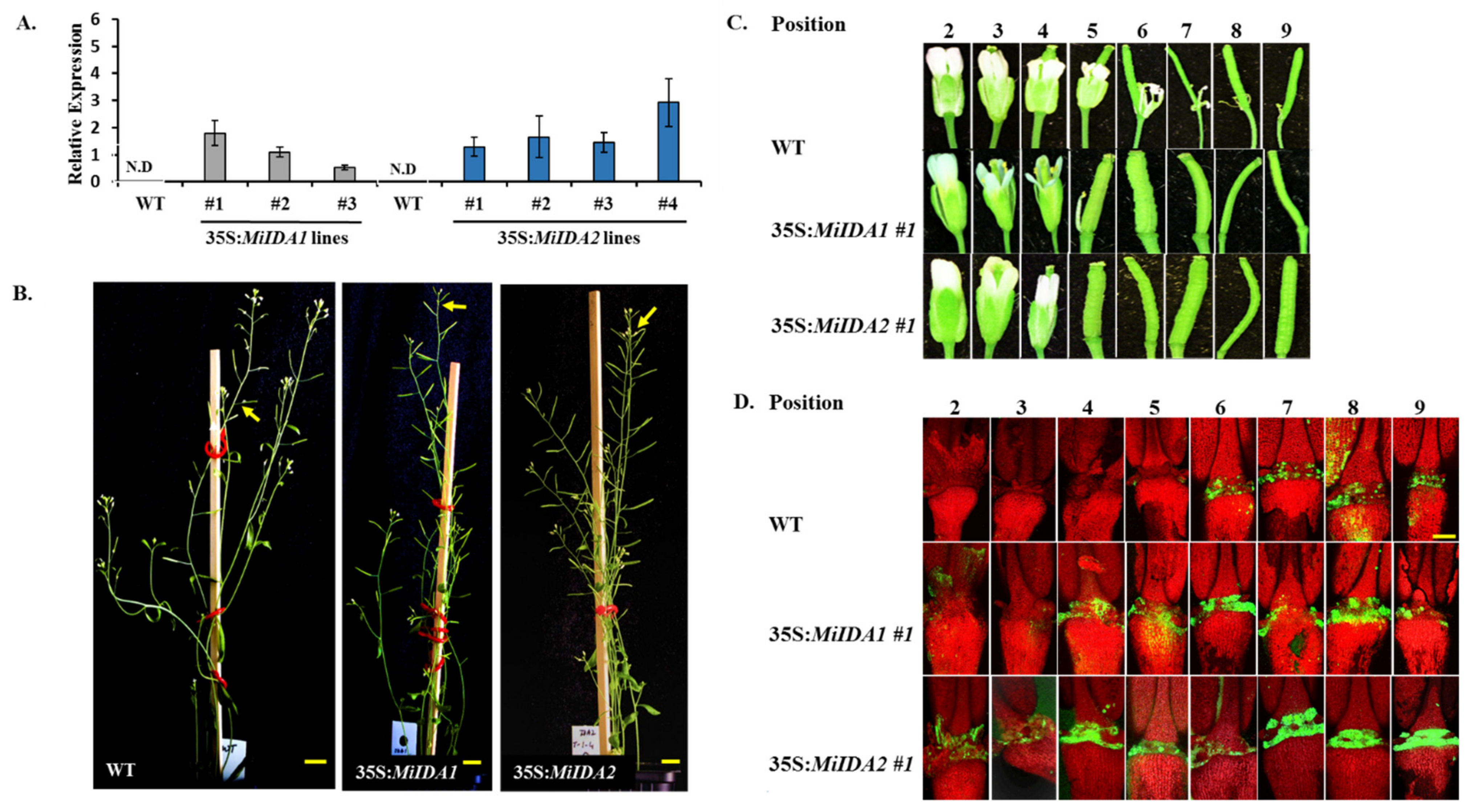
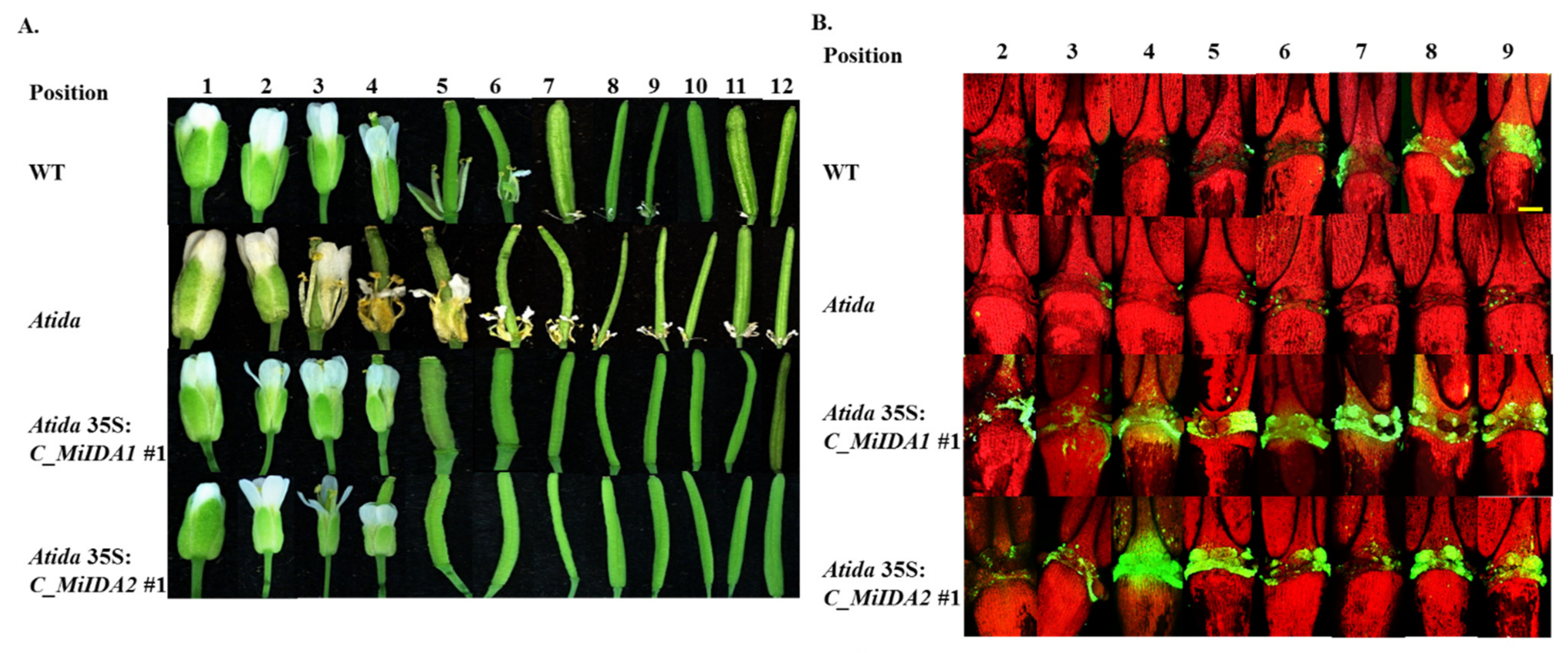
3.5. Overexpression of MiIDA1 and MiIDA2 in Arabidopsis Promoted Floral Organ Abscission, and Restored Floral Organ Abscission Ability in an Atida Mutant
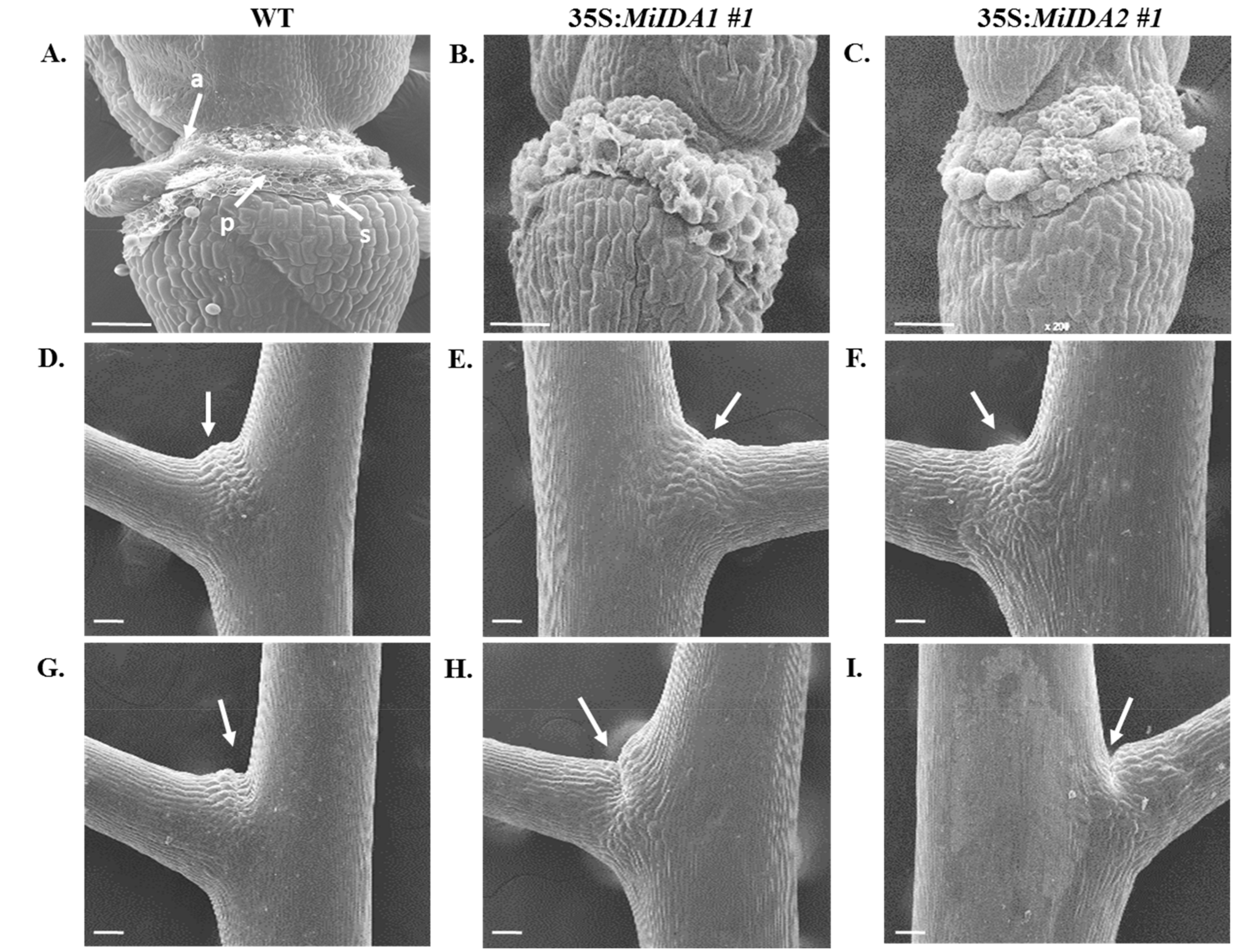
3.6. Overexpression of MiIDA1 and MiIDA2 in Arabidopsis Is also Associated with Activation of Cell Separation at the Base of Pedicel AZs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bonghi, C.; Ramina, A. Biochemical and molecular aspects of fruitlet abscission. Plant Growth Regul. 2000, 31, 35–42. [Google Scholar] [CrossRef]
- Estornell, L.H.; Agusti, J.; Merelo, P.; Talon, M.; Tadeo, F.R. Elucidating mechanisms underlying organ abscission. Plant Sci. 2013, 199, 48–60. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Sundaresan, S.; Philosoph-Hadas, S.; Yang, R.; Meir, S.; Tucker, M.L. Examination of the abscission-associated transcriptomes for soybean, tomato, and Arabidopsis highlights the conserved biosynthesis of an extensible extracellular matrix and boundary layer. Front. Plant Sci. 2015, 6, 1109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J. Four shades of detachment: Regulation of floral organ abscission. Plant Signal Behav. 2014, 9, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Patharkar, O.R.; Walker, J.C. Advances in abscission signaling. J. Exp. Bot. 2018, 69, 733–740. [Google Scholar] [CrossRef] [PubMed]
- Sawicki, M.; Aït Barka, E.; Clément, C.; Vaillant-Gaveau, N.; Jacquard, C. Cross-talk between environmental stresses and plant metabolism during reproductive organ abscission. J. Exp. Bot. 2015, 66, 1707–1719. [Google Scholar] [CrossRef] [Green Version]
- Meir, S.; Sundaresan, S.; Riov, J.; Agarwal, I.; Philosoph-Hadas, S. Role of auxin depletion in abscission control. Stewart Postharvest Rev. 2015, 11, 1–15. [Google Scholar]
- Basu, M.M.; Gonzalez-Carranza, Z.H.; Azam-Ali, S.; Tang, S.Y.; Shahid, A.A.; Roberts, J.A. The manipulation of auxin in the abscission zone cells of Arabidopsis flowers reveals that indoleacetic acid signaling is a prerequisite for organ shedding. Plant Physiol. 2013, 162, 96–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butenko, M.A.; Patterson, S.E.; Grini, P.E.; Stenvik, G.; Amundsen, S.S.; Mandal, A.; Aalen, R.B. INFLORESCENCE DEFICIENT IN ABSCISSION controls floral organ abscission in Arabidopsis and identifies a novel family of putative ligands in plants. Plant Cell 2003, 15, 2296–2307. [Google Scholar] [CrossRef] [Green Version]
- Stenvik, G.E.; Butenko, M.A.; Urbanowicz, B.R.; Rose, J.K.C.; Aalen, R.B. Overexpression of INFLORESCENCE DEFICIENT IN ABSCISSION activates cell separation in vestigial abcission zones in Arabidopsis. Plant Cell 2006, 18, 1467–1476. [Google Scholar] [CrossRef] [Green Version]
- Jinn, T.L.; Stone, J.M.; Walker, J.C. HAESA, an Arabidopsis leucine-rich repeat receptor kinase, controls floral organ abscission. Genes Dev. 2000, 14, 108–117. [Google Scholar]
- Sung, K.C.; Larue, C.T.; Chevalier, D.; Wang, H.; Jinn, T.L.; Zhang, S.; Walker, J.C. Regulation of floral organ abscission in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2008, 105, 15629–15634. [Google Scholar]
- Niederhuth, C.E.; Cho, S.K.; Seitz, K.; Walker, J.C. Letting go is never easy: Abscission and receptor-like protein kinases. J. Integr. Plant Biol. 2013, 55, 1251–1263. [Google Scholar] [CrossRef] [PubMed]
- Butenko, M.A.; Stenvik, G.E.; Alm, V.; Sæther, B.; Patterson, S.E.; Aalen, R.B. Ethylene-dependent and -independent pathways controlling floral abscission are revealed to converge using promoter::reporter gene constructs in the ida abscission mutant. J. Exp. Bot. 2006, 57, 3627–3637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Botton, A.; Ruperti, B. The yes and no of the ethylene involvement in abscission. Plants 2019, 8, 187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meir, S.; Philosoph-Hadas, S.; Riov, J.; Tucker, M.L.; Patterson, S.E.; Roberts, J.A. Re-evaluation of the ethylene-dependent and-independent pathways in the regulation of floral and organ abscission. J. Exp. Bot. 2019, 70, 1461–1467. [Google Scholar] [CrossRef]
- Vie, A.K.; Najafi, J.; Liu, B.; Winge, P.; Butenko, M.A.; Hornslien, K.S.; Kumpf, R.; Aalen, R.B.; Bones, A.M.; Brembu, T. The IDA/IDA-LIKE and PIP/PIP-LIKE gene families in Arabidopsis: Phylogenetic relationship, expression patterns, and transcriptional effect of the PIPL3 peptide. J. Exp. Bot. 2015, 66, 5351–5365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stenvik, G.E.; Tandstad, N.M.; Guo, Y.; Shi, C.L.; Kristiansen, W.; Holmgren, A.; Clark, S.E.; Aalen, R.B.; Butenko, M.A. The EPIP peptide of inflorescence deficient in abscission is sufficient to induce abscission in Arabidopsis through the receptor-like kinases HAESA and HAESA-LIKE2. Plant Cell 2008, 20, 1805–1817. [Google Scholar] [CrossRef] [Green Version]
- Stø, I.M.; Orr, R.J.S.; Fooyontphanich, K.; Jin, X.; Knutsen, J.M.B.; Fischer, U.; Tranbarger, T.J.; Nordal, I.; Aalen, R. Conservation of the abscission signaling peptide IDA during Angiosperm evolution: Withstanding genome duplications and gain and loss of the receptors HAE/HSL2. Front. Plant Sci. 2015, 6, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Shi, C.L.; Alling, R.M.; Hammerstad, M.; Aalen, R.B. Control of organ abscission and other cell separation processes by evolutionary conserved peptide signaling. Plants 2019, 8, 225. [Google Scholar] [CrossRef] [Green Version]
- Butenko, M.A.; Wildhagen, M.; Albert, M.; Jehle, A.; Kalbacher, H.; Aalen, R.B.; Felix, G. Tools and strategies to match peptide-ligand receptor pairs. Plant Cell 2014, 26, 1838–1847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tucker, M.L.; Yang, R. IDA-like gene expression in soybean and tomato leaf abscission and requirement for a diffusible stelar abscission signal. AoB Plants 2012, pls035. [Google Scholar] [CrossRef] [Green Version]
- Estornell, L.H.; Wildhagen, M.; Pérez-Amador, M.A.; Talón, M.; Tadeo, F.R.; Butenko, M.A. The IDA peptide controls abscission in Arabidopsis and citrus. Front. Plant Sci. 2015, 6, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ying, P.; Li, C.; Liu, X.; Xia, R.; Zhao, M.; Li, J. Identification and molecular characterization of an IDA-like gene from lychee, LcIDL1, whose ectopic expression promotes floral organ abscission in Arabidopsis. Sci. Rep. 2016, 6, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tranbarger, T.J.; Domonhédo, H.; Cazemajor, M.; Dubreuil, C.; Fischer, U.; Morcillo, F. The PIP peptide of INFLORESCENCE DEFICIENT IN ABSCISSION enhances Populus leaf and Elaeis guineensis fruit abscission. Plants 2019, 8, 143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilmowicz, E.; Kućko, A.; Ostrowski, M.; Panek, K. INFLORESCENCE DEFICIENT IN ABSCISSION-like is an abscission-associated and phytohormone-regulated gene in flower separation of Lupinus luteus. Plant Growth Regul. 2018, 85, 91–100. [Google Scholar] [CrossRef] [Green Version]
- Ventimilla, D.; Domingo, C.; González-Ibeas, D.; Talon, M.; Tadeo, F.R. Differential expression of IDA (INFLORESCENCE DEFICIENT IN ABSCISSION)-like genes in Nicotiana benthamiana during corolla abscission, stem growth and water stress. BMC Plant Biol. 2020, 20, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Litz, R.E. The Mango: Botany, Production and Uses, 2nd ed.; CABI: Oxfordshire, UK, 2009; pp. 1–680. [Google Scholar]
- Singh, Z.; Malik, A.U.; Davenport, T.L. Fruit drop in mango. Hortic Rev. (Am. Soc. Hortic. Sci.) 2005, 31, 111–153. [Google Scholar]
- Nunez-Elisea, R.; Davenport, T.L. Abscission of mango fruitlets as influenced by enhanced ethylene biosynthesis. Plant Physiol. 1986, 82, 991–994. [Google Scholar] [CrossRef] [Green Version]
- Ish-Shalom, M.; Dahan, Y.; Maayan, I.; Irihimovitch, V. Cloning and molecular characterization of an ethylene receptor gene, MiERS1, expressed during mango fruitlet abscission and fruit ripening. Plant Physiol. Bioch. 2011, 49, 931–936. [Google Scholar] [CrossRef]
- Hagemann, M.H.; Winterhagen, P.; Hegele, M.; Wunsche, J.N. Ethephon induced abscission in mango: Physiological fruitlet responses. Front. Plant Sci. 2015, 6, 706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winterhagen, P.; Hagemann, M.H.; Wuensche, J.N. Expression and interaction of the mango ethylene receptor MiETR1 and different receptor versions of MiERS1. Plant Sci. 2016, 246, 26–36. [Google Scholar] [CrossRef] [PubMed]
- Denisov, Y.; Glick, S.; Zviran, T.; Ish-Shalom, M.; Levin, A.; Faigenboim, A.; Cohen, Y.; Irihimovitch, V. Distinct organ-specific and temporal expression profiles of auxin-related genes during mango fruitlet drop. Plant Physiol. Bioch. 2017, 115, 439–448. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.; Puryear, J.; Cairney, J. A simple and efficient method for isolating RNA from pine trees. Plant Mol. Biol. Repo. 1993, 11, 113–116. [Google Scholar] [CrossRef]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [Green Version]
- Sundaresan, S.; Philosoph-Hadas, S.; Riov, J.; Belausov, E.; Kochanek, B.; Tucker, M.L.; Meir, S. Abscission of flowers and floral organs is closely associated with alkalization of the cytosol in abscission zone cells. J. Exp. Bot. 2015, 66, 1355–1368. [Google Scholar] [CrossRef] [Green Version]
- Hua, J.; Meyerowitz, E.M. Ethylene responses are negatively regulated by a receptor gene family in Arabidopsis thaliana. Cell 1998, 94, 261–271. [Google Scholar] [CrossRef] [Green Version]
- Mata, C.I.; Fabre, B.; Parsons, H.T.; Hertog, M.L.A.T.M.; Van Raemdonck, G.; Baggerman, G.; Van de Poel, B.; Lilley, K.S.; Nicolaï, B.M. Ethylene receptors, CTRS and EIN2 target protein identification and quantification through parallel reaction monitoring during tomato fruit ripening. Front. Plant Sci. 2018, 9, 1626. [Google Scholar] [CrossRef]
- Chen, Y.F.; Etheridge, N.; Schaller, G.E. Ethylene signal transduction. Ann. Bot. London 2005, 95, 901–915. [Google Scholar] [CrossRef]
- Niederhuth, C.E.; Patharkar, O.R.; Walker, J.C. Transcriptional profiling of the Arabidopsis abscission mutant hae hsl2 by RNA-Seq. BMC Genom. 2013, 17, 14–37. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Butenko, M.A.; Shi, C.-L.; Bolivar, J.L.; Winge, P.; Stenvik, G.-E.; Vie, A.K.; Leslie, M.E.; Brembu, T.; Kristiansen, W.; et al. NEVERSHED and INFLORESCENCE DEFICIENT IN ABSCISSION are differentially required for cell expansion and cell separation during floral organ abscission in Arabidopsis thaliana. J. Exp. Bot. 2013, 64, 5345–5357. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Chun, J.P.; Tucker, M.L. Transcriptional regulation of abscission zones. Plants 2019, 8, 154. [Google Scholar] [CrossRef] [Green Version]
- Santiago, J.; Brandt, B.; Wildhagen, M.; Hohmann, U.; Hothorn, L.A.; Butenko, M.A.; Hothorn, M. Mechanistic insight into a peptide hormone signaling complex mediating floral organ abscission. eLife 2016, 5, e15075. [Google Scholar] [CrossRef] [PubMed]
- Braidwood, L.; Breuer, C.; Sugimoto, K. My body is a cage: Mechanisms and modulation of plant cell growth. N. Phytol. 2014, 201, 388–402. [Google Scholar] [CrossRef] [PubMed]
- Hyung-Taeg, C.; Cosgrove, D.J. Altered expression of expansin modulates leaf growth and pedicel abscission in Arabidopsis thaliana. Proc. Nat. Acad. Sci. USA 2000, 97, 9783–9788. [Google Scholar]
- Wang, R.; Li, R.; Cheng, L.; Wang, X.; Fu, X.; Dong, X.; Mingfang, Q.M.; Jiang, C.-Z.; Xu, T.; Li, T. SlERF52 regulates SlTIP1;1 expression to accelerate tomato pedicel abscission. Plant Physiol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Bally, I.S.E.; Bombarely, A.; Chambers, A.H.; Cohen, Y.; Dillon, N.L.; Innes, D.J.; Islas-Osuna, M.A.; Kuhn, D.N.; Mueller, L.A.; Ophir, R.; et al. The ‘Tommy Atkins’ mango genome reveals candidate genes for fruit quality. BMC Plant Biol. 2021, 21, 108. [Google Scholar] [CrossRef]
- Wang, P.; Luo, Y.; Huang, J.; Gao, S.; Zhu, G.; Dang, Z.; Gai, J.; Yang, M.; Zhu, M.; Zhang, H.; et al. The genome evolution and domestication of tropical fruit mango. Genome Biol. 2020, 21, 60. [Google Scholar] [CrossRef] [Green Version]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rai, A.C.; Halon, E.; Zemach, H.; Zviran, T.; Sisai, I.; Philosoph-Hadas, S.; Meir, S.; Cohen, Y.; Irihimovitch, V. Characterization of Two Ethephon-Induced IDA-Like Genes from Mango, and Elucidation of Their Involvement in Regulating Organ Abscission. Genes 2021, 12, 439. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12030439
Rai AC, Halon E, Zemach H, Zviran T, Sisai I, Philosoph-Hadas S, Meir S, Cohen Y, Irihimovitch V. Characterization of Two Ethephon-Induced IDA-Like Genes from Mango, and Elucidation of Their Involvement in Regulating Organ Abscission. Genes. 2021; 12(3):439. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12030439
Chicago/Turabian StyleRai, Avinash Chandra, Eyal Halon, Hanita Zemach, Tali Zviran, Isaac Sisai, Sonia Philosoph-Hadas, Shimon Meir, Yuval Cohen, and Vered Irihimovitch. 2021. "Characterization of Two Ethephon-Induced IDA-Like Genes from Mango, and Elucidation of Their Involvement in Regulating Organ Abscission" Genes 12, no. 3: 439. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12030439