
Characteristics of the Diploid, Triploid, and Tetraploid Versions of a Cannabigerol-Dominant F1 Hybrid Industrial Hemp Cultivar, Cannabis sativa ‘Stem Cell CBG’

 and
and
Abstract
:1. Introduction
2. Materials and Methods
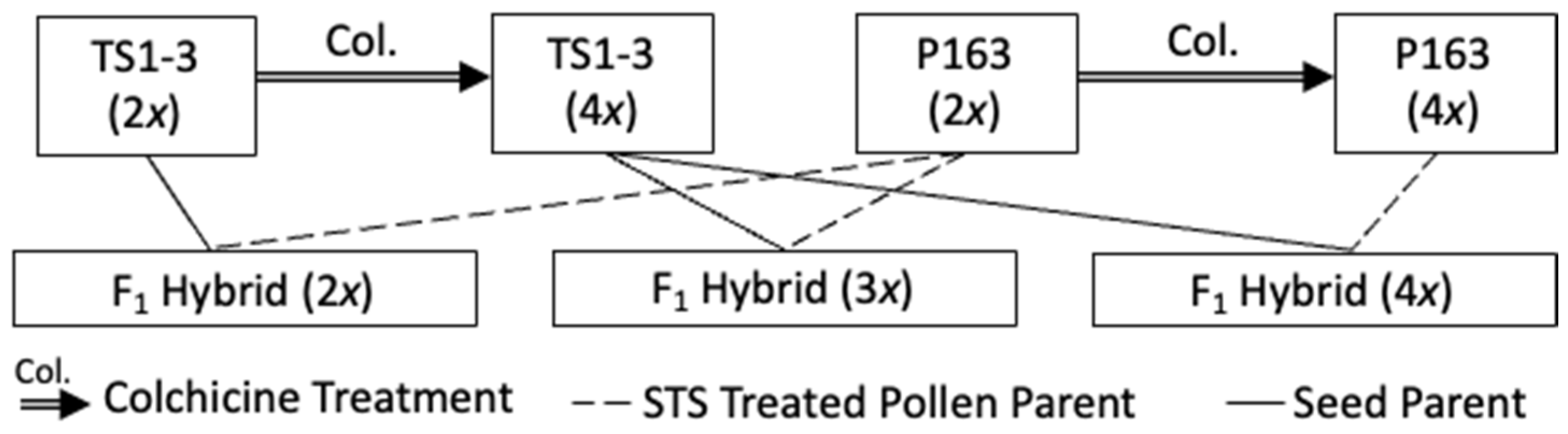
2.1. Plant Material
2.2. Fertility and Compatibility Experiments
2.3. Indoor Phenotyping Trials
2.4. Field Trials
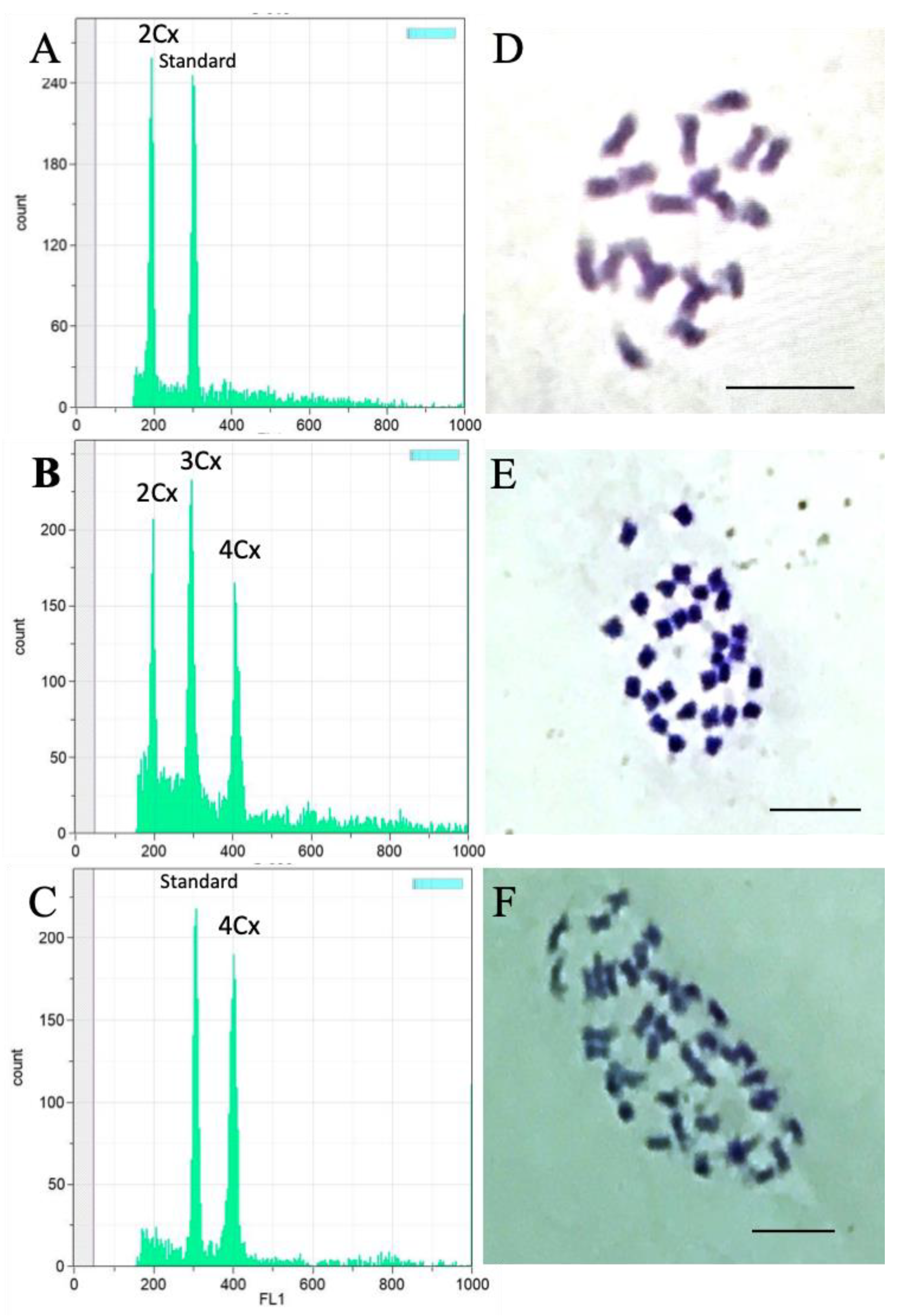
2.5. Flow Cytometry
2.6. Chromosome Squashes
3. Results
3.1. Ploidy Estimation
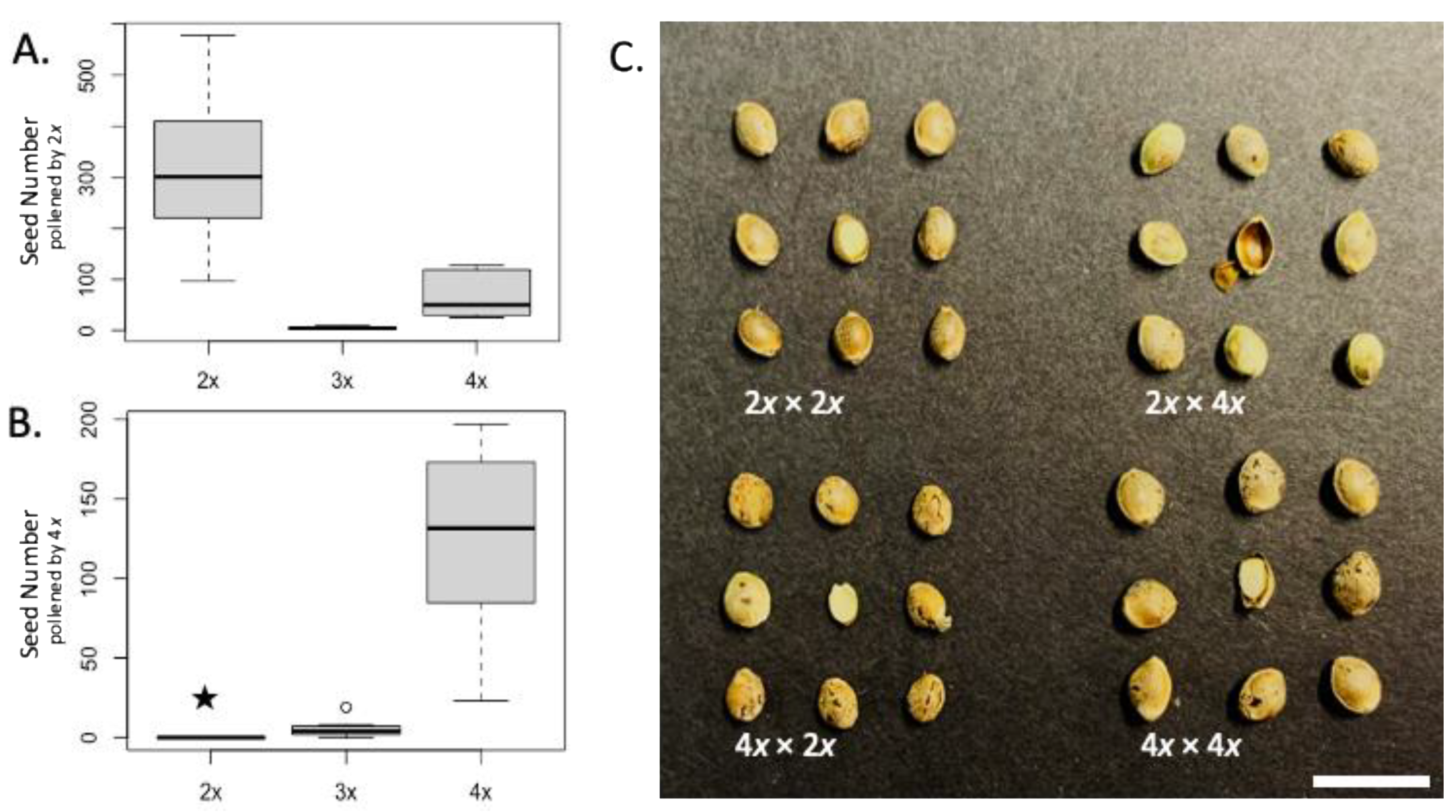
3.2. Fertility and Crossing Compatibility
3.3. Biomass and Inflorescence Weights and Plant Architecture
3.4. Cannabinoid Yield
4. Discussion
4.1. Fertility, Infertility, and Crossing Compatibility
4.2. Cannabinoid and Biomass Yield
4.3. Future Directions in Hemp Polyploid Manipulations
Author Contributions
Funding
Conflicts of Interest
References
- Henry, P.; Khatodia, S.; Kapoor, K.; Gonzales, B.; Middleton, A.; Hong, K.; Hilyard, A.; Johnson, S.; Allen, D.; Chester, Z. A single nucleotide polymorphism assay sheds light on the extent and distribution of genetic diversity, population structure and functional basis of key traits in cultivated north American cannabis. J. Cannabis Res. 2020, 2, 26. [Google Scholar] [CrossRef]
- McPartland, J.M. Cannabis systematics at the levels of family, genus, and species. Cannabis Cannabinoid Res. 2018, 3, 203–212. [Google Scholar] [CrossRef] [Green Version]
- Small, E. Evolution and classification of Cannabis sativa (Marijuana, Hemp) in relation to human utilization. Bot. Rev. 2015, 81, 189–294. [Google Scholar] [CrossRef]
- Sakamoto, K.; Akiyama, Y.; Fukui, K.; Kamada, H.; Satoh, S. Characterization; genome sizes and morphology of sex chromosomes in hemp (Cannabis sativa L.). Cytologia 1998, 63, 459–464. [Google Scholar] [CrossRef] [Green Version]
- Pate, D.W. Chemical ecology of Cannabis. J. Int. Hemp Assoc. 1994, 2, 32–37. [Google Scholar]
- Meier, C.; Mediavilla, V. Factors influencing the yield and the quality of hemp (Cannabis sativa L.) essential oil. J. Int. Hemp. Assoc. 1998, 5, 16–20. [Google Scholar]
- Toth, J.A.; Stack, G.M.; Cala, A.R.; Carlson, C.H.; Wilk, R.L.; Crawford, J.L.; Viands, D.R.; Philippe, G.; Smart, C.D.; Rose, J.K. Development and validation of genetic markers for sex and cannabinoid chemotype in Cannabis sativa L. Gcb. Bioenergy 2020, 12, 213–222. [Google Scholar] [CrossRef] [Green Version]
- Razumova, O.V.; Alexandrov, O.S.; Divashuk, M.G.; Sukhorada, T.I.; Karlov, G.I. Molecular cytogenetic analysis of monoecious hemp (Cannabis sativa L.) cultivars reveals its karyotype variations and sex chromosomes constitution. Protoplasma 2016, 253, 895–901. [Google Scholar] [CrossRef] [PubMed]
- Contreras, R.N.; Hoskins, T.C. Developing triploid maples. Horticulturae 2020, 6, 70. [Google Scholar] [CrossRef]
- Trojak-Goluch, A.; Skomra, U. Ploidy variation and agronomic performance of F1 hybrids of tetraploid and diploid forms of Humulus lupulus L. Breed. Sci. 2020, 70, 19102. [Google Scholar] [CrossRef] [Green Version]
- Trueblood, C.E.; Ranney, T.G.; Lynch, N.P.; Neal, J.C.; Olsen, R.T. Evaluating Fertility of triploid clones of Hypericum androsaemum L. for use as non-invasive landscape plants. HortScience 2010, 45, 1026–1028. [Google Scholar] [CrossRef]
- Rounsaville, T.J.; Touchell, D.H.; Ranney, T.G. Fertility and reproductive pathways in diploid and triploid Miscanthus sinensis. HortScience 2011, 46, 1353–1357. [Google Scholar] [CrossRef]
- Kurtz, L.E.; Brand, M.H.; Lubell-Brand, J.D. Production of tetraploid and triploid hemp. HortScience 2020, 55, 1703–1707. [Google Scholar] [CrossRef]
- Milo, J.; Levy, A.; Palevitch, D.; Ladizinsky, G. Thebaine content and yield in induced tetraploid and triploid plants of Papaver bracteatum Lindl. Euphytica 1987, 36, 361–367. [Google Scholar] [CrossRef]
- Jordanov, R.; Zheljazkov, V.; Tsevtkov Raev, R. Induced polyploidy in lavender. Acta Hortic. 1995, 426, 561–572. [Google Scholar]
- Xu, C.G.; Tang, T.X.; Chen, R.; Liang, C.H.; Liu, X.Y.; Wu, C.L.; Yang, Y.S.; Yang, D.P.; Wu, H. A comparative study of bioactive secondary metabolite production in diploid and tetraploid Echinacea purpurea (L.) Moench. Plant Cell Tiss. Org. 2014, 116, 323–332. [Google Scholar] [CrossRef]
- Pan, I.C.; Lu, Y.F.; Wen, P.J.; Chen, Y.M. Using colchicine to create poinsettia (Euphorbia pulcherrima × Euphorbia cornastra) mutants with various morphological traits. HortScience 2019, 54, 1667–1672. [Google Scholar] [CrossRef] [Green Version]
- González, R.L.; Iannicelli, J.; Coviella, A.; Bugallo, V.; Bologna, P.; Pitta-Álvarez, S.; Escandón, A.A. Protocol for the in vitro propagation and polyploidization of an interspecific hybrid of Glandularia (G. peruviana × G. scrobiculata). Sci. Hortic. 2015, 184, 46–54. [Google Scholar]
- Ye, Y.M.; Tong, J.; Shi, X.P.; Yuan, W.; Li, G.R. Morphological and cytological studies of diploid and colchicine-induced tetraploid lines of crape myrtle (Lagerstroemia indica L.). Sci. Hortic. 2010, 124, 95–101. [Google Scholar] [CrossRef]
- Defiani, M.R.; Astarini, I.A.; Kriswiyanti, E. Oryzalin and gamma radiation induced polyploidization in garden balsam plants (Impatiens balsamina L.) in vitro. Curr. Agric. Res. J. 2017, 5, 1–5. [Google Scholar] [CrossRef]
- Defiani, M.R.; Suprapta, D.; Sudana, I.; Ristiati, N. Oryzalin treatment modified plant morphology of Impatiens balsamina L. Curr. World Environ. 2013, 8, 23. [Google Scholar] [CrossRef] [Green Version]
- Singh, F.; Khoshoo, T. Chromosomal polymorphism within the Hibiscus rosa-sinensis complex. Caryologia 1970, 23, 19–27. [Google Scholar] [CrossRef] [Green Version]
- Bagheri, M.; Mansouri, H. Effect of induced polyploidy on some biochemical parameters in Cannabis sativa L. Appl. Biochem. Biotechnol. 2015, 175, 2366–2375. [Google Scholar] [CrossRef]
- Parsons, J.L.; Boudko, K.; Hepworth, S.; Martin, S.; James, T. Polyploidization for the genetic improvement of Cannabis sativa. Front. Plant Sci. 2019, 55, S24. [Google Scholar] [CrossRef]
- Garfinkel, A.R.; Otten, M.; Crawford, S. SNP in potentially defunct tetrahydrocannabinolic acid synthase is a marker for cannabigerolic acid dominance in Cannabis sativa L. Genes 2021, 12, 228. [Google Scholar] [CrossRef]
- Ram, H.Y.M.; Sett, R. Induction of fertile male flowers in genetically female Cannabis sativa plants by silver-nitrate and silver thiosulfate anionic complex. Theor. Appl. Genet. 1982, 62, 369–375. [Google Scholar]
- Vaclavik, L.; Benes, F.; Fenclova, M.; Hricko, J.; Krmela, A.; Svobodova, V.; Hajslova, J.; Mastovska, K. Quantitation of cannabinoids in cannabis dried plant materials, concentrates, and oils using liquid chromatography–diode array detection technique with optional mass spectrometric detection: Single-laboratory validation study, first action 2018.11. J. AOAC Int. 2019, 102, 1822–1833. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Chung, M.C.; Tsai, Y.C.; Wei, F.J.; Hsieh, J.S.; Hsing, Y.L.C. Distribution of new satellites and simple sequence repeats in annual and perennial Glycine species. Bot. Stud. 2015, 56, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lattier, J.D.; Chen, H.; Contreras, R.N. Improved method of enzyme digestion for root tip cytology. HortScience 2017, 52, 1029–1032. [Google Scholar] [CrossRef]
- Prunet, N.; Morel, P.; Negrutiu, I.; Trehin, C. Time to stop: Flower meristem termination. Plant Physiol. 2009, 150, 1764–1772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jibran, R.; Tahir, J.; Cooney, J.; Hunter, D.A.; Dijkwel, P.P. Arabidopsis AGAMOUS regulates sepal senescence by driving jasmonate production. Front. Plant Sci. 2017, 8, 2101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rogers, H.J. From models to ornamentals: How is flower senescence regulated? Plant Mol. Biol. 2013, 82, 563–574. [Google Scholar] [CrossRef] [PubMed]
- Contreras, R.N.; Ruter, J.M.; Hanna, W.W. An oryzalin-induced autoallooctoploid of Hibiscus acetosella ‘Panama Red’. J. Am. Soc. Hortic. Sci. 2009, 134, 553–559. [Google Scholar] [CrossRef] [Green Version]
- Ahanchede, A.; Poirier-Hamon, S.; Darmency, H. Why no tetraploid cultivar of foxtail millet? Genet. Resour. Crop Evol. 2004, 51, 227–230. [Google Scholar] [CrossRef]
- Graebner, R.C.; Chen, H.; Contreras, R.N.; Haynes, K.G.; Sathuvalli, V. Identification of the high frequency of triploid potato resulting from tetraploid× diploid crosses. HortScience 2019, 54, 1159–1163. [Google Scholar] [CrossRef] [Green Version]
- Stoute, A.I.; Varenko, V.; King, G.J.; Scott, R.J.; Kurup, S. Parental genome imbalance in Brassica oleracea causes asymmetric triploid block. Plant J. 2012, 71, 503–516. [Google Scholar] [CrossRef] [Green Version]
- Munzbergova, Z. Colchicine application significantly affects plant performance in the second generation of synthetic polyploids and its effects vary between populations. Ann. Bot. 2017, 120, 329–339. [Google Scholar] [CrossRef]
- DiMatteo, J.; Kurtz, L.; Lubell-Brand, J.D. Pollen Appearance and In Vitro Germination Varies for Five Strains of Female Hemp Masculinized Using Silver Thiosulfate. HortScience 2020, 55, 547–549. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tests | Ploidy | Plant Material 1 | Number of Plants |
|---|---|---|---|
| Fertility | 2x | F1 hybrid, TS1-3 (2x) × P163 (2x) 2 | 8 |
| 3x | F1 hybrid, TS1-3 (4x) × P163 (2x) 3 | 8 | |
| 4x | F1 hybrid, TS1-3 (4x) × P163 (4x) | 8 | |
| 2x | F1 hybrid, TS1-3 (2x) × P163 (2x) 2 | 8 | |
| 3x | F1 hybrid, TS1-3 (4x) × P163 (2x) | 8 | |
| 4x | F1 hybrid, TS1-3 (4x) × P163 (4x) | 8 | |
| Indoor test | 2x | F1 hybrid, TS1-3 (2x) × P163 (2x) 2 | 7 |
| 3x | F1 hybrid, TS1-3 (4x) × P163 (2x) 3 | 7 | |
| 4x | F1 hybrid, TS1-3 (4x) × P163 (4x) | 7 | |
| Field test | 2x | F1 hybrid, TS1-3 (2x) × P163 (2x) 2 | 15 |
| 3x | F1 hybrid, TS1-3 (4x) × P163 (2x) 3 | 15 |
| Seed Parent | Pollen Donor | Filled Seed Number 1,2 | Empty or Abnormal Seed Number | Total Seed Number | Compatibility 4 | Weight of 100 Seeds (g) 3 |
|---|---|---|---|---|---|---|
| 2x F1 hybrid | 2x F1 hybrid | 317.25 a | 36.75 | 354 | Yes | 1.25 b |
| 3x F1 hybrid | 2x F1 hybrid | 4.25 b | 4 | 8.25 | - | - |
| 4x F1 hybrid | 2x F1 hybrid | 68.63 c | 17.25 | 99.75 | Yes | 1.30 b |
| 2x F1 hybrid | 4x F1 hybrid | 0 A | 232.75 | 232.75 | No | 0.37 d |
| 3x F1 hybrid | 4x F1 hybrid | 5.63 B | 11.63 | 17.25 | - | - |
| 4x F1 hybrid | 4x F1 hybrid | 124.75 C | 10.13 | 134.88 | Yes | 2.07 a |
| Female Material | Number of Plants | Biomass Per Plant (g) 1 | Inflorescence Weight Per Plant (g) 1 | Total CBG Concentration (%) 1 | Total THC Concentration (%) 1 | Total Cannabinoid Concentration (%) 1 | CBG Yield Per Plant (g) | CBG: THC |
|---|---|---|---|---|---|---|---|---|
| 2x F1 hybrid-field | 15 | 2097.4 A | 1068.3 A | - | - | - | - | - |
| 3x F1 hybrid-field | 15 | 3311.3 B | 1312.1 B | - | - | - | - | - |
| 2x F1 hybrid-indoor | 7 | 57.7 a | 24.3 a | 7.78 a | 0.141 a | 8.66 a | 2.01 a | 64:1 |
| 3x F1 hybrid-indoor | 7 | 67.8 a | 30.7 b | 9.15 ab | 0.138 a | 10.18 ab | 3.11 a | 73:1 |
| 4x F1 hybrid-indoor | 7 | 58.0 a | 23.1 a | 11.23 b | 0.137 a | 12.38 b | 2.93 a | 84:1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Crawford, S.; Rojas, B.M.; Crawford, E.; Otten, M.; Schoenenberger, T.A.; Garfinkel, A.R.; Chen, H. Characteristics of the Diploid, Triploid, and Tetraploid Versions of a Cannabigerol-Dominant F1 Hybrid Industrial Hemp Cultivar, Cannabis sativa ‘Stem Cell CBG’. Genes 2021, 12, 923. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12060923
Crawford S, Rojas BM, Crawford E, Otten M, Schoenenberger TA, Garfinkel AR, Chen H. Characteristics of the Diploid, Triploid, and Tetraploid Versions of a Cannabigerol-Dominant F1 Hybrid Industrial Hemp Cultivar, Cannabis sativa ‘Stem Cell CBG’. Genes. 2021; 12(6):923. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12060923
Chicago/Turabian StyleCrawford, Seth, Brendan M. Rojas, Eric Crawford, Matthew Otten, Thecla A. Schoenenberger, Andrea R. Garfinkel, and Hsuan Chen. 2021. "Characteristics of the Diploid, Triploid, and Tetraploid Versions of a Cannabigerol-Dominant F1 Hybrid Industrial Hemp Cultivar, Cannabis sativa ‘Stem Cell CBG’" Genes 12, no. 6: 923. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12060923