
Detection of Genes in Arabidopsis thaliana L. Responding to DNA Damage from Radiation and Other Stressors in Spaceflight


Abstract
:1. Introduction
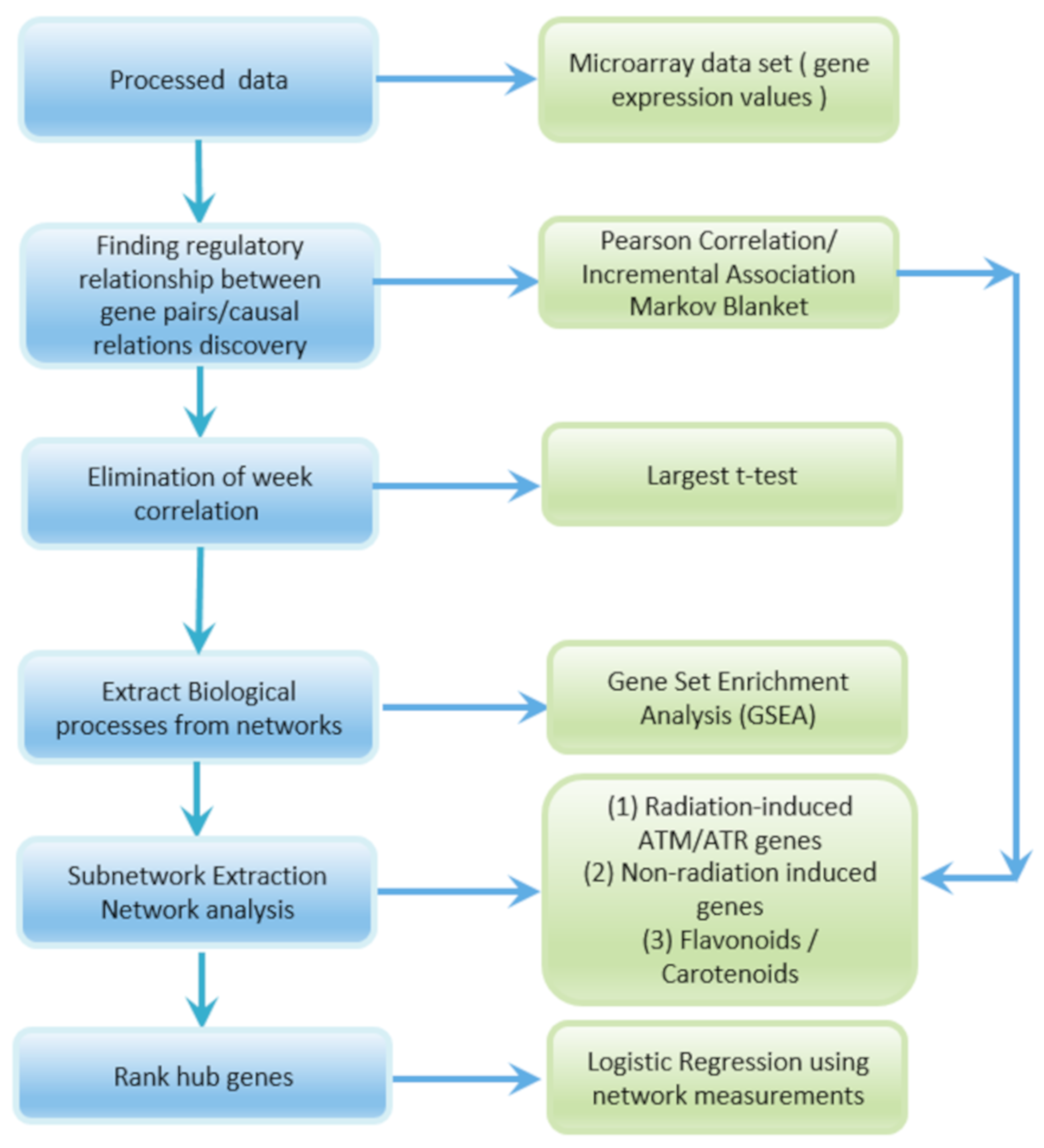
2. Materials and Methods
2.1. GeneLab Datasets
2.2. Gene Regulatory Network Inferencing Using Pearson Correlation
2.3. Causal Relations Discovery Using Incremental Association Markov Blanket (IAMB) Method
2.4. Computation of Network Measures
2.5. Logistic Regression-Based Gene Ranking
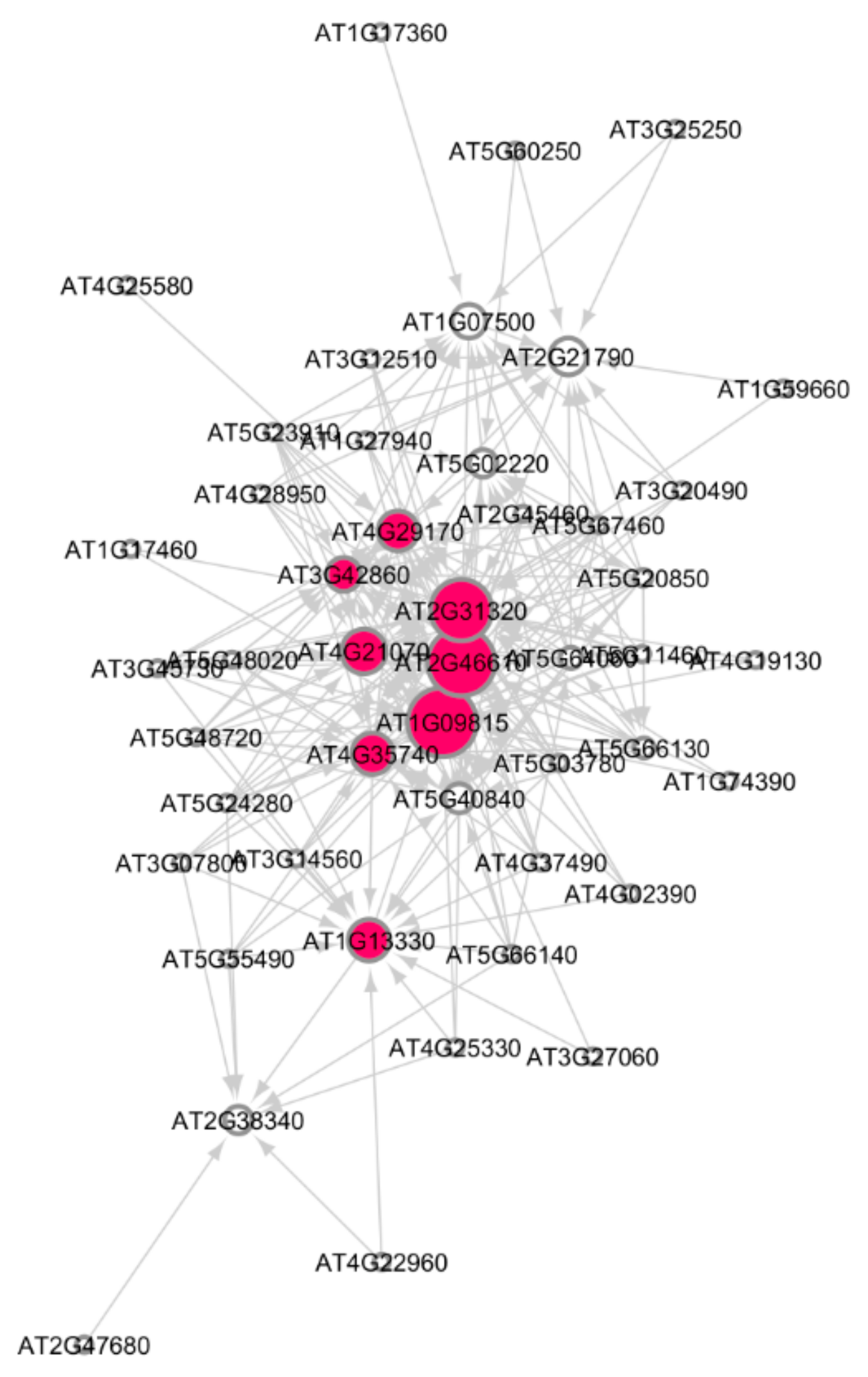
3. Results
3.1. Cellular Response to Stimulus
3.2. Cellular Response to Stress
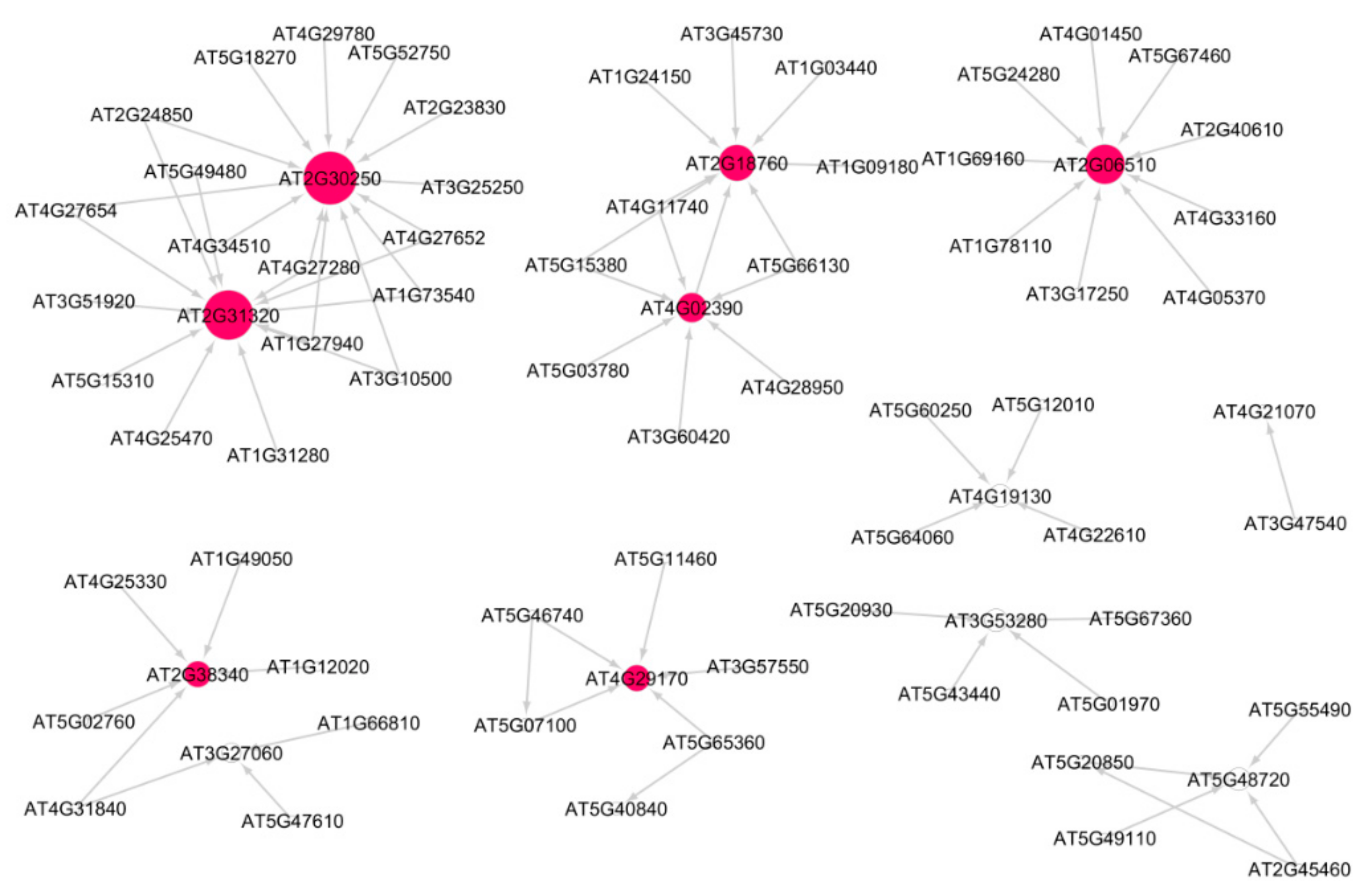
3.3. Cellular Response to DNA Damage Stimulus
3.4. DNA Metabolic Process
3.5. Flavonoid Biosynthesis and Carotenoid Catabolic Processes
3.6. Subnetwork Measurments for the Low, and Very High Radiation Dose DDR Processes
3.7. Jaccard Similarity Between Subnetworks
3.8. Logistic Regression Ranking of Hub Genes
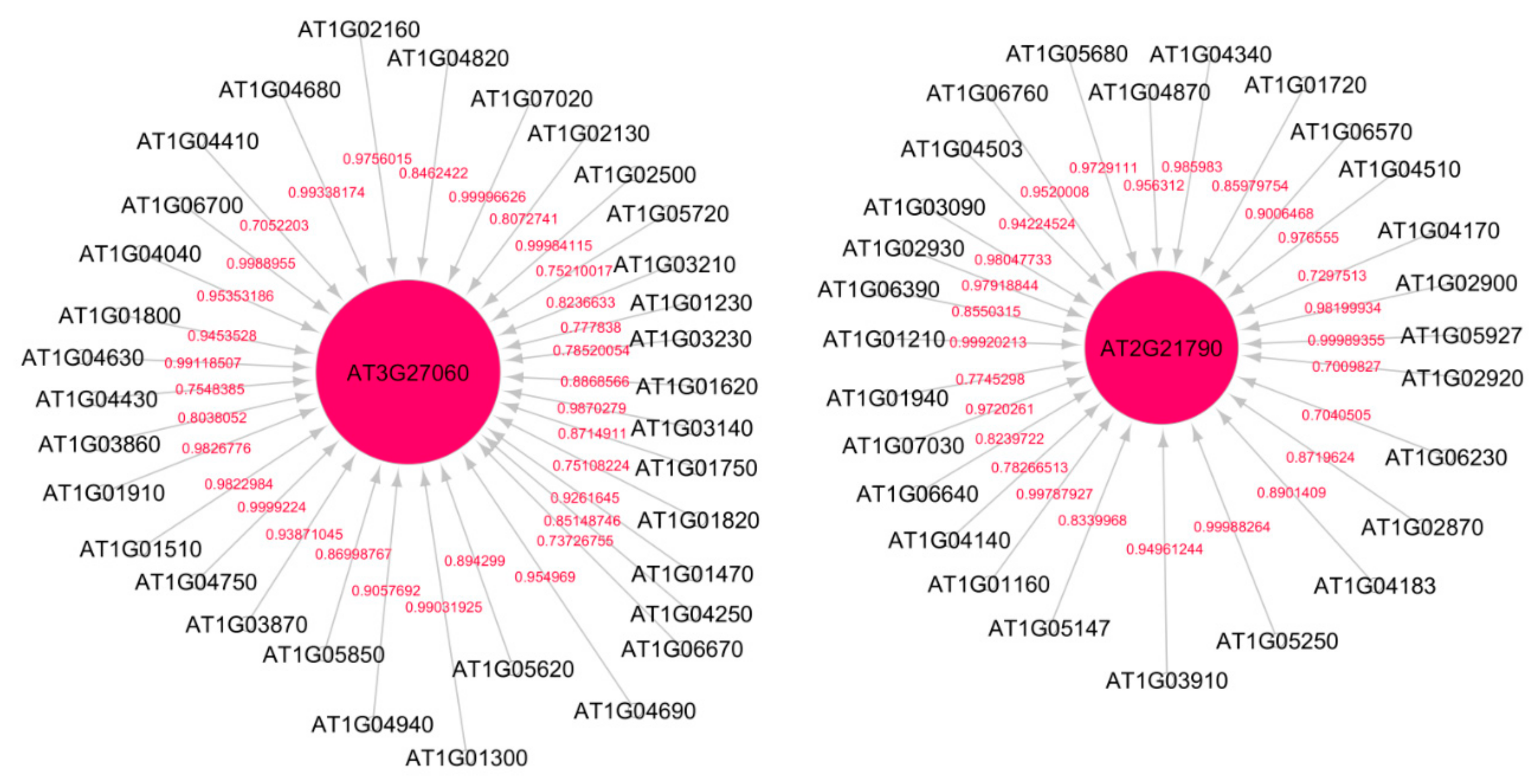
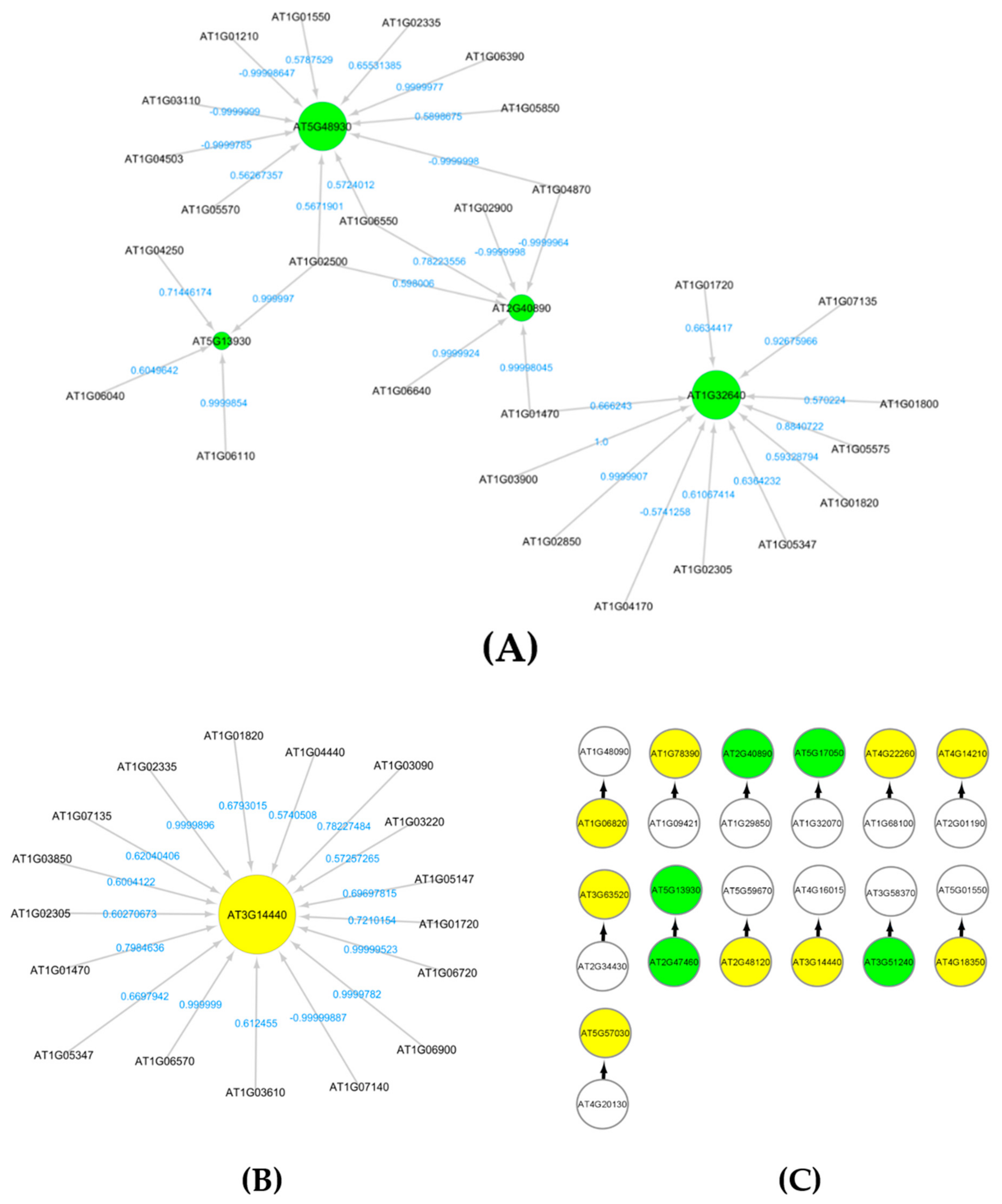
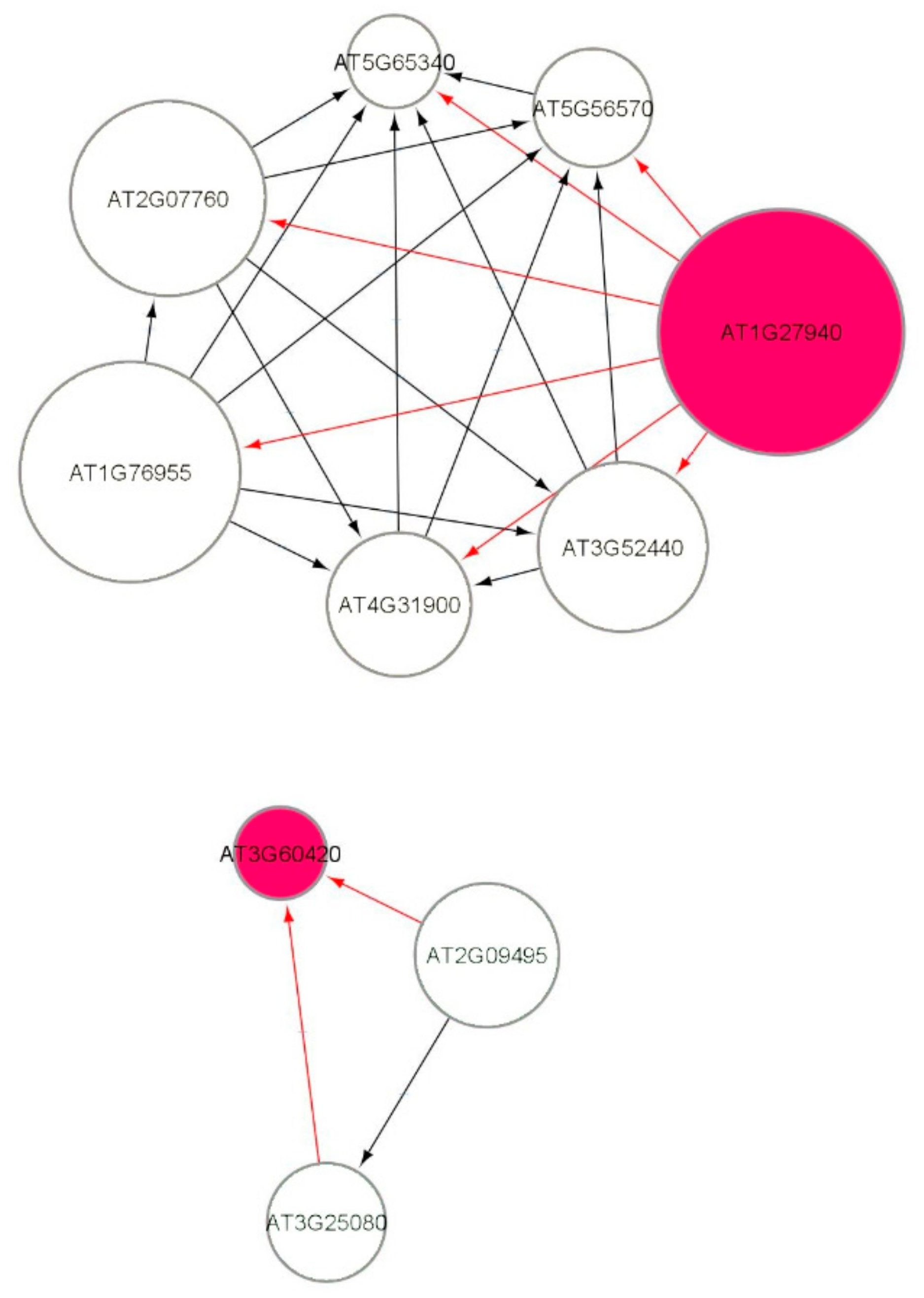
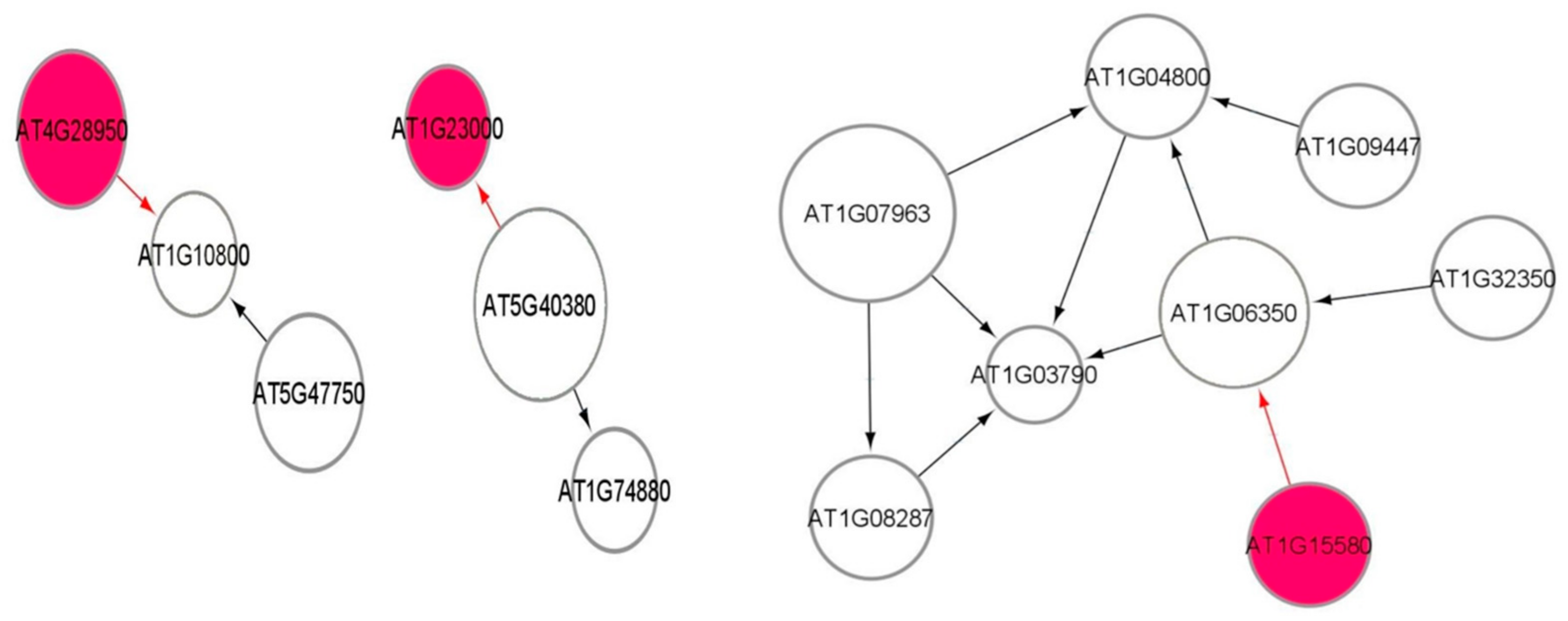
3.9. Causal Relational Network Analysis
4. Discussion
4.1. Computational Strategy
4.2. ATR/ATM Gene Interactions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nelson, G.A. Space Radiation and Human Exposures, A Primer. Radiat. Res. 2016, 185, 349–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- NASA. Why Space Radiation Matters. Available online: https://www.nasa.gov/analogs/nsrl/why-space-radiation-matters (accessed on 15 March 2021).
- McDonald, J.T.; Stainforth, R.; Miller, J.; Cahill, T.; Silveira, W.A.D.; Rathi, K.S.; Hardiman, G.; Taylor, D.; Costes, S.V.; Chauhan, V.; et al. NASA genelab platform utilized for biological response to space radiation in animal models. Cancers 2020, 12, 381. [Google Scholar] [CrossRef] [Green Version]
- Nikitaki, Z.; Pavlopoulou, A.; Holá, M.; Donà, M.; Michalopoulos, I.; Balestrazzi, A.; Angelis, K.J.; Georgakilas, A.G. Bridging plant and human radiation response and DNA repair through an in silico approach. Cancers 2017, 9, 65. [Google Scholar] [CrossRef]
- Field, M.; Growth, S.; Biology, A.P. NASA Facts Variable Gravity Plant Biology; NASA Report; National Aeronautics and Space Administration: Moffett Field, CA, USA, 2017.
- Shymanovich, T.; Kiss, J.Z. Growth and Development of Ecotypes of Arabidopsis thaliana: Preliminary Experiments to Prepare for a Moon Lander Mission. Gravit. Space Res. 2020, 8, 18–24. [Google Scholar] [CrossRef]
- Beisel, N.S.; Noble, J.; Barbazuk, W.B.; Paul, A.L.; Ferl, R.J. Spaceflight-induced alternative splicing during seedling development in Arabidopsis thaliana. NPJ Microgravity 2019, 5, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Caplin, N.M.; Halliday, A.; Willey, N.J. Developmental, Morphological and Physiological Traits in Plants Exposed for Five Generations to Chronic Low-Level Ionising Radiation. Front. Plant. Sci. 2020, 11, 1–14. [Google Scholar] [CrossRef]
- Maréchal, A.; Zou, L. DNA damage sensing by the ATM and ATR kinases. Cold Spring Harb. Perspect. Biol. 2013, 5, 1–17. [Google Scholar] [CrossRef]
- Ricaud, L.; Proux, C.; Renou, J.-P.; Pichon, O.; Fochesato, S.; Ortet, P.; Montané, M.-H. ATM-mediated transcriptional and developmental responses to γ-rays in Arabidopsis. PLoS ONE 2007, 2, e430. [Google Scholar] [CrossRef]
- Culligan, K.M.; Robertson, C.E.; Foreman, J.; Doerner, P.; Britt, A.B. ATR and ATM play both distinct and additive roles in response to ionizing radiation. Plant J. 2006, 48, 947–961. [Google Scholar] [CrossRef]
- Hase, Y.; Satoh, K.; Seito, H.; Oono, Y. Genetic Consequences of Acute/Chronic Gamma and Carbon Ion Irradiation of Arabidopsis thaliana. Front. Plant Sci. 2020, 11, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, Y.; Sasaki, N.; Ohmiya, A. Biosynthesis of plant pigments: Anthocyanins, betalains and carotenoids. Plant J. 2008, 54, 733–749. [Google Scholar] [CrossRef] [PubMed]
- Hirschberg, J. Carotenoid biosynthesis in flowering plants. Curr. Opin. Plant Biol. 2001, 4, 210–218. [Google Scholar] [CrossRef]
- Pizarro, L.; Stange, C. Light-dependent regulation of carotenoid biosynthesis in plants. Cienc. Investig. Agrar. 2009, 36, 143–162. [Google Scholar] [CrossRef]
- Ruiz-Sola, M.Á.; Rodríguez-Concepción, M. Carotenoid Biosynthesis in Arabidopsis: A Colorful Pathway. Arab. B 2012, 10, e0158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- TAIR—Reference Genome Sequence. Available online: https://www.arabidopsis.org/doc/portals/genAnnotation/gene_structural_annotation/ref_genome_sequence/11413 (accessed on 21 March 2020).
- Missirian, V.; Conklin, P.A.; Culligan, K.M.; Huefner, N.D.; Britt, A.B. High atomic weight, high-energy radiation (HZE) induces transcriptional responses shared with conventional stresses in addition to a core ‘DSB’ response specific to clastogenic treatments. Front. Plant Sci. 2014, 5, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Chowdhury, H.A.; Bhattacharyya, D.K.; Kalita, J.K. (Differential) Co-Expression Analysis of Gene Expression: A Survey of Best Practices. IEEE/ACM Trans. Comput. Biol. Bioinform. 2020, 17, 1154–1173. [Google Scholar] [CrossRef] [PubMed]
- NASA Gene Lab Data Repository. Available online: https://genelab.nasa.gov (accessed on 15 March 2021).
- Paul, A.L.; Zupanska, A.K.; Schultz, E.R.; Ferl, R.J. Organ-specific remodeling of the Arabidopsis transcriptome in response to spaceflight. BMC Plant Biol. 2013, 13, 112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paul, A.L.; Sng, N.J.; Zupanska, A.K.; Krishnamurthy, A.; Schultz, E.R.; Ferl, R.J. Genetic dissection of the Arabidopsis spaceflight transcriptome: Are some responses dispensable for the physiological adaptation of plants to spaceflight? PLoS ONE 2017, 12, e0180186. [Google Scholar] [CrossRef]
- Leister, D.; Schneider, A. From Genes to Photosynthesis in Arabidopsis thaliana. Int. Rev. Cytol. 2003, 228, 31–83. [Google Scholar] [CrossRef]
- Choi, W.G.; Barker, R.J.; Kim, S.H.; Swanson, S.J.; Gilroy, S. Variation in the transcriptome of different ecotypes of Arabidopsis thaliana reveals signatures of oxidative stress in plant responses to spaceflight. Am. J. Bot. 2019, 106, 123–136. [Google Scholar] [CrossRef]
- Kruse, C.P.S.; Basu, P.; Luesse, D.R.; Wyatt, S.E. Transcriptome and proteome responses in RNAlater preserved tissue of Arabidopsis thaliana. PLoS ONE 2017, 12, e0175943. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.P. Quantifying gene regulatory relationships with association measures: A comparative study. Front. Genet. 2017, 8, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Manian, V.; Orozco, J.; Gangapuram, H.; Janwa, H.; Agrinsoni, C. Network Analysis of Gene Transcriptions of Arabidopsis thaliana in Spaceflight Microgravity. Genes 2021, 12, 337. [Google Scholar] [CrossRef] [PubMed]
- Biggs, N. Algebraic Graph Theory, 2nd ed.; Cambridge University Press: Cambridge, UK, 1993. [Google Scholar]
- Gavai, A.K. Bayesian Networks for Omics Data Analysis. Ph.D. Thesis, Wageningen University, Wageningen, The Netherlands, 2009. [Google Scholar]
- Ainsworth, H.F.; Shin, S.Y.; Cordell, H.J. A comparison of methods for inferring causal relationships between genotype and phenotype using additional biological measurements. Genet. Epidemiol. 2017, 41, 577–586. [Google Scholar] [CrossRef] [Green Version]
- Ram, R.; Chetty, M. A Markov-blanket-based model for gene regulatory network inference. IEEE/ACM Trans. Comput. Biol. Bioinforma. 2011, 8, 353–367. [Google Scholar] [CrossRef]
- Yang, X.; Wang, Y.; Ou, Y.; Tong, Y. Three-Fast-Inter Incremental Association Markov Blanket learning algorithm. Pattern Recognit. Lett. 2019, 122, 73–78. [Google Scholar] [CrossRef]
- Ahmed, S.S.; Roy, S.; Kalita, J. Assessing the Effectiveness of Causality Inference Methods for Gene Regulatory Networks. IEEE/ACM Trans. Comput. Biol. Bioinforma. 2020, 17, 56–70. [Google Scholar] [CrossRef]
- Tsamardinos, I.; Aliferis, C.; Statnikov, A.; Statnikov, E. Algorithms for Large Scale Markov Blanket Discovery. FLAIRS Conf. 2003, 376–381. [Google Scholar] [CrossRef]
- Bui, A.T.; Jun, C.H. Learning Bayesian network structure using Markov blanket decomposition. Pattern Recognit. Lett. 2012, 33, 2134–2140. [Google Scholar] [CrossRef]
- Pellet, J.P.; Elisseeff, A. Using Markov blankets for causal structure learning. J. Mach. Learn. Res. 2008, 9, 1295–1342. [Google Scholar]
- Loscalzo, J.; Barabási, A.-L. Network Science, 1st ed.; Cambridge University Press: Cambridge, UK, 2016. [Google Scholar]
- Janwa, H.; Massey, S.E.; Velev, J.; Mishra, B. On the origin of biomolecular networks. Front. Genet. 2019, 10, 240–257. [Google Scholar] [CrossRef] [Green Version]
- Bass, J.I.F.; Diallo, A.; Nelson, J.; Soto, J.M.; Myers, C.L.; Walhout, A.J.M. Using networks to measure similarity between genes: Association index selection. Nat. Methods 2013, 10, 1169–1176. [Google Scholar] [CrossRef] [Green Version]
- Khunlertgit, N.; Yoon, B.J. Incorporating topological information for predicting robust cancer subnetwork markers in human protein-protein interaction network. BMC Bioinform. 2016, 17 (Suppl. 13), 143–152. [Google Scholar] [CrossRef] [Green Version]
- Opsahl, T.; Agneessens, F.; Skvoretz, J. Node centrality in weighted networks: Generalizing degree and shortest paths. Soc. Netw. 2010, 32, 245–251. [Google Scholar] [CrossRef]
- Cickovski, T.; Peake, E.; Aguiar-Pulido, V.; Narasimhan, G. ATria: A novel centrality algorithm applied to biological networks. BMC Bioinform. 2017, 18 (Suppl. 8), 1–10. [Google Scholar] [CrossRef] [Green Version]
- Mistry, D.; Wise, R.P.; Dickerson, J.A. DiffSLC: A graph centrality method to detect essential proteins of a protein-protein interaction network. PLoS ONE 2017, 12, e0187091. [Google Scholar] [CrossRef] [Green Version]
- Sartor, M.A.; Leikauf, G.D.; Medvedovic, M. LRpath: A logistic regression approach for identifying enriched biological groups in gene expression data. Bioinformatics 2009, 25, 211–217. [Google Scholar] [CrossRef] [PubMed]
- ShinyGO. Available online: http://bioinformatics.sdstate.edu/go/ (accessed on 15 March 2021).
- Gene Ontology and GO Annotations. Available online: https://www.ebi.ac.uk/QuickGO/ (accessed on 25 March 2021).
- Paul, A.-L.; Zupanska, A.K.; Ostrow, D.T.; Zhang, Y.; Sun, Y.; Li, J.-L.; Shanker, S.; Farmerie, W.G.; Amalfitano, C.E.; Ferl, R.J. Spaceflight transcriptomes: Unique responses to a novel environment. Astrobiology 2012, 12, 40–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, N.; Ogita, N.; Takahashi, T.; Taniguchi, S.; Tanaka, M.; Seki, M.; Umeda, M. A regulatory module controlling stress-induced cell cycle arrest in Arabidopsis. eLife 2019, 8, 1–27. [Google Scholar] [CrossRef]
- Shultz, R.W.; Tatineni, V.M.; Hanley-Bowdoin, L.; Thompson, W.F. Genome-wide analysis of the core DNA replication machinery in the higher plants Arabidopsis and rice1[W][OA]. Plant Physiol. 2007, 144, 1697–1714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tossi, V.E.; Regalado, J.J.; Iannicelli, J.; Laino, L.E.; Burrieza, H.P.; Escandón, A.S.; Pitta-Álvarez, S.I. Beyond Arabidopsis: Differential UV-B response mediated by UVR8 in diverse species. Front. Plant Sci. 2019, 10, 1–17. [Google Scholar] [CrossRef]
- Peng, W.; Wang, J.; Zhang, Z.; Wu, F.-X. Applications of Random Walk Model on Biological Networks. Curr. Bioinform. 2016, 11, 211–220. [Google Scholar] [CrossRef]
- Radiation Data for GLDS Studies on the International Space Station. Available online: https://genelab.nasa.gov/environmental/radiation/iss (accessed on 5 April 2021).
- Mohorianu, I.; Bretman, A.; Smith, D.T.; Fowler, E.K.; Dalmay, T.; Chapman, T. Comparison of alternative approaches for analysing multi-level RNA-seq data. PLoS ONE 2017, 12, e0182694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manian, V.; Gangapuram, H.; Orozco, J.; Janwa, H.; Agrinsoni, C. Network Analysis of Local Gene Regulators in Arabidopsis thaliana under Spaceflight Stress. Computers 2021, 10, 18. [Google Scholar] [CrossRef]
- Giscard, P.L.; Wilson, R.C. A centrality measure for cycles and subgraphs II. Appl. Netw. Sci. 2018, 3. [Google Scholar] [CrossRef] [Green Version]
- Estrada, E. About the Discriminant Power of the Subgraph Centrality and Other Centrality Measures About the Discriminant Power of the Subgraph Centrality and Other Centrality Measures (Working Paper). 2013. Available online: http://arxiv.org/abs/1305.6836 (accessed on 15 April 2021).
- Ouma, W.Z.; Pogacar, K.; Grotewold, E. Topological and statistical analyses of gene regulatory networks reveal unifying yet quantitatively different emergent properties. PLoS Comput. Biol. 2018, 14, e1006098. [Google Scholar] [CrossRef] [PubMed]
- Koutrouli, M.; Karatzas, E.; Paez-Espino, D.; Pavlopoulos, G.A. A Guide to Conquer the Biological Network Era Using Graph Theory. Front. Bioeng. Biotechnol. 2020, 8, 1–26. [Google Scholar] [CrossRef]
- Espinosa-Soto, C. On the role of sparseness in the evolution of modularity in gene regulatory networks. PLoS Comput. Biol. 2018, 14, e1006172. [Google Scholar] [CrossRef]
- Van den Broeck, L.; Gordon, M.; Inzé, D.; Williams, C.; Sozzani, R. Gene Regulatory Network Inference: Connecting Plant Biology and Mathematical Modeling. Front. Genet. 2020, 11, 1–12. [Google Scholar] [CrossRef]
- Menolfi, D.; Zha, S. ATM, ATR and DNA-PKcs kinases-the lessons from the mouse models: Inhibition = deletion. Cell Biosci. 2020, 10, 1–15. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Arabidopsis Gene Identifier for Hub Genes | Degree Distribution | Subgraph Centrality | ||
|---|---|---|---|---|
| LRD1 | LRD2 | LRD1 | LRD2 | |
| AT1G08260 | 1 | 1 | 1.5431 | 1.5431 |
| AT1G29440 | 1 | 1 | 1.5431 | 1.5431 |
| AT2G30360 | 1 | 4 | 1.5431 | 1.5431 |
| AT3G13380 | 12 | 7 | 15.9895 | 7.0825 |
| AT3G51920 | 6 | 1 | 5.8344 | 1.5431 |
| AT3G61630 | 2 | 13 | 2.1782 | 18.4146 |
| AT4G28950 | 1 | 3 | 1.5431 | 2.9146 |
| AT5G07100 | 1 | 9 | 1.5431 | 10.0677 |
| AT5G40840 | 4 | 4 | 3.7622 | 3.7622 |
| AT5G48720 | 1 | 20 | 1.5431 | 3.7622 |
| AT5G61600 | 1 | 15 | 2.1782 | 18.4146 |
| Arabidopsis Gene Identifier for Hub Genes | Degree Distribution | Subgraph Centrality | ||
|---|---|---|---|---|
| LRD3 | LRD4 | LRD3 | LRD4 | |
| AT3G27060 | 2 | 22 | 2.3811 | 6253.2089 |
| AT3G51920 | 4 | 40 | 4.9976 | 11,529.486 |
| AT1G70940 | 1 | 22 | 1.5907 | 12,412.744 |
| Arabidopsis Gene Identifier for Hub Genes | Degree Distribution | Subgraph Centrality | ||
|---|---|---|---|---|
| LRD1 | LRD2 | LRD1 | LRD2 | |
| AT1G08260 | 1 | 1 | 1.5431 | 1.5431 |
| AT5G40840 | 4 | 4 | 3.7622 | 3.7622 |
| AT5G07100 | 1 | 9 | 1.5431 | 10.0677 |
| AT5G48720 | 1 | 20 | 1.5431 | 43.7775 |
| DDR Processes | Degree Distribution | Subgraph Centrality | ||
|---|---|---|---|---|
| LRD3 | LRD4 | LRD3 | LRD4 | |
| Cellular response to stress | 3 | 22 | 54.4538 | 2.9737 |
| Cellular response to DNA damage stimulus | 2 | 33 | 2.1782 | 156.2451 |
| DNA metabolic process | 2 | 22 | 2.3811 | 57.2840 |
| Cellular Response to Stimulus | ||||
|---|---|---|---|---|
| Network Measurements | LRD3 | LRD4 | LRD1 | LRD2 |
| Spectral gap | 0.3347 | 6.172 | 2.013 | 0.099 |
| Density | 0.0489 | 0.0711 | 0.022 | 0.0037 |
| Diameter | 1 | 1 | 1 | 1 |
| Conn. Comps. | 3 | 2 | 3 | 28 |
| Cellular Response to Stress | ||||
| Spectral gap | 0.7923 | 4.69 | 0.99 | 0.099 |
| Density | 0.027 | 0.087 | 0.102 | 0.0104 |
| Diameter | 2 | 1 | 1 | 1 |
| Conn. Comps. | 9 | 1 | 5 | 17 |
| DNA Metabolic Process | ||||
| Spectral gap | 0.858 | 1.202 | 0.99 | 0.4112 |
| Density | 0.5714 | 0.038 | 0.166 | 0.0059 |
| Diameter | 1 | 1 | 1 | 2 |
| Conn. Comps | 6 | 2 | 3 | 69 |
| Cellular Response to DNA Damage Stimulus | ||||
| Spectral gap | 0.068 | 5.74 | 0.99 | 0.1399 |
| Density | 0.055 | 0.058 | 0.166 | 0.0074 |
| Diameter | 1 | 1 | 1 | 2 |
| Conn. Comps. | 7 | 1 | 3 | 55 |
| Network Measurements | DNA Metabolic Process | Nucleic Acid Response Process | ||
|---|---|---|---|---|
| HZE | γR | HZE | γR | |
| Density | 0.1782 | 0.2040 | 0.2465 | 0.2211 |
| Spectral gap | 7.5153 | 8.9726 | 12.2554 | 10.4919 |
| Diameter | 2 | 4 | 2 | 5 |
| Subnetwork with Hub-Gene | Jaccard Similarity Index between HZE and γR Subnetworks | |
| DNA Metabolic Process | Nucleic Acid Response Process | |
| AT1G09815 | 0.5789 | 0.5789 |
| AT2G31320 | 0.6176 | 0.6176 |
| AT4G21070 | 0.7619 | - |
| AT1G13330 | 0.4814 | 0.4285 |
| AT4G29170 | 0.4444 | 0.4444 |
| AT2G46610 | - | 0.6250 |
| Subnetwork with hub-gene | Jaccard similarity index between LRD3 and LRD4 subnetworks | |
| DNA metabolic process and Response to DNA damage to stimulus | DNA metabolic process & Response to DNA damage to stimulus | |
| AT3G27060 | 1.0 | 0.1224 |
| Arabidopsis Gene Identifier | Gene Coding Protein Description |
|---|---|
| AT3G13380 | Protein binding, protein serine kinase activity |
| AT3G61630 | Cotyledon development, embryo development ending in seed dormancy, leaf development, regulation of transcription, DNA-templated |
| AT5G61600 | Cell division, defense response to fungus, phloem or xylem histogenesis, positive regulation of transcription, DNA-templated |
| AT3G27060 | Directly involved in synthesis of deoxyribonucleotides, DNA repair, DNA replication, multicellular organism development, programmed cell death, regulation of cell cycle |
| AT3G51920 | Response to salt stress and water deprivation, calcium ion binding. |
| AT1G70940 | Positive gravitropism, regulation of root meristem growth, response to light stimulus, root development, root hair elongation, root hair initiation |
| AT5G48720 | DNA repair, female meiotic nuclear division, pollen development, response to X-ray |
| AT2G31320 | DNA ADP-ribosylation, DNA repair, double-strand break repair, protein ADP-ribosylation, protein poly-ADP-ribosylation, response to abscisic acid, response to oxidative stress |
| AT4G21070 | DNA recombination, DNA repair, cellular response to gamma radiation, double-strand break repair via homologous recombination, negative regulation of fatty acid biosynthetic process |
| AT2G46610 | mRNA splicing, via spliceosome, RNA binding, protein binding |
| AT3G21280 | Protein deubiquitination, ubiquitin-dependent protein catabolic process |
| AT1G07500 | Cellular response to DNA damage stimulus, negative regulation of mitotic nuclear division, regulation of DNA endoreduplication |
| AT5G66140 | Proteasomal ubiquitin-independent protein catabolic process |
| AT1G27940 | ATPase-coupled transmembrane transporter activity and nucleotide binding |
| AT3G60420 | Phosphoglycerate mutase family protein |
| AT4G28950 | Meiotic DNA repair, pollen development, and responds to X-ray |
| AT1G23000 | Heavy metal transport/detoxification superfamily protein involved in metal ion transport |
| AT1G15580 | Regulation of transcription, DNA-templated, response to auxin |
| Arabidopsis Gene Identifier | Gene Coding Protein Description |
|---|---|
| AT1G01470 | Induced in response to wounding and light stress. Might be involved in protection against desiccation. |
| AT1G06390 | This gene is involved in response to osmotic stress. This protein can interact with the BZR1 protein involved in brassinosteroid-mediated signaling in a Y2H assay and promotes BZR1 phosphorylation in protoplasts. |
| AT1G05850 | Essential for tolerance to heat, salt and drought stresses. Also involved in root hair development, cell expansion and response to cytokinin. |
| AT1G05680 | This enzyme can also transfer glycosyl groups to several compounds related to the explosive TNT when this synthetic compound is taken up from the environment. |
| AT1G05620 | Transcript levels for this gene are elevated in older leaves suggesting that it may play a role in purine catabolism during senescence. |
| AT3G22370 | Plays a role in shoot acclimation to low temperature. Also is capable of ameliorating reactive oxygen species production when the cytochrome pathway is inhibited. |
| AT5G43680 | The protein is localized to the inner mitochondrial membrane that is nuclear-encoded and is essential for plant growth and development. |
| AT2G19620 | Plays a role in dehydration stress response. |
| AT4G34410 | Direct participation in auxin biosynthesis leading to the plant’s ability to tolerate salt stress. |
| AT2G19620 | Plays a role in dehydration stress response. |
| AT4G31480 | Required for plant growth, salt tolerance, and maintenance of the structure of the Golgi apparatus. |
| AT1G72490 | It is expressed in roots and involved in leaf root architecture, specifically the orientation of lateral root angles |
| AT5G61020 | Involved in cell proliferation during plant organogenesis. |
| AT5G45420 | Plays a role in root hair elongation. |
| Important role in controlling root skewing and maintaining the microtubule network. | |
| AT4G35100 | Salt-stress-inducible Major Intrinsic Protein (MIP) |
| AT1G13900 | Encodes a dual-localized acid phosphatase (mitochondria and chloroplast) that modulates carbon metabolism. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manian, V.; Orozco-Sandoval, J.; Diaz-Martinez, V. Detection of Genes in Arabidopsis thaliana L. Responding to DNA Damage from Radiation and Other Stressors in Spaceflight. Genes 2021, 12, 938. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12060938
Manian V, Orozco-Sandoval J, Diaz-Martinez V. Detection of Genes in Arabidopsis thaliana L. Responding to DNA Damage from Radiation and Other Stressors in Spaceflight. Genes. 2021; 12(6):938. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12060938
Chicago/Turabian StyleManian, Vidya, Jairo Orozco-Sandoval, and Victor Diaz-Martinez. 2021. "Detection of Genes in Arabidopsis thaliana L. Responding to DNA Damage from Radiation and Other Stressors in Spaceflight" Genes 12, no. 6: 938. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12060938