
Two RECK Splice Variants (Long and Short) Are Differentially Expressed in Patients with Stable and Unstable Coronary Artery Disease: A Pilot Study


 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants’ Enrollment and Samples Collection
2.2. PBMCs Isolation
2.3. RNA Sequencing Study
2.4. RNA Sequencing Data Analysis
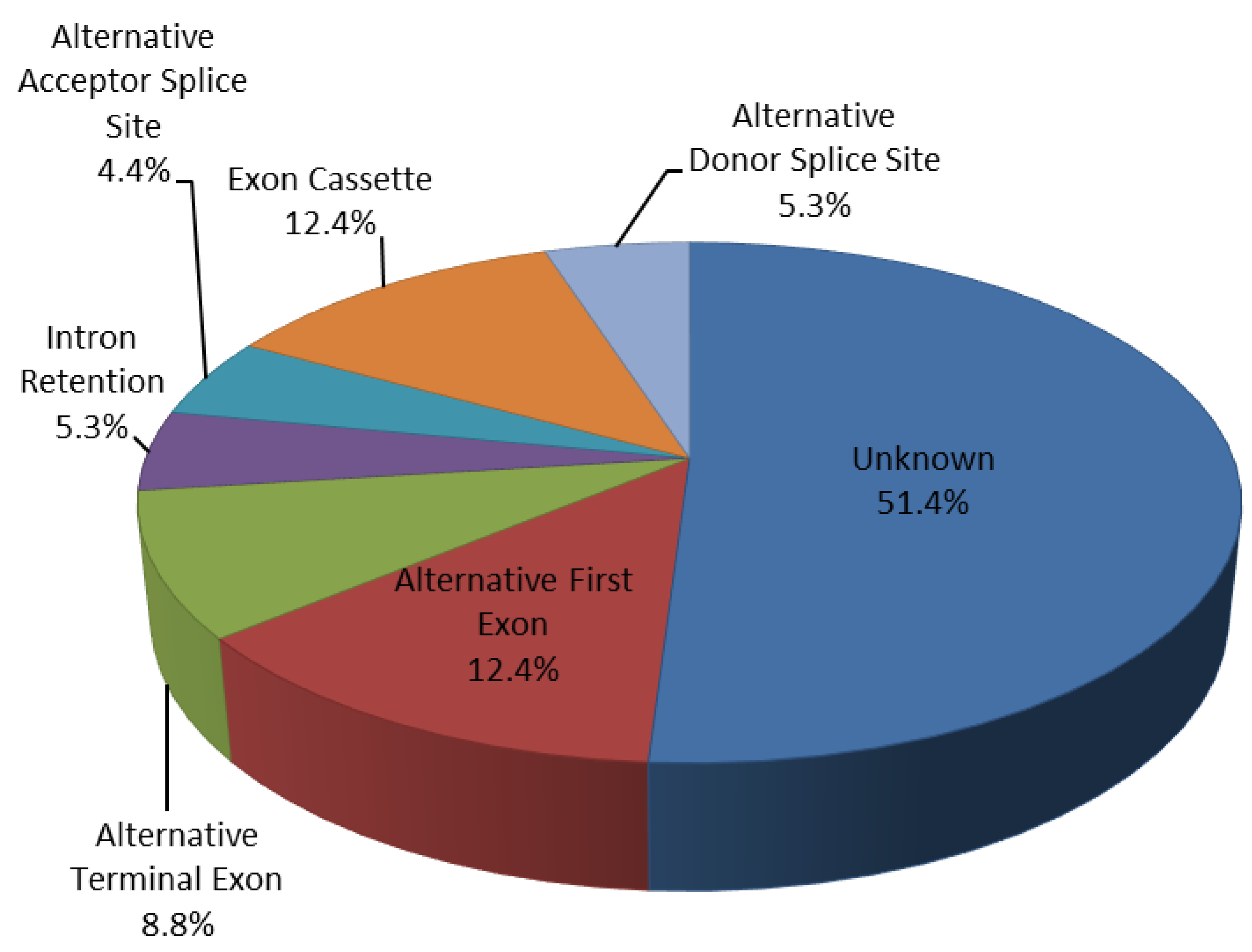
2.5. Regulated Patterns-Exons Analysis
2.6. RECK Splice Variants Analysis by qRT-PCR
- Primer pairs must be designed to discriminate between the two splice variants (Long and Short) of the same gene. Indeed, we have aligned the sequences of the five transcripts using the Clustal Omega software (www.ebi.ac.uk (accessed on 14 June 2021)) and selected the diversity between them, therefore each of the two primers pairs used are specific to amplify the corresponding transcript. For the Long splice variant (transcript variant 1) the primer pairs were designed with the forward (Fw) primer between the exons 17–18 while the reverse (Rev) primer was designed between the exons 18–19 in order to amplify the exon 18 involved in the AS event. For the Short splice variant (transcript variant 5) the primer pairs were designed with the forward (Fw) primer in exon 8 (whose sequence is common between the five transcripts) while the reverse (Rev) primer was designed in exon 9 (whose sequence is specific for this variant 5, Supplementary Figure S1);
- PCR product size must be about 100–230 bp;
- An evaluation on Ensembl Genome Browser (https://www.ensembl.org/index.html (accessed on 14 June 2021)) excluded common SNPs in the primers. Finally, another general requirement considered was the similar annealing temperatures and a balanced G/C content.
2.7. Statistical Analysis
3. Results
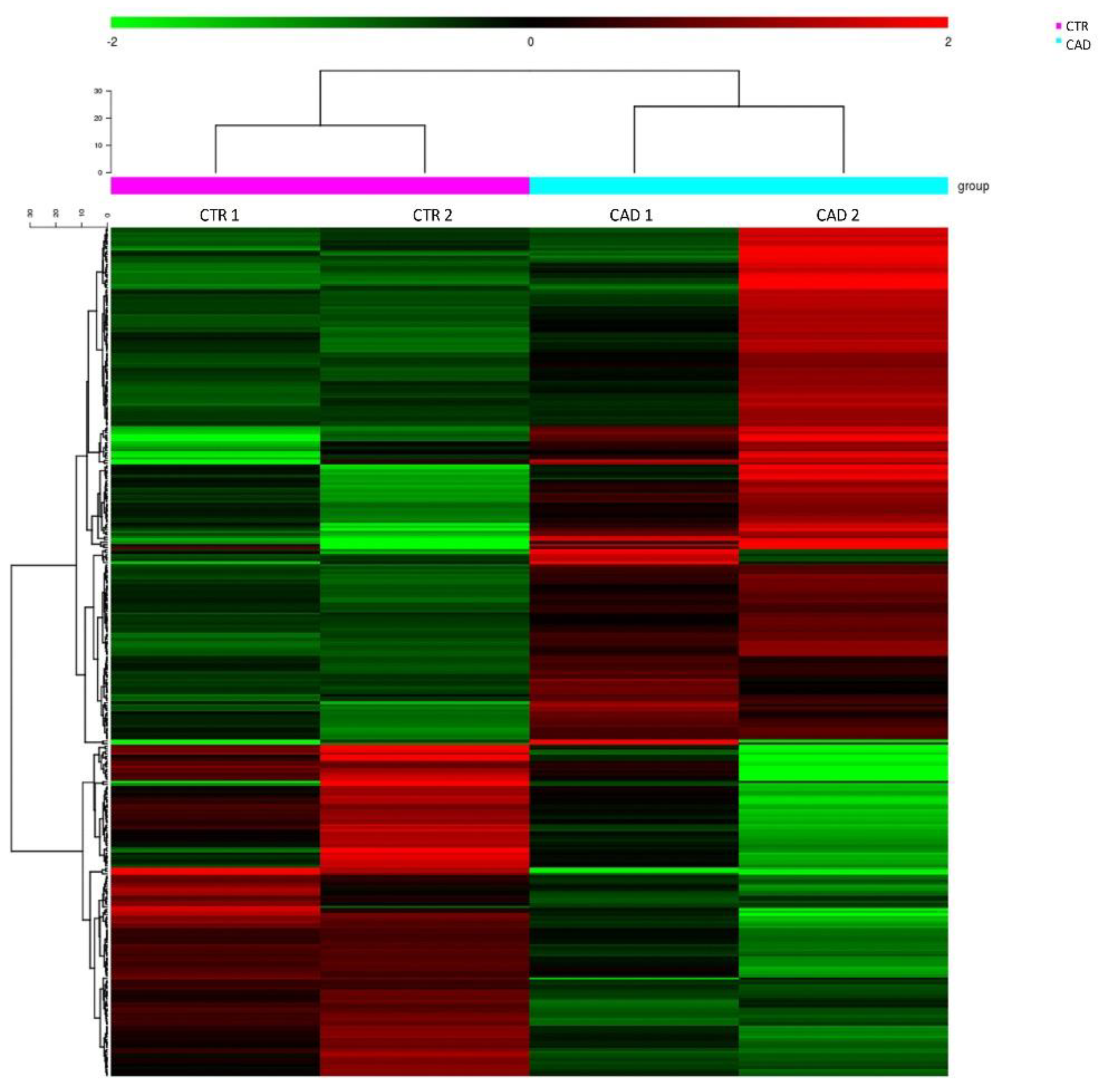
3.1. RNA Sequencing Study
3.2. Clinical Study
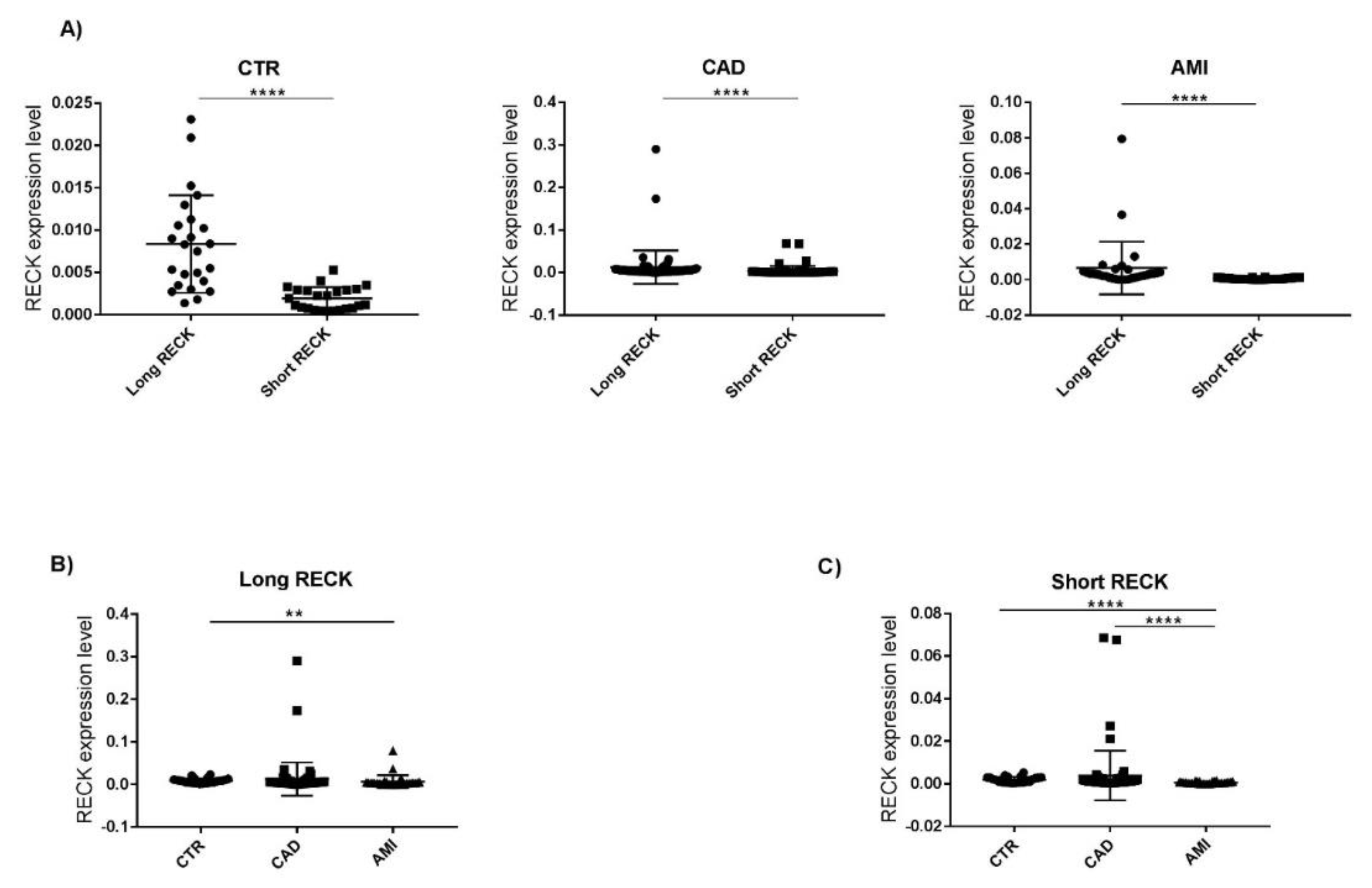
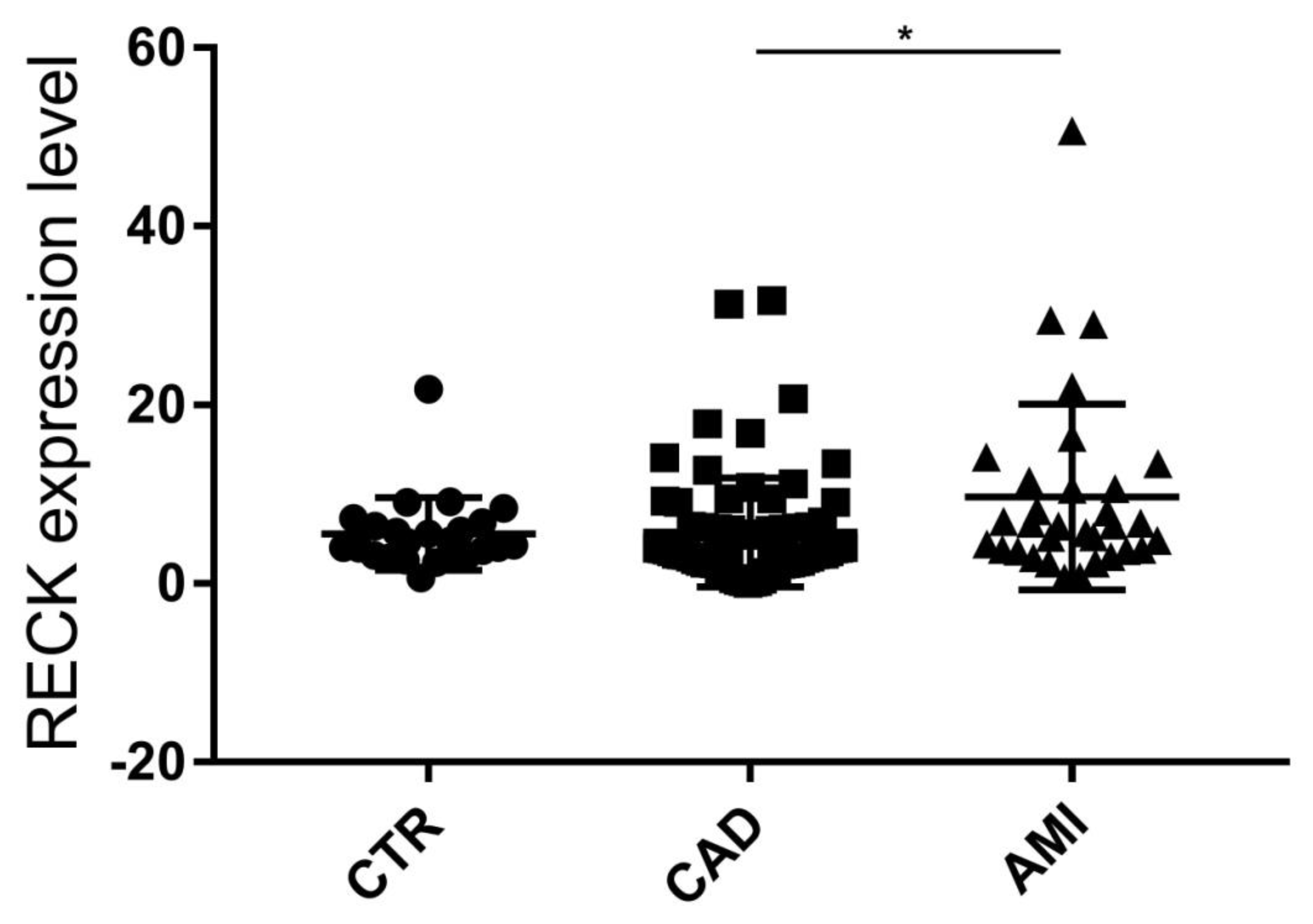
3.3. Evaluation of RECK Splice Variants in PBMCs of All Recruited Patients
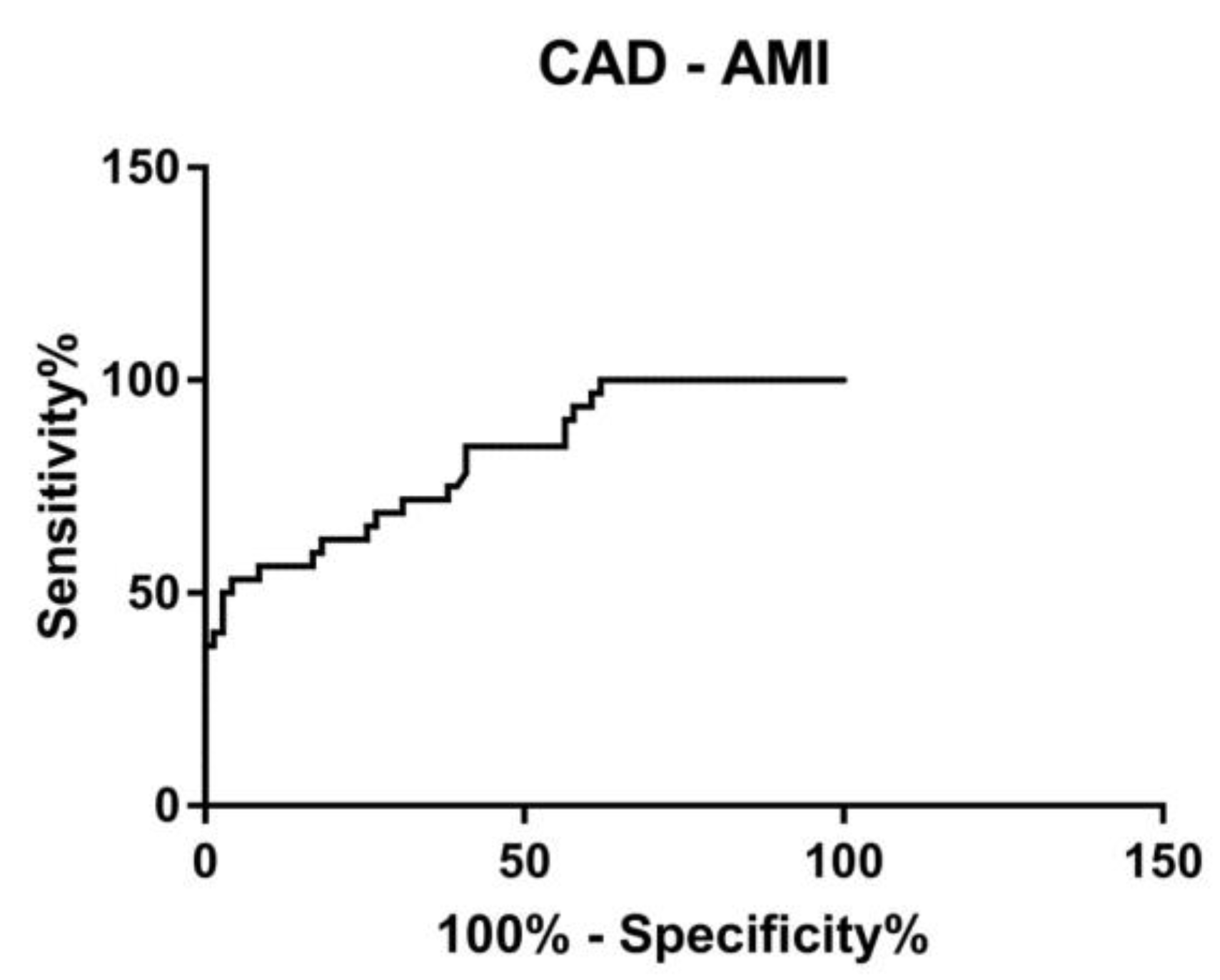
3.4. ROC Curve
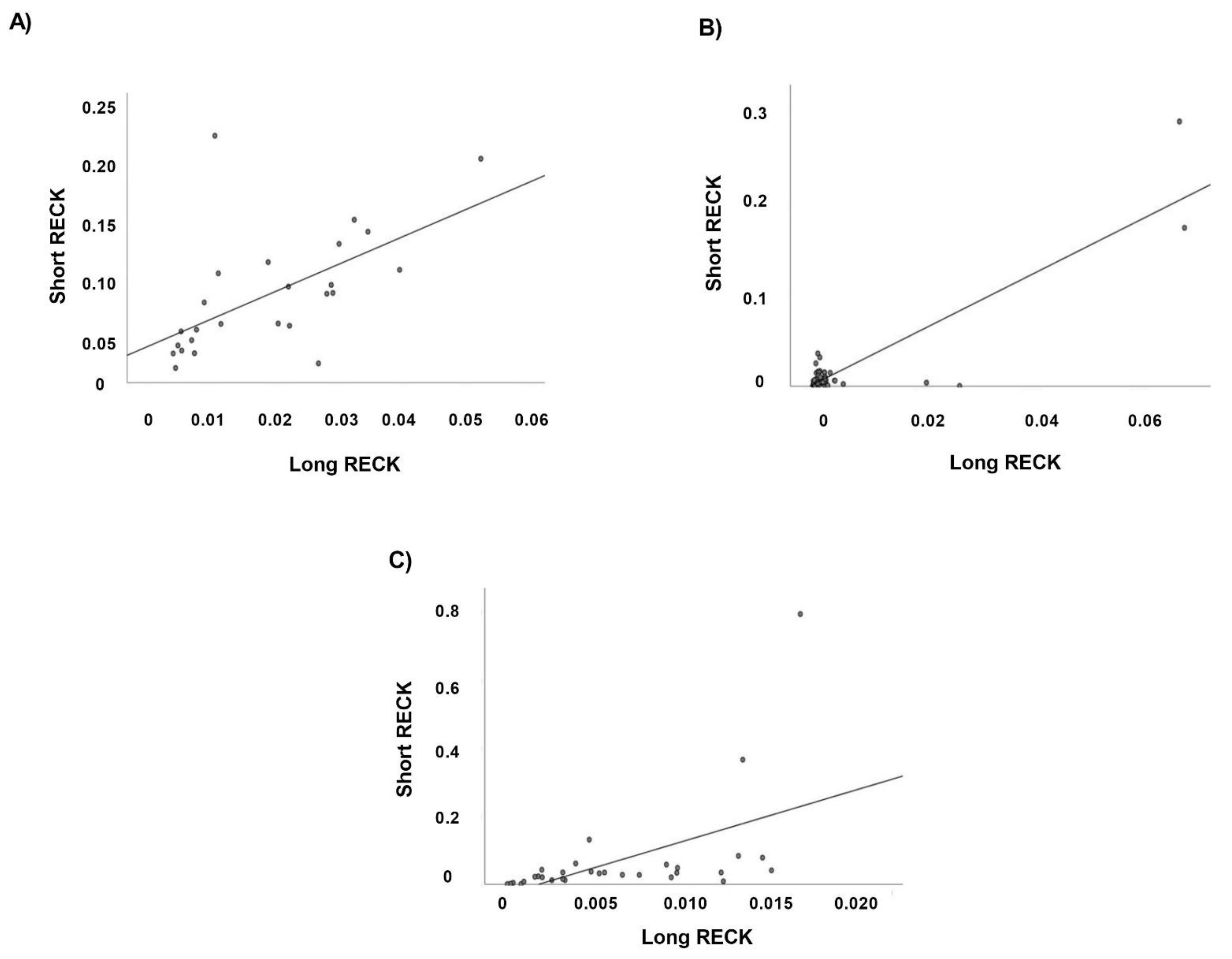
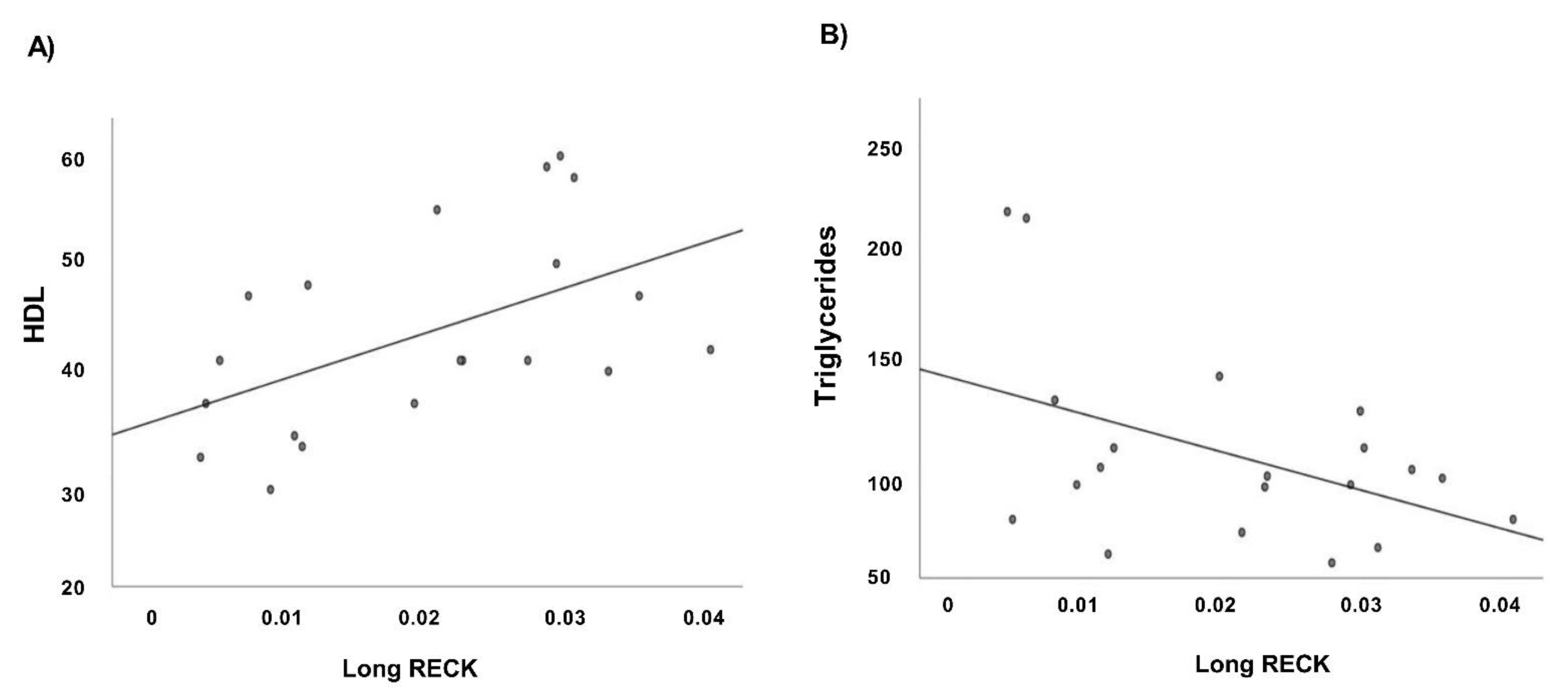
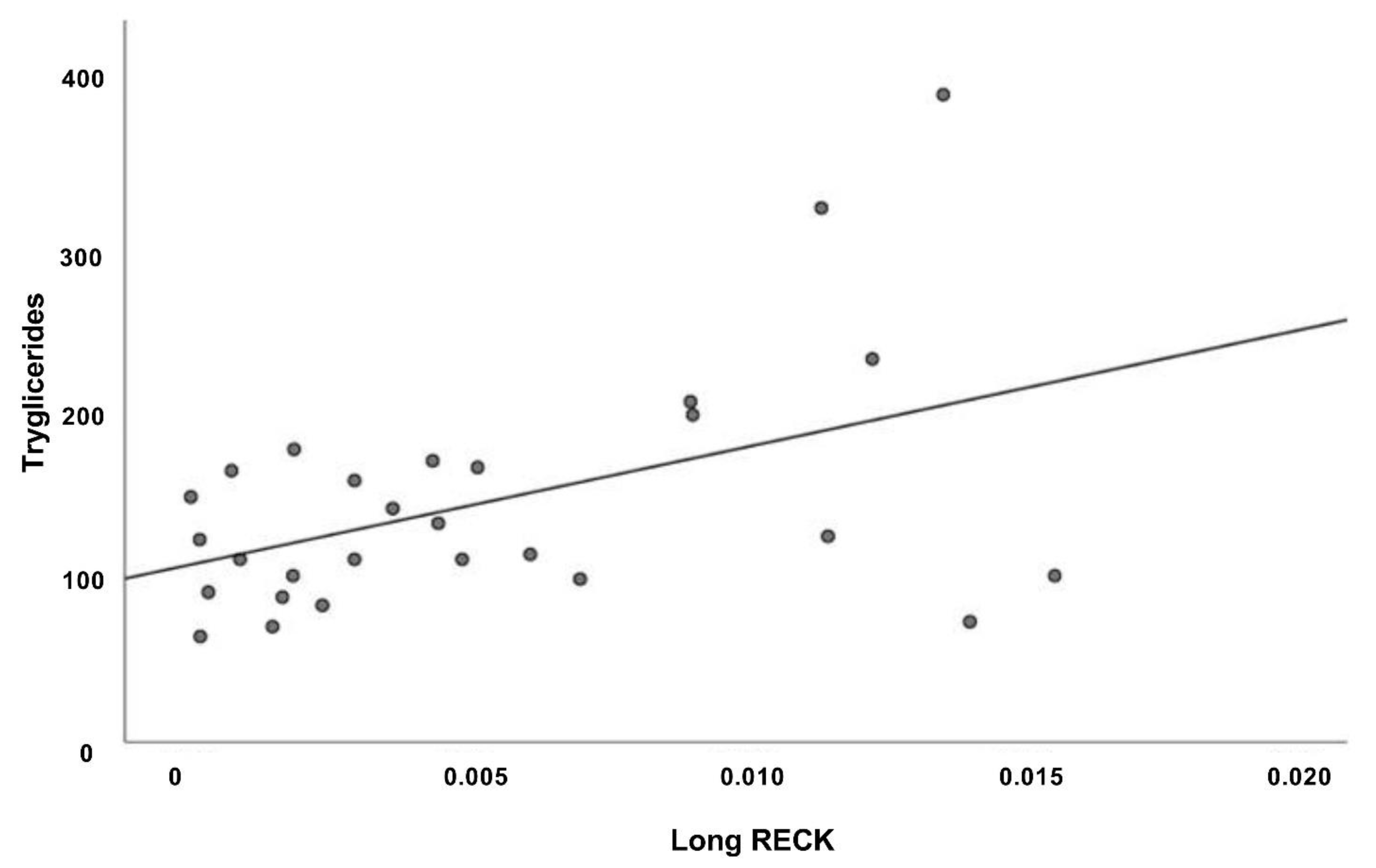
3.5. Correlation Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Roth, G.A.; Mensah, G.A.; Johnson, C.O.; Addolorato, G.; Ammirati, E.; Baddour, L.M.; Barengo, N.C.; Beaton, A.Z.; Benjamin, E.J.; Benziger, C.P.; et al. Global Burden of Cardiovascular Diseases and Risk Factors, 1990–2019: Update from the GBD 2019 Study. J. Am. Coll. Cardiol. 2020, 76, 2982–3021. [Google Scholar] [CrossRef]
- Rizzacasa, B.; Morini, E.; Mango, R.; Vancheri, C.; Budassi, S.; Massaro, G.; Maletta, S.; Macrini, M.; D’Annibale, S.; Romeo, F.; et al. MiR-423 is differentially expressed in patients with stable and unstable coronary artery disease: A pilot study. PLoS ONE 2019, 14, e0216363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Libby, P. Inflammation in Atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 2045–2051. [Google Scholar] [CrossRef] [Green Version]
- Khera, A.V.; Kathiresan, S. Is Coronary Atherosclerosis One Disease or Many? Setting Realistic Expectations for Precision Medicine. Circulation 2017, 135, 1005–1007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, E.; Goren, A.; Ast, G. Alternative splicing: Current perspectives. BioEssays 2008, 30, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.K.; Pham, M.H.C.; Ko, K.S.; Rhee, B.D.; Han, J. Alternative splicing isoforms in health and disease. Eur. J. Physiol. 2018, 470, 995–1016. [Google Scholar] [CrossRef]
- Baralle, F.E.; Giudice, J. Alternative splicing as a regulator of development and tissue identity. Nat. Rev. Mol. Cell Biol. 2017, 18, 437–451. [Google Scholar] [CrossRef] [PubMed]
- Feng, W.; Zhao, P.; Zheng, X.; Hu, Z.; Liu, J. Profiling Novel Alternative Splicing within Multiple Tissues Provides Useful Insights into Porcine Genome Annotation. Genes 2020, 11, 1405. [Google Scholar] [CrossRef]
- Chow, L.T.; Gelinas, R.E.; Broker, T.R.; Roberts, R. An amazing sequence arrangement at the 5′ ends of adenovirus 2 messenger RNA. Cell 1977, 12, 1–8. [Google Scholar] [CrossRef]
- Wang, E.T.; Sandberg, R.; Luo, S.; Khrebtukova, I.; Zhang, L.; Mayr, C.; Kingsmore, S.F.; Schroth, G.P.; Burge, C.B. Alternative isoform regulation in human tissue transcriptomes. Nature 2008, 456, 470–476. [Google Scholar] [CrossRef] [Green Version]
- Bhadra, M.; Howell, P.; Dutta, S.; Heintz, C.; Mair, W.B. Alternative splicing in aging and longevity. Hum. Genet. 2020, 139, 357–369. [Google Scholar] [CrossRef]
- Lara-Pezzi, E.; Gómez-Salinero, J.; Gatto, A.; García-Pavía, P. The Alternative Heart: Impact of Alternative Splicing in Heart Disease. J. Cardiovasc. Transl. Res. 2013, 6, 945–955. [Google Scholar] [CrossRef] [PubMed]
- Kalsotra, A.; Xiao, X.; Ward, A.J.; Castle, J.C.; Johnson, J.M.; Burge, C.B.; Cooper, T.A. A postnatal switch of CELF and MBNL proteins reprograms alternative splicing in the developing heart. Proc. Natl. Acad. Sci. USA 2008, 105, 20333–20338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, S.W.; Hu, Y.W.; Ho, J.W.; Ikeda, S.; Polster, S.; John, R.; Hall, J.L.; Bisping, E.; Pieske, B.; Dos Remedios, C.G.; et al. Heart Failure-Associated Changes in RNA Splicing of Sarcomere Genes. Circ. Cardiovasc. Genet. 2010, 3, 138–146. [Google Scholar] [CrossRef] [PubMed]
- Weeland, C.J.; Hoogenhof, M.V.D.; Beqqali, A.; Creemers, E.E. Insights into alternative splicing of sarcomeric genes in the heart. J. Mol. Cell. Cardiol. 2015, 81, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Rajan, S.; Jagatheesan, G.; Karam, C.N.; Alves, M.L.; Bodi, I.; Schwartz, A.; Bulcao, C.F.; D’Souza, K.M.; Akhter, S.A.; Boivin, G.P.; et al. Molecular and Functional Characterization of a Novel Cardiac-Specific Human Tropomyosin Isoform. Circulation 2010, 121, 410–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, P.; Yong, T.F.; Liang, M.C.; Yue, D.T.; Soong, T.W. Splicing for alternative structures of Cav1.2 Ca2+ channels in cardiac and smooth muscles. Cardiovasc. Res. 2005, 68, 197–203. [Google Scholar] [CrossRef] [Green Version]
- Splawski, I.; Timothy, K.W.; Decher, N.; Kumar, P.; Sachse, F.; Beggs, A.H.; Sanguinetti, M.C.; Keating, M.T. Severe arrhythmia disorder caused by cardiac L-type calcium channel mutations. Proc. Natl. Acad. Sci. USA 2005, 102, 8089–8096. [Google Scholar] [CrossRef] [Green Version]
- Omura, A.; Matsuzaki, T.; Mio, K.; Ogura, T.; Yamamoto, M.; Fujita, A.; Okawa, K.; Kitayama, H.; Takahashi, C.; Sato, C.; et al. RECK Forms Cowbell-shaped Dimers and Inhibits Matrix Metalloproteinase-catalyzed Cleavage of Fibronectin. J. Biol. Chem. 2009, 284, 3461–3469. [Google Scholar] [CrossRef] [Green Version]
- Russell, J.J.; Grisanti, L.A.; Brown, S.M.; Bailey, C.A.; Bender, S.B.; Chandrasekar, B. Reversion inducing cysteine rich protein with Kazal motifs and cardiovascular diseases: The RECKlessness of adverse remodeling. Cell. Signal. 2021, 83, 109993. [Google Scholar] [CrossRef]
- Lee, H.N.; Bosompra, O.A.; Coller, H.A. RECK isoforms differentially regulate fibroblast migration by modulating tubulin post-translational modifications. Biochem. Biophys. Res. Commun. 2019, 510, 211–218. [Google Scholar] [CrossRef]
- Lee, H.N.; Mitra, M.; Bosompra, O.; Corney, D.C.; Johnson, E.L.; Rashed, N.; Ho, L.D.; Coller, H.A. RECK isoforms have opposing effects on cell migration. Mol. Biol. Cell 2018, 29, 1825–1838. [Google Scholar] [CrossRef]
- Siddesha, J.M.; Valente, A.J.; Sakamuri, S.S.; Gardner, J.D.; Delafontaine, P.; Noda, M.; Chandrasekar, B. Acetylsalicylic Acid Inhibits IL-18-Induced Cardiac Fibroblast Migration through the Induction of RECK. J. Cell. Physiol. 2014, 229, 845–855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trombetta-Lima, M.; Assis-Ribas, T.; Cintra, R.C.; Campeiro, J.D.; Guerreiro, J.R.; Winnischofer, S.M.B.; Nascimento, I.C.C.; Ulrich, H.; Hayashi, M.A.F.; Sogayar, M.C. Impact of Reck expression and promoter activity in neuronal in vitro differentiation. Mol. Biol. Rep. 2021, 48, 1985–1994. [Google Scholar] [CrossRef]
- Lima, M.T.; Winnischofer, S.M.B.; Demasi, M.A.A.; Filho, R.A.; Carreira, A.C.O.; Wei, B.; Ribas, T.D.A.; Konig, M.S.; Bowman-Colin, C.; Oba-Shinjo, S.M.; et al. Isolation and characterization of novel RECK tumor suppressor gene splice variants. Oncotarget 2015, 6, 33120–33133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacomasso, T.; Ribas, H.T.; Lima, M.T.; Konig, M.S.; Trindade, E.D.S.; Martinez, G.R.; Sogayar, M.C.; Winnischofer, S.M.B. The alternatively spliced RECK transcript variant 3 is a predictor of poor survival for melanoma patients being upregulated in aggressive cell lines and modulating MMP gene expression in vitro. Melanoma Res. 2020, 30, 223–234. [Google Scholar] [CrossRef]
- Kraemer, R. Regulation of cell migration in atherosclerosis. Curr. Atheroscler. Rep. 2000, 2, 445–452. [Google Scholar] [CrossRef] [PubMed]
- Neglia, D.; Rovai, D.; Caselli, C.; Pietila, M.; Teresinska, A.; Aguadé-Bruix, S.; Pizzi, M.N.; Todiere, G.; Gimelli, A.; Schroeder, S.; et al. Detection of Significant Coronary Artery Disease by Noninvasive Anatomical and Functional Imaging. Circ. Cardiovasc. Imaging 2015, 8, e002179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thygesen, K.; Alpert, J.S.; Jaffe, A.S.; Simoons, M.L.; Chaitman, B.R.; White, H.D.; Katus, H.A.; Apple, F.S.; Lindahl, B.; Morrow, D.A.; et al. Third universal definition of myocardial infarction. Eur. Heart J. 2012, 33, 2551–2567. [Google Scholar] [CrossRef] [Green Version]
- Expert Panel on Detection, Evaluation, and Treatment of High Blood Cholesterol in Adults. Executive summary of the third report of the National Cholesterol Education Program (NCEP) expert panel on detection, evaluation, and treatment of high blood cholesterol in adults (adult treatment panel III). JAMA 2001, 285, 2486–2497. [Google Scholar] [CrossRef]
- Williams, B.; Mancia, G.; Spiering, W.; Rosei, E.A.; Azizi, M.; Burnier, M.; Clement, D.L.; Coca, A.; de Simone, G.; Dominiczak, A.F.; et al. 2018 ESC/ESH Guidelines for the management of arterial hypertension. Eur. Heart J. 2018, 39, 3021–3104. [Google Scholar] [CrossRef] [PubMed]
- American Diabetes Association. 2. Classification and Diagnosis of Diabetes: Standards of Medical Care in Diabetes—2019. Diabetes Care 2019, 42 (Suppl. S1), S13–S28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- De La Grange, P.; Dutertre, M.; Martin, N.; Auboeuf, D. FAST DB: A website resource for the study of the expression regulation of human gene products. Nucleic Acids Res. 2005, 33, 4276–4284. [Google Scholar] [CrossRef] [Green Version]
- Tiano, F.; Amati, F.; Cherubini, F.; Morini, E.; Vancheri, C.; Maletta, S.; Fortuni, S.; Serio, D.; Quatrana, A.; Luffarelli, R.; et al. Frataxin deficiency in Friedreich’s ataxia is associated with reduced levels of HAX-1, a regulator of cardiomyocyte death and survival. Hum. Mol. Genet. 2020, 29, 471–482. [Google Scholar] [CrossRef] [PubMed]
- Kralj, V.; Bilos, I.B. Morbidity and mortality from cardiovascular diseases. Cardiol. Croat. 2013, 8, 373–378. [Google Scholar] [CrossRef]
- Frančula Zaninović, S.; Nola, I.A. Management of Measurable Variable Cardiovascular Disease’ Risk Factors. Curr. Cardiol. Rev. 2018, 14, 153–163. [Google Scholar] [CrossRef]
- European Association for Cardiovascular Prevention & Rehabilitation; Reiner, Z.; Catapano, A.L.; De Backer, G.; Graham, I.; Taskinen, M.R.; Wiklund, O.; Agewall, S.; Alegria, E.; Chapman, M.J.; et al. ESC/EAS Guidelines for the management of dyslipidaemias: The Task Force for the management of dyslipidaemias of the European Society of Cardiology (ESC) and the European Atherosclerosis Society (EAS). Eur. Heart J. 2011, 32, 1769–1818. [Google Scholar]
- Buttar, H.; Li, T.; Ravi, N. Prevention of cardiovascular diseases: Role of exercise, dietary interventions, obesity and smoking cessation. Exp Clin Cardiol. 2005, 10, 229–249. [Google Scholar]
- Piepoli, M.F.; Hoes, A.W.; Agewall, S.; Albus, C.; Brotons, C.; Catapano, A.L.; Cooney, M.T.; Corrà, U.; Cosyns, B.; Deaton, C.; et al. 2016 European Guidelines on cardiovascular disease prevention in clinical practice: The Sixth Joint Task Force of the European Society of Cardiology and Other Societies on Cardiovascular Disease Prevention in Clinical Practice (constituted by representatives of 10 societies and by invited experts) Developed with the special contribution of the European Association for Cardiovascular Prevention & Rehabilitation (EACPR). Eur. Heart J. 2016, 37, 2315–2381. [Google Scholar]
- Cieply, B.; Carstens, R.P. Functional roles of alternative splicing factors in human disease. Wiley Interdiscip. Rev. RNA 2015, 6, 311–326. [Google Scholar] [CrossRef] [Green Version]
- Stastna, M.; Van Eyk, J.E. Analysis of protein isoforms: Can we do it better? Proteomics 2012, 12, 2937–2948. [Google Scholar] [CrossRef] [Green Version]
- Noda, M.; Kitayama, H.; Matsuzaki, T.; Sugimoto, Y.; Okayama, H.; Bassin, R.H.; Ikawa, Y. Detection of genes with a potential for suppressing the transformed phenotype associated with activated ras genes. Proc. Natl. Acad. Sci. USA 1989, 86, 162–166. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, C.; Sheng, Z.; Horan, T.P.; Kitayama, H.; Maki, M.; Hitomi, K.; Kitaura, Y.; Takai, S.; Sasahara, R.M.; Horimoto, A.; et al. Regulation of matrix metalloproteinase-9 and inhibition of tumor invasion by the membrane-anchored glycoprotein RECK. Proc. Natl. Acad. Sci. USA 1998, 95, 13221–13226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kazal, L.A.; Spicer, D.S.; Brahinsky, R.A. Isolation of a Crystalline Trypsin Inhibitor-Anticoagulant Protein from Pancreas. J. Am. Chem. Soc. 1948, 70, 3034–3040. [Google Scholar] [CrossRef] [PubMed]
- Laskowski, M.; Kato, I. Protein Inhibitors of Proteinases. Annu. Rev. Biochem. 1980, 49, 593–626. [Google Scholar] [CrossRef] [PubMed]
- Mendes, S.R.; Del Amo-Maestro, L.; Marino-Puertas, L.; De Diego, I.; Goulas, T.; Gomis-Rüth, F.X. Analysis of the inhibiting activity of reversion-inducing cysteine-rich protein with Kazal motifs (RECK) on matrix metalloproteinases. Sci. Rep. 2020, 10, 6317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutiérrez, J.; Droppelmann, C.A.; Salsoso, R.; Westermeier, F.; Toledo, F.; Salomón, C.; Sanhueza, C.; Pardo, F.; Leiva, A.; Sobrevia, L. A Hypothesis for the Role of RECK in Angiogenesis. Curr. Vasc. Pharmacol. 2016, 14, 106–115. [Google Scholar] [CrossRef]
- De Almeida, G.M.; Yamamoto, M.; Morioka, Y.; Ogawa, S.; Matsuzaki, T.; Noda, M. Critical roles for murine Reck in the regulation of vascular patterning and stabilization. Sci. Rep. 2015, 5, 17860. [Google Scholar] [CrossRef] [Green Version]
- Vitale, C.; Fini, M.; Speziale, G.; Chierchia, S. Gender differences in the cardiovascular effects of sex hormones. Fundam. Clin. Pharmacol. 2010, 24, 675–685. [Google Scholar] [CrossRef]
- Bucholz, E.M.; Beckman, A.L.; Kiefe, C.I.; Krumholz, H.M. Smoking status and life expectancy after acute myocardial infarction in the elderly. Heart 2016, 102, 133–139. [Google Scholar] [CrossRef]
- Mal, K.; Kumar, R.; Ejaz, M.; Fatima, K.; Shaukat, F. Comparison of Lipid Profile in Patients with and without Acute Myocardial Infarction. Cureus 2019, 11, e6467. [Google Scholar] [CrossRef]
- Balci, B. The Modification of Serum Lipids after Acute Coronary Syndrome and Importance in Clinical Practice. Curr. Cardiol. Rev. 2011, 7, 272–276. [Google Scholar] [CrossRef] [Green Version]
- Konstantinou, K.; Tsioufis, C.; Koumelli, A.; Mantzouranis, M.; Kasiakogias, A.; Doumas, M.; Tousoulis, D. Hypertension and patients with acute coronary syndrome: Putting blood pressure levels into perspective. J. Clin. Hypertens. 2019, 21, 1135–1143. [Google Scholar] [CrossRef] [Green Version]
- Majahalme, S.K.; Smith, D.E.; Cooper, J.V.; Kline-Rogers, E.; Mehta, R.H.; Eagle, K.A.; Bisognano, J.D. Comparison of patients with acute coronary syndrome with and without systemic hypertension. Am. J. Cardiol. 2003, 92, 258–263. [Google Scholar] [CrossRef]
- Collet, J.-P.; Thiele, H.; Barbato, E.; Barthélémy, O.; Bauersachs, J.; Bhatt, D.L.; Dendale, P.; Dorobantu, M.; Edvardsen, T.; Folliguet, T.; et al. 2020 ESC Guidelines for the management of acute coronary syndromes in patients presenting without persistent ST-segment elevation. Eur. Heart J. 2021, 42, 1289–1367. [Google Scholar] [CrossRef] [PubMed]
- Kerensky, R.A.; Wade, M.; Deedwania, P.; Boden, W.E.; Pepine, C.J.; Veterans Affairs Non-Q-Wave Infarction Strategies in-Hospital (VANQWISH) Trial Investigators. Revisiting the culprit lesion in non-Q-wave myocardial infarction. Results from the VANQWISH trial angiographic core laboratory. J. Am. Coll. Cardiol. 2002, 39, 1456–1463. [Google Scholar] [CrossRef] [Green Version]
- Park, D.-W.; Clare, R.M.; Schulte, P.J.; Pieper, K.S.; Shaw, L.K.; Califf, R.M.; Ohman, E.M.; Van De Werf, F.; Hirji, S.; Harrington, R.A.; et al. Extent, Location, and Clinical Significance of Non-Infarct-Related Coronary Artery Disease among Patients with ST-Elevation Myocardial Infarction. JAMA 2014, 312, 2019–2027. [Google Scholar] [CrossRef] [PubMed]
- Dixon, W.C.; Wang, T.Y.; Dai, D.; Shunk, K.A.; Peterson, E.D.; Roe, M.T.; Registry, N.C.D. Anatomic Distribution of the Culprit Lesion in Patients with Non-ST-Segment Elevation Myocardial Infarction Undergoing Percutaneous Coronary Intervention: Findings from the National Cardiovascular Data Registry. J. Am. Coll. Cardiol. 2008, 52, 1347–1348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mummidi, S.; Das, N.A.; Carpenter, A.J.; Yoshida, T.; Yariswamy, M.; Mostany, R.; Izadpanah, R.; Higashi, Y.; Sukhanov, S.; Noda, M.; et al. RECK suppresses interleukin-17/TRAF3IP2-mediated MMP-13 activation and human aortic smooth muscle cell migration and proliferation. J. Cell. Physiol. 2019, 234, 22242–22259. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.; Stone, N.J.; Ballantyne, C.; Bittner, V.; Criqui, M.H.; Ginsberg, H.N.; Goldberg, A.C.; Howard, W.J.; Jacobson, M.S.; Kris-Etherton, P.M.; et al. Triglycerides and cardiovascular disease: A scientific statement from the American Heart Association. Circulation 2011, 123, 2292–2333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reiner, Ž. Hypertriglyceridaemia and risk of coronary artery disease. Nat. Rev. Cardiol. 2017, 14, 401–411. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Accession Number | Sequence (5′-3′) | Annealing Temperature (°C) | Size (bp) | |
|---|---|---|---|---|---|
| RECK (Short splice variant) | NM_001316348 | Fw | GAACAGACTCTTCTCCTGGT | 56 | 208 |
| Rev | AGATATCAGGCTCTCTTCTCA | ||||
| RECK (Long splice variant) | NM_021111 | Fw | CCAGCCCTTTTGCAGAGCA | 60 | 221 |
| Rev | AAGCACCCGGTGGGATGAT | ||||
| GAPDH | NM_002046 | Fw | AAGGTCGGAGTCAACGGATTT | 59 | 100 |
| Rev | TGAAGGGGTCATTGATGGCA | ||||
| Alternative Event Type | N. of AS Regulated Events |
|---|---|
| Alternative First Exon | 14 |
| Alternative Terminal Exon | 10 |
| Exon Cassette | 14 |
| Alternative Acceptor Splice Site | 5 |
| Alternative Donor Splice Site | 6 |
| Intron Retention | 6 |
| unknown | 58 |
| Gene Symbol | Accession Number | Alternative Event Type | Involved Exon | Regulation Splicing Index | Splicing-Index Fold-Change | Splicing-Index p-Value | Event Coordinates (hg19) |
|---|---|---|---|---|---|---|---|
| CFAP44 | NM_001164496 | unknown | e25 | up | 3.51 | 4.87 × 10−2 | chr3:113052252-113052429 |
| PLCB2 | NM_004573 | unknown | i31 | down | 3.23 | 9.14 × 10−4 | chr15:40581119-40581471 |
| RECK | NM_021111 | unknown | e18 | down | 2.64 | 2.17 × 10−2 | chr9:36118754-36118964 |
| VTI1A | NM_145206 | unknown | i1 | up | 2.63 | 6.80 × 10−3 | chr10:114208248-114208639 |
| CD58 | NM_001779 | Alternative Terminal Exon | ae5 | down | 2.45 | 3.91 × 10−2 | chr1:117061321-117061851 |
| MIR4469 // RNF170 | NM_030954 | Alternative Terminal Exon | ae6 | down | 2.29 | 2.93 × 10−2 | chr8:42716503-42716886 |
| PTER | NM_001001484 | Exon Cassette | e2 | down | 2.21 | 2.41 × 10−2 | chr10:16479356-16479489 |
| GPATCH2L | NM_017926 | Alternative Terminal Exon | ae9 | down | 2.19 | 4.61 × 10−2 | chr14:76662316-76662739 |
| CLEC12A | NM_138337 | unknown | i1 | down | 2.13 | 1.23 × 10−2 | chr12:10124287-10131564 |
| BLNK | NM_013314 | Exon Cassette | e16 | down | 2.13 | 3.31 × 10−2 | chr10:97956663-97956735 |
| CTR Subjects | CAD Patients | AMI Patients | p-Value | |
|---|---|---|---|---|
| Age (years) | 67.5 ± 9.3 | 66.6 ± 9.8 | 62.2 ± 13 | n.s. |
| Gender | ||||
| Male (%) | 66.6 | 84.7 | 90.6 | * p < 0.05 |
| Ɨ p < 0.05 | ||||
| Hypertension (%) | 69.6 | 77.5 | 53.1 | ǂ p < 0.05 |
| Diabetes (%) | 21.7 | 39.4 | 28.1 | n.s. |
| Dyslipidemia (%) | 47.8 | 88.7 | 46.9 | * p < 0.0005 |
| ǂ p < 0.0005 | ||||
| Smoking history | ||||
| Present (%) | 17.4 | 23.9 | 75 | Ɨ p < 0.0005 |
| ǂ p < 0.0005 | ||||
| Past (%) | 43.5 | 39.4 | 3.1 | Ɨ p < 0.0005 |
| ǂ p < 0.0005 | ||||
| Number of affected vessels | ||||
| 1 vessel disease (%) | 0 | 45.1 | 46.9 | * p < 0.0005 |
| Ɨ p < 0.0005 | ||||
| 2 vessel disease (%) | 0 | 29.6 | 28.1 | * p < 0.005 |
| Ɨ p < 0.005 | ||||
| 3 vessel disease (%) | 0 | 25.4 | 24.2 | * p < 0.005 |
| Ɨ p < 0.005 | ||||
| Type of affected vessel | ||||
| LAD (%) | 0 | 54.9 | 62.5 | * p < 0.0005 |
| Ɨ p < 0.0005 | ||||
| CFX (%) | 0 | 31 | 37.6 | * p < 0.0005 |
| Ɨ p < 0.0005 | ||||
| RCA (%) | 0 | 38 | 5 | * p < 0.0005 |
| ǂ p < 0.0005 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vancheri, C.; Morini, E.; Prandi, F.R.; Alkhoury, E.; Celotto, R.; Romeo, F.; Novelli, G.; Amati, F. Two RECK Splice Variants (Long and Short) Are Differentially Expressed in Patients with Stable and Unstable Coronary Artery Disease: A Pilot Study. Genes 2021, 12, 939. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12060939
Vancheri C, Morini E, Prandi FR, Alkhoury E, Celotto R, Romeo F, Novelli G, Amati F. Two RECK Splice Variants (Long and Short) Are Differentially Expressed in Patients with Stable and Unstable Coronary Artery Disease: A Pilot Study. Genes. 2021; 12(6):939. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12060939
Chicago/Turabian StyleVancheri, Chiara, Elena Morini, Francesca Romana Prandi, Elie Alkhoury, Roberto Celotto, Francesco Romeo, Giuseppe Novelli, and Francesca Amati. 2021. "Two RECK Splice Variants (Long and Short) Are Differentially Expressed in Patients with Stable and Unstable Coronary Artery Disease: A Pilot Study" Genes 12, no. 6: 939. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12060939