
Comparison of the Transcriptomic and Epigenetic Profiles of Gonadal Primordial Germ Cells of White Leghorn and Green-Legged Partridgelike Chicken Embryos


Abstract
:1. Introduction
2. Materials and Methods
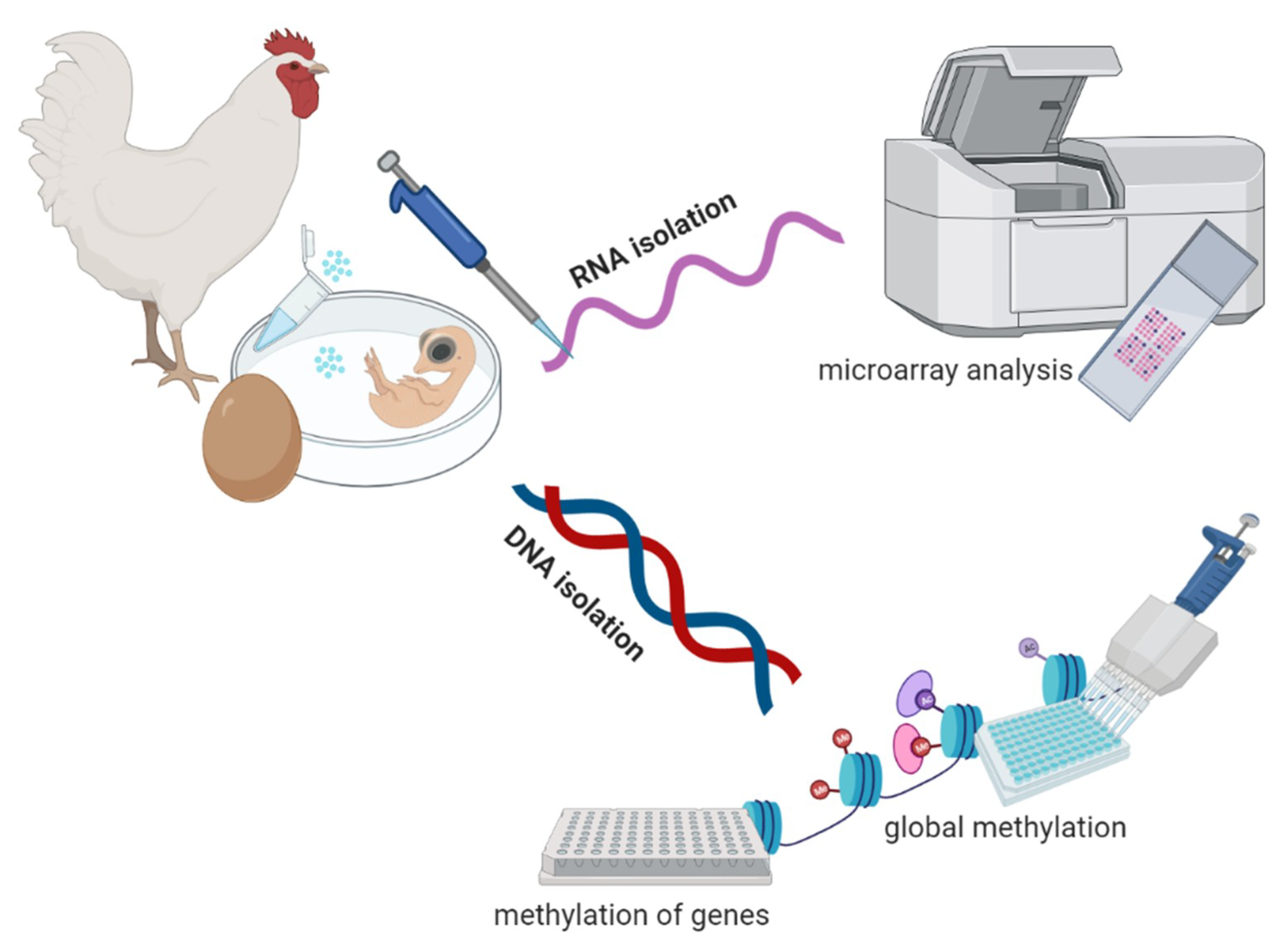
2.1. Gonadal PGC Isolation
2.2. RNA and DNA Isolation
2.3. Sex Determination of Embryos
2.4. Transcriptome Analysis
2.5. Global Methylation Analysis
2.6. Gene-Specific Methylation Analysis
3. Results
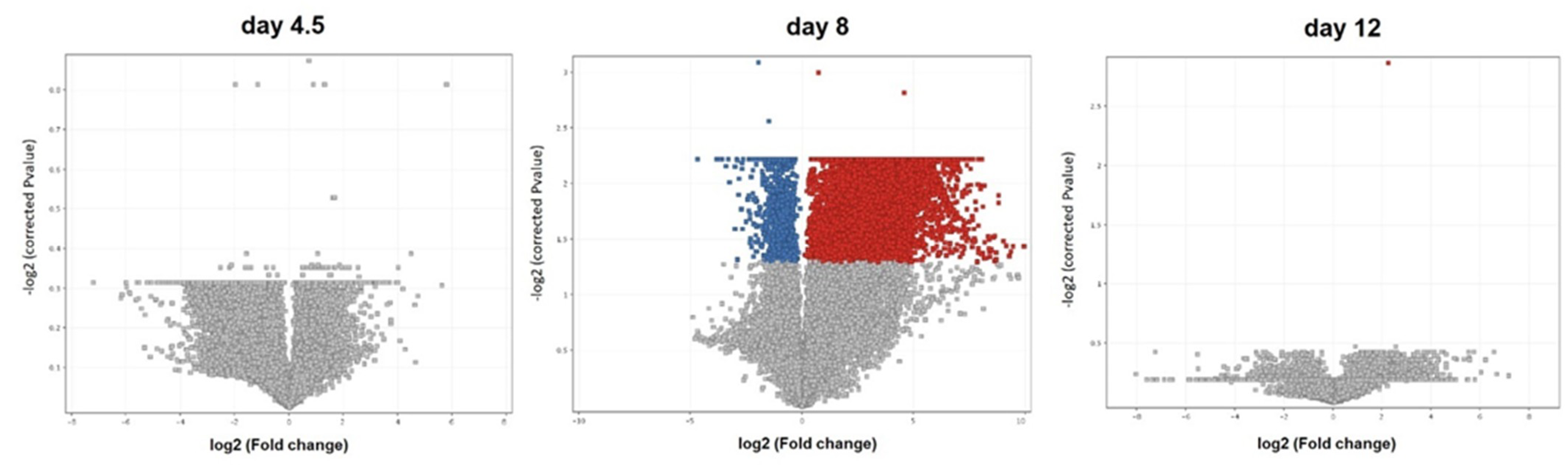
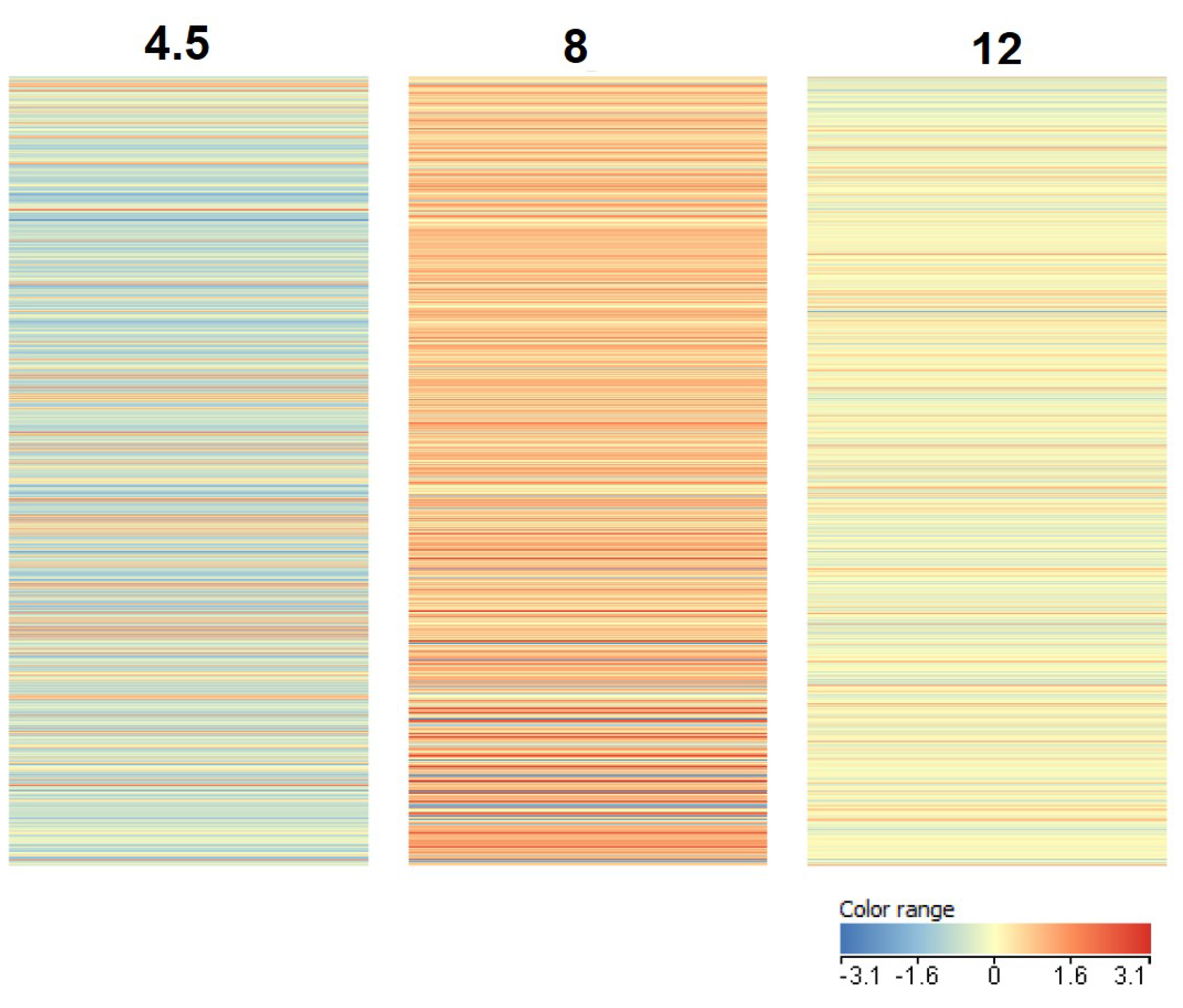
3.1. Transcriptome Analysis
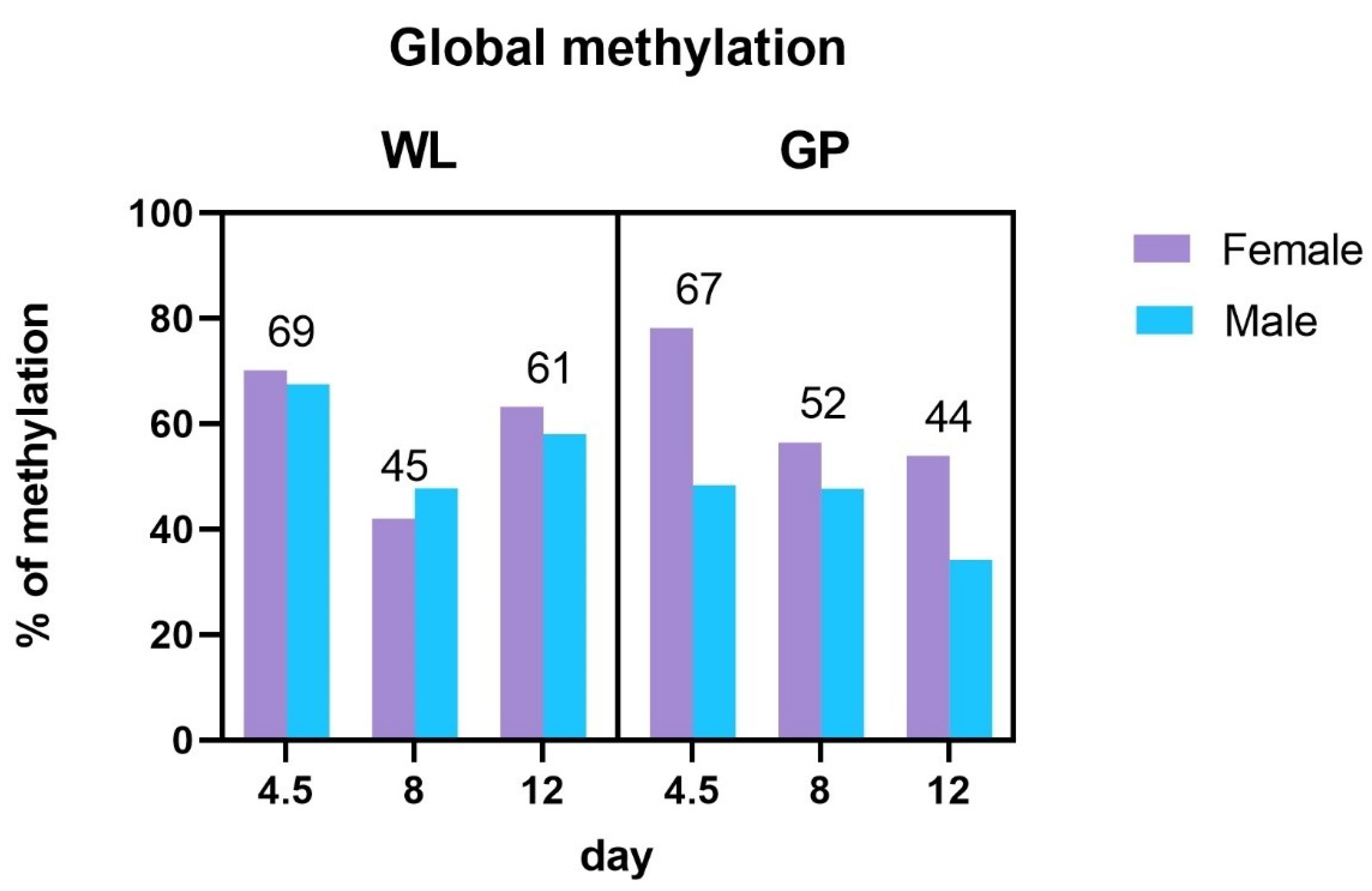
3.2. Global Methylation Analysis
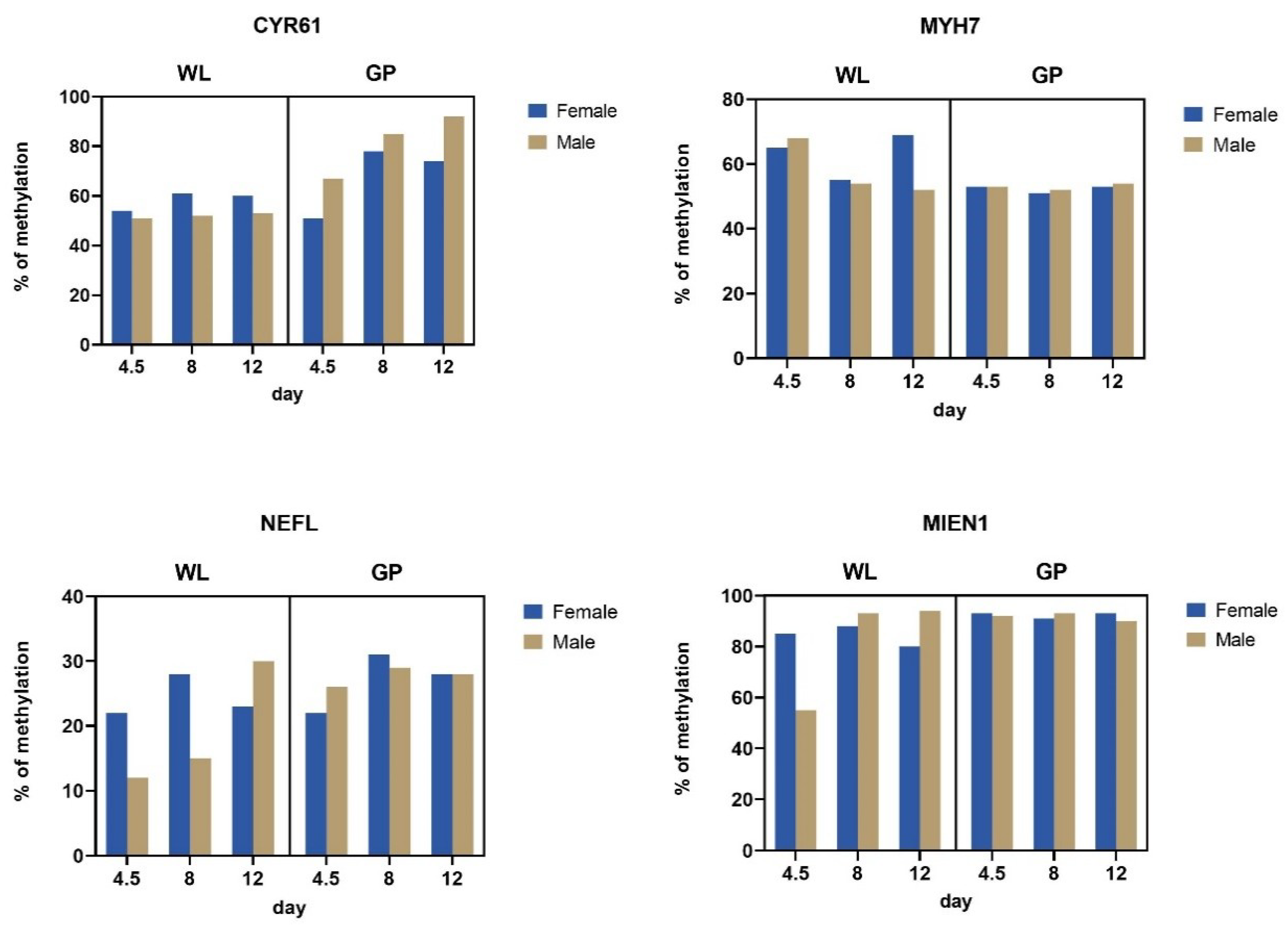
3.3. Methylation of Silenced Genes
4. Discussion
4.1. Transcriptome Analysis
4.2. Global Methylation Analysis
4.3. Gene-Specific Methylation Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Buzała, M.; Janicki, B.; Czarnecki, R. Consequences of different growth rates in broiler breeder and layer hens on embryogenesis, metabolism and metabolic rate: A review. Poult. Sci. 2014, 94, 728–733. [Google Scholar] [CrossRef]
- Bednarczyk, M.; Rosinski, A. Comparison of egg hatchability and in vitro survival of goose embryos of various origins. Poult. Sci. 1999, 78, 579–585. [Google Scholar] [CrossRef]
- Sawicka, D.; Samek, K.; Chojnacka-Puchta, L.; Witkowski, A.; Knaga, S.; Debowska, M.; Bednarczyk, M. Changes in quail blastodermal cell status as a result of selection. Folia Biol. 2015, 63, 63–67. [Google Scholar] [CrossRef] [PubMed]
- Sadeghi, R.; Pakdel, M.; Moradi Shabrbabak, M.; Moradi Shabrbabak, H. Divergent Selection Effect on Reproductive Trait in Japanese Quails. Res. J. Poult. Sci. 2013, 6, 18–22. [Google Scholar]
- Rashidi, H.; Sottile, V. The chick embryo: Hatching a model for contemporary biomedical research. BioEssays 2009, 31, 459–465. [Google Scholar] [CrossRef]
- Weeke-Klimp, A.; Bax, N.A.M.; Bellu, A.R.; Winter, E.M.; Vrolijk, J.; Plantinga, J.; Maas, S.; Brinker, M.; Mahtab, E.A.F.; Gittenberger-de Groot, A.C.; et al. Epicardium-derived cells enhance proliferation, cellular maturation and alignment of cardiomyocytes. J. Mol. Cell. Cardiol. 2010, 49, 606–616. [Google Scholar] [CrossRef] [PubMed]
- Cogburn, L.A.; Wang, X.; Carre, W.; Rejto, L.; Porter, T.E.; Aggrey, S.E.; Simon, J. Systems-wide chicken DNA microarrays, gene expression profiling, and discovery of functional genes. Poult. Sci. 2003, 82, 939–951. [Google Scholar] [CrossRef] [PubMed]
- Ribatti, D. Chicken chorioallantoic membrane angiogenesis model. Methods Mol. Biol. 2012, 843, 47–57. [Google Scholar]
- Dunislawska, A.; Slawinska, A.; Siwek, M. Hepatic DNA Methylation in Response to Early Stimulation of Microbiota with Lactobacillus Synbiotics in Broiler Chickens. Genes 2020, 11, 579. [Google Scholar] [CrossRef] [PubMed]
- Ginsburg, M.; Eyal-Giladi, H. Primordial Germ Cells of the Young Chick Blastoderm Originate from the Central Zone of the Area Pellucida Irrespective of the Embryo-Forming Process. Development 1987, 101, 209–219. [Google Scholar] [CrossRef]
- Chojnacka-Puchta, L.; Kasperczyk, K.; Płucienniczak, G.; Sawicka, D.; Bednarczyk, M. Primordial germ cells (PGCs) as a tool for creating transgenic chickens. Pol. J. Vet. Sci. 2012, 15, 181–188. [Google Scholar] [CrossRef] [Green Version]
- Mochizuki, K.; Matsui, Y. Epigenetic profiles in primordial germ cells: Global modulation and fine tuning of the epigenome for acquisition of totipotency. Dev. Growth Differ. 2010, 52, 517–525. [Google Scholar] [CrossRef]
- Dunislawska, A.; Szczerba, A.; Siwek, M.; Bednarczyk, M. Dynamics of the transcriptome during chicken embryo development based on primordial germ cells. BMC Res. Notes 2020, 13, 441. [Google Scholar] [CrossRef] [PubMed]
- Jang, H.J.; Seo, H.W.; Lee, B.R.; Yoo, M.; Womack, J.E.; Han, J.Y. Gene expression and DNA methylation status of chicken primordial germ cells. Mol. Biotechnol. 2013, 54, 177–186. [Google Scholar] [CrossRef]
- Nakajima, Y.; Minematsu, T.; Naito, M.; Tajima, A. A New Method for Isolating Viable Gonadal Germ Cells from 7-day-old Chick Embryos. J. Poult. Sci. 2011, 48, 106–111. [Google Scholar] [CrossRef] [Green Version]
- Hamburger, V.; Hamilton, H.L. A series of normal stages in the development of the chick embryo. J. Morphol. 1951, 88, 49–92. [Google Scholar] [CrossRef] [PubMed]
- Clinton, M.; Haines, L.; Belloir, B.; McBride, D. Sexing chick embryos: A rapid and simple protocol. Br. Poult. Sci. 2001, 42, 134–138. [Google Scholar] [CrossRef] [PubMed]
- Jawor, M.; Knaga, S.; Kozłowska, I.; Barna, J.; Váradi, É.; Kasperek, K.; Drobnyák, Á.; Bodzsár, N.; Várkonyi, E.P.; Jeżewska-Witkowska, G.; et al. Population Structure of Four Indigenous Chicken Breeds Undergoing In Situ Conservation. Anim. Sci. Pap. 2020, 38, 167–179. [Google Scholar]
- Szczerba, A.; Kuwana, T.; Bednarczyk, M. Concentration and total number of circulating primordial germ cells in Green-legged Partridgelike chicken embryos. Poult. Sci. 2021, 100, 319–324. [Google Scholar] [CrossRef]
- Siwek, M.; Wragg, D.; Sławińska, A.; Malek, M.; Hanotte, O.; Mwacharo, J.M. Insights into the genetic history of Green-legged Partridgelike fowl: MtDNA and genome-wide SNP analysis. Anim. Genet. 2013, 44, 522–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bednarczyk, M.; Lakota, P.; Slomski, R.; Plawski, A.; Lipinski, D.; Siemieniako, B.; Lisowski, M.; Czekalski, P.; Grajewski, B.; Dluzniewska, P. Reconstitution of a chicken breed by inter se mating of germline chimeric birds. Poult. Sci. 2002, 81, 1347–1353. [Google Scholar] [CrossRef]
- Kuwana, T.; Kawashima, T.; Naito, M.; Yamashita, H.; Matsuzaki, M.; Takano, T. Conservation of a Threatened Indigenous Fowl (Kureko Dori) Using the Germline Chimeras Transplanted from Primordial Germ Cells. J. Poult. Sci. 2006, 43, 60–66. [Google Scholar] [CrossRef] [Green Version]
- Kostaman, T.; Yusuf, T.L.; Fahrudin, M.; Setiadi, M.A. Isolation and number of circulated primordial germ cells (circulated-PGCs) on stages of embryonic development of Gaok chicken. Jurnal Ilmu Ternak dan Veteriner 2013, 18, 54–62. [Google Scholar] [CrossRef]
- Sopiyana, S.; Setiadi, M.A.; Fahrudin, M.; Supriatna, I. Isolation and number of gonadal primordial germ cells (gonadal PGCs) on the stages of early embryonic development of KUB chicken. Media Peternak. 2017, 40, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Zhao, D.-F.; Yamashita, H.; Matsuzaki, M.; Takano, T.; Abe, S.-I.; Naito, M.; Kuwana, T. Genetic Factors Affect the Number of Circulating Primordial Germ Cells in Early Chick Embryos. J. Poult. Sci. 2003, 40, 101–113. [Google Scholar] [CrossRef] [Green Version]
- Chojnacka-Puchta, L.; Sawicka, D.; Lakota, P.; Plucienniczak, G.; Bednarczyk, M.; Plucienniczak, A. Obtaining chicken primordial germ cells used for gene transfer: In vitro and in vivo results. J. Appl. Genet. 2015, 56, 493–504. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.M.; Han, J.Y. The early development of germ cells in chicken. Int. J. Dev. Biol. 2018, 62, 145–152. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Zhu, Y.; Zhi, L.; Han, X.; Shen, J.; Liu, Y.; Yao, J.; Yang, X. DNA Methylation Variation Trends during the Embryonic Development of Chicken. PLoS ONE 2016, 11, e0159230. [Google Scholar] [CrossRef] [Green Version]
- Messerschmidt, D.M.; Knowles, B.B.; Solter, D. DNA methylation dynamics during epigenetic reprogramming in the germline and preimplantation embryos. Genes Dev. 2014, 28, 812–828. [Google Scholar] [CrossRef] [Green Version]
- Gryzinska, M.; Blaszczak, E.; Strachecka, A.; Jezewska-Witkowska, G. Analysis of age-related global DNA methylation in chicken. Biochem. Genet. 2013, 51, 554–563. [Google Scholar] [CrossRef] [Green Version]
- Romanov, G.A.; Vanyushin, B.F. Methylation of reiterated sequences in mammalian DNAs Effects of the tissue type, age, malignancy and hormonal induction. BBA Sect. Nucleic Acids Protein Synth. 1981, 653, 204–218. [Google Scholar] [CrossRef]
- Dunislawska, A.; Slawinska, A.; Gryzinska, M.; Siwek, M. Interaction between early in ovo stimulation of the gut microbiota and chicken host—Splenic changes in gene expression and methylation. J. Anim. Sci. Biotechnol. 2021, 12, 73. [Google Scholar] [CrossRef]
- Piprek, R.P.; Kolasa, M.; Podkowa, D.; Kloc, M.; Kubiak, J.Z. Transcriptional profiling validates involvement of extracellular matrix and proteinases genes in mouse gonad development. Mech. Dev. 2018, 149, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Dunislawska, A.; Slawinska, A.; Bednarczyk, M.; Siwek, M. Transcriptome modulation by in ovo delivered Lactobacillus synbiotics in a range of chicken tissues. Gene 2019, 698, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Martinsen, B.J. Reference guide to the stages of chick heart embryology. Dev. Dyn. 2005, 233, 1217–1237. [Google Scholar] [CrossRef] [PubMed]
- Karunanithi, S.; Venkatachalapathy, L.; Sudhalakshmi, Y. Comparative In Silico Analysis of Hypertrophic Cardiomyopathy Heart in Human and Normal Chicken Heart. J. Adv. Lab. Res. Biol. 2010, 1, 60–63. [Google Scholar]
- Xu, Z.; Che, T.; Li, F.; Tian, K.; Zhu, Q.; Mishra, S.K.; Dai, Y.; Li, M.; Li, D. The temporal expression patterns of brain transcriptome during chicken development and ageing. BMC Genom. 2018, 19, 917. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene (NCBI No.) | Primer Sequences | GC% | Amplicon Size | Reference | |
|---|---|---|---|---|---|
| CYR61 (429089) | M | F: TTTGGTTTTAGTGTTTAAAGACGT R: TTATATTTACCTTCAAAAAAACGTA | 58.33 44.00 | 150 | [9] |
| U | F: TTTTGGTTTTAGTGTTTAAAGATGT R: TATTTATATTTACCTTCAAAAAAACATA | 56.00 42.86 | 154 | ||
| MYH7 (395350) | M | F: AGGGTTTTGTTTCGTGTTTTATTC R: CTCCCCCATCTCTATAATAACGAT | 70.83 62.50 | 100 | This study |
| U | F: GGGTTTTGTTTTGTGTTTTATTTGT R: CCTCCCCCATCTCTATAATAACAAT | 76.00 64.00 | 100 | ||
| NEFL (419528) | M | F: TTTTTTGTATTCGGTGGATAGTTTC R: TAAAATCCTACAACTAAACCCGCT | 68.00 70.83 | 104 | This study |
| U | F: TTTTTGTATTTGGTGGATAGTTTTG R: TTAAAATCCTACAACTAAACCCACT | 68.00 68.00 | 104 | ||
| MIEN1 (100858225) | M | F: GGGGTAGTTGAGAGTTATACGT R:TACAAAATAATACAAAAAAAACGAC | 68.18 68.00 | 125 | This study |
| U | F: TGTGGGGTAGTTGAGAGTTATATGT R: TACAAAATAATACAAAAAAAACAAC | 64.00 68.00 | 128 | ||
| Breed | Day | CYR61 | MYH7 | NEFL | MIEN1 |
|---|---|---|---|---|---|
| WL | 8 vs. 4.5 | –3.26 | –2.37 | nd | nd |
| 12 vs. 4.5 | –2.34 | –3.91 | –4.19 | –2.52 | |
| GP | 8 vs. 4.5 | –1.78 | 1.30 | 0.37 | nd |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dunislawska, A.; Siwek, M.; Stadnicka, K.; Bednarczyk, M. Comparison of the Transcriptomic and Epigenetic Profiles of Gonadal Primordial Germ Cells of White Leghorn and Green-Legged Partridgelike Chicken Embryos. Genes 2021, 12, 1090. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12071090
Dunislawska A, Siwek M, Stadnicka K, Bednarczyk M. Comparison of the Transcriptomic and Epigenetic Profiles of Gonadal Primordial Germ Cells of White Leghorn and Green-Legged Partridgelike Chicken Embryos. Genes. 2021; 12(7):1090. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12071090
Chicago/Turabian StyleDunislawska, Aleksandra, Maria Siwek, Katarzyna Stadnicka, and Marek Bednarczyk. 2021. "Comparison of the Transcriptomic and Epigenetic Profiles of Gonadal Primordial Germ Cells of White Leghorn and Green-Legged Partridgelike Chicken Embryos" Genes 12, no. 7: 1090. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12071090