
Autophosphorylation and Self-Activation of DNA-Dependent Protein Kinase


1
Graduate School of Science and Engineering, Gunma University, Kiryu 376-8515, Japan
2
Food and Health Science Education and Research Center, Gunma University, Kiryu 376-8515, Japan
Genes 2021, 12(7), 1091; https://0-doi-org.brum.beds.ac.uk/10.3390/genes12071091
Submission received: 15 June 2021
/
Revised: 11 July 2021
/
Accepted: 17 July 2021
/
Published: 19 July 2021
(This article belongs to the Special Issue Role of ATM and MRE11 in Genomic Stability and Oxidative Stress Responses)
{kind=link}
{kind=link}
{kind=link}
Abstract
:The DNA-dependent protein kinase catalytic subunit (DNA-PKcs), a member of the phosphatidylinositol 3-kinase-related kinase family, phosphorylates serine and threonine residues of substrate proteins in the presence of the Ku complex and double-stranded DNA. Although it has been established that DNA-PKcs is involved in non-homologous end-joining, a DNA double-strand break repair pathway, the mechanisms underlying DNA-PKcs activation are not fully understood. Nevertheless, the findings of numerous in vitro and in vivo studies have indicated that DNA-PKcs contains two autophosphorylation clusters, PQR and ABCDE, as well as several autophosphorylation sites and conformational changes associated with autophosphorylation of DNA-PKcs are important for self-activation. Consistent with these features, an analysis of transgenic mice has shown that the phenotypes of DNA-PKcs autophosphorylation mutations are significantly different from those of DNA-PKcs kinase-dead mutations, thereby indicating the importance of DNA-PKcs autophosphorylation in differentiation and development. Furthermore, there has been notable progress in the high-resolution analysis of the conformation of DNA-PKcs, which has enabled us to gain a visual insight into the steps leading to DNA-PKcs activation. This review summarizes the current progress in the activation of DNA-PKcs, focusing in particular on autophosphorylation of this kinase.
1. Introduction
Cells regulate the cell cycle and gene expression rapidly in response to both intracellular and extracellular stresses, during which signal transduction is primarily regulated via phosphorylation [1,2,3]. Within these signal transduction pathways, members of the PI3K-related kinase (PIKK) family, including ataxia-telangiectasia mutated (ATM), ataxia telangiectasia and Rad3-related (ATR), the DNA-dependent protein kinase catalytic subunit (DNA-PKcs), the mammalian target of rapamycin (mTOR), the suppressor with morphological effect on genitalia-1 (SMG-1) and the TRAF and TNF receptor-associated protein (TTRAP), are known to play central roles [4]. Although TTRAP lacks serine/threonine kinase activity, all of these proteins are characterized by the presence of HEAT, FAT and kinase domains [5,6,7]. Phylogenetic analyses of DNA-PKcs have recently indicated that many metazoans and plants have putative gene encoding DNA-PKcs. However, many model organisms such as Saccharomyces cerevisiae, Schizosaccharomyces pombe, Caenorhabditis elegans, Drosophila melanogaster and Arabidopsis thaliana lack an ortholog of DNA-PKcs. Therefore, DNA-PKcs appears to have been lost or its functions may have been diversified as organisms adapted to their environment during the course of evolution [8]. DNA-PKcs has been established as playing a central role in non-homologous end-joining (NHEJ), a DNA double-strand break (DSB) repair pathway, and its activation is dependent on its interactions with the Ku complex [9,10,11]. DNA-PKcs binds rapidly to DNA in the presence of Ku and it has been reported that the interaction of Akt1 and/or EGFR with DNA-PKcs promotes the activation of DNA-PK [12]. During NHEJ, the Ku70/Ku80 complex rapidly binds to DNA ends in response to DSBs; DNA-PKcs then binds to these DNA ends followed by the recruitment of NHEJ factors such as Artemis and DNA polymerase λ/μ, which subsequently process the DNA ends to generate ligatable ends [11,13]. In organisms that lack the gene encoding for DNA-PKcs, such as fission yeast (Schizosaccharomyces pombe), the Mre11/Rad50/Xrs2 complex interacts with NHEJ factors such as DNA ligase IV, is involved in the end-resection step of NHEJ and ensures NHEJ fidelity [14,15,16]. It has also been suggested that PAXX interacts with the Ku complex and stabilizes DNA ends [10,17,18] and that XRCC4/XLF filaments are formed within the vicinity of DNA ends to protect the DNA ends from further degradation [10,19,20,21,22]. However, the order and timing of DNA-PKcs binding and recruitment of these NHEJ factors have yet to be sufficiently established.
NHEJ is also involved in V(D)J recombination, which is responsible for the diversity of antigen recognition sites in antibodies and T-cell receptors [23]. In particular, DNA-PKcs is essential for opening the hairpin DNA formed after cleavage by the RAG1/RAG2 complex [24]. Artemis, which is the enzyme responsible for opening the hairpin DNA, has endonuclease activity [11,24]. However, when it forms a complex with DNA-PKcs and undergoes phosphorylation by DNA-PKcs, it acquires structure-specific exonuclease activity [11,24]. Therefore, an abnormality in the gene encoding DNA-PKcs results in radiosensitive severe combined immunodeficiency (RS-SCID) [23].
It is known that DNA-PKcs undergoes autophosphorylation and phosphorylates other NHEJ factors including Ku70, Ku80, Artemis, PNKP and XRCC4 and the roles of DNA-PKcs substrates and DNA-PKcs autophosphorylation in the DSB repair process have been well analyzed [25]. For example, it has been reported that autophosphorylation of DNA-PKcs is involved in end-processing, the pathway choice for DSB repair and normal mitotic progression [26,27,28,29]. Moreover, the phosphorylation status of DNA-PKcs is associated with a conformational change, which is required for self-activation [30,31,32,33,34,35]. The three-dimensional structure of the Ku complex was reported in detail in 2001 [36]. A visualization of the three-dimensional structure of DNA-PKcs was attempted by cryo-electron microscopy (Cryo-EM) in 1998 although the resolution was low [37,38,39]. More recently, there have been a number of higher resolution structural characterizations of DNA-PKcs using X-ray and Cryo-EM analyses [40,41,42,43]. Based on this information, in this review, the current understanding of autophosphorylation and the activation of DNA-PKcs has been summarized.
2. Self-Activation of a DNA-Dependent Protein Kinase Catalytic Subunit
DNA-PKcs primarily recognizes serine/glutamine and threonine/glutamine motifs in substrate proteins and phosphorylate serine and threonine residues in the presence of the Ku complex and double-stranded DNA (Figure 1) [44]. As shown in Figure 2, DNA-PKcs consists of the HEAT, FAT and kinase domains. The HEAT domain contains two major autophosphorylation clusters referred to as the S2056 (S2023–S2056) and T2609 (T2609–T2647) clusters, also known as the PQR and ABCDE clusters, respectively [7,26,33].
Based on the Cryo-EM analysis, the conformations of DNA-PKcs can be classified into three major groups based on its binding to the Ku complex and DNA and its corresponding activation state: (i) the DNA-PKcs-DNA complex, (ii) the DNA-PK holoenzyme in the inactivated state and (iii) the DNA-PK holoenzyme in the activated state [40]. In the DNA-PKcs-DNA complex, it was found that DNA-PKcs binds to DNA alone, although weakly; this is consistent with the weak kinase activity of DNA-PKcs in the absence of Ku but in the presence of DNA [45,46]. In addition to forming a DNA-PK holoenzyme, the self-activation of DNA-PK requires the physical interaction between Ku80 and DNA-PKcs as well as conformational changes in the FAT and kinase domains of DNA-PKcs. When the DNA-PK holoenzyme is in the activated state, a putative inositol 6-phosphate (IP6) binding site appears [40]. IP6 was reported by Hanakahi and West in 2000 as a factor that promotes end-joining in vitro [47]. Interestingly, IP6 binds to the Ku complex rather than to DNA-PKcs [24,48]. Consistent with this, it has been suggested that DNA-PKcs lacks an IP6 binding site unlike other PI3KKs [49]. To verify this contradiction, it is necessary to assess in detail whether IP6 binds to DNA-PKcs or the Ku complex.
With respect to autophosphorylation, there is yet to be a comprehensive characterization of PQR clusters (S2029, S2041, S2053 and S2056 in humans and S2026, S2038, S2050 and S2053 in mice), including the locations and structural changes associated with phosphorylation [40]. Of these phosphorylation sites, autophosphorylation of S2056 has been well analyzed. S2056 is phosphorylated in trans, at least in vitro, in response to ionizing radiation (IR) [50]. Phosphorylation of S2056 influences the pathway choice by restricting the end-processing by Artemis during the NHEJ reaction. It has been reported that the autophosphorylation efficiency of S2056 is related to the balance with O-linked β-N-acetylglucosamine (O-GlcNAc) modification, a post-translational modification of proteins, and the interaction of DNA-PKcs with casein kinase II and/or Akt1 [12,51,52]. However, even though all phosphorylation sites in the PQR cluster are replaced with alanine (hereafter referred to as DNA-PKcsPQR), phosphorylation of KAP-1 by DNA-PKcs occurs normally in response to IR. Furthermore, mouse embryonic fibroblasts (MEFs) and B-cells derived from DNA-PKcsPQR/PQR mice exhibited a mild sensitivity to IR [53]. Collectively, these findings indicate that phosphorylation at the PQR cluster is dispensable for the activation of DNA-PK.
In contrast to the PQR cluster, the ABCDE cluster plays an important role in the self-activation of DNA-PKcs [30,33,54]. Within the ABCDE cluster, six (in humans) or five (in mouse) serine and threonine residues (T2609, S2612, T2620, S2624, T2638 and T2647 in humans and T2605, S2614, S2616, T2634 and T2643 in mice) undergo phosphorylation in vitro. Four of these amino acids (T2609, S2612, T2638 and T2647) have also been found to undergo phosphorylation in vivo [32]. Mammalian cells and transgenic mice expressing DNA-PKcs with all phosphorylation sites or five (excluding the non-S/Q motif S2645 in all ABCDE clusters) or three threonine residues (T2609, T2638 and T2647 in humans; T2605, T2634 and T2463 in mice) of the ABCDE cluster replaced with alanine (hereinafter referred to as DNA-PKcs6A, DNA-PKcs5A and DNA-PKcs3A, respectively) are often used to analyze the role of the ABCDE cluster [30,31,32,34,35]. For example, analyses using V3 CHO cells expressing human DNA-PKcs6A and in vitro NHEJ assays using autophosphorylation mutant DNA-PKcs proteins revealed that DNA-PKcs remains bound to DNA ends in a Ku-dependent manner and that the NHEJ reaction does not proceed any further if the ABCDE cluster does not undergo phosphorylation [30,32]. Interestingly, two distinct types of conformational changes associated with phosphorylation have been reported in the ABCDE cluster. Briefly, in one scenario, the ABCDE cluster extends from within the HEAT domain to the DNA binding site and autophosphorylation at the ABCDE cluster occurs in trans. An alternative view suggests that the ABCDE cluster is located externally to the HEAT domain, adjacent to the substrate binding groove of the kinase domain, and that autophosphorylation at the ABCDE cluster occurs in cis (Figure 3) [32,40,55]. Consequently, it would be desirable to verify the precise location of the ABCDE cluster in future studies.
In addition to these clusters, it has been reported that autophosphorylation of the N-terminal cluster (S56 and S72) is involved in the inactivation of DNA-PKcs. In particular, phosphorylation of S72 has been suggested to destabilize the binding of DNA-PKs to DNA [28,40,56]. It has also been suggested that phosphorylation of the JK clusters (T946 and S1004) does not affect enzyme activity but is involved in the pathway choice [56]. Phosphorylation of the leucine zipper clusters (S1470 and S1546) and T1865 appears to have little effect on DNA-PK functions such as radiosensitivity, V(D)J recombination, the inhibition of homologous recombination and the assembly into Ku-bound DNA [56]. S3205 undergoes autophosphorylation modification by DNA-PKcs in vitro. However, it is phosphorylated by ATM in response to DNA damage in vivo [56]. Regarding T3950, as this residue is located within the activation loop of the kinase domain, its phosphorylation and dephosphorylation have been suggested to play a switching role with respect to kinase activity [40,42,57].
3. Effects of DNA-PKcs on Development and Differentiation Determined by Comparing Knockout and Mutant Mice
The importance of DNA-PKcs phosphorylation in the development of tissues and cells has been demonstrated using mouse models. For example, it has been found that while DNA-PKcs-/- mice develop normally, a DNA-PKcs kinase-dead mutant (DNA-PKcskd/kd), in which the aspartic acid at position 3922 of mouse DNA-PKcs is replaced by alanine, is embryonic lethal [58,59,60]. A histological analysis of the E14.5 brains of these DNA-PKcskd/kd mice revealed severe neuronal apoptosis, which was similar to that observed in Xrcc4 knockout mice. Neuronal apoptosis is primarily detected in the post-mitotic intermediate zone, thereby indicating that apoptosis occurs during the G0/G1 phase of the cell cycle in which NHEJ repair is predominant [59]. In addition, an accumulation of chromosomal breaks was detected in ES cells and MEFs derived from DNA-PKcskd/kd mice. Although ES cells derived from DNA-PKcs-/- were found to be only moderately sensitive to IR, those derived from DNA-PKcskd/kd mice were notably more sensitive. Although DNA-PK is also implicated in the pathway choice, a homologous recombination repair in ES cells derived from DNA-PKcskd/kd mice has been observed to be comparable with that in wild-type ES cells, thereby indicating that genomic instability caused by the kinase-dead mutation in DNA-PK is attributable to NHEJ defects. Although extrachromosomal end-ligation does not proceed in B-cells derived from DNA-PKcskd/kd mice, this end-ligation failure can be rescued by Ku deletion, consequently indicating that unphosphorylated DNA-PKcs blocks ligation [59]. Considering the fact that DNA-PKcsPQR/PQR mice are healthy and the aforementioned importance of the ABCDE cluster in DNA-PK activation, it appears that the fetal lethality observed in DNA-PKkd/kd mice is associated with an aberrant phosphorylation of the ABCDE cluster. Moreover, DNA-PKcs3A/3A mice, wherein three amino acids in the ABCDE cluster are replaced with alanine, have been found to have bone marrow and telomere abnormalities and die approximately 10 days after birth [35,61]. Accordingly, this phenotypic difference between DNA-PKcs3A/3A and DNA-PKcskd/kd mice would appear to indicate that in addition to phosphorylation of the ABCDE cluster, autophosphorylation of DNA-PKcs is an important process for the development (at least neurogenesis) of individual mice. Furthermore, given that the embryonic lethality characterizing DNA-PKkd/kd mice can be rescued by Ku deficiency, the effects of autophosphorylation such as those of S72 and T2950 (in human DNA-PKcs), which are required for the dissociation of DNA-PKcs from DNA, would not be negligible.
Gene knockout and kinase-dead mutations in DNA-PKcs also give rise to further phenotypic differences. Recently, it has been reported that DNA-PKcs is involved in rRNA processing and hematopoiesis in a Ku-dependent manner [34]. In mice, the loss of cNHEJ factors other than DNA-PKcs leads to the development of pro-B lymphoma in a TP53-deficient background [62,63,64], whereas, in the same background, most DNA-PKcs kinase-dead mutant mice die within 40 days without developing lymphoma [34]. An analysis of the bone marrow and spleen from DNA-PKcskd/kd TP53-/- mice revealed that these mice developed myelodysplastic syndrome. Studies focusing on autophosphorylation of DNA-PKcs have indicated that while DNA-PKcsPQR/PQR mice were healthy, DNA-PKcs5A/5A and DNA-PKcs3A/3A mice showed bone marrow failure [34,53]. The fact that Ku binds to both DNA and RNA [65,66], and that the Ku complex and DNA-PKcs localize in the nucleolus in the absence of DNA damage [67], suggests that DNA-PK may act in an RNA-dependent manner in the nucleolus. Indeed, an analysis using v-ABL kinase-transformed pro-B cell lines from DNA-PKcskd/kd and DNA-PKcs3A/3A mice revealed that translation is reduced in these cells [34]. Furthermore, the findings of a ChIRP-MS analysis indicated that the DNA-PK holoenzyme binds to U3 small nucleolar RNA (snoRNA), which is involved in the maturation of the 40S ribosomal subunit. Further in vitro phosphorylation assays have revealed that DNA-PKcs binds to the snoRNA stem-loop structure in a Ku-dependent manner, thereby causing autophosphorylation of the ABCDE cluster. Accordingly, these observations indicate that DNA-PKcskd/kd, DNA-PKcs5A/5A and DNA-PKcs3A/3A mice show Ku-dependent aberrant 18S rRNA processing, which results in an overall reduction in protein synthesis in hematopoietic cells [34]. However, although the role of the ABCDE cluster in rRNA processing remains to be conclusively established, Shao et al. speculated that the conformation of DNA-PKcs may differ depending on the identity of the binding nucleic acid (i.e., DNA or RNA) [34]. Indeed, an elucidation of the three-dimensional structure of DNA-PKcs bound to RNA is presumably the most important issue to be resolved regarding the role of DNA-PKcs in rRNA processing.
4. Conclusions
In this review, how the activation of DNA-PKcs is accompanied by autophosphorylation and conformational changes as well as the binding of IP6, the Ku complex and DNA have been described. The evidence also indicates that autophosphorylation of DNA-PKcs is important for protein synthesis in hematopoietic cells as well as in the development of neuronal and lymphoid cells. The findings reported herein highlight the continuing importance of studies based on animal models to clarify the roles of DNA-PKs. Although there have been notable advances in the structural analysis of macromolecular proteins by Cryo-EM, the structures of DNA-PKcs have yet to be sufficiently clarified. Consequently, the precise locations of the ABCDE and PQR clusters and the conformation of DNA-PKs in the ATP-bound state remain to be determined. In conclusion, it is envisaged that a complementary combination of cellular, molecular, biochemical and structural analyses will contribute to gaining a more comprehensive understanding of the mechanisms underlying DNA-PK activation, thereby facilitating a further elucidation of the multiple functions of DNA-PKcs.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data sharing is not applicable to this article.
Acknowledgments
I am deeply grateful to the Guest Editor of Genes, Junya Kobayashi, for giving me the opportunity to write this review article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Huang, R.X.; Zhou, P.K. DNA damage response signaling pathways and targets for radiotherapy sensitization in cancer. Signal Transduct. Target. Ther. 2020, 5, 60. [Google Scholar] [CrossRef]
- Jeggo, P.; Löbrich, M. Radiation-induced DNA damage responses. Radiat. Prot. Dosim. 2006, 122, 124–127. [Google Scholar] [CrossRef]
- Lanz, M.C.; Dibitetto, D.; Smolka, M.B. DNA damage kinase signaling: Checkpoint and repair at 30 years. EMBO J. 2019, 38, e101801. [Google Scholar] [CrossRef]
- Kidiyoor, G.R.; Kumar, A.; Foiani, M. ATR-mediated regulation of nuclear and cellular plasticity. DNA Repair 2016, 44, 143–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blackford, A.N.; Jackson, S.P. ATM, ATR, and DNA-PK: The Trinity at the heart of the DNA damage response. Mol. Cell 2017, 66, 801–817. [Google Scholar] [CrossRef] [Green Version]
- Elias-Villalobos, A.; Fort, P.; Helmlinger, D. New insights into the evolutionary conservation of the sole PIKK pseudokinase Tra1/TRRAP. Biochem. Soc. Trans. 2019, 47, 1597–1608. [Google Scholar] [CrossRef] [PubMed]
- Imseng, S.; Aylett, C.H.; Maier, T. Architecture and activation of phosphatidylinositol 3-kinase related kinases. Curr. Opin. Struct. Biol. 2018, 49, 177–189. [Google Scholar] [CrossRef]
- Lees-Miller, J.P.; Cobban, A.; Katsonis, P.; Bacolla, A.; Tsutakawa, S.E.; Hammel, M.; Meek, K.; Anderson, D.W.; Lichtarge, O.; Tainer, J.A.; et al. Uncovering DNA-PKcs ancient phylogeny, unique sequence motifs and insights for human disease. Prog. Biophys. Mol. Biol. 2021, 163, 87–108. [Google Scholar] [CrossRef] [PubMed]
- Ensminger, M.; Löbrich, M. One end to rule them all: Non-homologous end-joining and homologous recombination at DNA double-strand breaks. Br. J. Radiol. 2020, 93, 20191054. [Google Scholar] [CrossRef]
- Ghosh, D.; Raghavan, S.C. Nonhomologous end joining: New accessory factors fine tune the machinery. Trends Genet. 2021, 37, 582–599. [Google Scholar] [CrossRef]
- Kurosawa, A.; Adachi, N. Functions and regulation of Artemis: A goddess in the maintenance of genome integrity. J. Radiat. Res. 2010, 51, 503–509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodwin, J.F.; Knudsen, K.E. Beyond DNA repair: DNA-PK function in cancer. Cancer Discov. 2014, 4, 1126–1139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, H.H.Y.; Watanabe, G.; Gerodimos, C.A.; Ochi, T.; Blundell, T.L.; Jackson, S.P.; Lieber, M.R. Different DNA end configurations dictate which NHEJ components are most important for joining efficiency. J. Biol. Chem. 2016, 291, 24377–24389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwasaki, D.; Hayashihara, K.; Shima, H.; Higashide, M.; Terasawa, M.; Gasser, S.M.; Shinohara, M. The MRX complex ensures NHEJ fidelity through multiple pathways including Xrs2-FHA-dependent Tel1 activation. PLoS Genet. 2016, 12, e1005942. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Wang, J.; Zhou, G.; Lajeunesse, M.; Le, N.; Stawicki, B.N.; Corcino, Y.L.; Berkner, K.L.; Runge, K.W. Nonhomologous end-joining with minimal sequence loss is promoted by the Mre11-Rad50-Nbs1-Ctp1 complex in Schizosaccharomyces pombe. Genetics 2017, 206, 481–496. [Google Scholar] [CrossRef] [Green Version]
- Matsuzaki, K.; Shinohara, A.; Shinohara, M. Forkhead-associated domain of yeast Xrs2, a homolog of human Nbs1, promotes nonhomologous end joining through interaction with a ligase IV partner protein, Lif1. Genetics 2008, 179, 213–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ochi, T.; Blackford, A.N.; Coates, J.; Jhujh, S.; Mehmood, S.; Tamura, N.; Travers, J.; Wu, Q.; Draviam, V.M.; Robinson, C.V.; et al. DNA repair. PAXX, a paralog of XRCC4 and XLF, interacts with Ku to promote DNA double-strand break repair. Science 2015, 347, 185–188. [Google Scholar] [CrossRef] [Green Version]
- Xing, M.; Yang, M.; Huo, W.; Feng, F.; Wei, L.; Jiang, W.; Ning, S.; Yan, Z.; Li, W.; Wang, Q.; et al. Interactome analysis identifies a new paralogue of XRCC4 in non-homologous end joining DNA repair pathway. Nat. Commun. 2015, 6, 6233. [Google Scholar] [CrossRef] [Green Version]
- Andres, S.N.; Vergnes, A.; Ristic, D.; Wyman, C.; Modesti, M.; Junop, M. A human XRCC4-XLF complex bridges DNA. Nucleic Acids Res. 2012, 40, 1868–1878. [Google Scholar] [CrossRef] [Green Version]
- Hammel, M.; Yu, Y.; Fang, S.; Lees-Miller, S.P.; Tainer, J.A. XLF regulates filament architecture of the XRCC4.ligase IV complex. Structure 2010, 18, 1431–1442. [Google Scholar] [CrossRef] [Green Version]
- Mahaney, B.L.; Hammel, M.; Meek, K.; Tainer, J.A.; Lees-Miller, S.P. XRCC4 and XLF form long helical protein filaments suitable for DNA end protection and alignment to facilitate DNA double strand break repair. Biochem. Cell Biol. 2013, 91, 31–41. [Google Scholar] [CrossRef] [Green Version]
- Ropars, V.; Drevet, P.; Legrand, P.; Baconnais, S.; Amram, J.; Faure, G.; Marquez, J.A.; Pietrement, O.; Guerois, R.; Callebaut, I.; et al. Structural characterization of filaments formed by human Xrcc4-Cernunnos/XLF complex involved in nonhomologous DNA end-joining. Proc. Natl. Acad. Sci. USA 2011, 108, 12663–12668. [Google Scholar] [CrossRef] [Green Version]
- De Villartay, J.P. Congenital defects in V(D)J recombination. Br. Med. Bull. 2015, 114, 157–167. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Lieber, M.R. Binding of inositol hexakisphosphate (IP6) to Ku but not to DNA-PKcs. J. Biol. Chem. 2002, 277, 10756–10759. [Google Scholar] [CrossRef] [Green Version]
- Anisenko, A.N.; Gottikh, M.B. Role of cellular DNA repair systems in HIV-1 replication. Mol. Biol. 2019, 53, 355–366. [Google Scholar] [CrossRef]
- Cui, X.; Yu, Y.; Gupta, S.; Cho, Y.M.; Lees-Miller, S.P.; Meek, K. Autophosphorylation of DNA-dependent protein kinase regulates DNA end processing and may also alter double-strand break repair pathway choice. Mol. Cell. Biol. 2005, 25, 10842–10852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uematsu, N.; Weterings, E.; Yano, K.; Morotomi-Yano, K.; Jakob, B.; Taucher-Scholz, G.; Mari, P.O.; van Gent, D.C.; Chen, B.P.; Chen, D.J. Autophosphorylation of DNA-PKCS regulates its dynamics at DNA double-strand breaks. J. Cell Biol. 2007, 177, 219–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jette, N.; Lees-Miller, S.P. The DNA-dependent protein kinase: A multifunctional protein kinase with roles in DNA double strand break repair and mitosis. Prog. Biophys. Mol. Biol. 2015, 117, 194–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.J.; Lin, Y.F.; Chou, H.Y.; Yajima, H.; Fattah, K.R.; Lee, S.C.; Chen, B.P. Involvement of DNA-dependent protein kinase in normal cell cycle progression through mitosis. J. Biol. Chem. 2011, 286, 12796–12802. [Google Scholar] [CrossRef] [Green Version]
- Block, W.D.; Yu, Y.; Merkle, D.; Gifford, J.L.; Ding, Q.; Meek, K.; Lees-Miller, S.P. Autophosphorylation-dependent remodeling of the DNA-dependent protein kinase catalytic subunit regulates ligation of DNA ends. Nucleic Acids Res. 2004, 32, 4351–4357. [Google Scholar] [CrossRef] [Green Version]
- Crowe, J.L.; Wang, X.S.; Shao, Z.; Lee, B.J.; Estes, V.M.; Zha, S. DNA-PKcs phosphorylation at the T2609 cluster alters the repair pathway choice during immunoglobulin class switch recombination. Proc. Natl. Acad. Sci. USA 2020, 117, 22953–22961. [Google Scholar] [CrossRef]
- Ding, Q.; Reddy, Y.V.; Wang, W.; Woods, T.; Douglas, P.; Ramsden, D.A.; Lees-Miller, S.P.; Meek, K. Autophosphorylation of the catalytic subunit of the DNA-dependent protein kinase is required for efficient end processing during DNA double-strand break repair. Mol. Cell. Biol. 2003, 23, 5836–5848. [Google Scholar] [CrossRef] [Green Version]
- Meek, K. Activation of DNA-PK by hairpinned DNA ends reveals a stepwise mechanism of kinase activation. Nucleic Acids Res. 2020, 48, 9098–9108. [Google Scholar] [CrossRef]
- Shao, Z.; Flynn, R.A.; Crowe, J.L.; Zhu, Y.; Liang, J.; Jiang, W.; Aryan, F.; Aoude, P.; Bertozzi, C.R.; Estes, V.M.; et al. DNA-PKcs has KU-dependent function in rRNA processing and haematopoiesis. Nature 2020, 579, 291–296. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Yajima, H.; Huynh, H.; Zheng, J.; Callen, E.; Chen, H.T.; Wong, N.; Bunting, S.; Lin, Y.F.; Li, M.; et al. Congenital bone marrow failure in DNA-PKcs mutant mice associated with deficiencies in DNA repair. J. Cell. Biol. 2011, 193, 295–305. [Google Scholar] [CrossRef] [PubMed]
- Walker, J.R.; Corpina, R.A.; Goldberg, J. Structure of the Ku heterodimer bound to DNA and its implications for double-strand break repair. Nature 2001, 412, 607–614. [Google Scholar] [CrossRef] [PubMed]
- Chiu, C.Y.; Cary, R.B.; Chen, D.J.; Peterson, S.R.; Stewart, P.L. Cryo-EM imaging of the catalytic subunit of the DNA-dependent protein kinase. J. Mol. Biol. 1998, 284, 1075–1081. [Google Scholar] [CrossRef]
- Leuther, K.K.; Hammarsten, O.; Kornberg, R.D.; Chu, G. Structure of DNA-dependent protein kinase: Implications for its regulation by DNA. EMBO J. 1999, 18, 1114–1123. [Google Scholar] [CrossRef] [Green Version]
- Rivera-Calzada, A.; Maman, J.D.; Spagnolo, L.; Pearl, L.H.; Llorca, O. Three-dimensional structure and regulation of the DNA-dependent protein kinase catalytic subunit (DNA-PKcs). Structure 2005, 13, 243–255. [Google Scholar] [CrossRef]
- Chen, X.; Xu, X.; Chen, Y.; Cheung, J.C.; Wang, H.; Jiang, J.; de Val, N.; Fox, T.; Gellert, M.; Yang, W. Structure of an activated DNA-PK and its implications for NHEJ. Mol. Cell 2021, 81, 801–810 e803. [Google Scholar] [CrossRef]
- Sharif, H.; Li, Y.; Dong, Y.; Dong, L.; Wang, W.L.; Mao, Y.; Wu, H. Cryo-EM structure of the DNA-PK holoenzyme. Proc. Natl. Acad. Sci. USA 2017, 114, 7367–7372. [Google Scholar] [CrossRef] [Green Version]
- Sibanda, B.L.; Chirgadze, D.Y.; Ascher, D.B.; Blundell, T.L. DNA-PKcs structure suggests an allosteric mechanism modulating DNA double-strand break repair. Science 2017, 355, 520–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, X.; Liu, M.; Tian, Y.; Wang, J.; Xu, Y. Cryo-EM structure of human DNA-PK holoenzyme. Cell Res. 2017, 27, 1341–1350. [Google Scholar] [CrossRef] [Green Version]
- Anderson, C.W.; Lees-Miller, S.P. The nuclear serine/threonine protein kinase DNA-PK. Crit. Rev. Eukaryot. Gene. Expr. 1992, 2, 283–314. [Google Scholar] [PubMed]
- Cary, R.B.; Peterson, S.R.; Wang, J.; Bear, D.G.; Bradbury, E.M.; Chen, D.J. DNA looping by Ku and the DNA-dependent protein kinase. Proc. Natl. Acad. Sci. USA 1997, 94, 4267–4272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- West, R.B.; Yaneva, M.; Lieber, M.R. Productive and nonproductive complexes of Ku and DNA-dependent protein kinase at DNA termini. Mol. Cell. Biol. 1998, 18, 5908–5920. [Google Scholar] [CrossRef] [Green Version]
- Hanakahi, L.A.; Bartlet-Jones, M.; Chappell, C.; Pappin, D.; West, S.C. Binding of inositol phosphate to DNA-PK and stimulation of double-strand break repair. Cell 2000, 102, 721–729. [Google Scholar] [CrossRef] [Green Version]
- Hanakahi, L.A.; West, S.C. Specific interaction of IP6 with human Ku70/80, the DNA-binding subunit of DNA-PK. EMBO J. 2002, 21, 2038–2044. [Google Scholar] [CrossRef] [Green Version]
- Gat, Y.; Schuller, J.M.; Lingaraju, M.; Weyher, E.; Bonneau, F.; Strauss, M.; Murray, P.J.; Conti, E. InsP6 binding to PIKK kinases revealed by the cryo-EM structure of an SMG1-SMG8-SMG9 complex. Nat. Struct. Mol. Biol. 2019, 26, 1089–1093. [Google Scholar] [CrossRef]
- Chen, B.P.; Chan, D.W.; Kobayashi, J.; Burma, S.; Asaithamby, A.; Morotomi-Yano, K.; Botvinick, E.; Qin, J.; Chen, D.J. Cell cycle dependence of DNA-dependent protein kinase phosphorylation in response to DNA double strand breaks. J. Biol. Chem. 2005, 280, 14709–14715. [Google Scholar] [CrossRef] [Green Version]
- Lafont, F.; Fleury, F.; Benhelli-Mokrani, H. DNA-PKcs Ser2056 auto-phosphorylation is affected by an O-GlcNAcylation/phosphorylation interplay. Biochim. Biophys. Acta Gen. Subj. 2020, 1864, 129705. [Google Scholar] [CrossRef]
- Toulany, M.; Lee, K.J.; Fattah, K.R.; Lin, Y.F.; Fehrenbacher, B.; Schaller, M.; Chen, B.P.; Chen, D.J.; Rodemann, H.P. Akt promotes post-irradiation survival of human tumor cells through initiation, progression, and termination of DNA-PKcs-dependent DNA double-strand break repair. Mol. Cancer Res. 2012, 10, 945–957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, W.; Estes, V.M.; Wang, X.S.; Shao, Z.; Lee, B.J.; Lin, X.; Crowe, J.L.; Zha, S. Phosphorylation at S2053 in murine (S2056 in Human) DNA-PKcs is dispensable for lymphocyte development and class switch recombination. J. Immunol. 2019, 203, 178–187. [Google Scholar] [CrossRef]
- Reddy, Y.V.; Ding, Q.; Lees-Miller, S.P.; Meek, K.; Ramsden, D.A. Non-homologous end joining requires that the DNA-PK complex undergo an autophosphorylation-dependent rearrangement at DNA ends. J. Biol. Chem. 2004, 279, 39408–39413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neal, J.A.; Xu, Y.; Abe, M.; Hendrickson, E.; Meek, K. Restoration of ATM expression in DNA-PKcs-deficient cells inhibits signal end joining. J. Immunol. 2016, 196, 3032–3042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neal, J.A.; Dang, V.; Douglas, P.; Wold, M.S.; Lees-Miller, S.P.; Meek, K. Inhibition of homologous recombination by DNA-dependent protein kinase requires kinase activity, is titratable, and is modulated by autophosphorylation. Mol. Cell. Biol. 2011, 31, 1719–1733. [Google Scholar] [CrossRef] [Green Version]
- Neal, J.A.; Sugiman-Marangos, S.; VanderVere-Carozza, P.; Wagner, M.; Turchi, J.; Lees-Miller, S.P.; Junop, M.S.; Meek, K. Unraveling the complexities of DNA-dependent protein kinase autophosphorylation. Mol. Cell. Biol. 2014, 34, 2162–2175. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Chaudhuri, J.; Zhu, C.; Davidson, L.; Weaver, D.T.; Alt, F.W. A targeted DNA-PKcs-null mutation reveals DNA-PK-independent functions for KU in V(D)J recombination. Immunity 1998, 9, 367–376. [Google Scholar] [CrossRef] [Green Version]
- Jiang, W.; Crowe, J.L.; Liu, X.; Nakajima, S.; Wang, Y.; Li, C.; Lee, B.J.; Dubois, R.L.; Liu, C.; Yu, X.; et al. Differential phosphorylation of DNA-PKcs regulates the interplay between end-processing and end-ligation during nonhomologous end-joining. Mol. Cell 2015, 58, 172–185. [Google Scholar] [CrossRef] [Green Version]
- Taccioli, G.E.; Amatucci, A.G.; Beamish, H.J.; Gell, D.; Xiang, X.H.; Torres Arzayus, M.I.; Priestley, A.; Jackson, S.P.; Marshak Rothstein, A.; Jeggo, P.A.; et al. Targeted disruption of the catalytic subunit of the DNA-PK gene in mice confers severe combined immunodeficiency and radiosensitivity. Immunity 1998, 9, 355–366. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Matsunaga, S.; Lin, Y.F.; Sishc, B.; Shang, Z.; Sui, J.; Shih, H.Y.; Zhao, Y.; Foreman, O.; Story, M.D.; et al. Spontaneous tumor development in bone marrow-rescued DNA-PKcs(3A/3A) mice due to dysfunction of telomere leading strand deprotection. Oncogene 2016, 35, 3909–3918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frank, K.M.; Sharpless, N.E.; Gao, Y.; Sekiguchi, J.M.; Ferguson, D.O.; Zhu, C.; Manis, J.P.; Horner, J.; DePinho, R.A.; Alt, F.W. DNA ligase IV deficiency in mice leads to defective neurogenesis and embryonic lethality via the p53 pathway. Mol. Cell 2000, 5, 993–1002. [Google Scholar] [CrossRef]
- Nacht, M.; Strasser, A.; Chan, Y.R.; Harris, A.W.; Schlissel, M.; Bronson, R.T.; Jacks, T. Mutations in the p53 and SCID genes cooperate in tumorigenesis. Genes Dev. 1996, 10, 2055–2066. [Google Scholar] [CrossRef] [Green Version]
- Zhu, C.; Mills, K.D.; Ferguson, D.O.; Lee, C.; Manis, J.; Fleming, J.; Gao, Y.; Morton, C.C.; Alt, F.W. Unrepaired DNA breaks in p53-deficient cells lead to oncogenic gene amplification subsequent to translocations. Cell 2002, 109, 811–821. [Google Scholar] [CrossRef] [Green Version]
- Shadrina, O.; Garanina, I.; Korolev, S.; Zatsepin, T.; Van Assche, J.; Daouad, F.; Wallet, C.; Rohr, O.; Gottikh, M. Analysis of RNA binding properties of human Ku protein reveals its interactions with 7SK snRNA and protein components of 7SK snRNP complex. Biochimie 2020, 171–172, 110–123. [Google Scholar] [CrossRef]
- Yoo, S.; Dynan, W.S. Characterization of the RNA binding properties of Ku protein. Biochemistry 1998, 37, 1336–1343. [Google Scholar] [CrossRef]
- Britton, S.; Coates, J.; Jackson, S.P. A new method for high-resolution imaging of Ku foci to decipher mechanisms of DNA double-strand break repair. J. Cell Biol. 2013, 202, 579–595. [Google Scholar] [CrossRef] [Green Version]
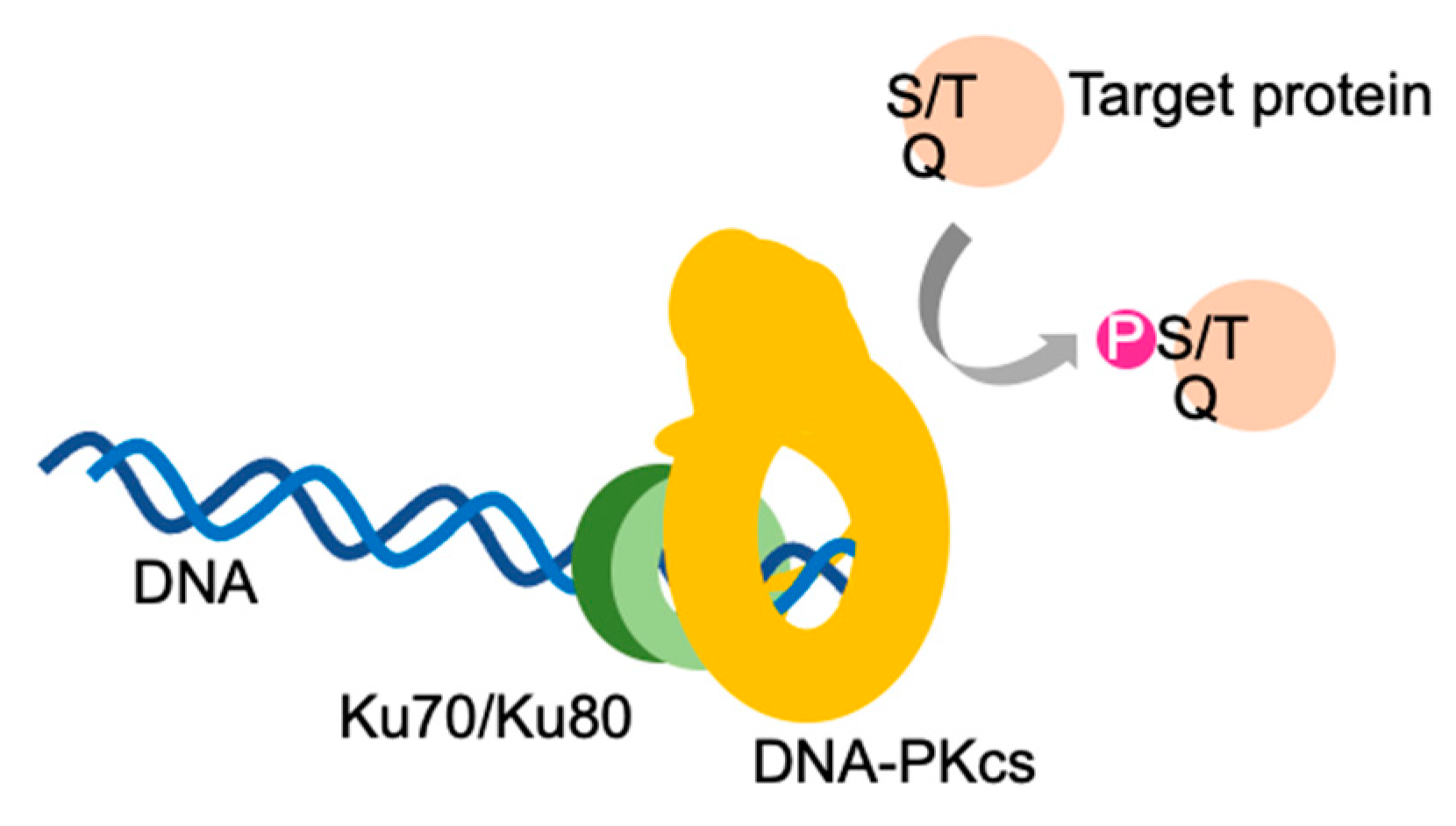
Figure 1.
DNA-dependent protein kinase. DNA-PK holoenzyme consists of a DNA-dependent protein kinase catalytic subunit (DNA-PKcs), the Ku complex (Ku70/Ku80) and double-stranded DNA. DNA-PKcs acquires kinase activity in the presence of the Ku complex and double-stranded DNA. DNA-PK holoenzymes preferentially phosphorylate the serine (S) and threonine (T) of serine /glutamine (Q) and threonine/glutamine motifs in target proteins.
Figure 1.
DNA-dependent protein kinase. DNA-PK holoenzyme consists of a DNA-dependent protein kinase catalytic subunit (DNA-PKcs), the Ku complex (Ku70/Ku80) and double-stranded DNA. DNA-PKcs acquires kinase activity in the presence of the Ku complex and double-stranded DNA. DNA-PK holoenzymes preferentially phosphorylate the serine (S) and threonine (T) of serine /glutamine (Q) and threonine/glutamine motifs in target proteins.

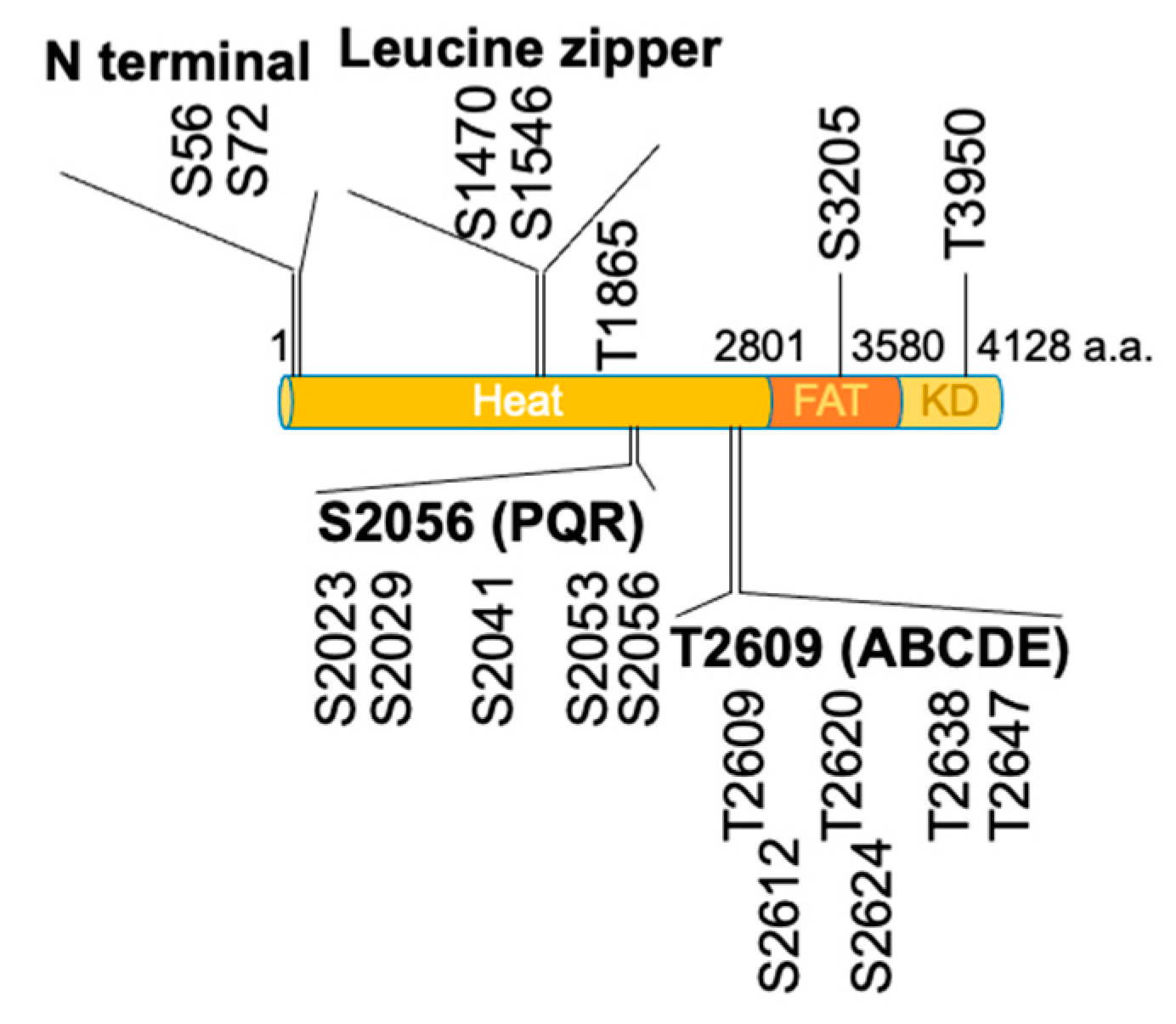
Figure 2.
DNA-PKcs consists of the HEAT domain, FAT domain and kinase domain (KD). There are four autophosphorylation clusters (N terminal, leucine zipper, S2056/PQR and T2609/ABCDE) and one autophosphorylation site in the HEAT domain. There is an autophosphorylation site in the FAT (S3205) and KD (T3950) domains, respectively.
Figure 2.
DNA-PKcs consists of the HEAT domain, FAT domain and kinase domain (KD). There are four autophosphorylation clusters (N terminal, leucine zipper, S2056/PQR and T2609/ABCDE) and one autophosphorylation site in the HEAT domain. There is an autophosphorylation site in the FAT (S3205) and KD (T3950) domains, respectively.

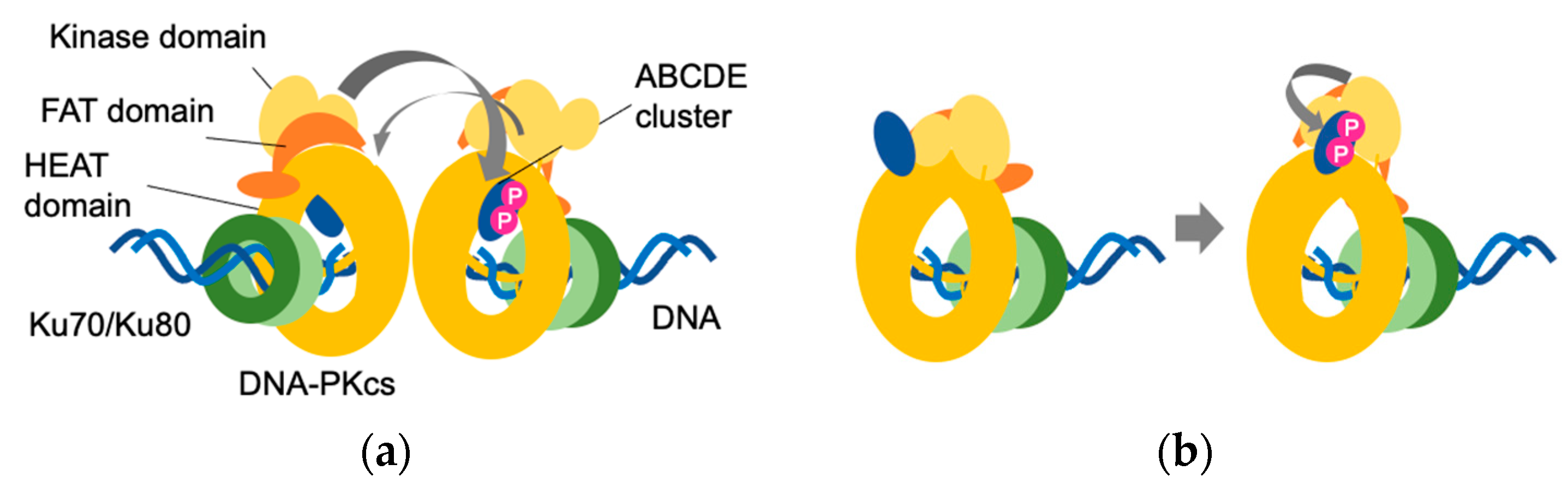
Figure 3.
Schematic representation of a putative ABCDE cluster. (a) A putative ABCDE cluster is located inside the HEAT domain. Autophosphorylation occurs in trans; (b) Another putative ABCDE cluster is located outside the HEAT domain. Autophosphorylation occurs in cis. A blue oval indicates the ABCDE domain; a P in white on a pink circle indicates the phosphate group.
Figure 3.
Schematic representation of a putative ABCDE cluster. (a) A putative ABCDE cluster is located inside the HEAT domain. Autophosphorylation occurs in trans; (b) Another putative ABCDE cluster is located outside the HEAT domain. Autophosphorylation occurs in cis. A blue oval indicates the ABCDE domain; a P in white on a pink circle indicates the phosphate group.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kurosawa, A. Autophosphorylation and Self-Activation of DNA-Dependent Protein Kinase. Genes 2021, 12, 1091. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12071091
AMA Style
Kurosawa A. Autophosphorylation and Self-Activation of DNA-Dependent Protein Kinase. Genes. 2021; 12(7):1091. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12071091
Chicago/Turabian StyleKurosawa, Aya. 2021. "Autophosphorylation and Self-Activation of DNA-Dependent Protein Kinase" Genes 12, no. 7: 1091. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12071091
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.