
Effects of Cocoa Genotypes on Coat Color, Platelets and Coagulation Parameters in French Bulldogs


 , and
, and {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Test Material
2.2. Clinical Parameters
2.3. Mepacrin Assay
2.4. Genotyping
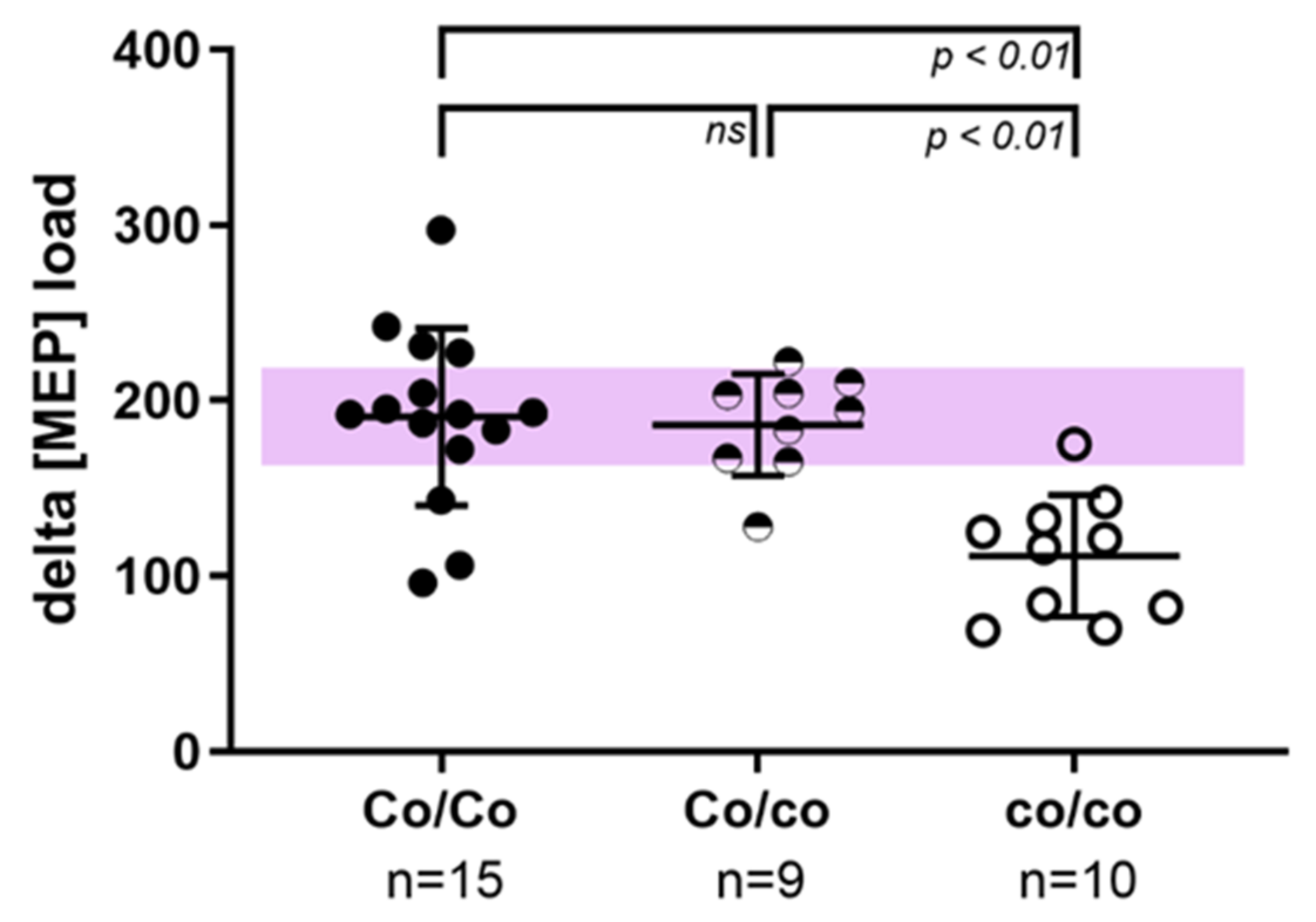
3. Results
3.1. Coat Colors
3.2. Hematological Investigations
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schmutz, S.M.; Berryere, T.G.; Goldfinch, A.D. TYRP1 and MC1R genotypes and their effects on coat color in dogs. Mamm. Genome 2002, 13, 380–387. [Google Scholar] [CrossRef]
- Turnova, E.H.; Majchrakova, Z.; Bielikova, M.; Soltys, K.; Turňa, J.; Dudas, A. A novel mutation in the TYRP1 gene associated with brown coat colour in the Australian Shepherd Dog Breed. Anim. Genet. 2017, 48, 626. [Google Scholar] [CrossRef] [PubMed]
- Van Buren, S.L.; Mickelson, J.R.; Minor, K.M. A novel TYRP1 mutation associated with brown coat color in Siberian huskies. Anim. Genet. 2021, 52, 245–246. [Google Scholar] [CrossRef] [PubMed]
- Wright, H.E.; Schofield, E.; Mellersh, C.S.; Burmeister, L.M. A novel TYRP1 variant is associated with liver and tan coat colour in Lancashire Heelers. Anim. Genet. 2019, 50, 783. [Google Scholar] [CrossRef] [PubMed]
- Kiener, S.; Kehl, A.; Loechel, R.; Langbein-Detsch, I.; Müller, E.; Bannasch, D.; Jagannathan, V.; Leeb, T. Novel Brown Coat Color (Cocoa) in French Bulldogs Results from a Nonsense Variant in HPS. Genes 2020, 11, 636. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Guiu, I.; Torregrosa, J.M.; Velasco, F.; Antón, A.I.; Lozano, M.L.; Vicente, V.; Rivera, J. Hermansky-Pudlak syndrome. Overview of clinical and molecular features and case report of a new HPS-1 variant. Hamostaseologie 2014, 34, 301–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Chemaly, S.; Young, L.R. Hermansky-Pudlak Syndrome. Clin. Chest Med. 2016, 37, 505–511. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, T.; Li, W.; Zhang, Q.; Novak, E.K.; Sviderskaya, E.V.; Wilson, A.; Bennett, D.C.; Roe, B.A.; Swank, R.T.; Spritz, R.A. The gene mutated in cocoa mice, carrying a defect of organelle biogenesis, is a homologue of the human Hermansky-Pudlak syndrome-3 gene. Genomics 2001, 78, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Novak, E.K.; Sweet, H.O.; Prochazka, M.; Parentis, M.; Soble, R.; Reddington, M.; Cairo, A.; Swank, R.T. Cocoa: A new mouse model for platelet storage pool deficiency. Br. J. Haematol. 1988, 69, 371–378. [Google Scholar] [CrossRef] [PubMed]
- Manukjan, G.; Eilenberger, J.; Andres, O.; Schambeck, C.; Eber, S.; Schulze, H. Functional Classification of Paediatric Patients with Non-syndromic Delta-Storage Pool Deficiency. Hamostaseologie 2018, 39, 383–391. [Google Scholar] [CrossRef]
- Gordon, N.; Thom, J.; Cole, C.; Baker, R. Rapid detection of hereditary and acquired platelet storage pool deficiency by flow cytometry. Br. J. Haematol. 1995, 89, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Huizing, M.; Malicdan, M.C.V.; Wang, J.A.; Pri-Chen, H.; Hess, R.A.; Fischer, R.; O’Brien, K.J.; Merideth, M.A.; Gahl, W.A.; Gochuico, B.R. Hermansky–Pudlak syndrome: Mutation update. Hum. Mutat. 2019, 41, 543–580. [Google Scholar] [CrossRef]
- Berber, I.; Erkurt, M.A.; Kuku, I.; Kaya, E.; Koroglu, M.; Nizam, I.; Gul, M.; Bentli, R. Hermansky-Pudlak Syndrome: A Case Report. Case Rep. Hematol. 2014, 2014, 249195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ambrosio, A.L.; Di Pietro, S.M. Storage pool diseases illuminate platelet dense granule biogenesis. Platelets 2017, 28, 138–146. [Google Scholar] [CrossRef] [PubMed]
- Hoareau, G.; Mellema, M. Pro-coagulant thromboelastographic features in the bulldog. J. Small Anim. Pract. 2014, 56, 103–107. [Google Scholar] [CrossRef] [PubMed]
- Crane, C.; Rozanski, E.A.; Abelson, A.L.; DeLaforcade, A. Severe brachycephalic obstructive airway syndrome is associated with hypercoagulability in dogs. J. Veter Diagn. Investig. 2017, 29, 570–573. [Google Scholar] [CrossRef] [PubMed]
- Selle, F.; James, C.; Tuffigo, M.; Pillois, X.; Viallard, J.F.; Alessi, M.C.; Fiore, M. Clinical and Laboratory Findings in Patients with δ-Storage Pool Disease: A Case Series. Semin. Thromb Hemost 2017, 43, 48–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Billio, A.; Moeseneder, C.; Donazzan, G.; Triani, A.; Pescosta, N.; Coser, P. Hermansky-Pudlak syndrome: Clinical presentation and confirmation of the value of the mepacrine-based cytofluorimetry test in the diagnosis of delta granule deficiency. Haematologica 2001, 86, 220. [Google Scholar] [PubMed]
- Andres, O.; Wiegering, V.; König, E.M.; Schneider, A.L.; Semeniak, D.; Stritt, S.; Klopocki, E.; Schulze, H. A novel two-nucleotide deletion in HPS6 affects mepacrine uptake and platelet dense granule secretion in a family with Hermansky-Pudlak syndrome. Pediatr. Blood Cancer 2017, 64, e26320. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Laukner, A.; Truchet, L.; Manukjan, G.; Schulze, H.; Langbein-Detsch, I.; Mueller, E.; Leeb, T.; Kehl, A. Effects of Cocoa Genotypes on Coat Color, Platelets and Coagulation Parameters in French Bulldogs. Genes 2021, 12, 1092. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12071092
Laukner A, Truchet L, Manukjan G, Schulze H, Langbein-Detsch I, Mueller E, Leeb T, Kehl A. Effects of Cocoa Genotypes on Coat Color, Platelets and Coagulation Parameters in French Bulldogs. Genes. 2021; 12(7):1092. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12071092
Chicago/Turabian StyleLaukner, Anna, Laura Truchet, Georgi Manukjan, Harald Schulze, Ines Langbein-Detsch, Elisabeth Mueller, Tosso Leeb, and Alexandra Kehl. 2021. "Effects of Cocoa Genotypes on Coat Color, Platelets and Coagulation Parameters in French Bulldogs" Genes 12, no. 7: 1092. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12071092