
Genomic Association vs. Serological Determination of ABO Blood Types in a Chinese Cohort, with Application in Mendelian Randomization


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design and Ethics
2.2. Data Analysis
2.3. Analytical Resources
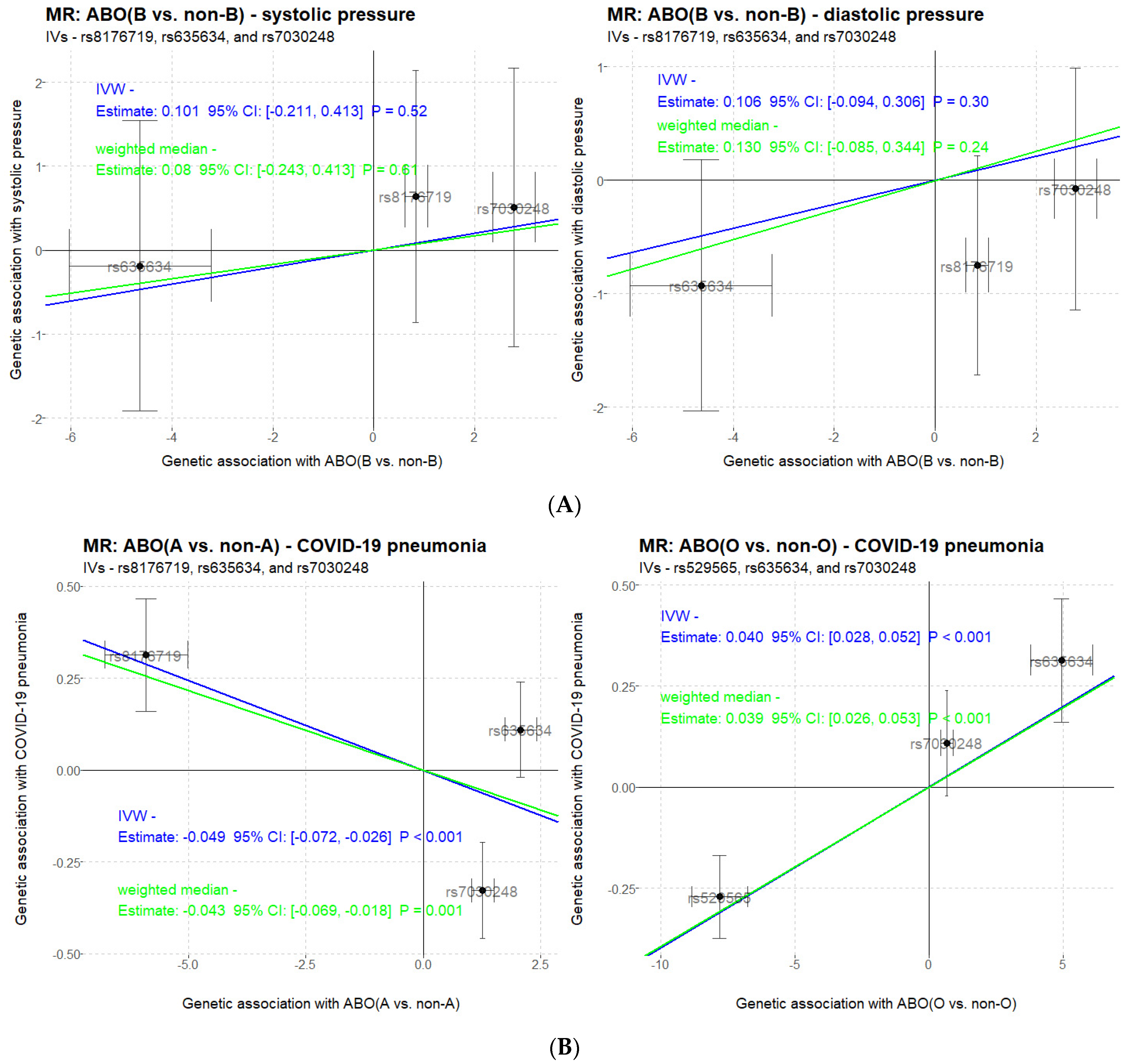
3. Results
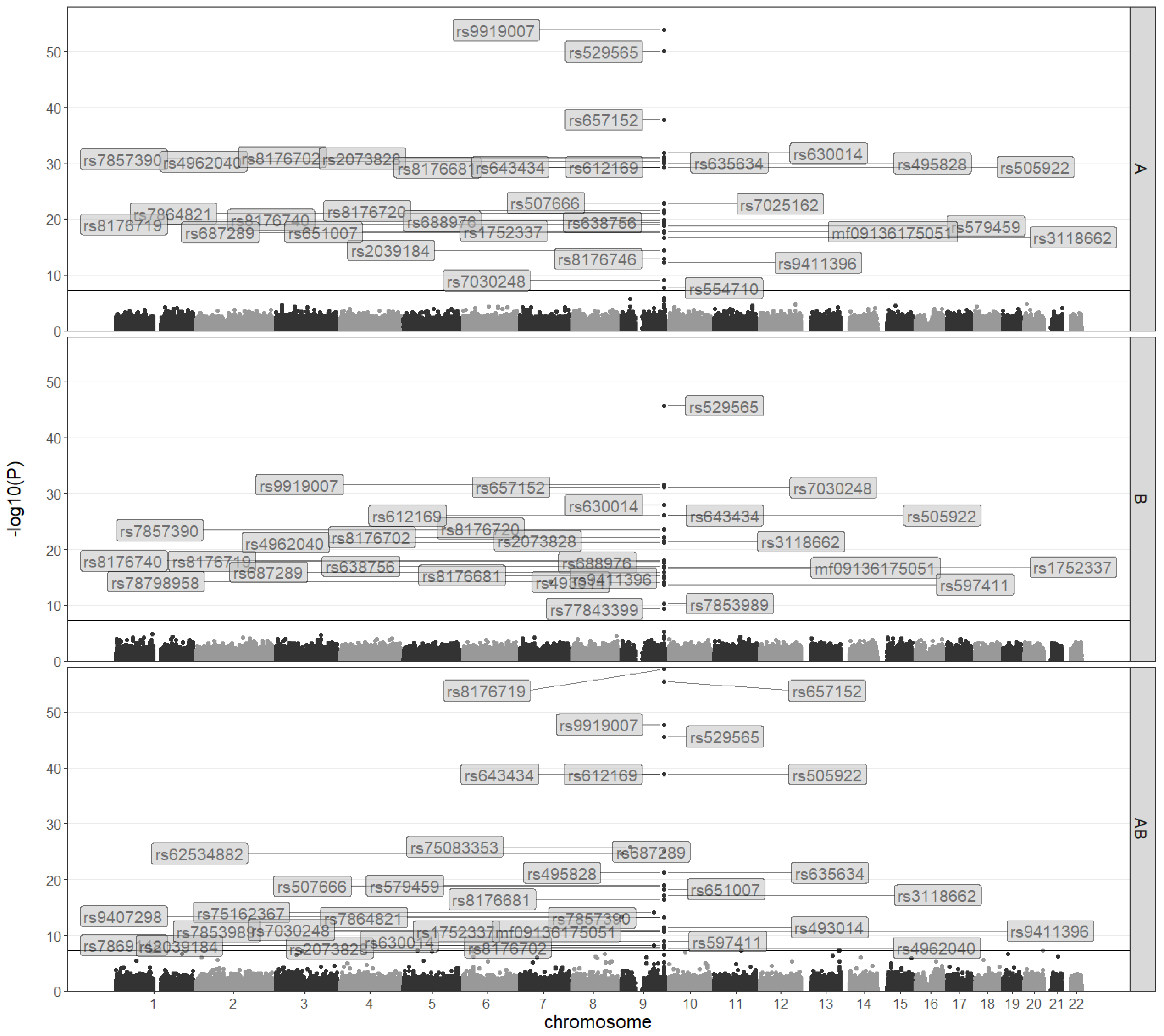
3.1. A GWAS on ABO Blood Types in a Chinese Population
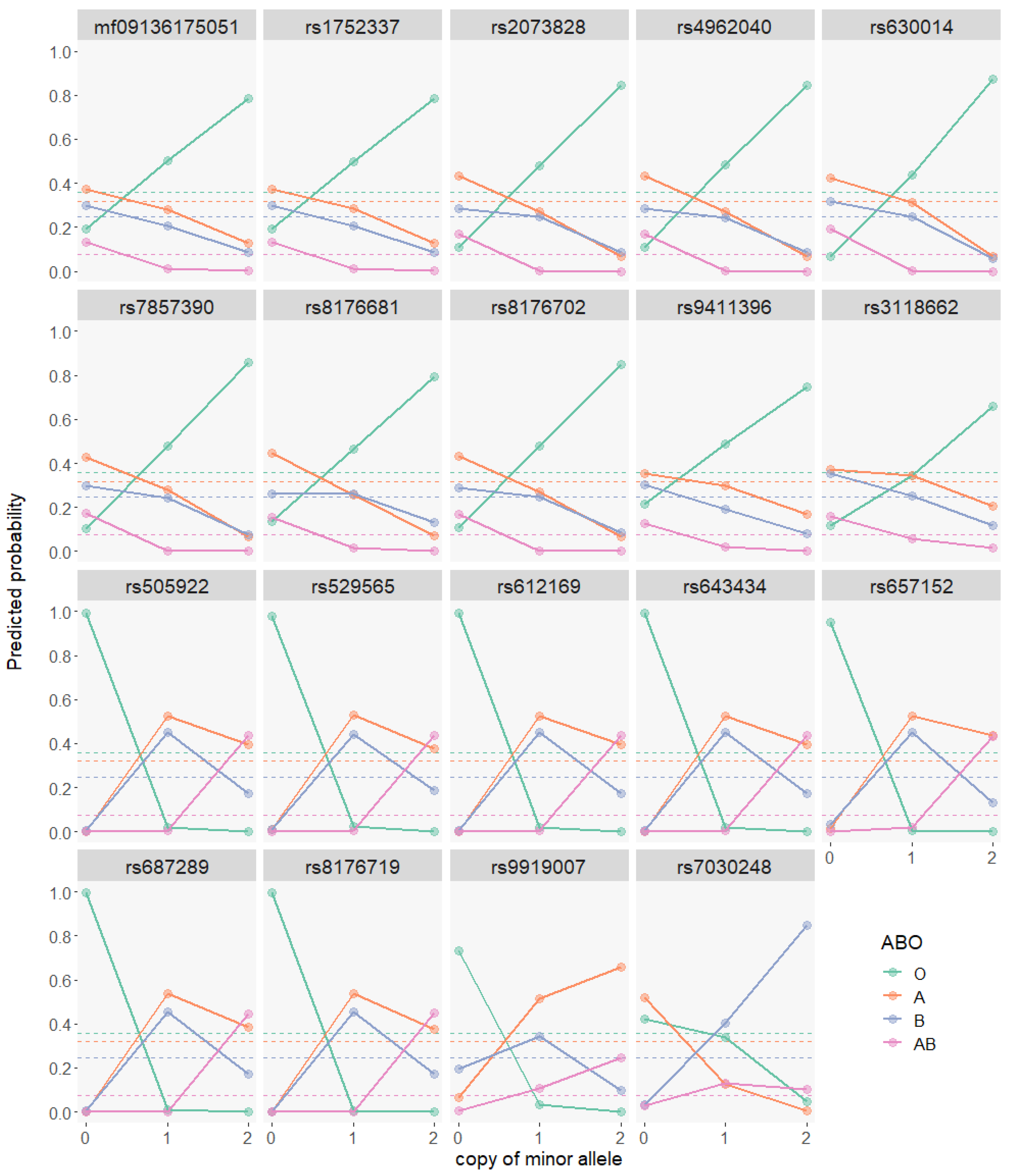
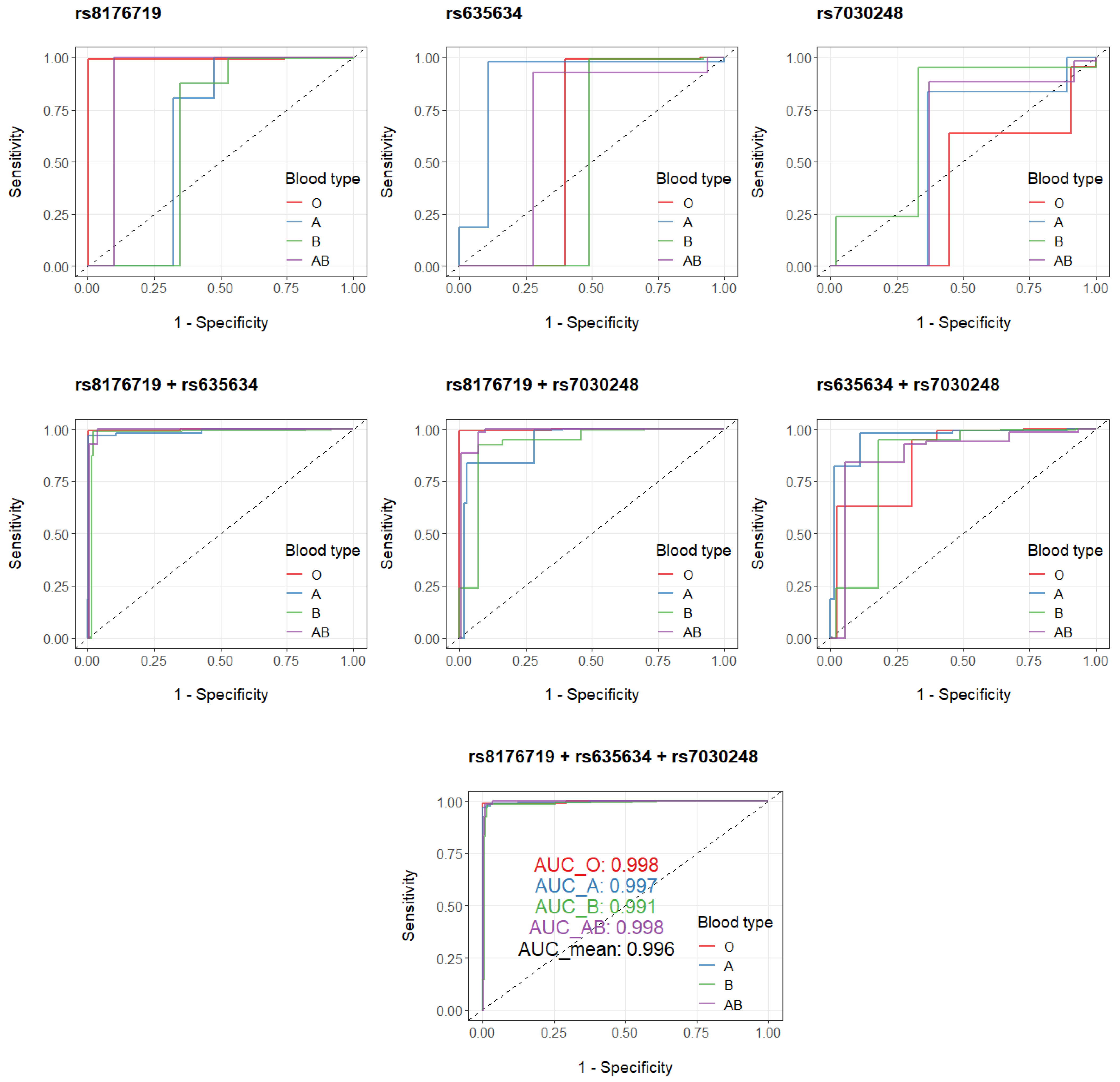
3.2. GWAS on ABO Blood Types Based on Binomial Models
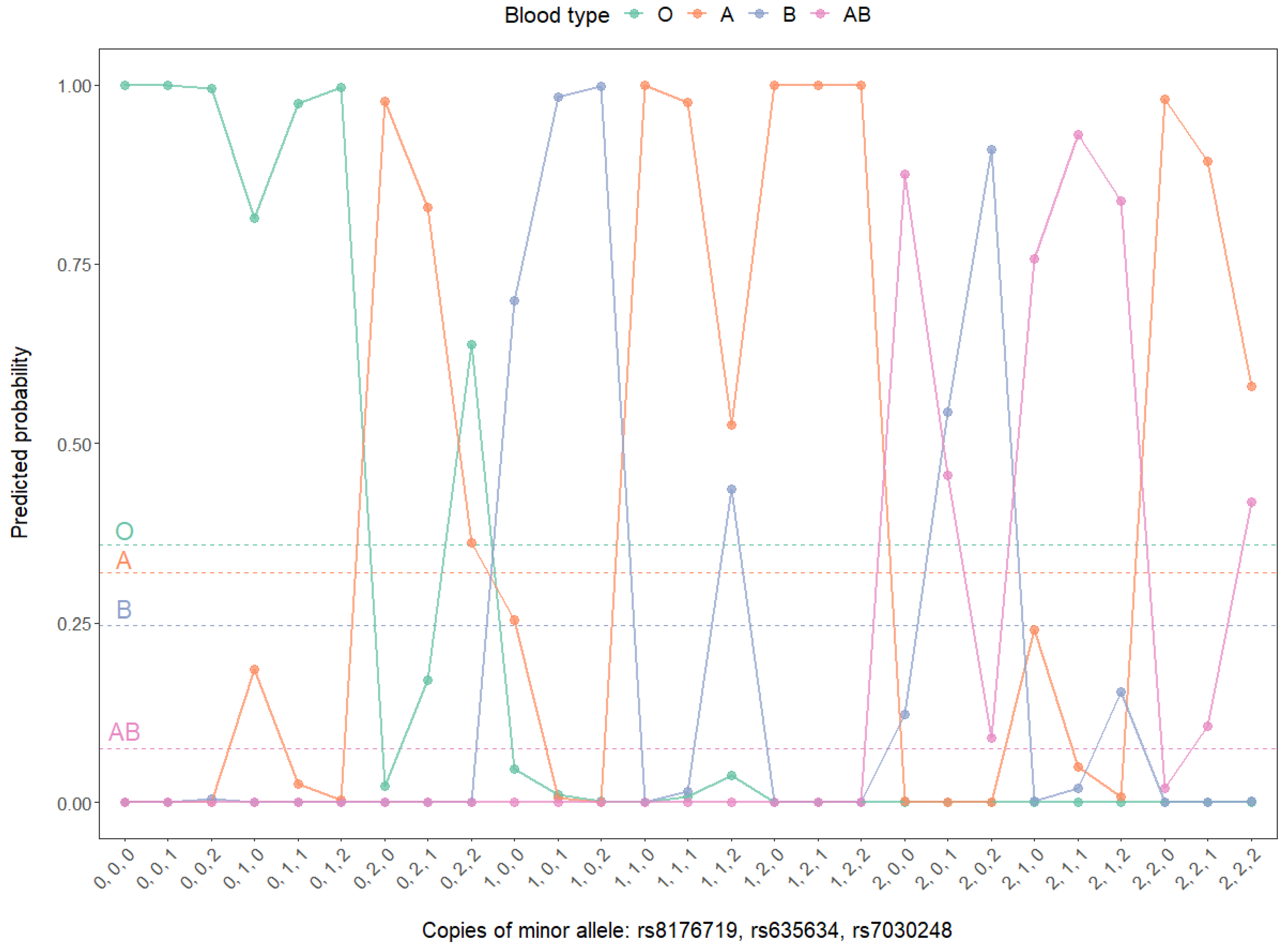
3.3. GWAS on ABO Blood Types Based on Multinomial Models
3.4. Causal Inference by Mendelian Randomization
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Karl Landsteiner-Biographical. NobelPrize.org. Nobel Media AB, 2020. Available online: https://www.nobelprize.org/prizes/medicine/1930/landsteiner/biographical/ (accessed on 8 March 2021).
- Clausen, H.; Bennett, E.P.; Grunnet, N. Molecular genetics of ABO histo-blood groups. Transfus. Clin. Biol. 1994, 1, 79–89. [Google Scholar] [CrossRef]
- Yamamoto, F.; Cid, E.; Yamamoto, M.; Blancher, A. ABO Research in the Modern Era of Genomics. Transfus. Med. Rev. 2012, 26, 103–118. [Google Scholar] [CrossRef] [PubMed]
- Fokkema, I.F.A.C.; Taschner, P.E.M.; Schaafsma, G.C.P.; Celli, J.; Laros, J.F.J.; den Dunnen, J.T. LOVD v.2.0: The next generation in gene variant databases. Hum. Mutat. 2011, 32, 557–563. [Google Scholar] [CrossRef]
- Groot, H.E.; Villegas Sierra, L.E.; Said, M.A.; Lipsic, E.; Karper, J.C.; van der Harst, P. Genetically Determined ABO Blood Group and its Associations with Health and Disease. Arterioscler. Thromb. Vasc. Biol. 2020, 40, 830–838. [Google Scholar] [CrossRef] [PubMed]
- Franchini, M.; Bonfanti, C. Evolutionary aspects of ABO blood group in humans. Clin. Chim. Acta 2015, 444, 66–71. [Google Scholar] [CrossRef]
- Hakomori, S. Antigen structure and genetic basis of histo-blood groups A, B and O: Their changes associated with human cancer. Biochim. Biophys. Acta 1999, 1473, 247–266. [Google Scholar] [CrossRef]
- Amundadottir, L.; Kraft, P.; Stolzenberg-Solomon, R.Z.; Fuchs, C.S.; Petersen, G.M.; Arslan, A.A.; Hoover, R.N. Genome-wide association study identifies variants in the ABO locus associated with susceptibility to pancreatic cancer. Nat. Genet. 2009, 41, 986–990. [Google Scholar] [CrossRef] [PubMed]
- Schunkert, H.; König, I.R.; Kathiresan, S.; Reilly, M.P.; Assimes, T.L.; Holm, H.; Cambien, F. Large-scale association analysis identifies 13 new susceptibility loci for coronary artery disease. Nat. Genet. 2011, 43, 333–338. [Google Scholar] [CrossRef]
- Dong, E.; Du, H.; Gardner, L. An interactive web-based dashboard to track COVID-19 in real time. Lancet Infect. Dis. 2020, 20, 533–534. [Google Scholar] [CrossRef]
- Lambert, H.; Gupte, J.; Fletcher, H.; Hammond, L.; Lowe, N.; Pelling, M.; Shanks, K. COVID-19 as a global challenge: Towards an inclusive and sustainable future. Lancet Planet. Health 2020, 4, e312–e314. [Google Scholar] [CrossRef]
- Li, J.; Wang, X.; Chen, J.; Cai, Y.; Deng, A.; Yang, M. Association between ABO blood groups and risk of SARS-CoV-2 pneumonia. Br. J. Haematol. 2020, 190, 24–27. [Google Scholar] [CrossRef] [PubMed]
- Golinelli, D.; Boetto, E.; Maietti, E.; Fantini, M.P. The association between ABO blood group and SARS-CoV-2 infection: A meta-analysis. PLoS ONE 2020, 15, e0239508. [Google Scholar]
- Wu, Y.; Feng, Z.; Li, P.; Yu, Q. Relationship between ABO blood group distribution and clinical characteristics in patients with COVID-19. Clin. Chim Acta 2020, 509, 220–223. [Google Scholar] [CrossRef]
- Severe Covid-19 GWAS Group. Genomewide Association Study of Severe Covid-19 with Respiratory Failure. N. Engl. J. Med. 2020, 383, 1522–1534. [Google Scholar] [CrossRef] [PubMed]
- Tam, V.; Patel, N.; Turcotte, M.; Bossé, Y.; Paré, G.; Meyre, D. Benefits and limitations of genome-wide association studies. Nat. Rev. Genet. 2019, 20, 467–484. [Google Scholar] [CrossRef]
- Li, Y.R.; Keating, B.J. Trans-ethnic genome-wide association studies: Advantages and challenges of mapping in diverse populations. Genome Med. 2014, 6, 91. [Google Scholar] [CrossRef] [PubMed]
- Fang, H.; Hui, Q.; Lynch, J.; Honerlaw, J.; Assimes, T.L.; Huang, J.; Tang, H. Harmonizing Genetic Ancestry and Self-identified Race/Ethnicity in Genome-wide Association Studies. Am. J. Hum. Genet. 2019, 105, 763–772. [Google Scholar] [CrossRef]
- Haga, S.B. Impact of limited population diversity of genome-wide association studies. Genet. Med. 2010, 12, 81–84. [Google Scholar] [CrossRef]
- Mills, M.C.; Rahal, C. The GWAS Diversity Monitor tracks diversity by disease in real time. Nat. Genet. 2020, 52, 242–243. [Google Scholar] [CrossRef]
- Zhao, X.; Hong, F.; Yin, J.; Tang, W.; Zhang, G.; Liang, X.; Li, X. Cohort profile: The China Multi-Ethnic cohort (CMEC) study. Int. J. Epidemiol. 2020. [Google Scholar] [CrossRef]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.; Bender, D.; Sham, P.C. PLINK: A Tool Set for Whole-Genome Association and Population-Based Linkage Analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef] [PubMed]
- Chandra, T.; Gupta, A. Association and Distribution of Hypertension, Obesity and ABO Blood groups in Blood Donors. Iran. J. Pediatr. Hematol. Oncol. 2012, 2, 140–145. [Google Scholar]
- Bugert, P.; Rink, G.; Kemp, K.; Klüter, H. Blood Group ABO Genotyping in Paternity Testing. Transfus. Med. Hemother. 2012, 39, 182–186. [Google Scholar] [CrossRef] [PubMed]
- Dahlgren, U.S.; Bennet, W. ABO-Incompatible Liver Transplantation—A Review of the Historical Background and Results. Int. Rev. Immunol. 2019, 38, 118–128. [Google Scholar] [CrossRef]
- Morath, C.; Zeier, M.; Döhler, B.; Opelz, G.; Süsal, C. ABO-Incompatible Kidney Transplantation. Front. Immunol. 2017, 8, 234. [Google Scholar] [CrossRef] [PubMed]
- Hobgood, D.K. Personality traits of aggression-submissiveness and perfectionism associate with ABO blood groups through catecholamine activities. Med. Hypotheses 2011, 77, 294–300. [Google Scholar] [CrossRef]
- Tsuchimine, S.; Saruwatari, J.; Kaneda, A.; Yasui-Furukori, N. ABO Blood Type and Personality Traits in Healthy Japanese Subjects. PLoS ONE 2015, 10, e0126983. [Google Scholar] [CrossRef] [PubMed]
- Burgess, S.; Labrecque, J.A. Mendelian randomization with a binary exposure variable: Interpretation and presentation of causal estimates. Eur. J. Epidemiol. 2018, 33, 947–952. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SNP | Accuracy | Sensitivity | Specificity | PPV | NPV | F1 | AUC |
|---|---|---|---|---|---|---|---|
| O vs. non-O | |||||||
| rs2039184 | 0.64 | 0.49 | 0.72 | 0.5 | 0.72 | 0.49 | 0.61 |
| rs76321958 | 0.63 | 0.38 | 0.77 | 0.48 | 0.69 | 0.43 | 0.58 |
| rs7864821 | 0.67 | 0.55 | 0.73 | 0.54 | 0.75 | 0.55 | 0.64 |
| rs9919007 | 0.86 | 0.96 | 0.81 | 0.73 | 0.97 | 0.83 | 0.88 |
| rs8176740 | 0.68 | 0.68 | 0.68 | 0.54 | 0.79 | 0.6 | 0.68 |
| rs7853989 | 0.69 | 0.99 | 0.52 | 0.54 | 0.99 | 0.7 | 0.75 |
| rs4962040 | 0.67 | 0.82 | 0.58 | 0.53 | 0.85 | 0.64 | 0.7 |
| rs529565 | 0.98 | 0.97 | 0.99 | 0.98 | 0.98 | 0.97 | 0.98 |
| rs630014 | 0.65 | 0.88 | 0.53 | 0.5 | 0.89 | 0.64 | 0.7 |
| rs651007 | 0.75 | 0.99 | 0.61 | 0.59 | 0.99 | 0.74 | 0.8 |
| rs7030248 | 0.58 | 0.64 | 0.55 | 0.44 | 0.73 | 0.52 | 0.59 |
| rs1752337 | 0.66 | 0.69 | 0.65 | 0.52 | 0.79 | 0.6 | 0.67 |
| rs493014 | 0.49 | 0.94 | 0.25 | 0.41 | 0.87 | 0.57 | 0.59 |
| rs9411396 | 0.65 | 0.66 | 0.65 | 0.51 | 0.77 | 0.57 | 0.65 |
| rs12763 | 0.59 | 0.58 | 0.6 | 0.45 | 0.72 | 0.51 | 0.59 |
| rs3118662 | 0.72 | 0.43 | 0.88 | 0.67 | 0.73 | 0.52 | 0.66 |
| full set | 0.99 | 0.99 | 0.99 | 0.99 | 1 | 0.99 | 0.99 |
| A vs. non-A | |||||||
| rs2039184 | 0.54 | 0.78 | 0.42 | 0.39 | 0.8 | 0.52 | 0.6 |
| rs7864821 | 0.56 | 0.8 | 0.45 | 0.4 | 0.82 | 0.54 | 0.62 |
| rs9919007 | 0.74 | 0.93 | 0.66 | 0.56 | 0.95 | 0.7 | 0.79 |
| rs7857390 | 0.6 | 0.56 | 0.62 | 0.41 | 0.75 | 0.47 | 0.59 |
| rs7853989 | 0.63 | 0.98 | 0.47 | 0.46 | 0.99 | 0.62 | 0.73 |
| rs8176720 | 0.77 | 0.62 | 0.83 | 0.63 | 0.83 | 0.63 | 0.73 |
| rs8176719 | 0.68 | 1 | 0.52 | 0.5 | 1 | 0.66 | 0.76 |
| rs630014 | 0.6 | 0.47 | 0.66 | 0.4 | 0.73 | 0.43 | 0.57 |
| rs635634 | 0.92 | 0.98 | 0.89 | 0.81 | 0.99 | 0.88 | 0.93 |
| rs7030248 | 0.7 | 0.84 | 0.64 | 0.52 | 0.89 | 0.64 | 0.74 |
| rs7025162 | 0.6 | 0.9 | 0.47 | 0.44 | 0.91 | 0.59 | 0.68 |
| rs13289928 | 0.59 | 0.61 | 0.59 | 0.41 | 0.76 | 0.49 | 0.6 |
| rs635634 + rs8176719 | 0.99 | 0.97 | 0.99 | 0.99 | 0.99 | 0.98 | 0.98 |
| full set | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| B vs. non-B | |||||||
| rs77843399 | 0.78 | 0.22 | 0.96 | 0.65 | 0.79 | 0.32 | 0.59 |
| rs8176720 | 0.81 | 0.47 | 0.92 | 0.65 | 0.84 | 0.54 | 0.69 |
| rs8176719 | 0.6 | 1 | 0.47 | 0.38 | 1 | 0.55 | 0.73 |
| rs635634 | 0.63 | 0.99 | 0.51 | 0.4 | 0.99 | 0.57 | 0.75 |
| rs7030248 | 0.74 | 0.95 | 0.67 | 0.48 | 0.98 | 0.64 | 0.81 |
| rs7025162 | 0.69 | 0.58 | 0.73 | 0.41 | 0.84 | 0.48 | 0.65 |
| rs493014 | 0.75 | 0.36 | 0.88 | 0.49 | 0.81 | 0.41 | 0.62 |
| rs3118662 | 0.67 | 0.37 | 0.77 | 0.34 | 0.79 | 0.35 | 0.57 |
| rs7030248 + rs635634 | 0.85 | 0.95 | 0.82 | 0.63 | 0.98 | 0.76 | 0.88 |
| full set | 0.99 | 0.99 | 0.99 | 0.96 | 1 | 0.97 | 0.99 |
| AB vs. non-AB | |||||||
| rs9919007 | 0.54 | 0.96 | 0.5 | 0.14 | 0.99 | 0.24 | 0.73 |
| rs8176746 | 0.75 | 1 | 0.73 | 0.23 | 1 | 0.38 | 0.87 |
| rs687289 | 0.91 | 0.99 | 0.9 | 0.44 | 1 | 0.61 | 0.94 |
| rs8176681 | 0.63 | 0.9 | 0.6 | 0.16 | 0.99 | 0.27 | 0.75 |
| rs507666 | 0.68 | 0.97 | 0.65 | 0.19 | 1 | 0.31 | 0.81 |
| rs3118662 | 0.74 | 0.55 | 0.76 | 0.16 | 0.95 | 0.24 | 0.66 |
| full set | 0.91 | 0.99 | 0.9 | 0.46 | 1 | 0.62 | 0.94 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, M.; Gao, J.; Liu, J.; Zhao, X.; Lei, Y. Genomic Association vs. Serological Determination of ABO Blood Types in a Chinese Cohort, with Application in Mendelian Randomization. Genes 2021, 12, 959. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12070959
Wang M, Gao J, Liu J, Zhao X, Lei Y. Genomic Association vs. Serological Determination of ABO Blood Types in a Chinese Cohort, with Application in Mendelian Randomization. Genes. 2021; 12(7):959. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12070959
Chicago/Turabian StyleWang, Mengqiao, Jiaqi Gao, Jin Liu, Xing Zhao, and Yi Lei. 2021. "Genomic Association vs. Serological Determination of ABO Blood Types in a Chinese Cohort, with Application in Mendelian Randomization" Genes 12, no. 7: 959. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12070959