
New Data on Comparative Cytogenetics of the Mouse-Like Hamsters (Calomyscus Thomas, 1905) from Iran and Turkmenistan

 , , , ,
, , , ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Compliance with Ethical Standards
2.2. Species Sampled
2.3. Chromosome Preparation and Chromosome Staining
2.4. Fluorescent in Situ Hybridization (FISH)
2.5. Microdissection, Probe Amplification, and Labeling
2.6. Image Acquisition and Processing
3. Results
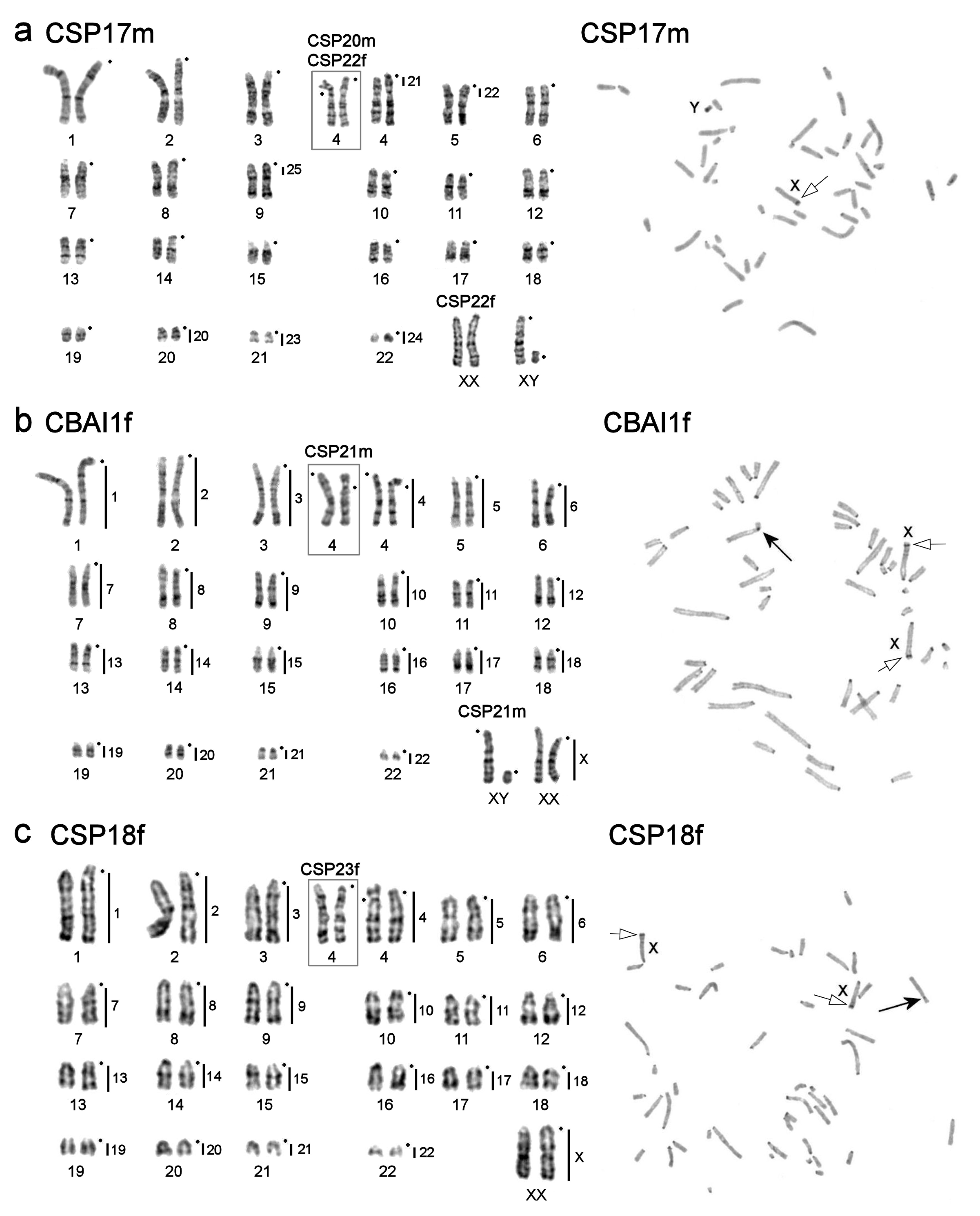
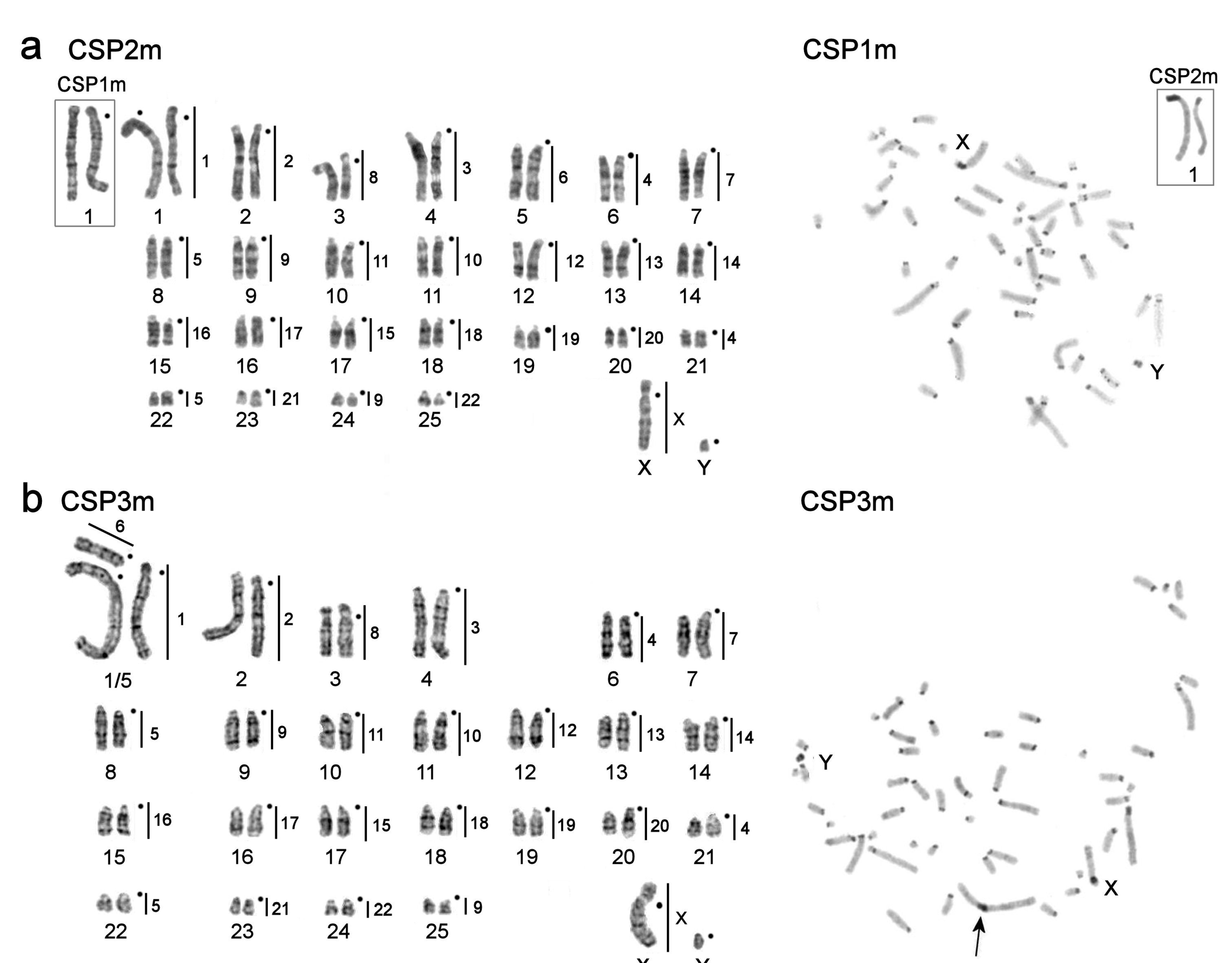
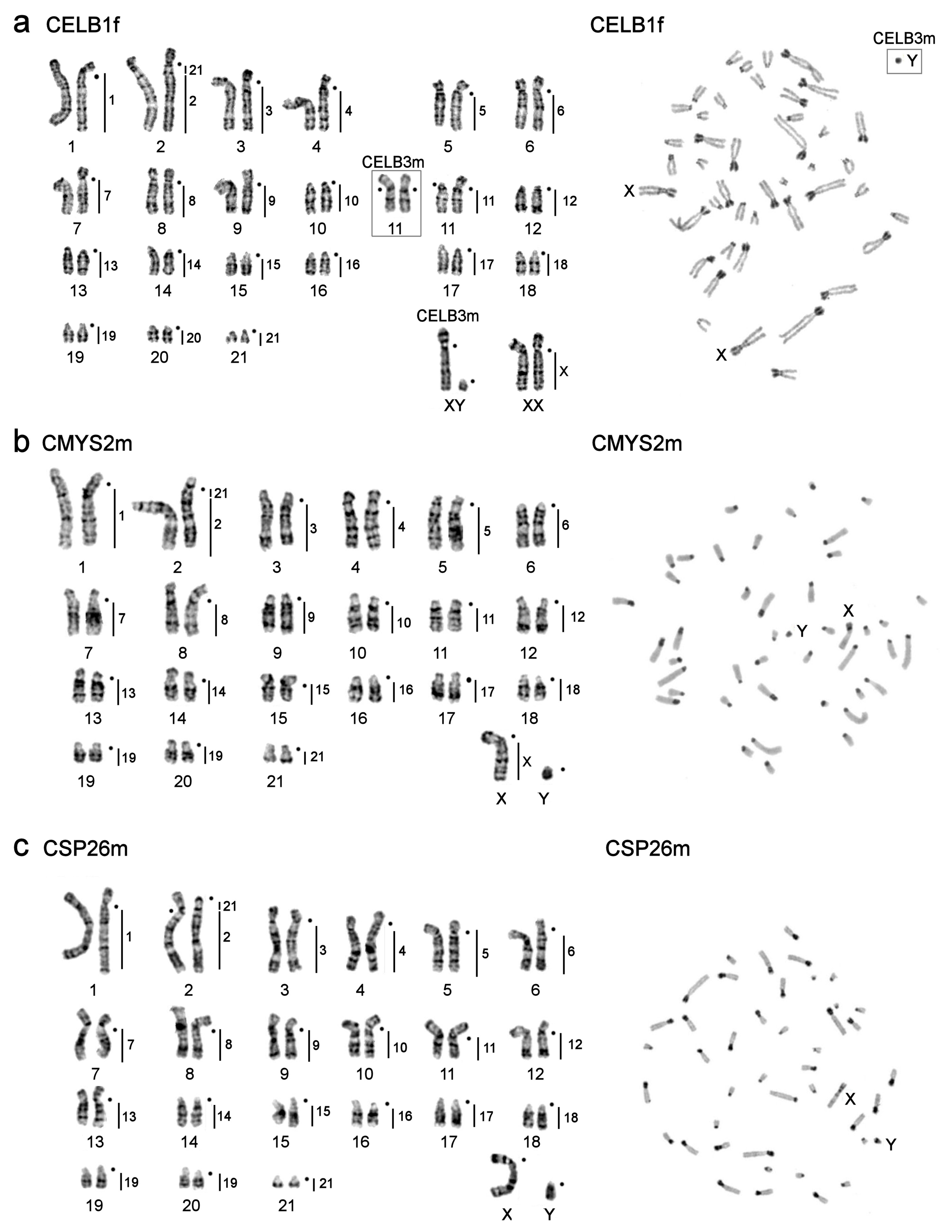
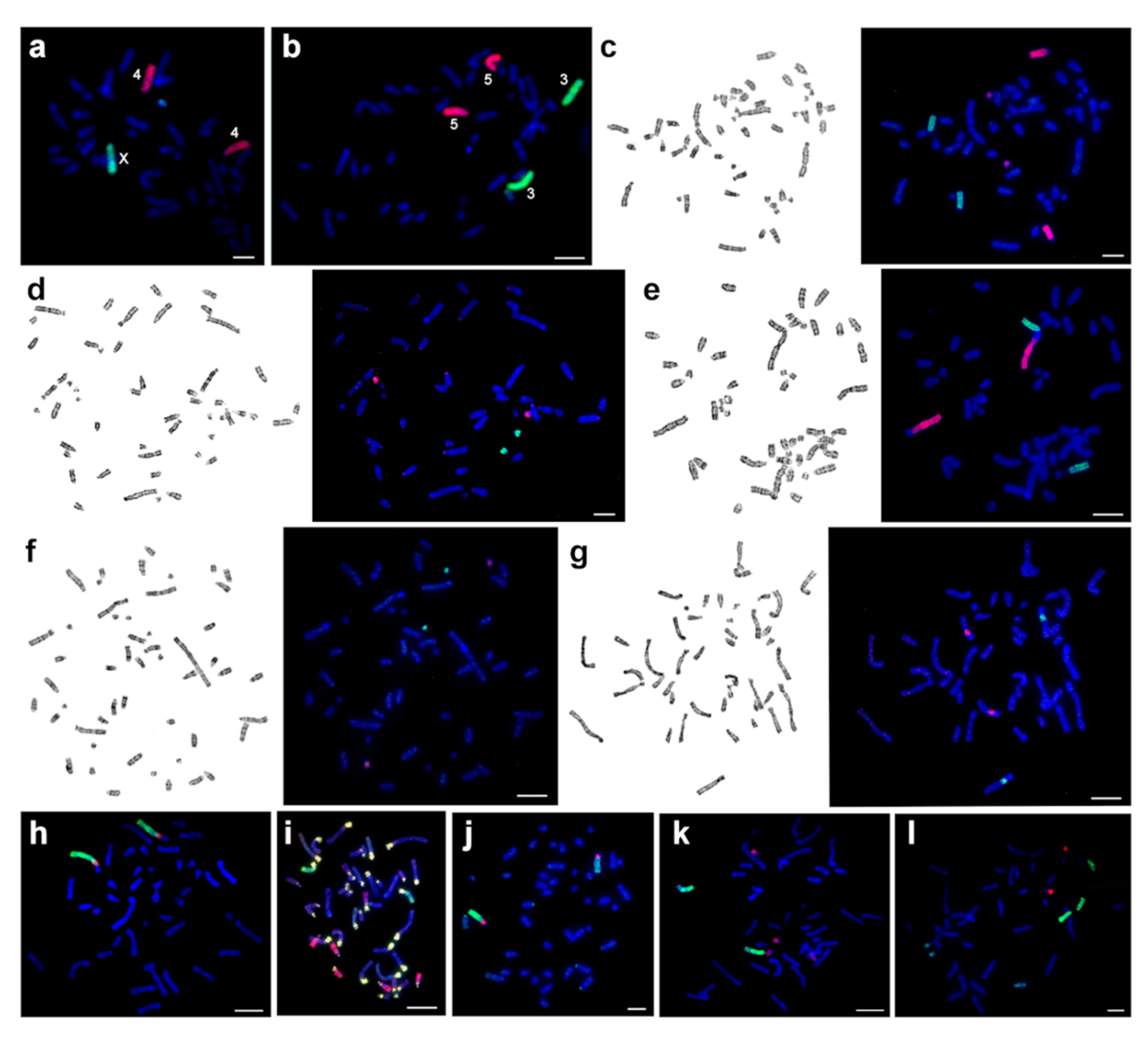
3.1. Karyotype Descriptions
3.2. The Results of Chromomycin A3-DAPI-after G-banding (CDAG)
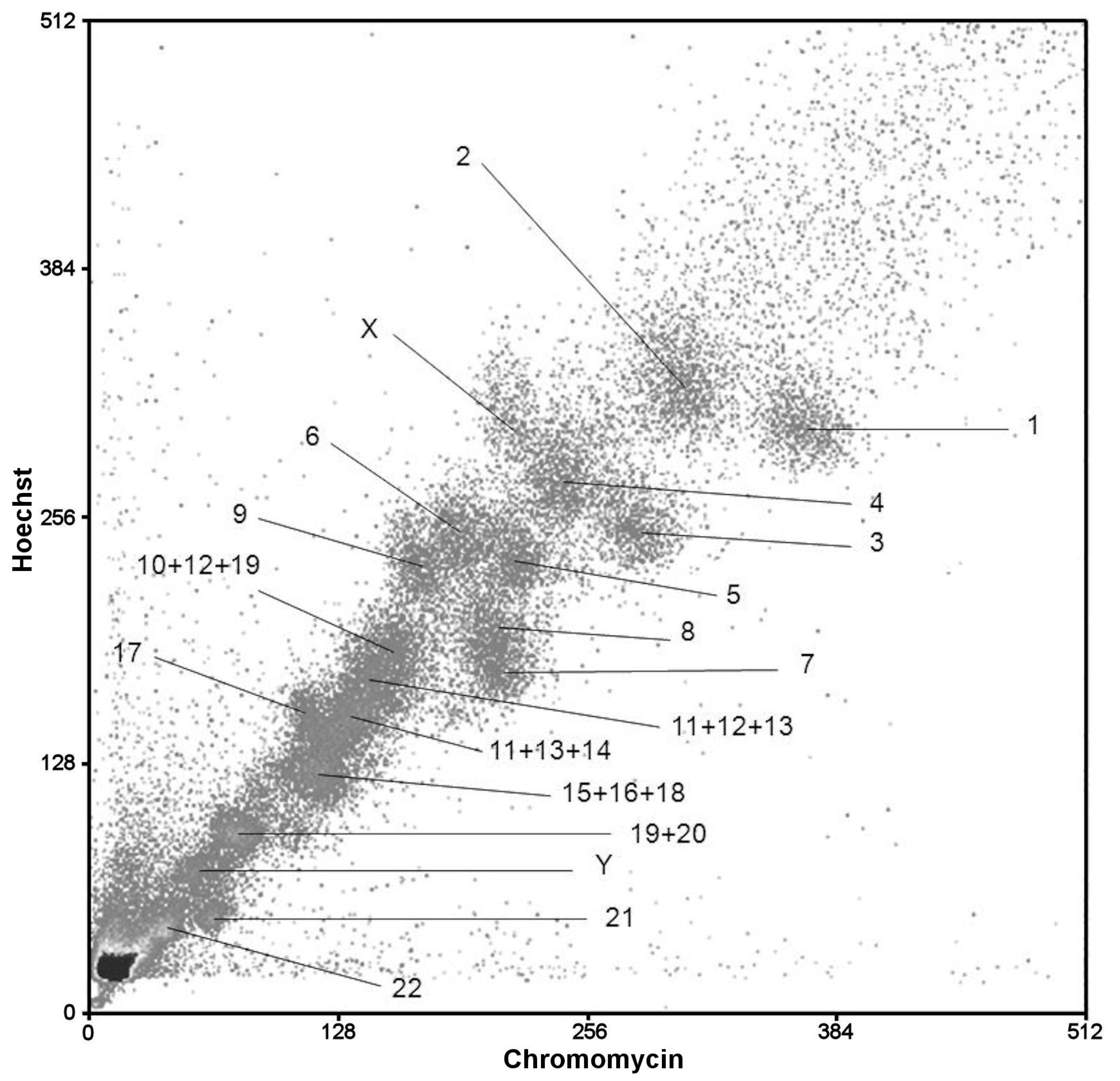
3.3. The Flow Karyotype of CSP17m
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Simpson, G.G. The principles of classification and a classification of Mammals. Bull. Am. Museum Nat. Hist. 1945, 1, 350. [Google Scholar] [CrossRef]
- Chaline, J.; Mein, P.; Petter, F. Les grandes lignes d’une classification évolutive des Muroidea. Mammalia 1977, 41. [Google Scholar] [CrossRef]
- Hooper, E.T. The Male Phallus in Mice of the Genus Peromyscus. Misc. Publ. Mus. Zool. 1958, 105, 1–24. [Google Scholar]
- Manville, R.H. The families of mammals. Science 1967, 157, 1421–1422. [Google Scholar] [CrossRef]
- Grassé, P.; Dekeyser, P.L. Ordre des Rongeurs. Trait. Zool. Anat. Syst. Biol. 1955, 17, 1321–1573. [Google Scholar]
- Vorontzov, N.N.; Potapova, E.G. Taxonomy of the genus Calomyscus (Cricetidae). Status of Calomyscus in the system of Cricetinae. Russ. J. Zool. 1979, 58, 1391–1397. [Google Scholar]
- Michaux, J.; Reyes, A.; Catzeflis, F. Evolutionary History of the Most Speciose Mammals: Molecular Phylogeny of Muroid Rodents. Mol. Biol. Evol. 2001, 18, 2017–2031. [Google Scholar] [CrossRef] [Green Version]
- Musser, G.G.; Carleton, M.D. Superfamily Muroidea. In Mammal Species of the World: A Taxonomie and Geographic Reference; John Hopkins University Press: Baltimore, MD, USA, 2005; pp. 894–1532. [Google Scholar]
- Thomas, O. Two new species of Calomyscus, (part B) scientific results from the mammal survey no. J. Bombay Nat. Hist. Soc. 1920, 26, 938–940. [Google Scholar]
- Kaschkarov, D. Materials to the knowledge of the rodents of the Turkestan. Trans. Sci. Soc. Turkestan Middle Asiat. Univ. 1925, 2, 43–56. [Google Scholar]
- Ognev, S.I.; Heptner, V.G. The mammals of the Central Kopet-Dag and the adjacent plain. Proc. Zool. Mus. Mosc. Univ. 1929, 3, 46–172. [Google Scholar]
- Argyropulo, A.I. The genera and species of Palearctic hamsters (Cricetinae). Proc. Zool. Inst. Acad. Sci. USSR 1933, 1, 239–248. [Google Scholar]
- Kalabuhov, N.I. The discovery of the mouse-like hamster in Transcaucasia. Nature 1939, 12, 83–84. [Google Scholar]
- Goodwin, G.G. Five New Rodents from the Eastern Elburz Mountains and a New Race of Hare from Teheran. American Museum Novitates; The American Museum of Natural History: New York, NY, USA, 1939. [Google Scholar]
- Ellerman, J.R. Key to the Rodents of South-West Asia in the British Museum Collection. Proc. Zool. Soc. London 1948, 765–816. [Google Scholar] [CrossRef]
- Ellerman, J.R.; Morrison-Scott, T.C.S. Checklist of Palaearctic and Indian Mammals 1758 to 1946; British Museum (Natural History): London, UK, 1951; 810p. [Google Scholar]
- Gromov, I.M.; Gureev, A.A.; Novikov, G.A.; Sokolov, I.I.; Strelkov, P.P.; Chaptskii, K.K. Order Rodentia. In Mammals of the USSR Fauna; AN SSSR: Moscow/Leningrad, Russia, 1963; pp. 244–638. [Google Scholar]
- Sokolov, V. Order Lagomorpha, Rodentia. In Systematics of Mammals; Vysshaya Shkola Press: Moscow, Russia, 1977. [Google Scholar]
- Corbet, G.B.; Hill, J.E. A World List of Mammalian Species; British Museum (Natural History) and Cornell University Press: London, UK; Ithaca: New York, NY, USA, 1980. [Google Scholar]
- Schlitter, D.A.; Setzer, H.W. New rodents (Mammalia: Cricetidae, Muridae) from Iran and Pakistan. Proc. Biol. Soc. Washingt. 1973, 86, 163–174. [Google Scholar]
- Peshev, D. The mouse like hamster (Calomyscus bailwardi Thomas, 1905), a new mammal, 1905), a new mammal for the Syrian fauna and the Arab peninsula. Mammalia 1989, 53, 109–112. [Google Scholar]
- Peshev, D. On the systematic position of the mouse-like hamster Calomyscus bailwardi Thomas, 1905 (Cricetidae, Rodentia) from the Near East and Middle Asia. Mammalia 1991, 55. [Google Scholar] [CrossRef]
- Matthey, R. Cytologie comparee des cricetinae palearctiques et d’Americanes. Rev. Suisse Zool. 1961, 68, 41–61. [Google Scholar]
- Vorontsov, N.N.; Kartavtseva, I.V.; Potapova, E.G. The systematic of mouse-like hamsters from Calomyscus (Cricetidae). The karyological differentiation of sibling species from Transcaucasia and Turkmenistan and review of species of Calomyscus genus. Russ. J. Zool. 1979, 58, 1213–1224. [Google Scholar]
- Graphodatsky, A.S.; Sablina, O.V.; Meyer, M.N.; Malikov, V.G.; Isakova, E.A.; Trifonov, V.A.; Polyakov, A.V.; Lushnikova, T.P.; Vorobieva, N.V.; Serdyukova, N.A.; et al. Comparative cytogenetics of hamsters of the genus Calomyscus. Cytogenet. Genom. Res. 2000, 88, 296–304. [Google Scholar] [CrossRef] [PubMed]
- Meyer, M.; Malikov, V.G. New species and subspecies of mouse-like hamsters of the genus Calomyscus (Rodentia, Cricetidae) from southern Turkmenistan. Russ. J. Zool. 2000, 79, 219–223. [Google Scholar]
- Lebedev, V.S.; Pavlinov, I. Ya.; Meyer, M.N.; Malikov, V.G. Craniometric analysis of mouse-like hamsters of the genus Calomyscus (Critecidae). Zool. Zhurnal 1998, 77, 721–731. [Google Scholar]
- Shahabi, S.; Aliabadian, M.; Darvish, J.; Kilpatrick, C.W. Molecular phylogeny of brush-tailed mice of the genus Calomyscus (Rodentia: Calomyscidae) inferred from mitochondrial DNA sequences (Cox1 gene). Mammalia 2013, 77, 425–431. [Google Scholar] [CrossRef]
- Lebedev, V.S.; Bannikova, A.A.; Neumann, K.; Ushakova, M.V.; Ivanova, N.V.; Surov, A.V. Molecular phylogenetics and taxonomy of dwarf hamsters Cricetulus Milne-Edwards, 1867 (Cricetidae, Rodentia): Description of a new genus and reinstatement of another. Zootaxa 2018, 4387, 331–339. [Google Scholar] [CrossRef]
- Norris, R.W.; Woods, C.A.; Kilpatrick, C.W. Morphological and molecular definition of Calomyscus hotsoni (Rodentia: Muroidea: Calomyscidae). J. Mammal. 2008, 89, 306–315. [Google Scholar] [CrossRef] [Green Version]
- Rezazadeh, E.; Aliabadian, M.; Darvish, J.; Ahmadzadeh, F. Diversification and evolutionary history of brush-tailed mice, Calomyscidae (Rodentia), in southwestern Asia. Org. Divers. Evol. 2020, 20, 155–170. [Google Scholar] [CrossRef]
- Stanyon, R.; Galleni, L. A rapid fibroblast culture technique for high resolution karyotypes. Bollet. Zool. 1991, 58, 81–83. [Google Scholar] [CrossRef] [Green Version]
- Romanenko, S.A.; Biltueva, L.S.; Serdyukova, N.A.; Kulemzina, A.I.; Beklemisheva, V.R.; Gladkikh, O.L.; Lemskaya, N.A.; Interesova, E.A.; Korentovich, M.A.; Vorobieva, N.V.; et al. Segmental paleotetraploidy revealed in sterlet (Acipenser ruthenus) genome by chromosome painting. Mol. Cytogenet. 2015, 8, 90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, F.; Graphodatsky, A.S. Animal probes and Zoo-FISH. In Fluorescence In Situ Hybridization (FISH)—Application Guide; Springer: Berlin/Heldelberg, Germany, 2017; pp. 395–416. [Google Scholar]
- Gladkikh, O.L.; Romanenko, S.A.; Lemskaya, N.A.; Serdyukova, N.A.; O’Brien, P.C.M.; Kovalskaya, J.M.; Smorkatcheva, A.V.; Golenishchev, F.N.; Perelman, P.L.; Trifonov, V.A.; et al. Rapid karyotype evolution in Lasiopodomys involved at least two autosome-Sex chromosome translocations. PLoS ONE 2016, 11, e0167653. [Google Scholar] [CrossRef]
- Seabright, M. A rapid banding technique for human chromosomes. Lancet 1971, 2, 971–972. [Google Scholar] [CrossRef]
- Lemskaya, N.A.; Kulemzina, A.I.; Beklemisheva, V.R.; Biltueva, L.S.; Proskuryakova, A.A.; Hallenbeck, J.M.; Perelman, P.L.; Graphodatsky, A.S. A combined banding method that allows the reliable identification of chromosomes as well as differentiation of AT- and GC-rich heterochromatin. Chromosom. Res. 2018, 26, 307–315. [Google Scholar] [CrossRef]
- Trifonov, V.A.; Vorobieva, N.V.; Serdyukova, N.A.; Rens, W. FISH with and Without COT1 DNA. In Fluorescence In Situ Hybridization (FISH)—Application Guide; Springer: Berlin/Heldelberg, Germany, 2017; pp. 123–135. [Google Scholar]
- Telenius, Håk.; Ponder, B.A.J.; Tunnacliffe, A.; Pelmear, A.H.; Carter, N.P.; Ferguson-Smith, M.A.; Behmel, A.; Nordenskjöld, M.; Pfragner, R. Cytogenetic analysis by chromosome painting using dop-PCR amplified flow-sorted chromosomes. Genes Chromosom. Cancer 1992, 4, 257–263. [Google Scholar] [CrossRef]
- Yang, F.; Carter, N.P.; Shi, L.; Ferguson-Smith, M.A. A comparative study of karyotypes of muntjacs by chromosome painting. Chromosoma 1995, 103, 642–652. [Google Scholar] [CrossRef]
- Sitnikova, N.A.; Romanenko, S.A.; O’Brien, P.C.M.; Perelman, P.L.; Fu, B.; Rubtsova, N.V.; Serdukova, N.A.; Golenishchev, F.N.; Trifonov, V.A.; Ferguson-Smith, M.A.; et al. Chromosomal evolution of Arvicolinae (Cricetidae, Rodentia). I. The genome homology of tundra vole, field vole, mouse and golden hamster revealed by comparative chromosome painting. Chromosom. Res. 2007, 15, 447–456. [Google Scholar] [CrossRef] [PubMed]
- Romanenko, S.A.; Lemskaya, N.A.; Trifonov, V.A.; Serdyukova, N.A.; O’Brien, P.C.M.; Bulatova, N.S.; Golenishchev, F.N.; Ferguson-Smith, M.A.; Yang, F.; Graphodatsky, A.S. Genome-wide comparative chromosome maps of Arvicola amphibius, Dicrostonyx torquatus, and Myodes rutilus. Chromosom. Res. 2016, 24, 145–159. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Trifonov, V.; Ng, B.L.; Kosyakova, N.; Carter, N.P. Generation of paint probes from flow-sorted and microdissected chromosomes. In Fluorescence In Situ Hybridization (FISH); Springer: Berlin/Heldelberg, Germany, 2017; pp. 63–79. [Google Scholar]
- Romanenko, S. Calomyscus bailwardi. In Atlas of Mammalian Chromosomes; Graphodatsky, A.S., Perelman, P.L., O’Brien, S.J., Eds.; Wiley: Hoboken, NJ, USA, 2020; p. 410. ISBN 9781119418030. [Google Scholar]
- Romanenko, S.A.; Volobouev, V.T.; Perelman, P.L.; Lebedev, V.S.; Serdukova, N.A.; Trifonov, V.A.; Biltueva, L.S.; Nie, W.; O’Brien, P.C.M.; Bulatova, N.S.; et al. Karyotype evolution and phylogenetic relationships of hamsters (Cricetidae, Muroidea, Rodentia) inferred from chromosomal painting and banding comparison. Chromosom. Res. 2007, 15, 283–297. [Google Scholar] [CrossRef] [PubMed]
- Romanenko, S. Calomyscus sp. In Atlas of Mammalian Chromosomes; Graphodatsky, A.S., Perelman, P.L., O’Brien, S.J., Eds.; Wiley: Hoboken, NJ, USA, 2020; p. 408. ISBN 9781119418030. [Google Scholar]
- Romanenko, S. Calomyscus elburzensis. In Atlas of Mammalian Chromosomes; Graphodatsky, A.S., Perelman, P.L., O’Brien, S.J., Eds.; Wiley: Hoboken, NJ, USA, 2020; p. 411. ISBN 9781119418030. [Google Scholar]
- Romanenko, S. Calomyscus mystax. In Atlas of Mammalian Chromosomes; Graphodatsky, A.S., Perelman, P.L., O’Brien, S.J., Eds.; Wiley: Hoboken, NJ, USA, 2020; p. 409. ISBN 9781119418030. [Google Scholar]
- Shahabi, S.; Zarei, B.; Sahebjam, B. Karyologic Study of Three Species of Calomyscus (Rodentia: Calomyscidae) from Iran. Iran. J. Anim. Biosyst. IJAB 2010, 6, 55–60. [Google Scholar]
- Radjabli, S.I.; Sablina, O.V.; Graphodatsky, A.S. Calomyscus bailwardi. In Atlas of Mammalian Chromosomes; Wiley Online Library: Hoboken, NJ, USA, 2006; p. 202. [Google Scholar]
- Lemskaya, N.A.; Kartavtseva, I.V.; Rubtsova, N.V.; Golenishchev, F.N.; Sheremetyeva, I.N.; Graphodatsky, A.S. Chromosome Polymorphism in Microtus (Alexandromys) mujanensis (Arvicolinae, Rodentia). Cytogenet. Genome Res. 2015, 146, 238–242. [Google Scholar] [CrossRef] [PubMed]
- Meyer, M.N.; Golenishchev, F.N.; Radjably, S.I.; Sablina, O.V. Voles (subgenus Microtus Schrank) of Russia and adjacent territories. Proc. Zool. Inst. RAS 1996, 232, 1–320. [Google Scholar]
- Shahabi, S.; Darvish, J.; Aliabadian, M.; Mirshamsi, O.; Mohammadi, Z. Cranial and dental analysis of mouse-like hamsters of the genus Calomyscus (Rodentia: Calomyscidae) from plateau of Iran. Hystrix It. J. Mamm. 2012, 22, 311–323. [Google Scholar]
- Akbarirad, S.; Darvish, J.; Aliabadian, M. Phylogeography of Calomyscus elburzensis (Calomyscidae, Rodentia) around the Central Iranian Desert with Description of a New Subspecies in Center of Iranian Plateau. J. Sci. Islam. Repub. Iran 2016, 27, 5–21. [Google Scholar]
- Rawson, B. Phylogenomic and Species Delimitation in the Brush-Tailed Mouse Genus Calomyscus Using DDRADSEQ data. In An Undergraduate Research Thesis; The Ohio State University: Columbus, OH, USA, 2019. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Calomyscus Species, Diploid Chromosome Number (2n) | Geographic Position in Figure 1 | Sex | Abbreviation | Locality | Coordinates/Comments |
|---|---|---|---|---|---|
| C. bailwardi (2n = 46) | 1 | ♀ | CBAI1f | Izeh, Khuzestan Province, Iran * | N 31°49′20.8″ E 49°50′09.6″, elevation: 1063 m |
| C. elburzensis (2n = 44) | 2 | ♀ | CELB1f | Korkhout Mt., North Khorasan Province, Bojnourd, Iran * | N 37°26′35.2″ E 56°31′30.0″, elevation: 1903 m |
| C. elburzensis (2n = 44) | 2 | ♂ | CELB3m | Korkhout Mt.,North Khorasan Province, Bojnourd, Iran * | N 37°26′35.2″ E 56°31′30.0″, elevation: 1903 m (laboratory colony of ZIN RAN) |
| C. mystax mystax (2n = 44) | 3 | ♂ | CMYS2m | Maly Balkan Gershi (Arlandag) Mt., Bolshoi Balkhan, Turkmenistan * | N 39°40′20.28″ E 54°32′35.61″, elevation: 1870 m (laboratory colony of ZIN RAN) |
| Calomyscus sp. 1 (2n = 52) | 4 | ♂ | CSP1m ** | Shahdad tunnel, Kerman Province, Iran | N 30°10′36.41″ E 57°24′44.24″, elevation: 2664 m (laboratory colony of Moscow Zoo) |
| Calomyscus sp. 1 (2n = 52) | 4 | ♂ | CSP2m ** | Shahdad tunnel, Kerman Province, Iran | N 30°10′36.41″ E 57°24′44.24″, elevation: 2664 m (laboratory colony of Moscow Zoo) |
| Calomyscus sp. 1 (2n = 51) | 4 | ♂ | CSP3m ** | Shahdad tunnel, Kerman Province, Iran | N 30°10′36.41″ E 57°24′44.24″, elevation: 2664 m (laboratory colony of Moscow Zoo) |
| Calomyscus sp. 2 (2n = 46) | 5 | ♂ | CSP17m | Cheshme Maiak, Shahr-e-Kord—Soreshjan, Chahar Mahal and Bakhtiaria Province, Iran | N 32°18′37.9″ E 50°37′34.3″, elevation 2161 m |
| Calomyscus sp. 2 (2n = 46) | 5 | ♀ | CSP18f | Aloni, Chahar Mahal and Bakhtiaria Province, Iran | N 31°33′21.3″ E 51°11′25.3″, elevation 1833 m |
| Calomyscus sp. 2 (2n = 46) | 5 | ♂ | CSP20m | Cheshme Maiak, Shahr-e-Kord—Soreshjan, Chahar Mahal and Bakhtiaria Province, Iran | N 32°18′37.9″ E 50°37′34.3″, elevation 2161 m |
| C. bailwardi (2n = 46) | 5 | ♂ | CSP21m | Izeh, Khozestan Province, Iran | N 31°49′20.8″ E 49°50′09.6″, elevation: 1063 m |
| Calomyscus sp. 2 (2n = 46) | 5 | ♀ | CSP22f | Cheshme Maiak, Shahr-e-Kord—Soreshjan, Bakhtiaria Province, Iran | N 32°18′37.9″ E 50°37′34.3″, elevation 2161 m |
| Calomyscus sp. 2 (2n = 46) | 5 | ♀ | CSP23f | Aloni, Chahar Mahal and Bakhtiaria Province, Iran | N 31°33′21.3″ E 51°11′25.3″, elevation 1833 m |
| Calomyscus sp. 3 (2n = 44) | 6 | ♂ | CSP26m | S-E ridge Zard-Kuh Mts., Isfahan Province, Iran | N 32°27′4.93″ E 50°2′28.36″, elevation 2885 m |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Romanenko, S.A.; Malikov, V.G.; Mahmoudi, A.; Golenishchev, F.N.; Lemskaya, N.A.; Pereira, J.C.; Trifonov, V.A.; Serdyukova, N.A.; Ferguson-Smith, M.A.; Aliabadian, M.; et al. New Data on Comparative Cytogenetics of the Mouse-Like Hamsters (Calomyscus Thomas, 1905) from Iran and Turkmenistan. Genes 2021, 12, 964. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12070964
Romanenko SA, Malikov VG, Mahmoudi A, Golenishchev FN, Lemskaya NA, Pereira JC, Trifonov VA, Serdyukova NA, Ferguson-Smith MA, Aliabadian M, et al. New Data on Comparative Cytogenetics of the Mouse-Like Hamsters (Calomyscus Thomas, 1905) from Iran and Turkmenistan. Genes. 2021; 12(7):964. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12070964
Chicago/Turabian StyleRomanenko, Svetlana A., Vladimir G. Malikov, Ahmad Mahmoudi, Feodor N. Golenishchev, Natalya A. Lemskaya, Jorge C. Pereira, Vladimir A. Trifonov, Natalia A. Serdyukova, Malcolm A. Ferguson-Smith, Mansour Aliabadian, and et al. 2021. "New Data on Comparative Cytogenetics of the Mouse-Like Hamsters (Calomyscus Thomas, 1905) from Iran and Turkmenistan" Genes 12, no. 7: 964. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12070964